
Abstract
Neuropathic pain is a significant clinical issue due to the limited efficacy and adverse effects of existing therapies. This study evaluated the antinociceptive effects of the GABA-derived short peptides pyroglutamylGABA, pyroglutamylGABA ethyl ester, and pyroglutamyl diGABA in combination with pregabalin in a rat model of neuropathic pain induced by chronic constriction injury. Pain-related behavior was assessed using the hot plate test by recording changes in the latency and frequency of licking, jumping, and locomotor activity. Co-administration of peptide–pregabalin combinations modulated nociceptive responses compared with control animals in a time- and compound-dependent manner. The combination of pyroglutamylGABA with pregabalin was associated with increased licking latency and reduced licking frequency, particularly at early time points after injury. The pyroglutamylGABA ethyl ester–pregabalin combination produced the most pronounced effects on jumping-related parameters. A delay in movements was recorded when a combination of pyroglutamyl diGABA and pregabalin was used, whereas all combinations demonstrated increased locomotor activity, suggesting that the antinociceptive effects were not accompanied by the motor suppression typically associated with pregabalin treatment. To explore the potential mechanisms, molecular docking studies were performed targeting key GABAergic system components, including the GAT1 transporter, GABA-AT enzyme, and GABA-B receptors. The investigated peptides demonstrated favorable binding affinities across these targets, suggesting that potential interactions with multiple targets are involved in GABAergic neurotransmission. These findings indicate that the combinations of GABA-derived peptides with pregabalin could be an effective approach in neuropathic pain management. However, further investigation is required to validate these effects and clarify the underlying mechanisms.
Plain language summary
Neuropathic pain is a chronic condition caused by damage of the nervous system. It affects millions of people worldwide and is often undertreated. Although medications such as pregabalin are commonly used to relieve neuropathic pain, they do not always provide sufficient effectiveness and can cause adverse effects such as drowsiness and sedation. Therefore, new treatment strategies that improve pain control while minimizing side effects are needed. In this study, we investigated analgesic effects of combinations of pregabalin with newly synthesized short peptides derived from GABA—pyroglutamylGABA, pyroglutamylGABA ethyl ester, and pyroglutamyl diGABA—in a rat model of neuropathic pain caused by chronic constriction injury of the sciatic nerve. Pain-related behavior was evaluated using the hot plate test, which measures responses to a painful heat stimulus. The results showed that peptide–pregabalin combinations can reduce different characteristics of pain-related behaviors compared with untreated animals. Importantly, these combinations did not reduce the animals’ overall motor activity, suggesting that the analgesic effects were not accompanied by the motor suppression often associated with pregabalin treatment. Computer-based molecular docking analyses further suggested that the peptides may interact with key targets of the GABAergic system, including GABA-B receptors, the GAT1 transporter, and the GABA-AT enzyme. Hence, these findings indicate that GABA-derived peptides combined with pregabalin may represent a promising strategy for improving neuropathic pain treatment. However, additional studies are needed to confirm these effects and better understand the underlying mechanisms.

Keywords
Introduction
The development of innovative approaches for the management of neuropathic pain (NP) and the improvement of pharmacological interventions remain urgent, unmet clinical needs owing to the significant medical, financial, and socio-economic burden on the healthcare system.1,2 Approximately 7%–10% of the world’s population suffers from NP,3,4 and the response of patients to various therapies is not always satisfactory, increasing in resistance to treatment and leading to severe mental health problems, depressed mood, and deterioration of the quality of life.5,6
Drug development for NP management is based on its complex etiopathogenic mechanisms, which involve several well-known neurotransmitters that maintain nervous system homeostasis. 7 The recently discovered molecular control of GABAergic plasticity related to glutamatergic NMDA receptors highlights the importance of GABA–glutamate balance in neuronal processing,8,9 suggesting the potential of GABAergic and glutamatergic agents in the management of NP.
Among the GABAergic drugs used to treat of NP, the GABA analog pregabalin (PGB) deserves special attention because of its ability to reverse NP symptoms, such as allodynia and hyperalgesia. 10 Preclinical studies using PGB in various NP models have investigated several molecular targets of action, such as Ca2+ channel-dependent release of neurotransmitters, activation of excitatory amino acid carrier proteins, activation of potassium channels, and reduction of neuroinflammation.11–13 Despite its effectiveness in treating NP, PGB has several adverse effects, including drowsiness and somnolence.14,15 The manifestation of undesirable side effects represents a major challenge in modern pharmacotherapy, 16 evidenced by the use of various well-known drugs with proven effectiveness. Therefore, it is important to develop new treatment approaches with novel agents and combination-based analgesic therapies that could potentiate the effectiveness of prescribed drugs and simultaneously reduce the risk of side effects.17,18 Hence, multimodal analgesia, in which two or more analgesics are used in reduced dosages, may provide additive or synergistic effects, resulting in greater effectiveness and fewer side effects than with a single drug therapy.19–22
Therefore, the effects of the combination of pregabalin with newly synthesized short peptides containing GABA and pyroglutamic acid—pyroglutamylGABA (pGlu-GABA), pyroglutamylGABA ethyl ester (pGlu-GABA ethyl ester), and pyroglutamyl diGABA (pGlu-diGABA) were investigated as pain-preventive agents for the management of NP without the expected side effects of PGB. Relevant drug combination strategies have also been used by various researchers. Thus, the development of tolerance and addiction to morphine can be reduced when morphine is used in combination with imidazoline2-receptor agonists, which are being investigated as new drugs for the treatment of chronic pain, as recently reported by Bektas et al. 23 Moreover, there are many studies on promising combination-based analgesic therapies involving PGB and other gabapentinoids, such as gabapentinoid-opioids and gabapentinoid-antidepressants,24–27 which aim to use lower doses to reduce side effects, but additional or cumulative activity is expected.
The choice of pGlu-GABA, pGlu-GABA ethyl ester, and pGlu-diGABA new short peptides for combination with pregabalin was based on their antinociceptive and cerebrovascular activities, as previously identified,28–32 as well as on the current interest in short peptides. Studies over the past few decades have identified short peptides as innovative therapeutic agents in various fields of medicine owing to their high pharmacological activity, specificity, safety, and great promise for treating various pathologies.33–36
Despite increasing interest in peptide-based neuromodulators, their role in neuropathic pain remains insufficiently explored. To the best of our knowledge, this study represents the first evaluation of GABA-derived short peptides in a chronic constriction injury (CCI) model of neuropathic pain. Specifically, the study aimed to investigate the antinociceptive effects of peptide–pregabalin combinations using in vivo behavioral assays and to explore potential mechanisms underlying these effects through in silico molecular docking analysis targeting key components of the GABAergic system. By integrating behavioral and computational approaches, this study provides preliminary evidence supporting the potential of GABA-derived peptides as adjuvant candidates for multimodal neuropathic pain therapy.
Materials and methods
In vivo studies
Experimental animals
Male inbred white rats (180–250 g, n = 24) were used in the experiments. The animals were kept under standard laboratory conditions, which included a 12-h light/dark cycle, a controlled temperature of 22 °C ± 1 °C, and free access to water and food. The experimental procedures were performed between 09:00 a.m. and 02:00 p.m. to reduce circadian influences on animal responses. All experiments were conducted in accordance with Directive 2010/63/EU concerning the protection of animals used for scientific purposes 37 and approved by the Ethics Committee of Yerevan State Medical University (approval no. 3-6/2023).
Induction of neuropathic pain in rats by chronic constriction injury
Chronic constriction injury (CCI) of the sciatic nerve in rats was performed to induce peripheral NP. 38 All possible measures were taken to minimize animal suffering and limit the number of animals required for the experiments. Surgical procedures were performed under anesthesia and the animals were monitored regularly for signs of distress. No unexpected death occurred during the study period.
Briefly, under deep anesthesia with chloral hydrate (400 mg/kg, intraperitoneal), the hair of the rat’s lower back and thigh was shaved off, and a skin incision was made on the lateral surface of the left thigh. To expose the sciatic nerve, an incision was made directly through the connective tissue between the gluteus superficialis muscle and the biceps femoris. The surgical procedures were performed under sterile conditions. After exposing the sciatic nerve, four weak ligatures were applied at an interval of 1 mm with a chromic gut suture (4–0). The skin was closed with a silk suture (6–0), followed by topical antibiotic application.
Behavioral examination
Antinociceptive activity was assessed by behavioral examination using the hot plate test as described by Eddy and Leimbach. 39 Pain behavior caused by thermal hyperalgesia on the surface of the hot plate was measured on the day before CCI surgery (on the day 0) in non-operated rats (pre-CCI baseline threshold) and on the third, fifth, and seventh days in CCI-subjected rats (post-CCI days). The rats were placed on the surface of the hot plate at a temperature of 50 °C ± 1 °C one at a time. To avoid tissue damage, the cut-off time was maintained at 60 s.
Heat, as a noxious stimulus in the hot plate, produces pain behavioral responses such as paw licking, jumping, and changes in locomotor activity (movements), which are estimated in terms of reaction times (latency) and their frequency during the observation period (60 s). After placing the animals on a hot plate, the following parameters were evaluated:
Licking latency—the latent period from the position of the animals on the hot plate to the licking of the hind paw,
Jumping latency—the latent period from the position of the animals on the hot plate to jumping off the plate,
Latency of movements—the latent period from the position of animals on the hot plate to the beginning of movements on a plate,
Licking frequency—the number of licks during the observation time on the hot plate,
Jumping frequency—the number of jumps during the observation time on the hot plate,
Number of movements—the number of movements in an observation time on the hot plate (general locomotor activity of animals).
Experimental protocol
In the experiment, the sample size was chosen at six animals per group based on commonly used group sizes in similar preclinical studies. Animals were allocated to the experimental groups in a structured manner to ensure comparable baseline characteristics; however, formal randomization procedures were not implemented.
The animals underwent an examination of pain behavior on the hot plate before NP modeling on day 0. The next day, neuropathic pain was induced by surgical intervention of the sciatic nerve with CCI, and after modeling, changes in pain behavior were observed on postoperative days 3, 5, and 7 (Figure 1). Although formal blinding was not performed, behavioral parameters were recorded using predefined criteria to minimize observer bias.

Experimental timeline of the study design. In the experiment, all animals (n = 24 rats) were tested for pain behavior in accordance with various characteristics (licking latency, licking frequency, jumping latency, jumping frequency, latency of movements, number of movements). Testing was performed at the indicated time point—before the CCI on preoperative day (0 day), and after the CCI on postoperative days (third, fifth, and seventh days). Treatment in all groups was started 24 h after CCI and was carried out twice a day, for 7 days, intraperitoneally, with 0.9% saline solution (in group 1—control) and combinations of each short peptide with pregabalin, both at a dose of 20 mg/kg (in groups 2, 3, and 4).
After 24 h from the CCI, animals were administered as follows:
Group 1 (control)—0.9% saline solution.
Group 2—combination of 20 mg/kg pGlu-GABA and 20 mg/kg PGB.
Group 3—combination of 20 mg/kg pGlu-GABA ethyl ester and 20 mg/kg PGB.
Group 4—combination of 20 mg/kg pGlu-diGABA and 20 mg/kg PGB.
The study design did not include single-compound treatment groups as their effects had already been identified in our previous series of experiments, 29 thus avoiding unnecessary animal use. All injections were administered intraperitoneally twice daily for 7 days. Short peptides and pregabalin were freshly dissolved in 0.9% saline solution and co-administered as a mixture in equal doses of 20 mg/kg, at a volume of 0.2 mL/100 g body weight of rats.
The PGB dose was selected based on its dose-efficacy relationship in various animal models of pain, as listed in the literature as a low effective dose. 40 The 20 mg/kg dose of the investigated short peptides was chosen based on our previous studies.28–32
In silico studies
Molecular docking analyses of protein–ligand interactions were performed using the AutoDock Vina docking program41,42 and AutoDock MGL Tools 1.5.6. PyMOL (PyMOL Molecular Graphics System, Version 2.0; Schrödinger, LLC) and Discovery Studio 2020 were used to visualize and modify the structures of the target proteins and ligands. Complete chemoinformation, preparation of the chemical structures of the ligand molecules, and file format conversion (mol2, sdf, pdb, pdbqt) were performed using ChemSketch, Open Babel 3-1.1, Discovery Studio 2020, and AutoDock MGL Tools 1.5.6. The crystal structures of the target proteins were downloaded from the RCSB Protein Data Bank (http://www.rcsb.org/) and encoded with the PDB codes.
Protein optimization was carried out by AutoDock MGL Tools 1.5.6 to separate inhibitors from the active sites, remove all water molecules from the proteins, prepare the required files for AutoDock Vina by assigning hydrogen polarities, and convert protein structures from the PDB file format to PDBQT format. A grid box for docking calculation was defined for each target protein, and the size and center coordinates of each grid box were determined.
After docking, AutoDock MGL Tools 1.5.6., Discovery Studio 2020, LigPlot+ v.2.2.5, and a protein–ligand interaction profiler (PLIP; https://plip-tool.biotec.tu-dresden.de/plip-web/plip/index, accessed on May 1, 2023) were used to visualize and analyze the output files. Ligand-binding affinities were predicted as negative Gibbs free energy (∆G), which were calculated based on the AutoDock Vina scoring function, and expressed as ligand–protein binding forces in kilocalories per mole (kcal/mol). The best conformation was selected from nine interaction models, which were characterized by the highest negative value of the binding energy, since in this case, the strongest binding force was observed. 43 Subsequently, the binding poses of each ligand were observed, their interactions with the protein were characterized, and the best and most energetically favorable conformations of each ligand were selected.
Post-docking analyses showed the sizes and locations of binding sites, hydrogen-bond interactions, hydrophobic interactions, and binding distances as interaction radii of <5 Å from the position of each docked ligand by the Protein–Ligand Interaction Profiler (PLIP) 44 online tools.
In this study, the GABA-B receptor (PDB code: 4MS4), 45 GAT1 (PDB code: 7Y7W), 46 and GABA-AT (PDB code: 1OHV) 47 were selected as target proteins, as they are important structures in GABAergic synaptic neurotransmission, and the GABA signaling system was proposed as a target for possible therapies aimed at relieving neuropathic pain. 48
The grid box of GAT1 for the docking calculation was centered on the active site of the protein (x = 101.054, y = 108.340, z = 107.739) and X = 50 Å, Y = 50 Å, Z = 50 Å size, encompassing all the amino acid residues. The configuration file defines the coordinates as exhaustiveness = 10 and energy ranges = 4.
The grid box of GABA-B receptors was centered on the active site of the protein (x = −23.286, y = 14.324, z = −29.638) and x = 60 Å, y = 60 Å, z = 60 Å size, with exhaustiveness = 10 and energy range = 4.
For GABA-AT, the grid size was set at 60 × 60 × 60 Å (x, y, and z) points, and the grid center was designated as x = 6.362, y = 0.796, and z = 27.875, with an energy range of 4 and exhaustiveness of 10.
The investigated ligand molecules were pGlu-GABA, pGlu-GABA ethyl ester, and pGlu-diGABA short peptides, and GABA, baclofen, tiagabine, and vigabatrin were selected as reference molecules.
Drugs and chemicals
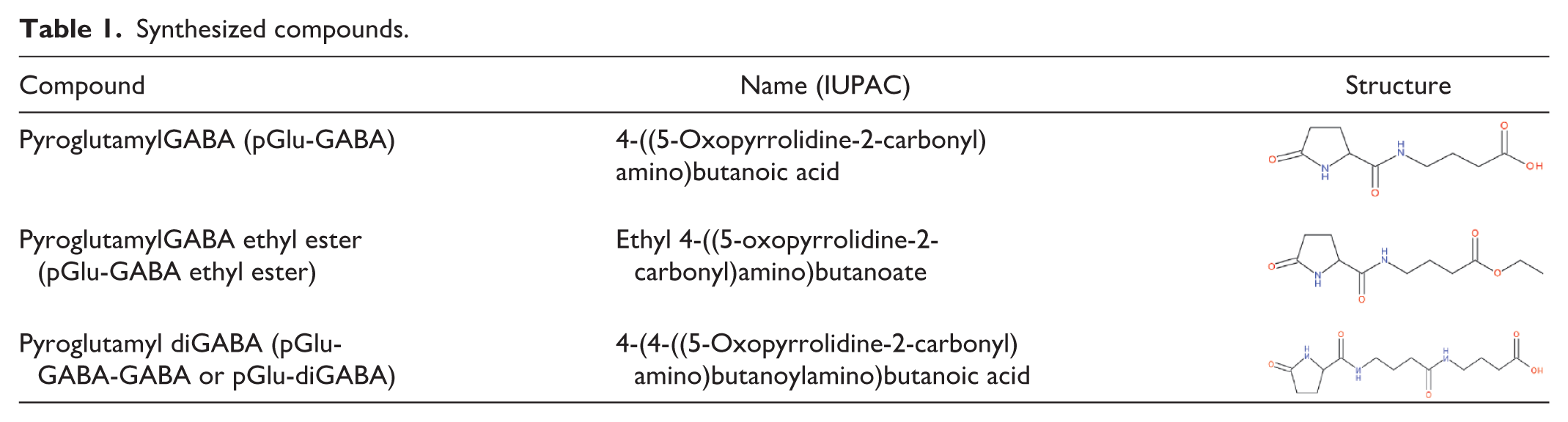
In this study, PGB was provided by the Scientific Center of Drug and Medical Technologies JSC of Armenia (SCDMTE), pGlu-GABA, pGlu-GABA ethyl ester, and pGlu-diGABA were synthesized in the scientific research laboratory “AZAD Pharmaceuticals LLC” (Yerevan, Armenia; Table 1).
Synthesized compounds.
Statistical data analysis
The results are presented as the mean ± standard error of the mean (SEM) of six animals per group. Data were analyzed using IBM SPSS Statistics version 27 and Microsoft Excel 2010. Comparisons of means were performed using two-way ANOVA, followed by Bonferroni’s test. p < 0.05 was considered statistically significant.
Results
Hot plate results
Combinations of the pGlu-GABA, pGlu-GABA ethyl ester, and pGlu-diGABA short peptides with PGB were tested in a rat model of CCI-induced NP. To evaluate the analgesic properties of the therapy, changes in pain behavior in the treated animals were compared with those in the control animals (group 1, n = 6). In these combinations, the dose of PGB was chosen based on the dose/efficacy ratio reported in animal pain models. 49 Here, a low effective dose of PGB was used in combinations to assess the possibility of additional effects.50,51 The peptide dose was selected based on our previous experiments.28–32
Thus, in the case of the combined administration of pGlu-GABA and pregabalin, the maximum increase in licking latency and the maximum decrease in licking frequency in animals were registered on the third day after injury compared with the pre-CCI values, as well as compared with the third day data in the control group (Figure 2(a) and (b)).
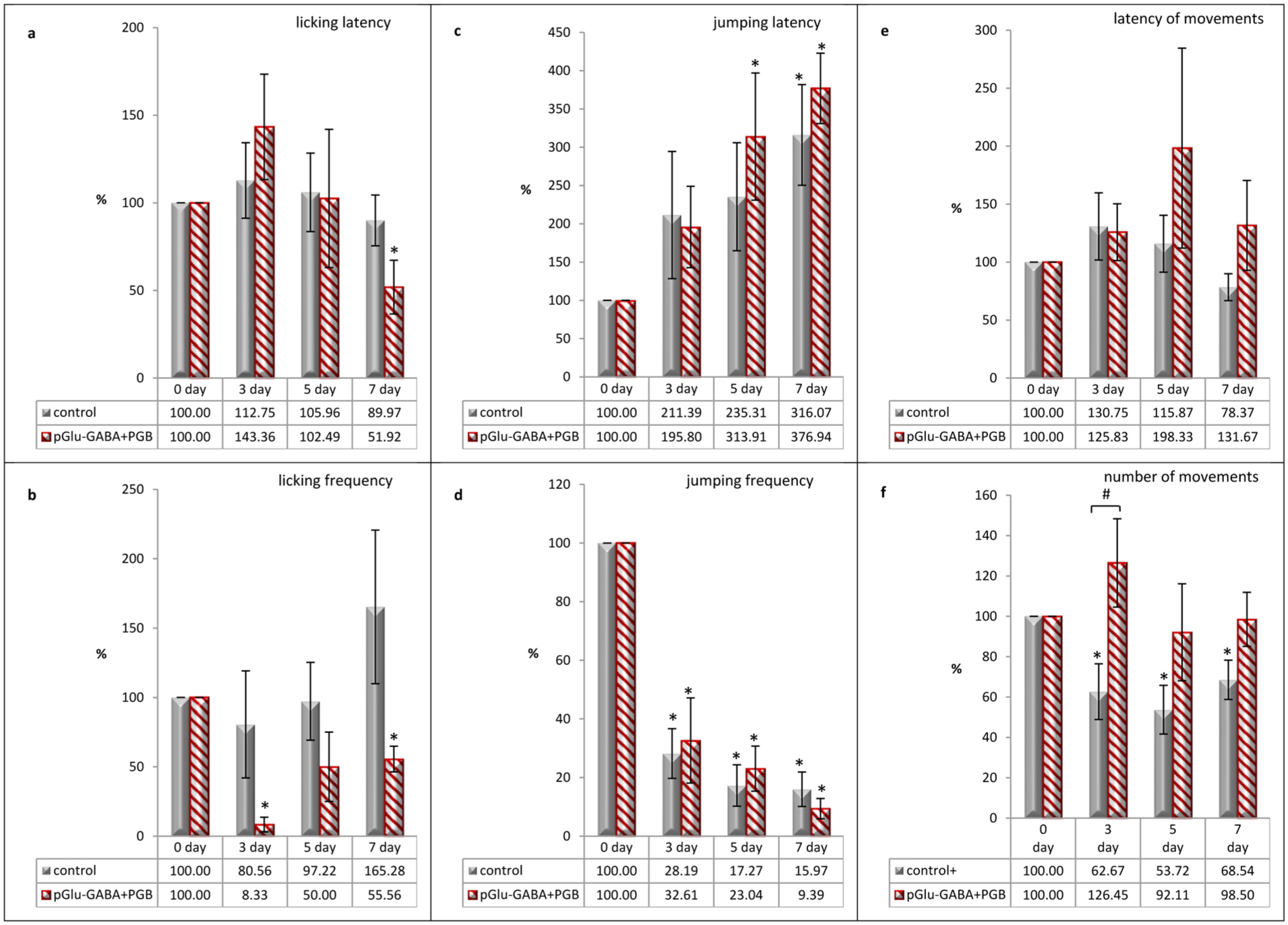
Effects of pGlu-GABA and PGB co-administration on the pain-related behaviors in rats in a neuropathic pain model caused by CCI, assessed using the hot plate test: (a) changes in licking latency, (b) changes in licking frequency, (c) changes in jumping latency, (d) changes in jumping frequency, (e) changes in latency of movements, and (f) changes in number of movements. Data are presented as a percentage of baseline values (day 0) for each day of observation (third, fifth, and seventh post-CCI days) and expressed as the mean ± SEM (n = 6 rats/group). *p < 0.05 versus day 0 (within-group comparison) and #p < 0.05 versus the control group (between-group comparison). Animals in the control group were injected with 0.9% saline solution at a volume of 0.2 mL/100 g body weight, twice a day for 7 post-CCI days, intraperitoneally. Animals in the treated group were injected with a combination of pGlu-GABA and PGB at a dose of 20 mg/kg of each compound, twice a day for 7 post-CCI days, intraperitoneally.
With the same treatment, the jumping latency was extended compared to the initial values on all days of observation, and even exceeded the results of the control group on the fifth and seventh days (Figure 2(c)). In the same group, a decrease in jumping frequency was recorded on all days of observation, but a more pronounced decrease was recorded only on the seventh day, compared with the control group (Figure 2(d)).
The locomotor activity (latency of movements and number of movements) after the combined administration of dipeptide and PGB differed markedly from that of the control group, indicating a weaker pain response. Thus, against the background of the prolonged latency of movements on the fifth and seventh days, compared with the control group, an increase in locomotor activity was recorded almost to the initial values of the indicator and even exceeded this value on the third day (Figure 2(e) and (f)).
Based on these results, it can be assumed that when pGlu-GABA is combined with PGB, there is a decrease in the pain behavior of animals without a reduction in their locomotor activity.
According to the results of the combined administration of pGlu-GABA ethyl ester with PGB, although the latency of licking did not increase compared with the control group (Figure 3(a)), a decrease in the frequency of licking on all observation days was recorded, compared with the initial values and corresponding control group data, which indicated a decrease in pain (Figure 3(b)).
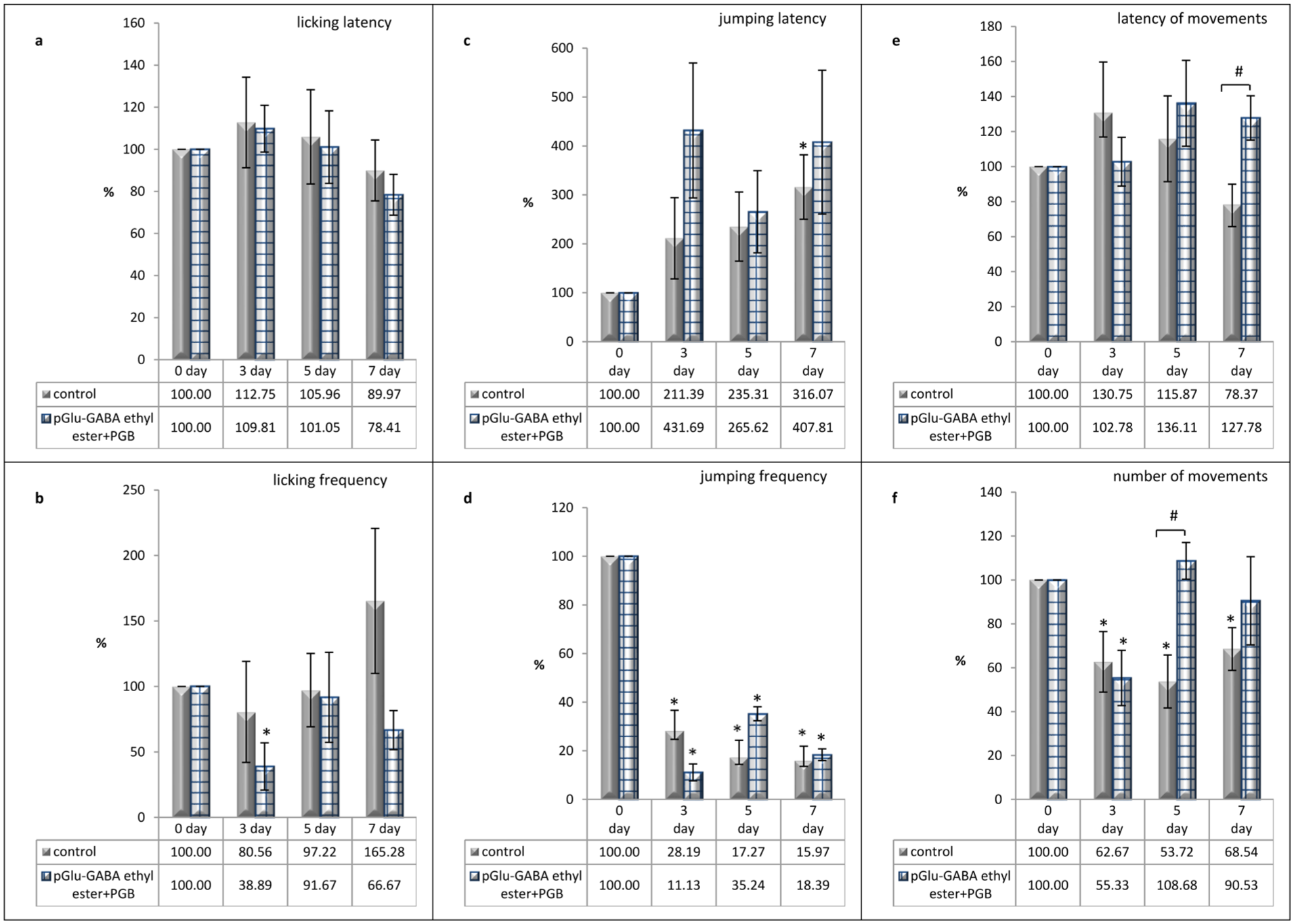
Effects of pGlu-GABA ethyl ester and PGB co-administration on the pain-related behaviors in rats in a neuropathic pain model caused by CCI, assessed using the hot plate test: (a) changes in licking latency, (b) changes in licking frequency, (c) changes in jumping latency, (d) changes in jumping frequency, (e) changes in latency of movements, and (f) changes in number of movements. Data are presented as a percentage of baseline values (day 0) for each day of observation (third, fifth, and seventh post-CCI days) and expressed as the mean ± SEM (n = 6 rats/group). *p < 0.05 versus day 0 (within-group comparison) and #p < 0.05 versus the control group (between-group comparison). Animals in the control group were injected with 0.9% saline solution at a volume of 0.2 mL/100 g body weight, twice a day for 7 post-CCI days, intraperitoneally. Animals in the treated group were injected with a combination of pGlu-GABA ethyl ester and PGB at a dose of 20 mg/kg of each compound, twice a day for 7 post-CCI days, intraperitoneally.
In the changes in jumping latency, the observed indicator exceeded the initial and control group values on all days of registration (Figure 3(c) and (d)). At the same time, both the maximum increase in latency and the maximum decrease in the frequency of jumps were recorded on the third day of observation, compared with the control group.
Under the same conditions, there was a noticeable increase in locomotor activity due to the number of movements, particularly on days 5 and 7, compared to the control group (Figure 3(f)). Moreover, on the same days of observation, an extension in movements latency was recorded, whereas in the control group, the changes in the observed indicator showed a downward tendency (Figure 3(e)).
Thus, the combination of pGlu-GABA ethyl ester and PGB produced an analgesic effect, as assessed by pain behavior.
Monitoring of changes in licking latency in the group of animals treated with pGlu-diGABA and pregabalin did not reveal any increase in licking latency or decrease in licking frequency compared with the control group (Figure 4(a) and (b)). However, there was a dynamic increase in licking latency from the third to the seventh postoperative days in the treated animals, whereas the reverse dynamic of changes was recorded in the control group.

Effects of pGlu-diGABA and PGB co-administration on the pain-related behaviors in rats in a neuropathic pain model caused by CCI, assessed using the hot plate test: (a) changes in licking latency, (b) changes in licking frequency, (c) changes in jumping latency, (d) changes in jumping frequency, (e) changes in latency of movements, and (f) changes in number of movements. Data are presented as a percentage of baseline values (day 0) for each day of observation (third, fifth, and seventh post-CCI days) and expressed as the mean ± SEM (n = 6 rats/group). *p < 0.05 versus day 0 (within-group comparison) and #p < 0.05 versus the control group (between-group comparison). Animals in the control group were injected with 0.9% saline solution at a volume of 0.2 mL/100 g body weight, twice a day for 7 post-CCI days, intraperitoneally. Animals in the treated group were injected with a combination of pGlu-diGABA and PGB at a dose of 20 mg/kg of each compound, twice a day for 7 post-CCI days, intraperitoneally.
When the effect of the combination of pGlu-diGABA with PGB on jumping was examined, it was found that, while the latency of jumping in the treated animals was increased compared to that in the control group on days fifth and seventh (Figure 4(c)), the frequency of jumping did not differ significantly between these two groups (Figure 4(d)).
A comparison of the changes in the locomotor activity of animals treated with a combination of pGlu-diGABA and PGB showed that the maximum prolongation of the latency of movements was recorded on the seventh day of observations (Figure 4(e)), and a significant increase in the number of movements was observed on the fifth day, compared with the same indicator in the control group (Figure 4(f)). The results indicate that the analgesic effect of the combination of pGlu-diGABA and PGB in the NP model due to changes in jumping and locomotor activity is characterized, especially under conditions of prolonged treatment.
Docking analysis results
The experimental results were supplemented in silico by molecular docking studies, which revealed protein–ligand interactions by analyzing the binding abilities of pGlu-GABA, pGlu-GABA-ethyl ester, and pGlu-diGABA studied short peptides (ligands) with target molecules (proteins) in the GABAergic system, such as the GABA transporter type 1 (GAT1 transporter), GABA-AT enzyme, and GABA-B receptors (Table 2), since their activity is crucial for GABAergic synaptic transmission. As a reference structures, the native ligand GABA was selected for all targets, as well as tiagabine for GAT1 as the main inhibitor of the transport protein, vigabatrin for GABA-AT as the main inhibitor of the enzyme, and baclofen for GABA-B as an agonist of the receptors.
Binding energy in the GAT1, GABA-AT, and GABA-B targets.

By the docking of each protein–ligand interaction, the structure of the active site in the binding pocket was revealed, the amino acids included in the site were noted, the distance of interactions and the type of bonds between the ligand and amino acid residues were established, and an energetically favorable conformation (complex-1) based on the binding affinity (expressed in ΔG kcal/mol) was marked. Moreover, the protein–ligand interactions of the reference molecules and the studied GABA-derived short peptides were compared to predict possible similarities in interactions.
Based on the fact that in the GAT1 transporter, the active sites of GABA (Figure 5(a)) and tiagabine (Figure 5(b)) have different localizations, the binding pockets of the studied peptides in GAT1 were analyzed from the point of view of interpreting their possible competitive (GABA-like) and non-competitive (tiagabine-like) interactions.

Binding conformations of the reference molecules and the investigated short peptides in the active site of GAT1: (a) interaction of the reference molecule GABA in complex-1, (b) interaction of the reference molecule tiagabine in complex-1, (c) interaction of pGlu-GABA in complex-1, (d) interaction of pGlu-GABA in complex-2, (e) interaction of pGlu-GABA ethyl ester in complex-1, (f) interaction of pGlu-GABA ethyl ester in complex-6, and (g) interaction of pGlu-diGABA in complex-1.
From the interaction models of short peptides, not only the energetically advantageous complexes-1 of GAT1—pGlu-GABA (Figure 5(c)), GAT1—pGlu-GABA ethyl ester (Figure 5(e)), and GAT1—pGlu-diGABA (Figure 5(g)) were analyzed, but also GAT1—pGlu-GABA complex-2 (Figure 5(d)), and GAT1—pGlu–GABA ethyl ester complex-6 (Figure 5(f)).
In complex-1 of the GAT1—pGlu-GABA interaction (Figure 5(c)), the ligand was fixed at a location other than the binding sites of the reference molecules, which was also observed in the GAT1—pGlu-diGABA interaction complex-1 (Figure 5(g)). A tiagabine-like binding pocket was formed in GAT1—pGlu-GABA ethyl ester complex-6 (Figure 5(f)). In other protein–ligand models, although the interactions do not occur in the same active center as in the GABA, they have a localization close to it.
It was found that the investigated peptides could form GABA-like and tiagabine-like binding pockets with GAT1. These types of interactions may enable the prediction of competitive or non-competitive inhibition of the GABA transporters, resulting in an increase in the synaptic level of GABA and the subsequent pain relief.
The protein–ligand interactions of GABA-B receptors with the pGlu-GABA, pGlu-GABA-ethyl ester, and pGlu-diGABA studied compounds were analyzed and compared with the complexes-1 of the reference molecules GABA (Figure 6(a)) and baclofen (Figure 6(b)). In experimental and clinical practice, baclofen effectively relieves NP as an orthosteric agonist of the metabotropic GABA-B receptors, which are widely present in areas involved in pain processing. 52

Binding conformations of the reference molecules and studied short peptides in the GABA-B active site: (a) interaction of GABA in complex-1, (b) interaction of baclofen in complex-1, (c) interaction of pGlu-GABA in complex-1, (d) interaction of pGlu-GABA in complex-4, (e) interaction of pGlu-GABA ethyl ester in complex-1, and (f) interaction of pGlu-diGABA in complex-1.
From the top nine models of GABA-B and pGlu-GABA interactions, complex-1 was selected, which showed a GABA-like binding ability (Figure 6(c)), and complex-4, which demonstrated baclofen-like binding ability (Figure 6(d)).
In the models of GABA-B receptors with pGlu-GABA ethyl ester and pGlu-diGABA, GABA-like interactions were indicated in complexes-1 (Figure 6(e) and (f)). This findings may support the assumption that peptides could activate GABA-B receptors in a GABA-like manner.
The results of the molecular docking of the investigated short peptides with the GABA-AT enzyme were analyzed and compared with the docking of the reference molecules GABA (Figure 7(a)) and vigabatrin (Figure 7(b)).

Binding conformations of the reference molecules and studied short peptides in the GABA-AT active site: (a) interaction of GABA in complex-1, (b) interaction of vigabatrin in complex-1, (c) interaction of pGlu-GABA in complex-1, (d) interaction of pGlu-GABA ethyl ester in complex-1, and (e) interaction of pGlu-diGABA in complex-1.
Complexes-1 for each protein–ligand interaction were selected from the top nine ligand poses (Figure 7(c)–(e)). In all binding pockets of short peptides and the GABA-AT enzyme, amino acid residues were fixed, which were not included in the active center of GABA, but were present in the vigabatrin active site.
Therefore, it was hypothesized that modulation of the GABA-AT enzyme can be realized by the GABA-derived short peptides, which may contribute to the reduction of NP by increasing the level of GABA in the synaptic cleft. These results allowed us to conclude that all studied compounds interactions with the GABA-AT enzyme were created close to the active center of vigabatrin, which was chosen as the reference molecule because it is an FDA-approved inhibitor of human GABA-AT. 53 These findings suggest that the studied GABA-derived short peptides may serve as promising therapeutic agents for targeting GABA-AT and reducing NP.
These results suggested that there was no competition between the studied compounds and GABA for the active center of GABA-AT. However, interactions around the binding pocket of vigabatrin were registered in all models, suggesting that the studied peptides appear to have vigabatrin-like interactions and have an inhibitory effect on GABA-AT.
Discussion
This study demonstrated that short peptides derived from GABA and pyroglutamic acid, when administered in combination with PGB, produce significant antinociceptive effects in a CCI-induced NP model.
The investigated peptides are novel compounds synthesized by our group, and their chemical design and detailed characterization have been reported previously.28–32 The selected dose was determined during preliminary experimental studies. Previous studies demonstrated the antinociceptive activity of these peptides and PGB in acute nociception models using the tail flick test.29,32 Both agents, administered at 20 mg/kg, produced antinociceptive effects as monotherapies, whereas their combined administration resulted in an augmentation of the effect compared to individual treatments. 29 These findings provided a rationale for the presented study, which focused on evaluating peptide–PGB combinations in the NP model. Combination pharmacotherapy is increasingly recognized as an effective strategy for pain management, as it may enhance therapeutic efficacy while reducing the required doses and limiting drug-related adverse effects.49,54 While both the tail flick and hot plate tests evaluate responses to thermal stimuli, they involve partially different underlying mechanisms and may reflect distinct aspects of nociceptive processing.
Behavioral assessments conducted during the established phase of NP following CCI, which is characterized by stable thermal hypersensitivity, showed that peptide–PGB combinations consistently attenuated nociceptive responses, as evidenced by the increased latency and reduced frequency of nociceptive responses. Notably, different peptide combinations produced distinct behavioral profiles, suggesting that structural differences among peptides may influence their pharmacological activity.
Importantly, the evaluated combinations did not exacerbate the motor suppression commonly associated with PGB treatment. Although locomotor activity increased across treatment groups with delayed movement onset, these findings should be interpreted cautiously, as locomotor parameters alone are insufficient to exclude sedative effects. Nevertheless, the absence of overt motor impairment suggests a potentially favorable safety profile, warranting further investigation using more specific behavioral assays.
Across all tested combinations, antinociceptive effects were evident but varied in magnitude and consistency. The pGlu-GABA–PGB combination demonstrated the most robust effects, particularly on licking parameters during the early postoperative period, whereas the pGlu-GABA ethyl ester–PGB combination predominantly affected jumping behavior, and the pGlu-diGABA–PGB combination exhibited more selective effects on movement latency measures.
Overall, the data suggest that peptide-PGB combinations alleviated NP-related behaviors in the CCI model. It is noteworthy that all the investigated combinations in the NP model did not appear to exacerbate motor suppression associated with PGB therapy.55,56 These findings suggest that peptide-based adjuvants can modulate the pharmacological profile of PGB and improve its therapeutic potential.
Although the hot plate test provides a validated measure of thermal nociception, it does not fully capture the complexity of NP. More specific assays, such as the von Frey and Hargreaves tests, may provide complementary information by assessing mechanical allodynia and thermal hypersensitivity, respectively. The selection of the hot plate test in the present study was based on its established sensitivity to centrally mediated nociceptive responses and its suitability for detecting pharmacological modulation in this experimental setting. Nevertheless, the absence of additional modality-specific assessments limits the generalizability of the findings across different pain modalities.
Furthermore, the 7-day observation period, while sufficient to evaluate early-phase NP following CCI, remains relatively short for a chronic pain model. As such, it does not permit assessment of long-term efficacy, potential tolerance development, or sustained behavioral effects. Future studies should incorporate extended observation periods and additional behavioral paradigms to provide a more comprehensive evaluation of the therapeutic potential of the investigated peptide–pregabalin combinations. In addition, the absence of formal randomization and blinding procedures may have introduced bias, particularly in observer-dependent behavioral assessments. Although predefined criteria were used to standardize the measurements, the possibility of residual bias cannot be excluded.
In addition to traditional in vivo assays, in silico investigations have also been conducted using virtual models of protein–ligand interactions, as a widely used approach to drug discovery.57–59 Considering the well-established role of the GABAergic pathway in pain signaling in response to peripheral nerve damage,60–62 and the presence of a structural fragment of GABA in the molecule of the studied compounds, the possible targets of pharmacological action were selected from GABAergic synapses. In view of this, the binding affinities of pGlu-GABA, pGlu-GABA ethyl ester, and pGlu-diGABA short peptides were analyzed using the GAT1 transporter, GABA-AT enzyme, and GABA-B receptors. GABA-A receptors were not included in the docking analysis because of the structural complexity of ligand-gated ion channels and current limitations in accurately modeling peptide–receptor interactions in such systems. In contrast, G protein-coupled receptors, such as GABA-B, are more amenable to computational docking approaches because of their relatively well-defined binding pockets, thereby supporting their prioritization in the present docking analysis.
The docking analysis suggested that the pharmacological effects of the investigated peptides may be mediated by GABA-B, GAT1, and GABA-AT. Although molecular docking results provide valuable insights into the potential mechanisms of action, they remain predictive in nature and require experimental validation.
This study provides the first evidence of the analgesic potential of the investigated peptide–PGB combinations in a CCI NP model. Some methodological limitations should be considered, including the absence of monotherapy groups, the short observation period, and the lack of formal randomization and blinding, which may introduce observer bias despite standardized assessment criteria.
The integration of in vivo behavioral data with in silico docking analysis provides a multimodal perspective on the potential mechanisms of action of these GABA-derived peptides. Taken together, these findings support the concept that peptide-based adjuvants may enhance the pharmacological profile of established analgesics and represent a promising strategy for developing multimodal therapies for NP.
Conclusions
This is the first study to demonstrate the therapeutic potential of investigated peptides against neuropathic pain. When combined with PGB, GABA-derived short peptides produced antinociceptive effects in a rat model of NP. These combinations attenuated pain-related behaviors without inducing the pronounced motor suppression associated with PGB treatment.
These findings suggest that peptide-based adjuvants may improve the therapeutic profile of PGB by enhancing its efficacy while limiting its adverse effects. However, further preclinical studies are required to clarify the mechanisms of action, evaluate the long-term efficacy, and determine the safety profile of these peptide–PGB combinations.
Patents
Naira H Adamyan, Marine G Balasanyan, Hakob V Topchyan, Varduhi H Poghosyan, ML Movsisyan, TA Margaryan, and LJ Nikoghosyan (GABA and pyroglutamic acid peptides with antinociceptive and analgesic activity). RA. Patent N3400A, IPC index C07D 207/00, AM20200039, August 17, 2020.
Footnotes
Abbreviations
The following abbreviations are used in this manuscript:
GABA Gamma-aminobutyric acid
PGB Pregabalin
NP Neuropathic pain
CCI Chronic constriction injury
pGlu-GABA PyroglutamylGABA
pGlu-GABA ethyl ester PyroglutamylGABA ethyl ester
pGlu-diGABA Pyroglutamyl diGABA
GAT1 GABA transporter type 1
GABA-AT GABA aminotransferase
SEM Standard error of the mean
ANOVA Analysis of variance
ΔG Gibbs free energy
PDB Protein Data Bank
Author contributions
All the authors have read and agreed to the published version of this manuscript. Conceptualization: Naira H Adamyan and Marine G Balasanyan. Methodology: Naira H Adamyan, Marine G Balasanyan, Hakob V Topchyan, Arusyak G Zhamharyan, and Tigran K Davtyan. Software: Arusyak G Zhamharyan and Tigran K Davtyan. Validation: Naira H Adamyan, Marine G Balasanyan, and Hakob V Topchyan. Formal analysis: Naira H Adamyan and Marine G Balasanyan. Investigation: Naira H Adamyan and Marine G Balasanyan. Resources: Naira H Adamyan and Marine G Balasanyan. Data curation; Naira H Adamyan and Marine G Balasanyan. Writing—original draft preparation: Naira H Adamyan and Marine G Balasanyan. Writing—review and editing: Naira H Adamyan, Marine G Balasanyan, Varduhi H Poghosyan, Knarik M Shamilyan, Sona H Grigoryan, Anush G Tananyan, Alisa A Chitchiyan, and Maria K Verdyan. Visualization: Arusyak G Zhamharyan and Tigran K Davtyan. Supervision; Naira H Adamyan and Marine G Balasanyan. Project administration, Marine G Balasanyan.
Data availability statement
The data presented in this study are available upon request from the corresponding author owing to privacy concerns.
Declaration of conflicting interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The authors received no financial support for the research, authorship, and/or publication of this article.
Institutional review board statement
The animal study protocol was approved by the Yerevan State Medical University Ethics Committee (no. 3-6/2023).
Informed consent statement
Not applicable.