
Abstract
Loneliness is a negative and distressing emotional state that arises from a discrepancy between one’s desired and achieved levels of social connectedness. The evolutionary theory of loneliness (ETL) posits that experiencing loneliness is an inherited adaptation that signals that salutary social relations are endangered or damaged and prompts people to reconnect to significant others. The basic tenets of the ETL has led researchers to examine the genetic underpinnings of loneliness. The current review provides an updated overview of genetic studies on loneliness and discusses the importance of genetic research for the ETL. The most recent studies suggest that the many genes that contribute to a small degree to differences in loneliness partially overlap with genes that contribute to neuroticism, but not with depression. In addition, the genetic studies discussed in this review show that genes are unlikely to have a direct effect on loneliness. Instead, environmental factors determine in a dynamic fashion how genes that contribute to loneliness are expressed. Future research on epigenetic processes, such as DNA methylation, can further elucidate the dynamic interplay between genes and the environment and how this interplay contributes to loneliness.
Loneliness is a negative and distressing emotional state that arises from a discrepancy between one’s desired and perceived levels of social connectedness (Perlman & Peplau, 1981). A large number of studies examining loneliness have focused on behavior and the environment (Heinrich & Gullone, 2006; Spithoven, Bijttebier, & Goossens, 2017). Although the number of studies examining genetic contributions to loneliness has increased, the available evidence on the role of genes in loneliness is still limited (Goossens et al., 2015). Because neither the environment nor genes operate in isolation (Manuck, 2010), our understanding of loneliness would benefit from increased insight into the complex interaction between genes and the environment. In this review, we focus on the evolutionary theory of loneliness (ETL; J. T. Cacioppo & Patrick, 2008), which assigns a crucial role to genes, the social environment, and their interaction. We describe how research in genetics can explain why some individuals experience feelings of loneliness more quickly than others. In addition, research into genetics can elucidate the association between loneliness on the one hand and neuroticism or depression on the other. Finally, we discuss the theoretical and practical implications of genetic research for the ETL. As research into the genetics of loneliness is still quite preliminary, this review concentrates in large part on directions for future research.
The Loneliness Phenotype
The definition of loneliness, as mentioned above, emphasizes the subjective character of the experience. As a result, it is important to distinguish loneliness from being alone or in solitude, which is an objective state. Being alone or in solitude is not a prerequisite for the experience of loneliness because not everyone feels lonely when they are alone. Loneliness can also be experienced in the presence of other people (e.g., Heinrich & Gullone, 2006). Not surprisingly, the qualitative or subjective appraisals of social relations, and friendships in particular, are more important predictors of loneliness than the quantitative aspects of social relations (Asher & Paquette, 2003; Heinrich & Gullone, 2006; Marangoni & Ickes, 1989; Vanhalst, Luyckx, & Goossens, 2014).
Loneliness has been assessed using two different approaches: unidimensional and multidimensional (Russell, 1982; Shaver & Brennan, 1991). Researchers who adopt the unidimensional approach consider loneliness to be a construct that varies mainly in intensity. They argue that there are important common themes in all experiences of loneliness across various contexts (Russell, 1982, 1996). Researchers who adopt the multidimensional approach, by contrast, distinguish between different types of loneliness as experienced in different relationships. They argue that different relationships (e.g., parents vs. peers) fulfill different social needs (e.g., nurturance and guidance vs. a sense of integration; Weiss, 1973). According to these researchers, a single type of relationship cannot satisfy all needs, and specific forms of loneliness emerge when a specific social need is not being met (DiTommaso & Spinner, 1997; Weiss, 1973). Factor analyses on the various unidimensional and multidimensional measures have revealed that unidimensional measures of loneliness load on a peer-related loneliness factor rather than on a separate, general loneliness factor (Cramer & Barry, 1999; Goossens et al., 2009).
Loneliness has been related to various psychosocial, mental, and physical problems (e.g., Heinrich & Gullone, 2006). The strong relation with other internalizing problems, such as depressive symptomatology (e.g., Mahon, Yarcheski, Yarcheski, Cannella, & Hanks, 2006), has led some researchers to doubt whether loneliness is a distinct construct and to suggest that it represents a symptom of depression. However, increasing evidence suggests that loneliness and depression are two distinct constructs because items of well-established measures of both concepts load on different latent factors (Anderson & Harvey, 1988; J. T. Cacioppo, Hawkley, et al., 2006; Weeks, Michela, Peplau, & Bragg, 1980). Moreover, loneliness and depression are differentially related to constructs such as personality (Vanhalst, Klimstra, et al., 2012), suicidal ideation (Lasgaard, Goossens, & Elklit, 2011), friendship quality and quantity, and self-esteem and happiness (Spithoven, Lodder, et al., 2017). Regarding personality, loneliness consistently shows a positive association with neuroticism and a negative association with extraversion (e.g., Asendorpf & Van Aken, 2003; J. T. Cacioppo, Hawkley, et al., 2006; J. T. Cacioppo, Hawkley, & Thisted, 2010; Vanhalst, Klimstra, et al., 2012). Because extraversion predisposes the individual for positive affect and neuroticism predisposes the individual for negative affect (Costa & McCrae, 1980; Larsen & Ketelaar, 1991), it is not surprising that they are related to a construct such as loneliness, which is characterized by high negative affect.
Theories of Loneliness
Major theoretical approaches to loneliness include the social needs perspective and the cognitive discrepancy model (Heinrich & Gullone, 2006; Marangoni & Ickes, 1989). The social needs perspective posits a direct relationship between objective social deficits and the subjective experience of loneliness (e.g., Heinrich & Gullone, 2006; Marangoni & Ickes, 1989; Sullivan, 1953). Adherents of this perspective claim that relationship-specific types of loneliness arise when a relationship does not satisfy its underlying set of social needs (DiTommaso & Spinner, 1997; Weiss, 1973). These social needs change across the life span, and experiences of loneliness are expected to fluctuate accordingly (Sullivan, 1953).
Unlike the social needs perspective, the cognitive discrepancy model focuses on people’s subjective evaluation of their relationships rather than the needs fulfilled by these relationships (e.g., Heinrich & Gullone, 2006; Marangoni & Ickes, 1989; Russell, Cutrona, McRae, & Gomez, 2012). All individuals are assumed to use an internal standard when evaluating their relationships (de Jong-Gierveld, van Tilburg, & Dykstra, 2006; Marangoni & Ickes, 1989; Perlman & Peplau, 1981; Russell et al., 2012). This internal standard refers to the desired quantity and quality of social relationships (de Jong-Gierveld et al., 2006) and is based in part on a comparison between one’s own social relationships and the relationships of similar others (Russell et al., 2012). Loneliness is thought to emerge when people experience a discrepancy between their internal standard and the perceived quantity or quality of their social relations (de Jong-Gierveld et al., 2006; Marangoni & Ickes, 1989; Perlman & Peplau, 1981; Russell et al., 2012). So the cognitive discrepancy model concentrates on subjective deficits in social relationships rather than objective ones.
Both the social needs perspective and cognitive discrepancy model emphasize the role of the social environment in the experience of loneliness (Marangoni & Ickes, 1989). Moreover, these theories focus on the negative side of loneliness. Loneliness is assumed to be a negative emotion that is associated with negative experiences in one’s social environment. Another theory that has recently gained prominence in research on loneliness is the ETL, which is discussed in detail in the next section. This theory states that both genes and the social environment play an important role in the experience of loneliness. In addition, it focuses on both the negative and positive sides of loneliness.
Evolutionary Theory of Loneliness
Establishing and maintaining social relations with conspecifics is essential for reproduction and the survival of social species, including humans. For example, social groups can share the responsibility for acquiring food, caring for infants, and protecting against predators. To ensure that the individual acts in ways that increase the likelihood of survival, biological warning mechanisms have evolved (J. T. Cacioppo, Cacioppo, & Boomsma, 2014; J. T. Cacioppo & Patrick, 2008). According to the ETL, the perception of being socially isolated (i.e., feeling lonely), even in the company of others, serves as a biological warning system, found across all social species, that signals that salutary social relations are endangered or damaged (J. T. Cacioppo, Cacioppo, Capitanio, & Cole, 2015; J. T. Cacioppo, Cacioppo, Cole, et al., 2015; S. Cacioppo, Capitanio, & Cacioppo, 2014). The aversive nature of loneliness motivates one to repair or replace deficient salutary bonds, and the negative affect relief that results from doing so is rewarding. Moreover, salutary social interactions are positive and rewarding experiences in themselves (J. T. Cacioppo, Hawkley, et al., 2006; S. Cacioppo, 2018). The aversive nature of loneliness and the positive rewarding value of social contact result in a two-fold motivation to resolve loneliness.
According to the ETL, however, the neurocognitive and adaptive behavioral effects of loneliness do not stop there. Intraspecies aggression represents a significant threat to reproductive success among anthropoid primates (e.g., Dunbar, 2011; Silk, 2007), especially humans (Georgiev, Klimczuk, Traficonte, & Maestripieri, 2013; Harari, 2015). The unfettered motivation to form trusting relationships with others in such contexts could therefore prove fatal. The ELT specifies that the perception of being socially isolated (e.g., absent mutual aid and protection) is not only deleterious for one’s health and well-being but also dangerous (J. T. Cacioppo & Patrick, 2008). Accordingly, the ELT posits that loneliness not only increases a motivation to attend to and approach others but also promotes an emphasis on short-term self-preservation, including (a) an increase in implicit vigilance for social threats and, at least in humans, (b) an increased concern for the individual’s own interests and welfare (i.e., increase in self-centeredness). There is now some evidence for both hypotheses (J. T. Cacioppo, Chen, & Cacioppo, 2017; S. Cacioppo et al., 2016; Spithoven, Bijttebier, et al., 2017). The resulting cooccurrence of increased approach motives with increased avoidance motives allows for a behavioral strategy that reduces the likelihood of negative social events while also increasing the likelihood of positive ones (Nikitin & Freund, 2008).
According to the ETL, practices such as shunning, ostracism, or time-out have been developed to facilitate the socialization of new members to a group or culture. For instance, when someone (e.g., a child) violates a social norm (e.g., sharing and playing nicely with other children), the reactions of significant others (e.g., placing the child in time-out or shunning) can increase the person’s perception of social isolation. The resulting aversive reaction, in turn, contributes to the person adopting more socially appropriate behavior in the future (J. T. Cacioppo & Patrick, 2008).
The ETL also posits that loneliness is influenced by both environmental factors and genetic inheritance (J. T. Cacioppo et al., 2014; J. T. Cacioppo & Patrick, 2008). The latter term refers to the process by which parents pass genetic information on to their offspring. Because loneliness serves an adaptive function as a signal that prompts individuals to reconnect with significant others, genetic variants that influence loneliness are maintained in the population through this intergenerational transmission. This line of reasoning implies that the genetic contribution to loneliness is not limited to the evolution of a single gene. Rather, the ETL predicts that multiple adaptive benefits are derived from the formation and maintenance of social bonds that ensure mutual aid and protection (J. T. Cacioppo et al., 2014; J. T. Cacioppo & Hawkley, 2009). As a result, many genes contribute to loneliness. This proposition is consistent with recent findings that many behavioral phenotypes are polygenic (Visscher, Brown, McCarthy, & Yang, 2012).
The ETL further states that some people are more likely to experience loneliness than others. This individual variability in loneliness is affected by both genes and the environment or, to be more precise, by their interaction. This interaction is described in psychology as differential susceptibility to the environment and is captured in biology and evolutionary genetics by the concept of reaction norm. According to the differential susceptibility hypothesis (Belsky, Bakermans-Kranenburg, & Van IJzendoorn, 2007), individuals who carry specific genetic variants are highly sensitive to the environment. They develop negative outcomes (e.g., loneliness) in adverse environments and positive outcomes (e.g., social connectedness) in beneficial environments. By contrast, individuals who do not carry these specific genetic variants are less sensitive to the environment. They show the same outcomes regardless of the adverse or beneficial environments in which they find themselves (Belsky et al., 2007). The reaction norm is the phenotypic expression of a single genotype across a range of environments, which is sometimes also referred to as plasticity (Goldberger, 1979; Manuck, 2010). Figure 1 provides a graphical representation of three reaction norms for three types of individuals with different genetic backgrounds, one highly plastic (i.e., the solid line) and two less plastic ones (i.e., the two dashed lines). What is inherited, according to the ETL, are genes that determine the range of loneliness levels that are exhibited in reaction to the different social environments to which one is exposed. Put simply, it is not the level of loneliness but the reaction norm that is inherited (J. T. Cacioppo et al., 2014).
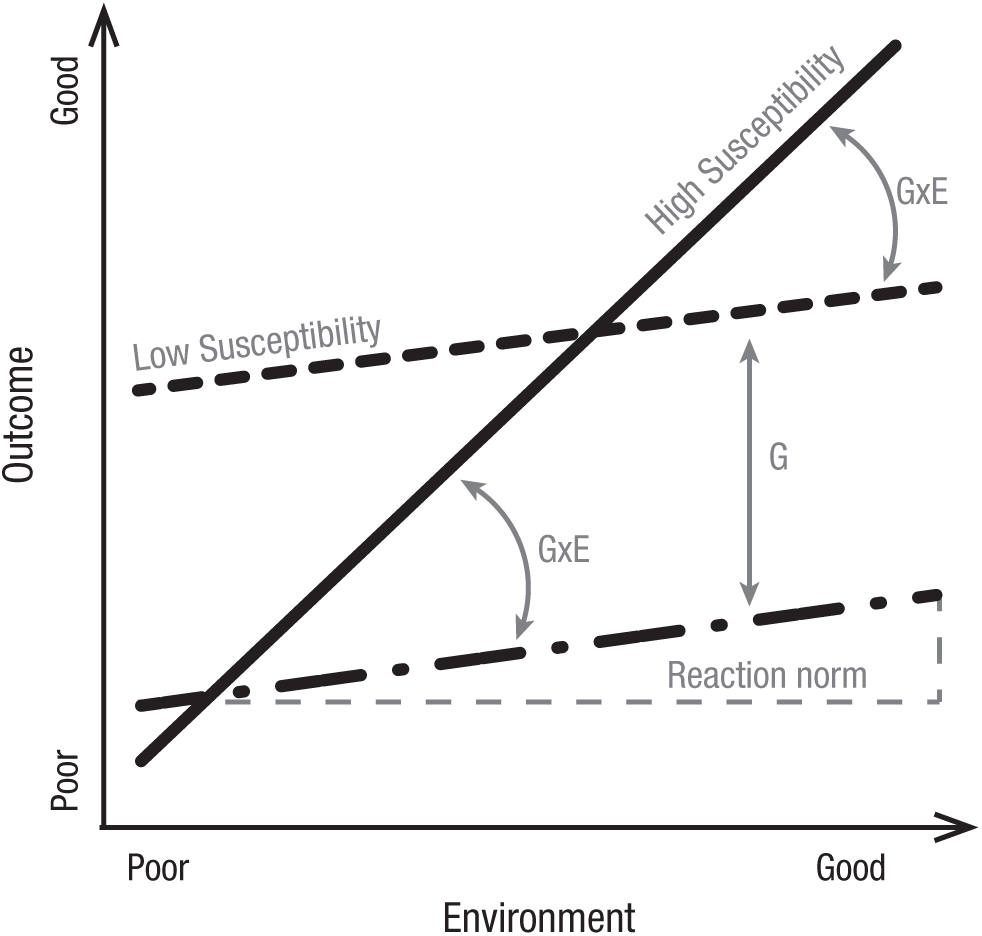
Conceptual representation of reaction norms, genetic effects, Genetic (G) × Environmental (E) effects, and susceptibility to the environment. The lines represent three different genotypes. Reaction norms are the phenotypic expression of a single genotype across a range of environments, which is represented by the slope of the lines. Individuals with a large reaction norm are more susceptible to environmental effects than individuals with a smaller reaction norm. The difference between the two dashed lines, which have a similar slope, represents a genetic effect. The difference between the solid line and the dashed lines represents G × E effects.
According to the ETL, individual variability in an individual’s sensitivity to the aversiveness of and subsequent negativity from loneliness has adaptive effects at the level of the population (J. T. Cacioppo et al., 2014). Individuals who are relatively insensitive to the aversiveness of loneliness are more likely to leave their social group and explore opportunities elsewhere, yet their relatively mild aversion to being absent from their salutary relationships predisposes them to return to the group to share their expertise and insights gained outside the group. Those individuals who are sensitive to the experience of loneliness, by contrast, are more likely to remain in and defend their social group. Moreover, the ETL posits that the depressive behaviors associated with loneliness decrease the likelihood that the individual will forcefully try to regain entry in the social group, and the depressive vocal tone and expressive and postural movements represent a safe call for anyone who is willing to reconnect to do so (J. T. Cacioppo et al., 2014).
Research on rodents provides evidence for the hypothesis that the genetic component of loneliness is, at least in part, the variability in experiencing negativity after the loss of salutary relations (see J. T. Cacioppo et al., 2014). Rat pups respond with a cry when separated from their mother, thereby expressing their distress (Wiedenmayer, Goodwin, & Barr, 2000). A pup’s separation cry elicits searching and retrieving behavior from the mother but might also attract predators. Thus, depending on the environment (i.e., the presence or absence of a predator), the separation cry might be either adaptive or maladaptive. As a result, rat pups show variability in their intensity of crying, in which some pups cry frequently in reaction to isolation whereas others do not (Brunelli & Hofer, 2007). When pups that cry frequently and those who do not cry frequently are selectively bred, after a few generations they will start to show differences in their behavior (Brunelli & Hofer, 2007). These findings suggest that the degree to which pups respond to being separated from their mother is inherited across generations. Similar trends across generations can be assumed to occur in humans (J. T. Cacioppo, Cacioppo, Cole, et al., 2015). Differences in the behavior between the two lines of rats after 25 years of breeding were reminiscent of some of the differences observed in humans who experience high compared with low levels of loneliness: The high relative to low ultrasonic vocalization (USV) line showed more distress to isolation as an infant. In addition, these individuals showed a greater latency to play as adolescents and greater depression-like behaviors, greater anxiety-like behaviors, greater latency to social interactions, greater startle, and diminished learning as adults. In other words, the high USV line indicated anxiousness and passivity, whereas the low USV line indicated exploratory, active, and aggressive behavior (Hofer, 2009).
In summary, according to the ETL, loneliness has an evolutionary adaptive value because it protects the individual against losing salutary social relations that are important for survival. The aversiveness of loneliness motivates people to attend to and approach others to repair and maintain their social relations. However, individuals differ in the extent to which they show a sensitivity to the experience of loneliness. The ETL states that there is a substantial genetic component to this sensitivity in addition to the widely acknowledged environmental component. More specifically, both genes and the environment exert their effects on loneliness through their interaction or interplay, which can be captured by the notions of differential susceptibility to the environment and differential reaction norms.
In this article, we review findings on loneliness from all traditional domains of genetic inquiry and their implications for the ETL. These domains are referred to as behavioral, molecular, and functional genetics, respectively. The main findings of all the studies reviewed are summarized in Table 1.
Overview of the Studies Included in This Review and Their Main Results
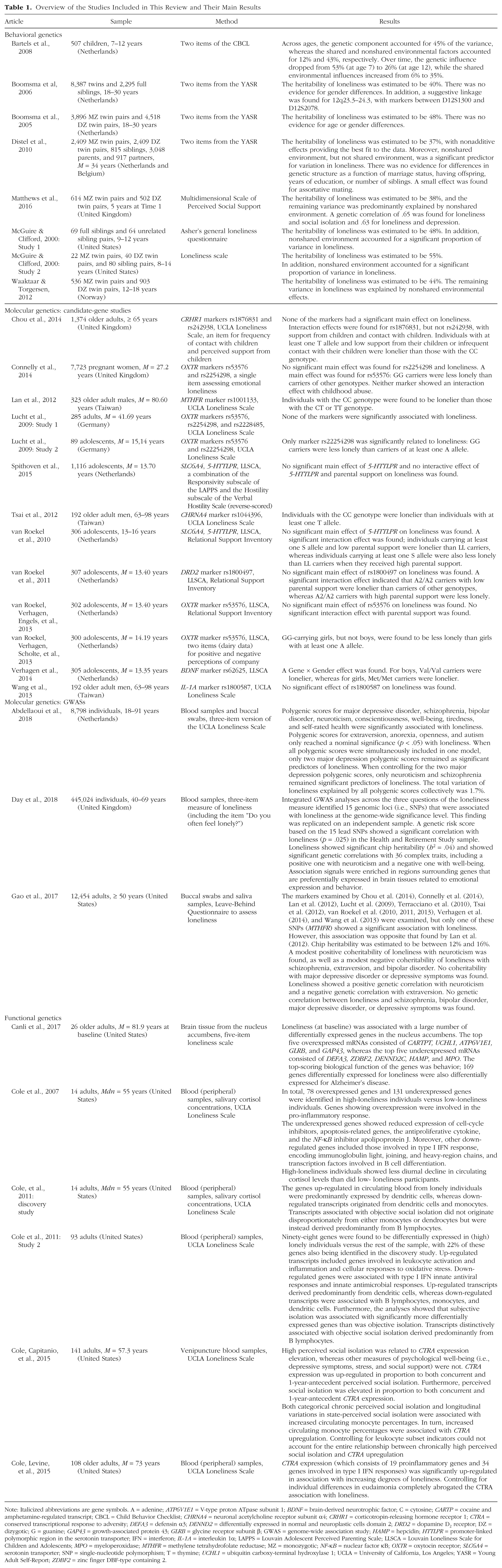
Note: Italicized abbreviations are gene symbols. A = adenine; ATP6V1E1 = V-type proton ATpase subunit 1; BDNF = brain-derived neurotrophic factor; C = cytosine; CARTP = cocaine and amphetamine-regulated transcript; CBCL = Child Behavior Checklist; CHRNA4 = neuronal acetylcholine receptor subunit α4; CRHR1 = corticotropin-releasing hormone receptor 1; CTRA = conserved transcriptional response to adversity; DEFA3 = defensin α3; DENND2 = differentially expressed in normal and neuroplastic cells domain 2; DRD2 = dopamine D2 receptor; DZ = dizygotic; G = guanine; GAP43 = growth-associated protein 43; GLRB = glycine receptor subunit β; GWAS = genome-wide association study; HAMP = hepcidin; HTTLPR = promoter-linked polymorphic region in the serotonin transporter; IFN = interferon; IL-1A = interleukin 1α; LAPPS = Louvain Adolescent Perceived Parenting Scale; LLSCA = Louvain Loneliness Scale for Children and Adolescents; MPO = myeloperoxidase; MTHFR = methylene tetrahydrofolate reductase; MZ = monozygotic; NF-κB = nuclear factor κB; OXTR = oxytocin receptor; SLC6A4 = serotonin transporter; SNP = single-nucleotide polymorphism; T = thymine; UCHL1 = ubiquitin carboxy-terminal hydroxylase 1; UCLA = University of California, Los Angeles; YASR = Young Adult Self-Report; ZDBF2 = zinc finger DBF-type containing 2.
Behavioral Genetics and Loneliness
In behavioral genetics, individual variation is separated into genetic versus environmental components. Broad-sense heritability (h2) is the proportion of phenotypic variance attributable to genetic factors, including the effects of single genes, epigenetics, and gene-gene interactions. The degree of h2 for loneliness, pitting the genome as a whole against the environment as a whole, can be estimated empirically—for instance, through twin and family studies. In this type of research, no DNA has to be collected or analyzed because the aggregate effects of genes are inferred from a comparison of groups with varying degrees of biological relatedness (e.g., monozygotic and dizygotic twins).
Heritability is a proportion, so estimates of heritability range from 0 (no genetic contributions to phenotypic variance) to 1 (genetic factors account for all phenotypic variance; Plomin, DeFries, Knopik, & Neiderhiser, 2013; Visscher, Hill, & Wray, 2008). Heritability is often falsely interpreted as the degree to which genetic factors are responsible for a phenotype rather than the degree to which genetic factors are responsible for phenotypic variation. Heritability refers to the latter, the individual differences observed at a given point in time in a phenotype (Goldberger, 1979; Plomin et al., 2013). For example, although complex adaptations, such as organs and tissues (e.g., heart or skin), are determined by genes, the heritability of the presence or absence of these complex adaptations is zero. These organs and tissues are universal and functionally identical because we would not survive without them. Note that deviations in their functioning or appearance, such as heart-muscle functioning or skin color, do have a heritability greater than zero (J. T. Cacioppo & Freberg, 2018; Plomin et al., 2013; Visscher et al., 2008). It should also be noted that heritability estimates depend on the total effect size of not only genetic factors that contribute to individual differences in a phenotype but also environmental factors. This is because heritability is defined as the proportion of phenotypic variance that results from genetic factors, and total phenotypic variance typically has both genetic and environmental components. Heritability estimates, therefore, can change if the contributions of genetic factors or environmental factors change (Goldberger, 1979; Plomin et al., 2013; Visscher et al., 2008).
High heritability does not imply that a phenotype is not modifiable by an environmental intervention either (Goldberger, 1979). Consider the case of phenylketonuria (PKU), a disease determined by a series of mutations in the genome, resulting in a heritability of 1. Yet, with a modified diet, one can prevent PKU from leading to intellectual disabilities, seizures, and behavioral problems. PKU is a so-called developmentally contingent trait, which means that the individual might initially be genetically predisposed to develop a range of phenotypes and that the phenotype becomes fixed after exposure to a certain environment (Mealey, 2001).
Research in behavioral genetics indicates that the h2 for loneliness ranges from .37 to .55 (see Table 1; Boomsma, Cacioppo, Slagboom, & Posthuma, 2006; Boomsma, Willemsen, Dolan, Hawkley, & Cacioppo, 2005; Distel et al., 2010; Matthews et al., 2016; McGuire & Clifford, 2000; Waaktaar & Torgersen, 2012). This work also suggests that the heritability estimates of loneliness decrease somewhat with age (Bartels, Cacioppo, Hudziak, & Boomsma, 2008), with higher heritability estimates found in children compared with adults (Goossens et al., 2015).
Shared and nonshared environmental influences account for significant variation in loneliness during childhood (Bartels et al., 2008; McGuire & Clifford, 2000), whereas nonshared rather than shared environmental influences account for individual differences in loneliness in adulthood (Boomsma et al., 2005; Distel et al., 2010). Shared environmental influences in childhood include the home, food, resources, marital stability of parents, and so forth, whereas nonshared environmental influences in childhood include differential parental treatment, sibling interactions, nonsystematic factors (e.g., falling ill), and perceptions of the environment, through which events that are shared between individuals (e.g., divorce) can have a nonshared environmental influence (Plomin & Daniels, 2011). Children spend most of their time with family members when they are young but spend increasingly more time with peers and alone as they get older (e.g., Larson & Richards, 1991), eventually leaving their shared environment (e.g., the parental home) altogether. It can thus be expected that the nonshared environment becomes more important in explaining variations in loneliness in adulthood (Boomsma et al., 2005, 2006; Distel et al., 2010; Matthews et al., 2016; McGuire & Clifford, 2000; Waaktaar & Torgersen, 2012).
In summary, the findings from behavioral genetics, which are based on samples of related individuals, are in line with the predictions derived from the ETL. That is, in line with the idea of loneliness being a heritable, innate response, genetic factors account for a substantial portion of the variance in loneliness. Moreover, behavioral genetic studies have shown that the amount of variance explained by environmental factors, particularly nonshared environmental factors, increases during adulthood. Clarifying the nature of these influences remains an important task for a life-span theory of loneliness (see Qualter et al., 2015).
Molecular Genetics and Loneliness
The field of molecular genetics relates the genome to a phenotype or phenotypic variation in a population. The goal in molecular genetics is to specify the genes or gene sets (i.e., segments that code for the production of RNA molecules and proteins) that are responsible for the phenotype of interest. Every individual has two copies of each gene, called alleles, one of which is provided by each parent. The genes are composed of nucleotides, of which four varieties exist (i.e., adenine, cytosine, guanine, and thymine). Genetic variation between individuals can occur when there is a difference in one nucleotide, called a single-nucleotide polymorphisms (SNP), or in a sequence of nucleotides (Manuck & McCaffery, 2014; Visscher et al., 2017). There are two types of research within the field of molecular genetics: candidate-gene studies and genome-wide association studies (GWASs). Candidate-gene studies examine a specific part of the genome (e.g., a specific sequence of nucleotides) in relation to the phenotype of interest, whereas GWASs examine SNPs across the whole genome in relation to the phenotype of interest (Visscher et al., 2017). Although the goal is the same for both types of studies, the methods and findings can differ significantly. The GWAS approach is generally preferred over candidate-gene studies (see also Manuck & McCaffery, 2014; Munafò, 2006).
Candidate-gene studies
Several candidate-gene studies have examined the association between specific genetic variants and individual differences in loneliness. Most have examined the main effects of specific genetic variations on loneliness (see Table 1; Chou, Cacioppo, Kumari, & Song, 2014; Connelly et al., 2014; Lan et al., 2012; Lucht et al., 2009; Tsai et al., 2012; van Roekel, Goossens, Scholte, Engels, & Verhagen, 2011; van Roekel, Scholte, Verhagen, Goossens, & Engels, 2010; van Roekel, Verhagen, Engels, Goossens, & Scholte, 2013; van Roekel, Verhagen, Scholte, et al., 2013; Verhagen, van Roekel, & Engels, 2014). However, only a few studies have found a significant relation of a particular gene to loneliness (Chou et al., 2014; Connelly et al., 2014; Lan et al., 2012; Tsai et al., 2012; van Roekel, Verhagen, Scholte, et al., 2013; Verhagen et al., 2014), and direct replication attempts have yielded inconsistent results (Lucht et al., 2009; van Roekel, Verhagen, Engels, et al., 2013).
Candidate-gene studies were first introduced to identify risk variants for a particular disease (Kwon & Goate, 2000) and were later applied to psychopathology. However, two obstacles were encountered when the methods of candidate-gene studies were applied to identifying the genes underlying psychopathological phenomena, including loneliness. First, the relevance of individual genes or the biological effects of specific genes to a complex phenotype, such as loneliness, is not known. Second, the size of the association between any single SNP and a complex phenotype, such as loneliness, is expected to be small, perhaps very small in light of evidence from behavioral genetics suggesting that loneliness is polygenic. This means that candidate-gene studies have been statistically underpowered, which leads to an exaggeration of true effect sizes (i.e., effect inflation), a high rate of false positives (i.e., type I error) relative to true positives and a high rate of misses (i.e., type II error), and the unlikelihood that true as well as false effects will be replicated (Asendorpf et al., 2013; Button et al., 2013).
A higher-power replication attempt conducted within the GWAS framework is discussed in greater detail later in this review. None of the previously published associations between variants within candidate genes (i.e., brain-derived neurotrophic factor, or BDNF; oxytocin receptor; retinoic acid receptor-related orphan receptor α; metabotropic glutamate receptor 8; neuronal acetylcholine receptor subunit α4; interleukin 1α; corticotropin-releasing hormone receptor 1; methylase tetrahydrofolate reductase; dopamine receptor D2; and apolipoprotein E) and loneliness have been replicated, despite a much larger sample size (Gao et al., 2017). The inability of GWASs to replicate the significant effects of candidate-gene studies is not surprising in light of the statistical issues in underpowered designs, and the inability to replicate candidate-gene studies is a common finding in the psychopathology literature (Duncan & Keller, 2011; Munafò, 2006), for instance, in research on depression (Bosker et al., 2011). However, although GWASs have not replicated the significant genetic effects of candidate-gene studies, it does not necessarily mean that none of these genetic variations has an effect on loneliness. Whereas candidate-gene studies are plagued by false positives as a result of being underpowered, GWASs are likely to produce false negatives as a result of the conservative correction for multiple testing, especially if the effect size for an individual gene is small (see Ioannidis, Tarone, & McLaughlin, 2011).
A subset of candidate-gene studies has also examined whether the effect of the environment on loneliness depends on genetic variation (see Table 1; Chou et al., 2014; Connelly et al., 2014; van Roekel et al., 2011; van Roekel et al., 2010; van Roekel, Verhagen, Engels, et al., 2013; van Roekel, Verhagen, Scholte, et al., 2013). Several of these Gene × Environment (G × E) studies reported significant interactions between genetic variations and both positive and negative environments. The findings are in accordance with the differential susceptibility hypothesis in that carriers of particular genotypes experience more loneliness when encountering adverse environments than noncarriers (see Goossens et al., 2015). However, these findings should also be considered preliminary. Duncan and Keller (2011) have estimated, without correcting for publication bias, that only 27% of the replication studies on G × E interactions have been successful. Indeed, to our knowledge, only one study has attempted to replicate these G × E interaction effects, and the interactions were not replicated (Spithoven et al., 2015). To date, candidate-gene studies provide no convincing evidence for the involvement of a single gene in loneliness. These results, then, also favor the notion that multiple genes each make small, possibly very small, individual contributions to loneliness.
GWASs
GWASs are used to detect associations between common variations across the whole genome and the trait of interest (Visscher et al., 2017), such as loneliness. Unlike candidate-gene studies, GWASs do not rely on hypotheses regarding underlying mechanisms because the technique is statistically driven (Manuck & McCaffery, 2014; Wilkening, Chen, Bermejo, & Canzian, 2009). The large number of SNPs tested and the large sample sizes are more likely to detect reliable genetic effects than is the case for candidate-gene studies, even when the genetic effects are small and despite strict statistical corrections (Manuck & McCaffery, 2014). GWASs can also be used to estimate the heritability of a trait. Heritability is defined in GWASs in a narrow sense; that is, it is defined as the proportion of phenotypic variance due to the additive effects from common genetic variants (i.e., genetic variations that occur in > 5% of the population; Manolio et al., 2009; Visscher et al., 2017). This more restricted focus explains why estimates of heritability in GWASs are lower than they are in behavioral genetics studies, a phenomenon referred to as missing heritability. Missing heritability in GWASs may result from not taking into account the effects of rare genetic variations (i.e., genetic variations that occur in < 5% of the population), dominance (i.e., the effect of one allele is masked by the presence of the second allele), and/or epistasis (i.e., G × G interactions; Manolio et al., 2009).
Gao et al. (2017) published the first GWAS on loneliness using data from the population-based Health and Retirement Study (HRS; N = 10,760). They estimated the h2 for loneliness to range from .14 to .27. Although a significant amount of variation in loneliness was explained, no single SNP reached genome-wide significance. In line with the conclusion of our earlier review (Goossens et al., 2015), this finding suggests that the separate SNPs have a very small effect on loneliness. In addition to estimating the amount of variance explained by genetic variation, Gao et al. (2017) also examined the coheritability of loneliness with other psychopathology and personality characteristics. Loneliness showed a positive coheritability with neuroticism and a negative coheritability with extraversion (see also Table 1; Gao et al., 2017). No statistically significant association was found in the GWAS for the coheritability of loneliness and psychopathology (e.g., major depressive disorder, bipolar disorder, schizophrenia). Because loneliness and depression have repeatedly been found to have reciprocal influences (e.g., J. T. Cacioppo et al., 2010; J. T. Cacioppo, Hughes, Waite, Hawkley, & Thisted, 2006; Vanhalst, Luyckx, Raes, & Goossens, 2012; Vanhalst, Luyckx, Teppers, & Goossens, 2012), the question of whether loneliness and major depressive disorder were coheritable was especially important. The GWAS findings suggest that the shared variance between loneliness and depression is the result of phenotypical expression, not common genetics.
Abdellaoui et al. (2018) also performed a GWAS on loneliness using data from twins and family members from the Netherlands Twin Register (N = 4,222). Results replicated the findings that loneliness has significant chip heritability (h2 ~ .14), no single SNP reached genome-wide significance, and both neuroticism and extraversion were coheritable with loneliness. Additional analyses showed that the association between loneliness and neuroticism decreased only slightly (and remained statistically significant) when correcting for extraversion. The association between loneliness and extraversion that decreased substantially to less than 1% explained the variance when correcting for neuroticism. These results suggest that the coheritability between loneliness and neuroticism reflects a set of genes that contribute to both neuroticism and loneliness. By contrast, the apparent coheritability between loneliness and extraversion is spurious because of the correlation between extraversion and neuroticism. In other words, these findings suggest that the shared variance of loneliness and neuroticism is the result, at least partly, of genetic overlap. Abdellaoui et al. (2018) suggested that the coheritability between loneliness and neuroticism may be related to the fact that loneliness is associated with a stronger sensitivity to negative social stimuli specifically, whereas neuroticism is associated with a stronger sensitivity to negative stimuli in general. This interpretation is consistent with the ETL, which states that some people are more likely to feel lonely than others because they show a stronger tendency to experience negative emotions when they find themselves in a social situation that is less supportive. The coheritability with the broader trait of neuroticism probably represents biological processes underlying negative affect, but it is still unclear what these processes are.
The sample sizes of these two GWASs (Abdellaoui et al., 2018; Gao et al., 2017) were still rather modest. A recent GWAS (Day, Ong, & Perry, 2018) conducted on a large sample of the United Kingdom Biobank study (N = 445,024) significantly expanded on these studies. Many SNPs showed nominal significance in their associations with loneliness, and 15 of these SNPs reached genome-wide significance. This finding suggests that GWASs can effectively find strong signals of genetic association with large sample sizes. A genetic risk score based on these 15 SNPs showed a significant correlation with loneliness in the HRS sample (as used in Gao et al., 2017), which attests to the validity of the approach adopted. Most of the findings of the two previous GWASs regarding heritability and coheritability were confirmed. Loneliness showed significant chip heritability (h2 = .04), and neuroticism was coheritable with loneliness. Additional analyses revealed that the strongest signals for genetic associations with loneliness were found for genes related to emotional expression and behavior. The latter finding is again consistent with the ETL because differences in the expression of negative emotions, particularly in low support environment, are thought to underlie different levels of loneliness according to that theory.
Conclusion
The findings on molecular genetics, which are based on samples of nonrelated individuals, provide some support for several predictions derived from the ETL. Recent GWAS findings, in particular, have revealed that many genes contribute, to a small degree, to differences in loneliness. These studies have also shown that loneliness does not share this genetic basis with depression. However, the genetic basis for the personality trait of neuroticism did show significant overlap with the genetic background of loneliness. Despite the clear convergence across all GWASs published so far (Abdellaoui et al., 2018; Day et al., 2018; Gao et al., 2017), it is important to point out that more work is needed to replicate and expand these findings. Another important prediction derived from the ETL—that environmental factors moderate the genetic effects on loneliness—has yet to be substantiated in empirical research. It is very important for future studies to incorporate environmental factors, especially as some genetic effects might appear only in the presence of certain environmental exposures (Jorgensen et al., 2009; Manuck & McCaffery, 2014).
The fact that GWASs are generally preferred over candidate-gene studies does not imply that the hypothesis-driven approach of candidate-gene studies is not viable. The inductive or statistically driven approach of GWASs does not necessarily target functional SNPs of interest despite the a priori hypotheses one might have about relevant SNPs. Rather than testing hypotheses at the level of single SNPs for candidate-gene studies, a more interesting approach is to test hypotheses at the level of biological pathways and to include all of the SNPs of a given pathway in the analyses (Jorgensen et al., 2009). An advantage of such an approach is that the targeted selection of SNPs allows researchers to examine rare genetic variations that occur in < 5% of the population (Wilkening et al., 2009). However, it should be noted that it is also possible to include a priori hypotheses about SNPs of interest into GWASs (for more information, see Eskin, 2008). Another interesting avenue for future research would be to use polygenic risk scores (PRSs; Purcell et al., 2009) representing each individual’s genetic vulnerability to loneliness, as derived from GWAS data, and to examine how this genetic vulnerability factor interacts with environmental risk factors, such as poor relations with parents or peers, to affect loneliness.
Functional Genetics and Loneliness
Behavioral and molecular genetics examine the statistical relation between the genomic variation and a phenotype, but it remains unclear how or why these genomic variations would be related to a phenotype such as loneliness. That is, the genome does not directly produce a phenotype. Rather, information from the genome is synthesized, through a process called transcription, into a product (e.g., RNA or proteins) that, in turn, might contribute to the phenotype (Meaney, 2010). Several processes have been identified that influence the expression of genes and subsequent synthesis of their products. Some studies within the field of functional genetics have focused on the gene-expression process itself, more specifically, whether the production of specific gene end products is increased (i.e., upgraded) or decreased (i.e., downgraded) for the phenotype of interest. However, it is important to note that this gene expression might differ depending on the examined tissue, phenotype of interest, and genotype. Other studies within the field of functional genomics examine mechanisms that control gene expression, even though these mechanisms are not encoded within the genome itself.
Gene expression
Several studies have found evidence for a downgraded expression of genes involved in anti-inflammatory and antiviral responses and an upgraded expression of genes involved in proinflammatory responses in lonely individuals (see Table 1; Cole, Capitanio, et al., 2015; Cole, Hawkley, Arevalo, & Cacioppo, 2011; Cole et al., 2007; Cole, Levine, et al., 2015; Moieni et al., 2015). It has been suggested that this profile of differentially expressed inflammatory genes, which is called the conserved transcriptional response to adversity (CTRA; Cole, 2014), represents an adaptive response of the immune system to social adversity. That is, social adversity might put the individual at risk for different diseases than social prosperity, as social contact with others increases the risk for viral infections, whereas social isolation increases the risk for wounds and bacterial infections. This suggests that the proinflammatory response is more important for socially isolated individuals than the antiviral response, which is reflected by the pattern of downgraded and upgraded genes (Cole et al., 2011). However, it has been suggested that this profile influences behavior above and beyond eliciting sickness behaviors (see Felger & Lotrich, 2013). More specifically, inflammation elicits a response from the social group. That is, inflammation implies the presence of a disease that can be transmitted to others within the social group. To decrease the risk of disease transmission, individuals experiencing inflammation are socially isolated (Lopes, 2016). Not surprisingly, the relationship between loneliness and CTRA was reciprocal: Loneliness predicted an increased CTRA 1 year later, and CTRA predicted loneliness levels 1 year later (Cole, Capitanio, et al., 2015).
Only one study has examined genetic expression postmortem in an area of the brain, the nucleus accumbens, that is involved in processing social stimuli and rewards (Canli et al., 2017). In previous studies, loneliness has been related to weaker activation in this area in response to social stimuli (J. T. Cacioppo, Norris, Decety, Monteleone, & Nusbaum, 2009). In accordance with predictions of the ELT, within this brain area a large number of genes were differentially expressed between lonely and nonlonely individuals (Canli et al., 2017). These genes have been associated previously, among other things, with behavioral processes, such as emotional and anxiety behaviors, neurological diseases, and psychological disorders (including depression and mood disorders). The genes that were differentially expressed in lonely and nonlonely older individuals significantly overlapped with genes differentially expressed in Alzheimer’s disease (Canli et al., 2017). However, it should be noted that these individuals participated in a study on common chronic conditions of aging. In other words, these results concern older participants (average age of death 85 years), of which the lonely individuals were more likely to have Alzheimer’s disease. The finding that differentially expressed genes in loneliness and Alzheimer’s disease overlap should thus be interpreted with caution, and replication is needed.
Epigenetics
Epigenetics refers to “processes and mechanisms that affect the activity of the DNA but do not change the DNA itself” (Lester, Conradt, & Marsit, 2016, p. 29). Over the past decade, researchers’ interest in these processes and mechanisms has increased exponentially. One reason for this flurry of interest is that epigenetic processes, in contrast to the DNA sequence, which does not change over time, are dynamic and reversible (Meaney, 2010; Tsankova, Renthal, Kumar, & Nestler, 2007). Because epigenetic processes appear to be reversible, they are of interest for both medical and therapeutic treatment (Szyf, 2001). A common hypothesis is that all of the chemical processes of epigenetics reflect environmental influences on the functional genome or a form of gene-environment interplay. In other words, the effect of environmental factors on the phenotype is hypothesized to be mediated by epigenetic mechanisms. The chemical machinery of the genes is negatively affected by the dangers of an unfavorable environment or opens up to the opportunities of a favorable environment (Boyce & Kobor, 2015). It is important to note, however, that, like genetic expression, epigenetic mechanisms are tissue- and cell-specific. Moreover, various epigenetic processes influence and depend on one another (Rivera & Bennett, 2010). The best-understood epigenetic mechanisms, which are discussed in more detail in the paragraphs that follow, are DNA methylation and histone modification (Rivera & Bennett, 2010).
DNA methylation is a process by which a methyl group is added to a cytosine nucleotide (i.e., one of the four basic units in the DNA sequence; Jones, 2012; Tsankova et al., 2007). More specifically, a methyl group is added to a cytosine nucleotide (denoted as “C”) that lies adjacent to a guanine nucleotide (denoted as “G”) and is connected to it by a phosphate group (denoted as “p”). If such CpG units cluster together (i.e., occur in a high density) in a particular part of the genome, they are referred to as CpG islands. Such CpG islands are found in 70% of the promoter regions of genes (i.e., the part of the gene that initiates the transcription process through which DNA is copied into RNA) but are relatively uncommon in other parts of the genome (Boyce & Kobor, 2015). Attaching a methyl group to a CpG unit inhibits the binding of transcription factors. That is, the attached methyl group makes it harder for the transcription factors to recognize the DNA sequence of the promoter region. DNA methylation thereby reduces transcriptional efficiency (Klose & Bird, 2006; Tsankova et al., 2007). The larger the number of methylated CpG units or islands in transcription starting sites of a particular gene, the more the activity of the gene is dimmed.
Histone modifications refer to the remodeling of the way in which the DNA is packaged to fit into the nucleus of the cell. DNA is tightly wrapped around proteins known as histones. This complex of DNA and its associated histone proteins is referred to as chromatin. Histones have protruding tails, and the chromatin structure can be modified by adding chemical compounds to specific locations on these histone tails (Boyce & Kobor, 2015; Tsankova et al., 2007). If the chromatin structure changes, the way in which DNA is wrapped around the histones is changed, and gene expression is either positively or negatively affected (Lester et al., 2016).
Two of these histone modifications are histone acetylation and histone methylation (not to be confused with DNA methylation). In histone acetylation, an acetyl group is added to the histone tails, which leads to a relaxation of the chromatin structure (Tsankova et al., 2007). Because DNA is wrapped less tightly around the histone proteins, the binding of transcription factors is facilitated (Meaney, 2010). That is, the “open state” of the overall structure, as a result of the acetyl group on the histone tails, makes it easier to “read” and transcribe the DNA. In histone methylation, a methyl group is added to the histone tails. The role of histone methylation is more complex, as it can both activate and repress transcription (Rivera & Bennett, 2010). In the latter case, DNA is wrapped more tightly around the histone proteins, which hampers the binding of transcription factors. This “closed state” of the overall structure makes it more difficult to read and transcribe the DNA. The complex role of histone methylation, that it can both activate and repress transcription, can be explained through the complexity of the underlying chemical processes. The effect of methylation depends on the specific location on the histone tail to which the methyl group is added and on the degree of methylation, as one, two, or three methyl groups can be added (Greer & Shi, 2012; Tsankova et al., 2007).
Although no epigenetic studies with humans have been conducted for loneliness, some animal studies suggest that epigenetic processes might play a role in loneliness. For example, it has been found that social isolation resulted in enhanced global DNA methylation, histone methylation, and histone acetylation in the midbrain of adult mice (Siuda et al., 2014). Moreover, Li, Du, Shao, and Wang (2016) examined histone acetylation in adolescent rats in a specific gene, BDNF, that is important for neuronal plasticity and connectivity as well as the proliferation, differentiation, and survival of neuronal cells. Social isolation was related to increased histone acetylation of the BDNF gene in the medial prefrontal cortex but decreased histone acetylation of that same gene in the hippocampus (Li et al., 2016). Assuming that animal models can inform human models, one can expect that loneliness in humans is associated with epigenetic processes such as DNA methylation, histone methylation, and histone acetylation.
Studying epigenetic processes for loneliness in humans is a promising avenue for future research. That is, epigenetics are likely the mediating mechanisms for the interactions between genetics and the environment that are now commonly studied (Barciszewski, Gurda, Paluchowski, Hornowska, & Jasielska, 2016; McGowan & Szyf, 2010). From this line of reasoning, it follows that the most general question of interest is how the environmental factors that influence loneliness are biologically embedded. On the one hand, it has been suggested that epigenetic effects can be found in particular neural pathways depending on the disorder of interest (see McGowan & Szyf, 2010). The ETL would suggest that for loneliness these effects can predominantly be found in systems or brain areas involved in social information processing (e.g., J. T. Cacioppo, Cacioppo, Capitanio, et al., 2015; J. T. Cacioppo & Hawkley, 2009; S. Cacioppo et al., 2014). On the other hand, it has been suggested that some epigenetic mechanisms, such as gene-specific DNA methylation, underlie various forms of psychopathology through their effects on “intermediate levels of function such as activity of the Hypothalamic–Pituitary–Adrenal (HPA) axis that regulates the stress response” (see McGowan & Szyf, 2010, p. 70). It is important for future epigenetic studies to take various internalizing problems into account to disentangle the unique epigenetic mechanisms for loneliness from the shared epigenetic mechanisms.
Conclusion
Gene-expression studies seem to provide further evidence for the ETL hypothesis that loneliness is associated with health risks. More specifically, gene-expression studies may explain why loneliness is associated with both increased inflammation and elevated cortisol levels. Moreover, some studies have suggested that several epigenetic processes, including DNA methylation, histone acetylation, and histone methylation, do have a role in animal models of loneliness. These animal studies therefore suggest that examining epigenetic processes might provide an interesting avenue for future research, including studies on humans.
Discussion
A recent analysis of 70 prospective studies involving more than 3 million people over an average of 7 years shows that loneliness increases the odds of mortality by 26%, which is about the same as the effect size observed for people living with obesity (Holt-Lunstad et al., 2015). The additional fact that more than 1 in 3 people in industrialized nations are dealing at least occasionally with loneliness and 1 in 10 are living in the grips of nearly constant loneliness makes it imperative that we more fully understand the risk factors for and processes through which loneliness operates. Investigations of the evolutionary origins of and genetic contributions to loneliness, including G × E interactions and correlations, have much to contribute.
Several important insights from genetic research on loneliness are highlighted in this review. It has become increasingly clear that many individual genes or SNPs on their own contribute only to a small degree to differences in loneliness. Moreover, the genes contributing to loneliness seem to partially overlap with genes contributing to neuroticism, but not with genes contributing to depression. Genetic factors, which include epigenetic effects and G × G interactions in addition to the effects of single genes, contribute substantially to loneliness. However, this does not imply that genes are the only risk factor for loneliness. In addition, the genetic studies discussed in this review show that genes are unlikely to have a direct effect on loneliness. That is, environmental factors determine in a dynamic fashion how genes are expressed.
As the field of genetics advances rapidly (Dick, Riley, & Latendresse, 2011), there is much that can still be learned about the genetics of loneliness. Genetics research, and epigenetics in particular, could facilitate our understanding of the biological mechanisms underlying the various developmental trajectories of loneliness. Greater insight into how genes and the environment interact to contribute to loneliness might thereby be obtained. However, future genetic studies should pay attention to power issues. The statistical power of genetic studies is a function of, among other things, the way in which the phenotype is coded (e.g., case control or continuous), minor allele frequency, type I error rate, and whether genetic main effects, G × E interactions, or G × G interactions are examined (Dick et al., 2011). Because of the large samples that are often needed for genetic studies, researchers compromise the quality of their phenotypical and environmental measures. Extensive measures of phenotype and environment are needed (Dick et al., 2011).
It should be stressed again that genes are not deterministic and do not have a direct causal role in loneliness. That is, the environment greatly influences the way genes are expressed. Genes and the environment should be thought of as complementary factors contributing to loneliness rather than competing factors. Accounting for the role of genes in loneliness is important when trying to obtain a comprehensive understanding of loneliness. Just as the role of the environment should not be ignored in genetic studies, the role of genes should not be ignored in developmental studies (Dick et al., 2011). It follows from this plea for an integrative approach to the nature-nurture issue that the substantial contribution of genes to loneliness by no means suggests that only medical treatment of loneliness would be effective. A meta-analysis on psychological interventions designed to alleviate loneliness indicated that efforts designed to change individuals’ social appraisal or their maladaptive social cognitions were most effective (Masi, Chen, Hawkley, & Cacioppo, 2011). The fact that recent research in genetics has suggested that lonely people’s tendency to interpret social situations in an overly negative way is partially under genetic control opens up new possibilities to increase treatment effectiveness. As we gradually begin to understand which specific genes contribute to loneliness, research in genetics may suggest ways in which psychological treatments could be tailored to the specific needs of subtypes of people with different genetic backgrounds (Bakermans-Kranenburg & Van IJzendoorn, 2015).
Footnotes
Acknowledgements
J. T. Cacioppo passed away in March 2018. He was an incredibly inspiring person and a great researcher. This article is dedicated to his memory.
Action Editor
Marjorie Rhodes served as action editor and June Gruber served as interim editor-in-chief for this article.
Declaration of Conflicting Interests
The author(s) declared that there were no conflicts of interest with respect to the authorship or the publication of this article.