
Abstract
Several labels, such as neuroticism, negative emotionality, and dispositional negativity, indicate a broad dimension of psychopathology. However, largely separate, often disorder-specific research lines have developed that focus on different cognitive and affective characteristics that are associated with this dimension, such as perseverative cognition (worry, rumination), reduced autobiographical memory specificity, compromised fear learning, and enhanced somatic-symptom reporting. In this article, we present a theoretical perspective within a predictive-processing framework in which we trace these phenotypically different characteristics back to a common underlying “better-safe-than-sorry” processing strategy. This implies information processing that tends to be low in sensory-perceptual detail, which allows threat-related categorical priors to dominate conscious experience and for chronic uncertainty/surprise because of a stagnated error-reduction process. This common information-processing strategy has beneficial effects in the short term but important costs in the long term. From this perspective, we suggest that the phenomenally distinct cognitive and affective psychopathological characteristics mentioned above represent the same basic processing heuristic of the brain and are only different in relation to the particular type of information involved (e.g., in working memory, in autobiographical memory, in the external and internal world). Clinical implications of this view are discussed.
Keywords
One of the most challenging problems for clinicians and researchers of psychopathology is to capture the large variability in symptom profiles into a parsimonious, comprehensive, and clinically useful classification system. Categorical descriptive schemas, such as the American Psychological Association’s Diagnostic and Statistical Manual of Mental Disorders (DSM) and the World Health Organization’s International Statistical Classification of Diseases and Related Health Problems, have historically dominated this endeavor. However, categorical models of psychopathology and psychiatric disorders are increasingly criticized for several reasons, such as the use of consensus- rather than evidence-based categories, arbitrary thresholds to delineate a categorical diagnosis, and the large heterogeneity of symptoms and processes within categories and their limited reliability and substantial comorbidity (Kotov et al., 2017).
On the other hand, a quantitative nosology based on empirical data analysis is rapidly developing, showing that the observed variation in psychopathology can be parsimoniously modeled using a limited number of hierarchically arranged dimensions (Caspi et al., 2014; Kotov et al., 2017). For example, analyzing the correlational structure among 11 first-order dimensions of psychopathology, Lahey, Krueger, Rathouz, Waldman, and Zald (2017) suggested one psychobiological dimension that increases the risk for psychopathology and stated that “negative emotionality lies at the heart of the general factor of psychopathology” (p. 161). Investigations using a broader database suggested a large p-factor (Caspi et al., 2014; Caspi & Moffitt, 2018) as a dimension that indicates the liability for any mental disorder, as well as for comorbidity, persistence, and symptom severity with disordered thought at the extreme end. Conway et al. (2019) recently proposed a dimensional hierarchical taxonomy of psychopathology (HiTOP) to guide mental health research. Their HiTOP describes a general psychopathology factor at the top and large spectra at the level below it that reflect dimensions of individual differences as identified in personality research. These spectra in turn aggregate syndromes and disorders and—further down the hierarchy—more narrow signs, symptoms, and components of psychopathological functioning to allow different degrees of specificity when describing the pathological state of an individual.
In the current article, we focus on the internalization spectrum as described in the HiTOP structure, which in personality research corresponds to different traits such as neuroticism (Eysenck, 1947; Zinbarg et al., 2016), negative emotionality (Eisenberg et al., 2005), negative affectivity (Watson & Clark, 1984), and dispositional negativity (N/NE; Shackman et al., 2016a, 2016b). There are several good reasons to focus on this last trait. First, N/NE is associated with less favorable conditions and outcomes in almost every domain of life, including education, work, relational stability, and various indicators of physical health, resulting in substantial individual and societal burden and excessive costs (Cuijpers et al., 2010; Shackman et al., 2016b). Second, N/NE is also critically involved in causing and maintaining psychopathology: N/NE is not only associated with a large array of disorders such as anxiety, mood, and substance-abuse disorders; amount of comorbidity; and a less favorable prognosis—it is also a powerful predictor for the development of these disorders in longitudinal studies (Hur, Stockbridge, Fox, & Shackman, 2019; Shackman et al., 2016b). Third, recent meta-analytic evidence has shown that N/NE can be altered with different types of therapy with similar moderate effect sizes (e.g., Cohen’s d = 0.50–0.60 for cognitive-behavioral, supportive, or mixed therapies; Roberts, Luo, et al., 2017). This is interesting because targeting a single transdiagnostic dimension such as N/NE is more efficient than developing separate treatments for a panoply of comorbid categorical disorders. At the same time, it suggests that a clear understanding of the processes involved in this transdiagnostic trait is extremely important for progress in preventing its consequences and improving treatment effects.
The goal of the current article is to use a novel theoretical perspective on how the brain processes information to suggest a processing heuristic that is at the core of N/NE. We call this heuristic a “better-safe-than-sorry” (BSTS) processing strategy. The label “BSTS” is not new in this context: It has been used occasionally in previous research to indicate the rationale behind some N/NE-related mechanisms such as attentional and interpretational bias. However, it has not been understood as the central organizing principle at the core of N/NE, as is shown by the absence of this concept in several important conceptual articles on N/NE (Barlow, Ellard, Sauer-Zavala, Bullis & Carl, 2014; Barlow, Sauer-Zavala, Carl, Bullis, & Ellard, 2014; Hur et al., 2019; Shackman et al., 2016a, 2016b). By taking a predictive-processing perspective, we aim to show that BSTS causes a stagnated error-reduction process that—depending on the specific type of information involved—results in different phenomenal characteristics of N/NE that up to now have been investigated along largely separate research lines. By putting BTST at the core of N/NE and by demonstrating how separate phenomena can be understood as representing the same processing heuristic of the brain, we aim to contribute to a deeper and more parsimonious understanding of N/NE. We also aim to show how this way of understanding may improve treatment by suggesting which treatment components are critical and why. Before elaborating on this predictive-processing perspective and its potential implications in understanding N/NE, we briefly summarize the current views on this trait.
Conceptualizations of N/NE as a Dimension of Psychopathology
The nature of N/NE
Resulting from both genetic and environmental factors and their interactions, N/NE appears as a broad and stable disposition that leads one to appraise situations as threatening, to hold negative anticipations about oneself and the world, to experience negative mood states and emotions, and to show poor emotion regulation (Barlow, Ellard, et al., 2014; McCrae & Costa, 2003; Ormel et al., 2013; Ormel, Riese, & Rosmalen, 2012; Watson & Clark, 1984). N/NE is different from a negative affective state in response to environmental factors. In general, individuals with high trait N/NE show more affective instability; that is, they show more moment-to-moment affective fluctuations, even between feelings of different quality (Kuppens, Van Mechelen, Nezlek, Dossche, & Timmermans, 2007). Within a constructionist view on emotion (Barrett, 2017), N/NE should be seen as a valenced background state reflecting a rather long-term imbalance between the necessary and actual/anticipated resources to secure basic needs (e.g., growth, survival, reproduction) rather than as a categorical constructed emotion.
N/NE is associated with more vigilance toward potentially negative information; behavioral inhibition; a greater intolerance for uncertain, ambiguous, and uncontrollable situations; and a lower threshold for reacting with avoidance and escape responses to threat and negative information, including one’s own negative emotions (Barlow, Sauer-Zavala, et al., 2014; Gray & McNaughton, 1996). N/NE is also associated with an array of behavioral characteristics such as attentional (Shackman et al., 2016a, 2016b) and interpretational (Mathews, Ridgeway, Cook, & Yiend, 2007) bias; worrying, ruminating, and catastrophizing (Ehring & Watkins, 2008); enhanced symptom reporting (Van den Bergh, Witthöft, Petersen, & Brown, 2017); reduced autobiographical memory specificity (Walker, Yancu, & Skowronski, 2014); and compromised fear-learning mechanisms, such as poor safety learning, enhanced generalization of threat perception, and poor extinction learning (Gazendam, Kamphuis, & Kindt, 2013; Haaker et al., 2015). It is likely that several of these characteristics rely partly on deficits in executive function and cognitive control (Hur et al., 2019).
Both animal and human research have shown that N/NE is associated with altered neural structure and function (Depue, 2009; Hariri, 2009; Ormel et al., 2013). Recent comprehensive reviews (Hur et al., 2019; Shackman et al., 2016a, 2016b) document elevated responding to threat in several brain areas, such as the amygdala, hippocampus, insula, bed nucleus of the stria terminalis (BNST), midcingulate and orbitofrontal cortex, and periaqueductal gray. Consistent evidence further suggests that the amygdala is a coordinating brain area involved in both resting-state and threat-induced responses that are associated with elevated N/NE. On the one hand, the amygdala receives input from sensory (thalamic), contextual (hippocampal), and evaluative/regulatory (prefrontal/insular) structures that flows from the ventral areas of the amygdala to the central and dorsal/posterior parts and to the BNST. On the other hand, the latter structures (central, BNST) coordinate the behavioral (e.g., avoidance, inhibition), physiological (e.g., autonomic, neuroendocrine), and cognitive (e.g., hypervigilance, attentional bias) responses characterizing negative affective states.
Acute stressors alter amygdala functional connectivity and potentiate amygdala responses to threat (Cousijn et al., 2010; Hermans et al., 2017), and these effects are stronger in individuals with N/NE (Everaerd, Klumpers, van Wingen, Tendolkar, & Fernández, 2015), suggesting sensitization of the amygdala. Repeated acute and/or chronic stress may also lead to neural changes in other brain areas, including the hippocampus and prefrontal cortex (McEwen & Gianaros, 2011; McEwen, Nasca, & Gray, 2016). These changes might affect larger brain networks subserving executive function and cognitive control such as the frontoparietal (dorsolateral prefrontal cortex, intraparietal sulcus) and cingulo-opercular network (midcingulate cortex, anterior insula, frontal operculum; Q. Li et al., 2017). Emerging evidence suggests that frontoparietal areas such as the dorsolateral prefrontal cortex are less efficient in allocating resources to execute function tasks in individuals with N/NE (for a review, see Hur et al., 2019).
N/NE and psychopathology
A major scientific challenge remains, however, to parsimoniously conceptualize N/NE as a general transdiagnostic psychobiological dimension to solve the multifinality problem (how a transdiagnostic factor causes different disorders) and the problem of divergent trajectories (why different individuals with the same transdiagnostic factor develop different disorders; Nolen-Hoeksema & Watkins, 2011). Nolen-Hoeksema and Watkins (2011) suggested solving these problems by sorting a large number of separate processes into a coherent framework consisting of distal, proximal, and moderating (risk) factors. In another attempt at solving these problems, Barlow, Ellard, et al. (2014b) described a complex of vulnerability factors (triple-vulnerability theory) involving (a) a biological (heritable) vulnerability factor predisposing emotion-related brain structures to hyperexcitability, (b) a (learned) psychological vulnerability factor representing “a pervasive sense of unpredictability and uncontrollability in relation to life events and a perceived inability to cope with negative outcomes from such life events” (p. 484), and (c) a (learned) factor that determines why an individual becomes concerned about a particular type of threat and thus develops a particular type of disorder. The Research Domain Criteria project also tries to identify and integrate dimensional constructs at multiple levels of measurement (from genes to self-reports) in different domains of functioning (Cuthbert & Insel, 2013). For example, Lang and coworkers suggested a dimension of psychophysiologic responding in the negative-valence system that cuts across DSM-based clinical anxiety diagnoses representing diminishing defensive reactivity (or blunting of the defensive system) with decreasing focal fear and increasing negative affectivity, functional interference, and distress of the patients (Lang, McTeague, & Bradley, 2016; McTeague & Lang, 2012).
These approaches have created coherent conceptual schemas that elucidate how general threat sensitivity may lead to various psychopathological phenomena through the operation of an array of empirically established processes. However, in this article we suggest that taking a new and more radical functional perspective may deepen our understanding in a more parsimonious way. From an evolutionary perspective, N/NE as a trait can be seen as an adaptive response to repeated experiences with threat involving a recalibration of the threshold for threat detection and concomitant vigilance and hyperarousal. An optimal threshold for threat detection depends on two factors: the probability of an aversive event and the relative payoffs of the four possible outcomes of detection (i.e., true and false positives and negatives; Nettle & Bateson, 2012). Given the typically much larger costs of false negatives (missing threat) compared with false positives (metabolic expenditure), it is adaptive to recalibrate thresholds to lower levels when aversive experiences are likely, representing a BSTS strategy to cope with threat. Because of the autocorrelated nature of the person-environment system within individuals and across generations, it is not surprising that recalibration to lower thresholds has been observed in animals and humans, especially after repeated (early) adverse experiences, and that it can be transferred to subsequent generations (Baker, Cesa, Gatz, & Mellins, 1992; Hanson, Hariri, & Williamson, 2015; Hanson, Nacewicz, et al., 2015; Hariri & Holmes, 2006; Kendler, Prescott, Myers, & Neale, 2003; McEwen et al., 2016; Nettle & Bateson, 2012). Recalibration occurs in both animals and humans, suggesting a pervasive change in the way living animals construe and behave in the world that is not always captured by assessing cognitive operations and/or verbal expressions. In addition, it suggests that bias should not be considered irrational and dysfunctional but rather quite rational and adaptive in view of a person’s history (Gilbert, 1998; Nemeroff, 2013).
In line with this functional perspective, the generalized unsafety theory of stress (GUTS; Brosschot, Verkuil, & Thayer, 2016, 2017, 2018) suggests that the default state of organisms is to expect and prepare for threat and that only the perception of safety can inhibit this default state. Individuals with high trait N/NE would be unable to perceive safety, resulting in a chronic state of threat, even when none is present. Brosschot and colleagues consider different behavioral characteristics such as attentional bias, negative interpretation biases, perseverative thinking, poor safety learning, and chronic stress to represent a strategy of “erring on the safe side.” However, whereas GUTS describes why these phenomena should be interpreted this way, it does not explain how information is processed representing this strategy and how precisely this leads to the behavioral phenomena that can be observed in clinical practice.
Clarifying this “how” in terms of processing heuristics in the brain is the purpose of the current article. We suggest that interpreting N/NE and associated cognitive, affective, and behavioral phenomena within a predictive-processing perspective elucidates the centrality and communality of a BSTS strategy in a way that would not be apparent without these new conceptualizations. Predictive-processing accounts entail a radical constructivism in which the brain is seen as an organ that actively constructs a model of reality from noisy input using information it already has. Prediction signals from models in the brain are matched with sensory input, resulting in prediction errors that are fed back to improve the adaptivity of these models when making perceptual inferences and actively navigating the environment. In a broad sense, we suggest that individuals who are highly sensitive to threat tend to abort this error-reduction process in a premature phase. Although this strategy may be rewarded by beneficial effects in the short term because it helps individuals to quickly classify information about threat and negative valence, it ultimately leads to insufficient updating of their model of reality, resulting in persistent deviations between expected and actual input. We suggest that such a stagnated error-reduction process is the core of the psychobiological dimension of N/NE and that phenomenally different clinical characteristics that are associated with it are at a more fundamental level implementations of one and the same process. Before elaborating on this point, we briefly introduce predictive-processing models (for more elaborate introductions, see Barrett & Simmons, 2015; Clark, 2013; Friston, 2010, 2013; Friston, FitzGerald, Rigoli, Schwartenbeck, & Pezzulo, 2017; Hohwy, 2012, 2013; Seth, 2013; Stephan et al., 2016; Van den Bergh et al., 2017; Wiese & Metzinger, 2017).
A Predictive-Processing Model
Rooted in theories of inference and control in biological systems, new conceptualizations of the brain have emerged in computational neuroscience, the implications of which are currently being explored in a growing number of areas (Barrett, 2017; Stephan et al., 2016). Conceiving of the brain as “an ever-active hierarchical prediction machine” (Clark, 2015, p. 3) striving to minimize prediction error has important implications for a functional interpretation of perception, attention, emotion, thought, language, and action and more widely for our understanding of mind, experience, and agency, leading several authors to qualify this perspective as a paradigm shift (Barrett, 2017; Clark, 2013; Hohwy, 2013; Lupyan & Clark, 2015).
Predictive processing, perception, and action
According to the predictive-processing framework, a basic task of the brain is to construct an adaptive model of the (external and internal) world, although its only source of information to do so is the spatial and temporal patterning of its own neural activity. Models are considered adaptive when they allow one to quickly and efficiently infer the sources of stimulation (perception) as well as to predict future states and consequences of actions (planning and action). To achieve this goal, the brain uses information not only from neural activity that is triggered by peripheral input (sense organs and receptors in the peripheral body) but also from neural activity generated by the brain itself, reflecting previous experiences and “built-in” information that act as predictions. The theory of predictive coding or processing specifies how the brain makes sense of the stimulation it receives given certain expectations using principles akin to Bayesian inference.
Input from the (external or internal) world leads to two counterflowing streams of neural activation across several hierarchical levels of the brain: Stimulation by peripheral input (“likelihood”) interacts with activations generated by the brain (“generative model”) that act as probabilistic predictions of the input (“prior beliefs”) within a specific context—that is, they estimate a likelihood of what the new input will be given previous experience. It should be noted, however, that prior beliefs are implicit assumptions of the brain that in most cases are not accessible to consciousness. Thus, they can be quite different from conscious beliefs. The discrepancy between predicted and actual inputs across multiple hierarchical levels—from low-level sensory input to high-level abstractions—results in prediction errors that are propagated throughout the system in a process of error minimization. The brain will eventually settle on/infer a posterior model that represents the most likely model of the stimulation. For example, if one is waiting for Jeff in a crowded street, the brain generates neural patterns acting as prior beliefs that will facilitate spotting Jeff in the crowd (example from Pezzulo, Maisto, Barca, & Van den Bergh, 2019, p. 3).
The system realizes error minimization in three ways: by adapting the prior beliefs to accommodate the actual input (belief update), by actively operating on the world and generating input that fits the prior beliefs (“active inference”), and by changing how the brain samples (or attends to) sensory input (Barrett & Simmons, 2015). Active inference acknowledges that perception does not passively wait for sensory input but depends on action (e.g., active sampling) to produce input. For example, waiting for Jeff may prompt the person to move toward a location that provides a better overview of the passing crowd and/or increases the scanning rate, generating more detailed information to help spot him (Pezzulo et al., 2019). In a broader sense, however, it refers to any kind of perceptual, somatovisceral, and behavioral (goal-directed) responses to produce input that is consistent with the expectations specified in the generative model to minimize prediction error. Active inference therefore involves “policy selection” to reach epistemic and/or pragmatic goals (Friston, FitzGerald, Rigoli, Schwartenbeck, & Pezzulo, 2016). This process of error minimization eventually settles on posterior beliefs that best account for the prediction errors and that reflect the combined influence of priors and the actual input. Conscious experience is thought to correspond to the posterior beliefs that are the most likely explanation of what is happening in the world (Hohwy, 2012).
An interesting consequence of this perspective is that it integrates perception, action, and physiological regulation within the same theoretical account (for a hierarchical predictive coding model describing how multiple neurovisceral interactions regulate heart rate variability in interactions with physiological and psychological demands, see R. Smith, Thayer, Khalsa, & Lane, 2017).
Precision and precision control
Priors, prediction errors, and posterior beliefs are conceived of as probability distributions that represent statistical regularities in neural activity with a mean and a variance. The inverse of the variance of these distributions is its precision. Highly precise priors and prediction errors reflect a neural pattern that has a high probability of being associated with a particular input. The opposite is true for low precise priors and prediction errors. For example, if Jeff is unusually tall, both priors and prediction errors representing Jeff’s height are highly precise, resulting in a quick and reliable recognition of Jeff. The relative impact of the distributions representing prior beliefs (i.e., predictions) versus the distributions representing the input (i.e., the likelihood of what is present) on the posterior beliefs (i.e., what the system concludes is present) will be determined by the precision of the distributions. For example, when it is dark, there is a high probability of recognizing Jeff in any tall person, reflecting a strong effect of the prior on the eventual perception. Conversely, on a sunny day it is less likely to take any tall person for Jeff, and this likelihood is even further reduced if one is not waiting for Jeff.
Because the brain cannot know whether any residual prediction error represents random information or is amenable to further minimization, it must learn the conditions under which particular models are likely to be adaptive. This is accomplished by developing context-dependent expectations about the precision of its inputs, which in turn determine how much weight is given to the prediction errors in the process (“precision optimization”; Hohwy, 2012). The implication is that contextual cues may have an important impact on the eventual posterior model that corresponds to conscious experience. In the case of Jeff, the perceptual information related to Jeff’s height is highly precise, and the brain will learn to consider height to be a highly precise prior for recognizing Jeff. In a more general way, precision control is extremely important in creating adaptive models of the stimulation that reaches the brain (Parr & Friston, 2018). Figure 1 illustrates how the relative precision of priors and prediction errors (likelihood) affects the eventual posterior model.
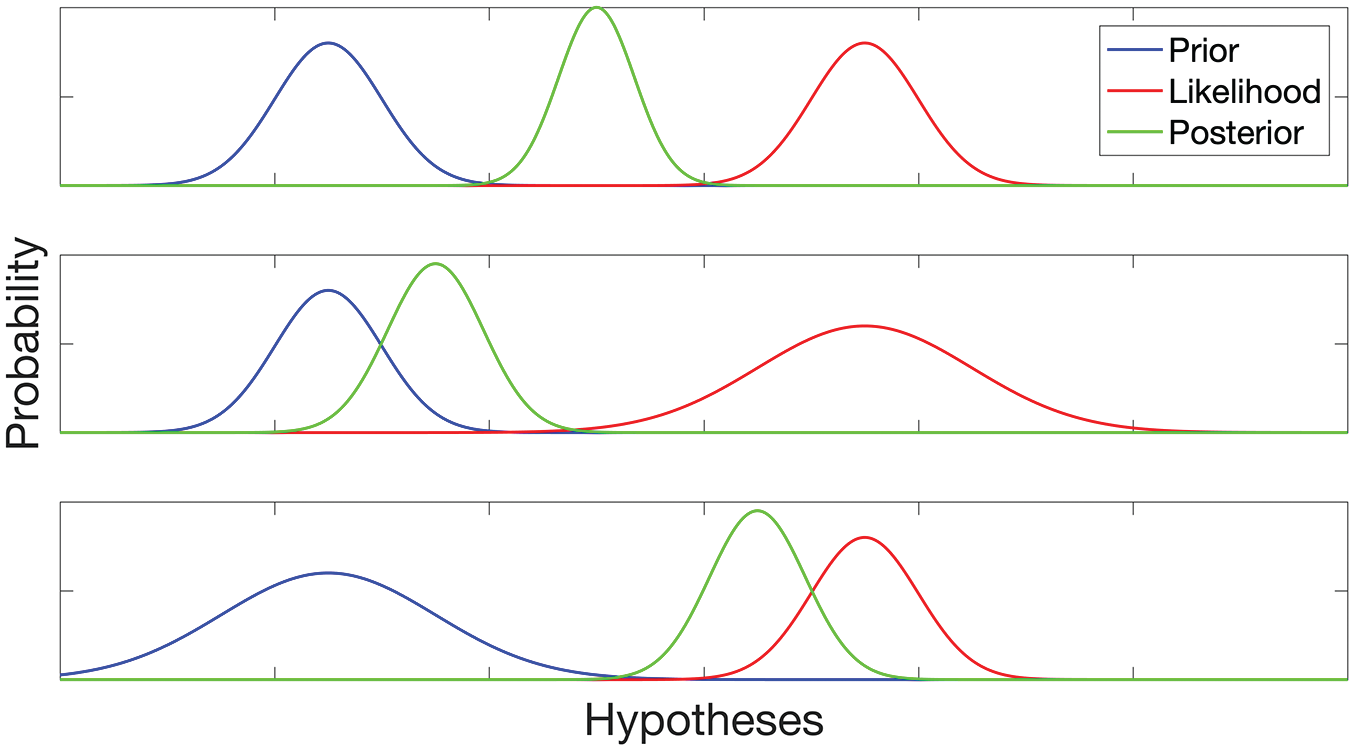
Integration of prior knowledge and sensory evidence (likelihood) in Bayesian inference. The top panel shows that if the prior and likelihood have the same precision (i.e., inverse variance of the Gaussian distribution), the posterior belief is in between. The middle and bottom panels show that a higher-precision prior and likelihood “attract” the posterior, respectively. Note that in all cases the precision of the posterior increases compared with the prior. Reproduced from Pezzulo, Maisto, Barca, & Van den Bergh (2019), which was published under a Creative Commons Attribution (CC BY) 4.0 International License.
Major advances have been made in relating these predictive-processing concepts to a neuroanatomical architecture and in operationalizing them in formal computational ways (see Friston, Rosch, Parr, Price, & Bowman, 2018; Parr & Friston, 2018; J. A. Roberts, Friston, & Breakspear, 2017).
Some implications
Several implications follow from this way of understanding the functionality of the brain. First, conscious experience always reflects prior expectations to some extent, but the degree to which this happens can vary. Prior beliefs with high precision in the context of imprecise inputs are likely to have a strong impact on the posterior beliefs, whereas the reverse is true when high-precision inputs are processed while prior beliefs are imprecise. In both cases, however, the same compelling sense of being “real” or “true” is produced. Second, the criterion for the brain to settle on particular posterior beliefs is not accuracy but adaptiveness or usefulness. Although accurate beliefs are mostly adaptive, it can be more adaptive to be biased (Lynn & Barrett, 2014). This suggests that it might be more fruitful not to consider bias a mistake or “being wrong” that should be corrected. Instead, trying to understand its usefulness and modifying conditions to make it less adaptive might be a more fruitful approach for modifying the information-processing “heuristics.” Third, and most importantly, predictive processing does not assume a self or an agent that organizes the information-processing traffic. Instead, the brain is considered to self-organize following the principle of free-energy minimization, and the sense of self or agency is a product of processing heuristics themselves producing perceiver and percept at the same time (Friston, 2010; Hume, 1739/2007). It follows that N/NE should not be considered to be something that a self “has” or to be some force that affects and skews “normal” information processing and should be controlled or regulated by a superseding self. Rather, we suggest that N/NE should be considered inherent to the data-processing heuristics themselves (Petersen, von Leupoldt, & Van den Bergh, 2015). The implication is that different characteristics of N/NE should not be seen as separate phenomena that are associated with N/NE but as a representation of these very data-processing heuristics. In this way, we suggest a more parsimonious interpretation of N/NE and its associated phenomena with potentially important clinical implications. We expand on this perspective below.
N/NE in a Predictive-Processing Model
If N/NE is considered an adaptive trait that involves recalibrating the threshold for threat detection representing a BSTS strategy, the question then becomes how a BSTS strategy is conceptualized in a predictive-processing model. Because input to the brain as represented in neural distributions can in principle be categorized in an infinite number of patterns, the system must apply a decision rule to settle on a categorical posterior model (to solve the “infinity problem”; Chater & Vitányi, 2003). The decision rule should ideally be set in such a way that not only is important information not missed but also details are not too manifold and/or too specific to impede generalizing and learning from prior experience (Petersen et al., 2015; Shepard, 1987; Rigoli, Pezzulo, Dolan, & Friston, 2017). Threat processing by individuals with high trait N/NE can be seen as involving a decision rule that has shifted toward oversimplifying input. The benefit may be greater speed in categorizing input as threat at the expense of the level of detail by which the input is processed.
Applying a decision rule that reduces detailed processing of the prediction errors while maintaining highly precise threat-related priors results in perceptions that are more informed by categorical threat-related priors than by actual input. This process (a form of “jumping to conclusions”) has the advantage of reducing uncertainty in the short term, but the cost is that prediction errors tend to remain imprecise, reducing the evidence needed to update the prior beliefs and enabling high-level (threat-related) priors to further dominate one’s conscious experiences. Reduced detail in processing information from the inner and outer world and poor updating of prior beliefs will also result in generative models with a low level of detail that, according to computational simulations, increases the precision of predictions (Kwisthout, Bekkering, & Van Rooij, 2017), leading to chronic conditions of uncertainty/surprise in the long term (i.e., new unpredicted input will always have to be dealt with). This suggests that a stagnated error-reduction process is at the core of high trait N/NE 1 and involves persistent deviations between model-based prior expectations and actual evidence, which is “the hallmark of a bad model” (Stephan et al., 2016, p. 6).
It is important to note that from a predictive-coding perspective N/NE is not only an affective quality added to the error-processing dynamics. Rather, emotional valence is considered to emerge from these dynamics at work (Joffily & Coricelli, 2013; Van de Cruys, 2017). 2 For example, unresolved mismatch (“surprise”) 3 between predicted and actual stimulation may characterize feelings of curiosity and interest and generate positive feelings of mastery as long as predictive progress is being made (Van de Cruys, 2017). However, when predictive progress stagnates, the persistent deviations between model-based prior expectations and evidence may engender unproductive coping behaviors (e.g., chronically elevated vigilance) and inappropriate physiological activations that characterize N/NE, which in turn may further compromise updating of nonadaptive prior beliefs (Stephan et al., 2016).
It is currently not clear how this processing strategy is implemented in the brain and maps onto amygdala overactivation and deficient cortical control that characterizes N/NE as described above. In any case, a predictive-processing account goes beyond a focus on particular brain structures and considers the process of descending predictions, ascending prediction errors, and error minimization across hierarchical layers of the brain to involved information flow through large-scale functional networks (see, e.g., Barrett, 2017; Barrett & Simmons, 2015; Park & Friston, 2013). How functionality emerges from the structural architecture of the brain still remains to a large extent “a mystery in neuroscience” (Park & Friston, 2013, p. 1). Studies on how N/NE is implemented in large-scale functional neural networks are emerging, but the picture up to now has been fragmented because the focus of these studies has been on (a) specific aspects of N/NE (e.g., working memory in N/NE individuals) and (b) the different brain-connectivity analysis methods and metrics used. The results are thus rarely consistent (see Dima, Friston, Stephan, & Frangou, 2015; Gentili et al., 2017; B. J. Li et al., 2018; Ueda et al., 2018).
In sum, we suggest that high trait N/NE as a general vulnerability factor for psychopathology has a BSTS strategy at its core that implies a stagnated error-reduction process when processing input. Input is processed with low detail, resulting in low-precision prediction errors and allowing prior threat-related beliefs to dominate the immediate experience. However, in the long term it also leads to poor updating of prior beliefs, the maintenance of highly precise prior beliefs, and thus chronic uncertainty/surprise. We suggest that this pervasive way of processing input underlies a wide variety of disparate phenomena and that the phenomenal differences result from the different content of information rather than involving different processes. This is not to say that the phenomena we describe below must always show up together within one individual. Context and individual concerns and experiences can moderate which of the phenomena below will be more pronounced in a particular case (Barlow, Ellard, et al., 2014; Lahey et al., 2017; Nolen-Hoeksema & Watkins, 2011).
Exemplars of a General BSTS Processing Strategy
Several cognitive biases related to N/NE have been extensively documented and occasionally interpreted as examples of a BSTS strategy (Cisler & Koster, 2010; Forbes, Purkis, & Lipp, 2011; Mathews & MacLeod, 2005; Williams, Mathews, & MacLeod, 1996). Typical examples include attentional bias toward threat detection and negative emotional information and greater difficulty in disengaging from these biases. In addition, bias toward negatively interpreting meanings and drawing negatively valenced inferences has been well documented (Heinrichs & Hofmann, 2001; Mathews et al., 2007). Negatively biased recall of information is also a robust finding, particularly when the information relates to the self (Rusting, 1998). Because it is rather obvious that attentional and interpretational biases are examples of lowering the threshold for threat detection, these cognitive biases will not be reviewed here, although they have not yet been conceptualized within a predictive-processing framework. In the following sections, we address other N/NE-related psychological characteristics that are less obvious examples of a BSTS strategy and discuss how a predictive-processing perspective reveals the BSTS strategy at its core.
Perseverative cognition
Perseverative cognition, or the repetitive, sustained activation of cognitive representations of past stressful events or feared events in the future, refers to a class of cognitive activities subsuming depressive rumination and anxious worry that is both prospectively and cross-sectionally associated with anxiety disorders and depression (Brosschot, Gerin, & Thayer, 2006; Drost, Van der Does, van Hemert, Penninx, & Spinhoven, 2014). It can therefore be considered a transdiagnostic feature of internalizing disorders. Although perseverative cognition does not directly cause depression and anxiety, it acts as a mediator between N/NE and these disorders, enhancing their probability. Perseverative cognition is also associated with increased allostatic load on the cardiovascular (blood pressure, heart rate, heart rate variability) and endocrine (e.g., cortisol) systems (Ottaviani et al., 2016). Perseverative cognition is part of a broader class of repetitive thinking that includes “thinking attentively, repetitively or frequently about one’s self and one’s world” (Watkins, 2008, p. 163), but the maladaptive variant typically has a negatively valent content.
Perseverative cognition is apparently triggered by negative affective states, more precisely when such states are elicited by the awareness of a difference between the current state and a target state (J. M. Smith & Alloy, 2009). For example, depressive rumination can be seen as a cognitive elaboration of one’s current sad state and its potential negative consequences in attaining a more desired state, as well as an elaboration of the reasons for the discrepancy (“why” questions).
Perseverative cognition is characterized by an abstract level of construal (Watkins, 2008) that is described as forming general, superordinate, and decontextualized mental representations that convey the “essential gist and meaning” of events and actions, whereas concrete low-level construals include contextual, specific, and incidental details of events and actions (Watkins, 2008, p. 187). It is assumed that abstract construal is selected as an emotion-focused coping style because it allows cognitive avoidance of a thorough experience of negative affective states. This results in a short-term benefit (avoiding intense negative affect) but also a long-term problem because more adaptive processing of negative affect and active problem solving is impeded (J. M. Smith & Alloy, 2009). The overall effect is prolonged stress and negative affect, which promotes a vicious cycle with perseverative cognition as the motor that keeps it going.
The idea that perseverative cognition is a recurrent attempt to reduce discrepancies between actual and desired goals is central to control theory (Martin & Tesser, 1989, 1996) and in an extensive elaboration of it by Watkins (2008). Because both control theory and predictive processing are rooted in theories of inference and control in biological systems, it is not surprising that there are striking similarities with the current predictive-processing account of N/NE: The option to select a short-term benefit by avoiding more elaborate processing of negative content at the expense of a larger problem in the long term represents a BSTS strategy that is characterized by a stagnated error-reduction process. Rather than processing specific and contextualized events and actions that are rich in specific and concrete evidence (prediction errors) and to accommodate higher-order mental representations accordingly (error minimization), abstract construals that largely reflect prior assumptions about the individual in interaction with the world dominate mental activity, compromising adaptiveness of the mental models of oneself and the world in the long term and leading to chronic uncertainty. As noted by Watkins (2008, p. 192), this strategy is particularly disadvantageous in conditions of novelty, unfamiliarity, difficulty, or stress, that is, conditions that are typically threatening for individuals with high trait N/NE.
Although both ways of interpreting perseverative cognition are quite similar, the benefit of understanding perseverative cognition within a predictive-coding perspective is that it reveals identical mechanisms that underlie both perseverative cognition and N/NE. This suggests that perseverative cognition is not just a separate phenomenon that is associated with N/NE. Rather, perseverative cognition is at the cognitive level a very instance of the same processing style that constitutes N/NE. Put differently, it is N/NE at work when dealing with discrepancies between actual and desired goals. In this way, a predictive-processing account is more parsimonious.
Reduced autobiographical memory specificity
Reduced autobiographical memory specificity is typically assessed with the autobiographical memory test (Raes, Hermans, Williams, & Eelen, 2007; Williams & Broadbent, 1986), in which individuals are given an emotional cue word and are asked to recall specific autobiographical memories in relation to that cue word. A specific memory refers to a particular event that lasted less than a day, whereas an overgeneral memory is more generic and typically includes a class of events (categoric memories) or an extended period of time (extended memories; Griffith et al., 2009). The difficulty in retrieving specific personal memories of a past event is found in a broad range of psychopathological disorders, most notably in depression and posttraumatic stress disorder but also in acute stress disorder and somatic symptom disorder (Barry, Chiu, Raes, Ricarte, & Lau, 2018; Walentynowicz, Raes, Van Diest, & Van den Bergh, 2017). Reduced autobiographical memory specificity is also a marker of an unfavorable course of psychopathology that affects the severity of symptoms, illness duration, and treatment success (Sumner, Griffith, & Mineka, 2010); in a community sample, it predicted a gradual increase in depressive symptoms over the course of 18 months (Van Daele, Griffith, Van den Bergh, & Hermans, 2014).
Reduced autobiographical memory specificity is associated with impaired executive function (e.g., deficits in inhibitory control, updating and maintaining information in working memory, verbal fluency) and with avoidance of negative affect (Barry et al., 2018; Sumner et al., 2014). Several areas of the brain involved in processing emotional salience and self-relevance as well as in executive control, emotion regulation, and memory have been associated with reduced autobiographical memory specificity, and these effects may be due in part to abnormalities, chronic or otherwise, in cortisol. The activation patterns in brain areas are not systematically replicated across diagnostic groups with the same behavioral effects, suggesting that reduced autobiographical memory specificity may result from a number of different neurocognitive mechanisms (Barry et al., 2018). A large-scale neural-network analysis might extend and further clarify how reduced autobiographical memory specificity is implemented in the brain.
One of the important factors that is assumed to account for reduced autobiographical memory specificity is capture and rumination (CaR-FA-X model; Williams et al., 2007), which implies that memory retrieval is “captured” at a general level, thereby blocking the retrieval of specific memories, whereas ruminating keeps processing of the information at an analytical and abstract level (see above). These deficits obviously make adaptive processing of negative information less likely, leaving one stuck at the level of overgeneral, self-related information. The choice of this level of processing is assumed to result from functional avoidance of episodes of negative affect that might be triggered by emotional cues. In the long term, however, this strategy is maladaptive because it inhibits emotional processing and active problem solving related to the sources of the negative affect. An unresolved issue is whether this strategy is already in play at the encoding phase, resulting in poor specific and detailed memories of negatively valent or threatening information in the first place (Raes, 2005).
From a predictive-processing perspective, this way of processing information in memory is consistent with the BSTS strategy described above: Little detailed processing of memory input from autobiographical events results in low-precision prediction errors, allowing strong prior beliefs representing (negatively valent) generalized memory information to determine the conscious recall of self-referential events. Consequently, error-minimization stagnates at a rather abstract level, leaving generalized prior beliefs unchanged and contributing to a vicious cycle of negative affect and low autobiographical memory specificity. Again, it is clear that reduced autobiographical memory specificity, when viewed in this way, is not just a phenomenon that is associated with N/NE. It is actually the core of N/NE at work when processing self-relevant memory information.
Compromised fear learning
Associative fear learning typically involves repeated pairing of a (relatively) neutral stimulus with an aversive fear-inducing stimulus (unconditioned stimulus), after which the previously neutral but now conditioned stimulus (CS) elicits fear by itself. Extinction learning implies repeated unreinforced exposures to the CS, leading to reduced responding to it. Associative fear learning is widely used as a laboratory model for understanding pathological fear (Craske, Hermans, & Vervliet, 2018). Although strong conclusions about the effect of individual-difference variables are hampered by the “noise” induced by different methodologies, some evidence suggests specificities in associative fear learning that are associated with N/NE and the vulnerability to develop and/or suffer from actual pathological anxiety (Lonsdorf & Merz, 2017). First, anxiety-prone individuals tend to show poor extinction learning, that is, a less steep decline of fear responses during extinction and/or a higher level of remaining fear after extinction (Shechner, Hong, Britton, Pine, & Fox, 2014). Put differently, for an equal amount of expectancy violations, the CS remains categorized as potentially dangerous for a longer period of time. Poor extinction learning in the laboratory has been shown to predict the development of anxiety symptoms in real-life situations (Guthrie & Bryant, 2006).
Second, anxiety-prone individuals show impaired safety learning in a differential-learning paradigm that involves the presentation of two neutral stimuli, one of which is paired with the aversive stimulus (CS+), and the other is never paired with it (CS−), turning the neutral stimuli into, respectively, a danger cue and safety cue. Impaired safety learning, emerging as elevated fear responding to the CS−, has frequently been observed in individuals scoring high on trait anxiety (Gazendam et al., 2013; Kindt & Soeter, 2014), in individuals with subclinical levels of anxiety (Arnaudova et al., 2013; Chan & Lovibond, 1996; Haddad, Pritchett, Lissek, & Lau, 2012), and in individuals with anxiety disorders (Grillon & Morgan, 1999; Jovanovic et al., 2013; Lissek et al., 2009; Winslow, Noble, & Davis, 2008) and depression (Pollak et al., 2008). Other data have shown that patients with panic disorder had higher danger expectancy when safety stimuli were presented (Lenaert, Boddez, Vervliet, Schruers, & Hermans, 2015). Poor danger-safety discrimination also predicts the return of fear after treatment (Staples-Bradley, Treanor, & Craske, 2018). In other words, individuals who are vulnerable to or with affective psychopathology show reduced discrimination between danger and safety in a danger context, illustrating a bias toward categorizing a safe cue as dangerous (see also Garcia & Zoellner, 2017).
Third, overgeneralization of fear has also been observed in threat-sensitive individuals. Generalization occurs when fear has been learned in response to one stimulus and subsequently emerges in response to stimuli that are similar but have never been paired with the aversive stimulus. Overgeneralization, then, occurs when fear responding to generalization stimuli remains relatively higher than healthy controls across a decreasing gradient of similarity with the original CS. This suggests that threat-sensitive individuals are less sensitive to differences between a new stimulus and danger cues, leading to a higher probability of categorizing new stimuli as dangerous. Overgeneralization of learned fear has been found in several groups with anxiety disorders (Lissek et al., 2010, 2014; Lissek & Grillon, 2012; Lenaert et al., 2014).
Fourth, a general sense of unpredictability and uncontrollability contributes to the development of neuroticism. This may result from a lack of safety learning and/or repeated experiences with unpredictable and uncontrollable aversive life events and from harsh, intrusive, and overcontrolling parenting styles. It is associated with alterations in neural function and neuroendocrine regulation of the hypothalamic-pituitary-adrenal axis (see Barlow, Ellard, et al., 2014).
In sum, individuals prone to developing emotional disorders are less likely to develop inhibitory fear learning, as indicated by poor extinction, poor safety learning, and overgeneralization, and they experience the world as less predictable and controllable. Put simply, they are more likely to categorize a safe stimulus as potentially dangerous, need more expectancy violations to shift a danger cue into a category of safe ones, and—more so than less threat-sensitive individuals—tend to categorize as dangerous both a safe cue within a dangerous context as well as a new cue that resembles a dangerous one.
How do these effects come about according to a predictive-processing framework? As suggested above, these effects may result from applying a perceptual decision rule that has shifted toward oversimplifying input (impoverished sampling), allowing highly precise threat-related priors to have a stronger impact on the eventual categorical danger perception. The obvious benefit is a higher likelihood to categorize input as threat at the expense of the level of detail by which the input is processed. This can be considered an instance of ecologically situated perception (Linson & Friston, 2019). Evidence consistent with this interpretation has been found in relation to fear overgeneralization in individuals with anxiety. Rather than resulting from postperceptual-choice behavior, it reflects errors resulting from an altered perception of simple stimulus features and causes less discrimination between danger and safety cues (Laufer, Israeli, & Paz, 2016; see also Struyf, Zaman, Vervliet, & Van Diest, 2015; Zaman, Ceulemans, Hermans, & Beckers, 2019). Other evidence in support of this interpretation may be found in studies linking these learning abnormalities to memory specificity (see above). Accurate discrimination learning relies on sufficiently detailed and specific memory: Without detailed memory, experiences will not be represented in memory as unique, specific events, reducing the ability to differentiate danger from safety. Lenaert et al. (2015) showed that individuals with anxiety who had low memory specificity also showed poor discrimination learning in a differential-conditioning paradigm. It may also be relevant in this context that animal studies have shown that stress-related changes in the hippocampal area lead to deficient “pattern separation” during memory encoding, making distinct patterns of information (e.g., CS+ vs. CS−) less distinguishable in memory (Tronel et al., 2012).
The overall picture of these learning abnormalities suggests that individuals with high trait N/NE use an abstract, general way of construing sensory input, leaving conscious experience and behavior relatively more influenced by categorical threat-related priors than by actual sensory evidence. Again, this is compatible with a BSTS strategy when processing aversive information: Stimuli are more likely classified as aversive or dangerous at the costs of more errors “on the safe side.” This interpretation again suggests that N/NE-related learning abnormalities are not just phenomena associated with N/NE. They actually are the N/NE-related processing heuristics in action during fear learning that lead to altered phenomenal learning characteristics.
Symptom perception in individuals with somatization and medically unexplained symptoms
A pervasive positive relationship has been established between trait N/NE and symptom reports unrelated to physiological dysfunction, or so-called medically unexplained symptoms (MUS; Van den Bergh et al., 2017). Such symptoms are highly prevalent throughout the health-care system (Haller, Cramer, Lauche, & Dobos, 2015): They appear in individuals who are not consulting health care providers, such as those with high trait N/NE, who tend to have more symptoms in daily life; in primary care, in which patients with MUS are characterized by elevated levels of anxiety and depression; and in secondary care, in which functional somatic disorders (or somatic symptom disorder) are associated with elevated psychiatric comorbidity (Henningsen, Zipfel, & Herzog, 2007; Wessely, Nimnuan, & Sharpe, 1999; Witthöft & Hiller, 2010).
A clue to understanding MUS may be found in the fact that neurobiological and psychometric evidence (Walentynowicz, Witthöft, Raes, Van Diest, & Van den Bergh, 2018) suggest that the experience of a symptom entails a sensory-perceptual component and an affective-motivational component that are intuitively integrated into one symptom report. Individuals with high trait N/NE show enhanced affective-motivational responding to aversive somatic sensations as well as reduced detail in sensory-perceptual processing, blurring a clear distinction between an emotional and a somatic state (Van den Bergh & Walentynowicz, 2016). This results in little precise somatic input (prediction errors), allowing for a large impact of symptom-related categorical priors that are constantly activated in these individuals. In other words, in individuals with MUS, categorical perception of the somatic state is largely determined by somatic priors and less or not at all by distinct sensory input (Van den Bergh et al., 2017).
Less detailed sensory-perceptual processing of somatic episodes is suggested by a number of recent findings:
First, untreated individuals with high rates of MUS and patients with somatoform disorders show diminished correspondence between induced physiological dysfunction and self-reported symptoms, particularly when in a negative affective context (Bogaerts et al., 2005; Van den Bergh et al., 2004).
Second, patients with somatoform disorders do not exhibit a peak-end memory bias after an induced aversive somatic episode (Walentynowicz et al., 2018). The peak-end bias implies that retrospective memory of an event is typically determined by the experience at the peak and at the end. Its absence in retrospective symptom reporting in habitual symptom reporters and in patients with somatic symptom disorder indicates that their memory of a symptom episode is minimally affected by the sensory-perceptual changes during a symptom episode.
Third, when given health-related cue words for autobiographical memory retrieval, patients with somatic symptom disorder exhibit reduced autobiographical memory specificity after controlling for depression and rumination (Walentynowicz et al., 2017). This finding suggests that these patients process and encode health-related episodes in memory with minimal detail.
Fourth, more in-depth analyses of how anxious individuals with high rates of habitual symptoms process and categorize ambiguous interoceptive stimuli shows that they more often misclassify interoceptive stimuli of low intensities into a high-intensity category (Petersen et al., 2015).
Fifth, several impairments in learning to fear pain have been observed in patients with somatic symptom disorder that are in line with the examples mentioned above (see Compromised Fear Learning), such as slower acquisition of pain-related fear to specific cues, more learning of the fear of pain in response to the context, more nondifferential fear generalization, and impaired extinction of generalized fear (Meulders, Boddez, Blanco, Van Den Houte, & Vlaeyen, 2018; Meulders, Jans, & Vlaeyen, 2015; Meulders, Meulders, Stouten, De Bie, & Vlaeyen, 2017). These findings consistently suggest less sensitivity to specific sensory-perceptual evidence and a greater impact of symptom-related priors in patients with a somatic symptom disorder.
Other evidence is compatible with the idea that input from somatic and affective states is not very well distinguished in individuals with MUS, making them highly vulnerable to somatic priors when in a negative affective state. For example, when inducing negative affect through picture viewing followed by a symptom questionnaire that activates symptom-related priors, elevated symptom reports are elicited in these individuals, and this effect is mediated by the difficulty of identifying feelings (Van Den Houte et al., 2017). Moreover, harmless cues that have been associated with symptom episodes in a conditioning paradigm can elicit elevated symptoms by themselves. This happens particularly when the cues have a negative affective valence and in individuals with high trait N/NE and individuals with somatic symptom disorder (Devriese et al., 2000; Van den Bergh, Stegen, & Van de Woestijne, 1997).
In sum, the above pattern of results converges on the interpretation that individuals with high trait N/NE are more affected by the aversive affective-motivational component of a somatic episode and process sensory-perceptual information of a somatic episode in a less detailed way. This results in fewer precise prediction errors and leads to persistently active symptom-related priors that produce symptoms that are poorly related or unrelated to physiological dysfunction. We suggest that this is an instance of a BSTS strategy: Despite little sensory evidence, the brain jumps to the conclusion of an aversive somatic symptom being present. The benefit is a reduction of uncertainty at an early stage of the type “better the devil you know than the one you don’t” (Van den Bergh et al., 2017, p. 197). The downside, however, is little sensitivity to error correction by means of updating the prior expectation, which leads to persistent physical symptoms unrelated to actual physiological dysfunction (Henningsen et al., 2018). Again, MUS are in this respect not a separate phenomenon that is associated with high trait N/NE. It actually is the N/NE-related processing heuristics at work when processing somatic information.
Integration
Using a predictive-processing perspective, we interpreted four behavioral phenomena that are substantially associated with N/NE as exemplars of a BSTS strategy. We chose these phenomena because, unlike classic attentional and misinterpretation biases, they are not typically considered straightforward examples of such a strategy. However, looking at these phenomena from a predictive-processing perspective suggests that this strategy may also underlie these examples, allowing a more parsimonious interpretation of a larger range of behavioral facets of N/NE. Indeed, each of these examples can be reframed as emanating from the same deeply embedded processing heuristic that constitutes N/NE and produces the conscious experience of the person in the world (Petersen et al., 2015). We propose that this processing heuristic of individuals with N/NE gives more weight (precision weighting) to the affective-motivational aspects of the input at the expense of detailed sensory-perceptual processing. 4 Depending on the source and focus of threat, this general processing strategy finds its expression in different behavioral phenotypes: Perseverative cognition emerges in relation to information in working memory, reduced autobiographical memory specificity emerges in relation to information in long-term memory, and compromised fear learning and somatization emerges in relation to information from exteroceptive and interoceptive threats, respectively.
The obvious short-term benefit of this strategy goes along with long-term costs in the form of a stagnated error-reduction process: Little detailed sensory-perceptual processing results in low-precision prediction errors, which allows precise threat-related priors to dominate conscious experience and leads to insufficient error minimization when the priors are updated. Conversely, active inference (see above) leads perceptual systems to sample threat-related information consistent with prior expectations. This keeps the system in a chronic state of stress produced by expecting and perceiving threat with little chance for corrective experiences. 5
It is obvious that the term “strategy” should not be considered the result of a deliberate decision by a conscious self to avoid negative states but a more fundamental way of handling information that in some way appears threatening. This psychobiological strategy is genetically engraved in a substantial part of the population (individuals with high trait N/NE) and is epigenetically turned on in individuals with early and/or chronic adverse experiences (McEwen et al., 2016). It implies enhanced amygdala function and elevated neurohumoral levels (glucocorticoids, noradrenaline) that are associated with impaired prefrontal cortex function (Phelps, Lempert & Sokol-Hessner, 2014). Eventually, this strategy may gradually develop from goal-directed to habit-based (e.g., epistemic habit; Friston et al., 2016). This pervasive psychobiological strategy should be considered adaptive in view of both the biological history of our species and the history of individuals with early adverse experiences. Repeated unpredictable and/or chronic threats may have changed both the structural and functional properties of the brain to process threat-relevant information in such a way that it promotes the short-term benefit of rapid threat categorization and preparation for action without being outweighed by the less dramatic long-term negative “side effects.”
Clinical Implications
Considering N/NE to be a transdiagnostic trait variable that implies both elevated vulnerability for and a psychobiological dimension underlying internalizing psychopathologies leads to important questions such as how malleable the trait is, whether the trait and its consequences can be prevented and treated, and whether this strategy would be more proficient than targeting specific disorders and symptoms. Overall, it seems that N/NE is more malleable than generally believed and slowly changes over time. In both children and adults, significant small to moderate changes in N/NE have been observed that resulted from cognitive-behavioral therapy (CBT)-based interventions that targeted N/NE, and changes in N/NE have been shown to predict changes in symptoms and functional impairment but not vice versa (Barlow, Sauer-Zavala, et al., 2014a; Carl, Gallagher, Sauer-Zavala, Bentley, & Barlow, 2014; Farchione et al., 2012; Sauer-Zavala, Wilner, & Barlow, 2017). Recent meta-analytic evidence provides more solid evidence suggesting that N/NE can be altered by treatment with a moderate effect size (Cohen’s d = 0.50–0.60). The type of therapy did not matter much, and the change reached its maximum effects between 4 and 8 weeks (Roberts, Luo, et al., 2017).
Although these findings are promising, it remains important to know which components are critical and why and to design focused intervention protocols to further improve treatment effects. The Unified Protocol for Transdiagnostic Treatment of Emotional Disorders developed by Barlow and coworkers is intended to do this (Bullis, Fortune, Farchione, & Barlow, 2014; Ehrenreich-May et al., 2017; Ellard, Fairholme, Boisseau, Farchione, & Barlow, 2010). Conceiving N/NE as characterized by aversive reactions to emotional experiences and attempts to avoid and escape them, the treatment modules focus on extinguishing distress in response to strong emotions and the reduction of avoidant emotion-regulation strategies. The latter comes close to what would be needed within the current predictive-processing perspective. Assuming that N/NE reflects a stagnated error-reduction process, the goal then is to guide individuals to overcome the point at which error reduction gets stuck. This implies processing information that is associated with aversiveness thoroughly and with openness for detailed sensory-perceptual threat-related elements. This will enable prediction errors to modify threat-related priors and, in the long term, facilitate a more complete error-minimization process. Once modified, functionally more adaptive priors will in turn act as predictors for new input, allowing more efficient error minimization and explaining away unresolved mismatches between predicted and actual input. In other words, a more adaptive generative model about the world is formed that is characterized by less unresolved mismatches between expected and actual input.
However, high-level threat-related priors that are innate or learned to fit a survival goal may be quite strong and not easily changed. A predictive-processing framework considers priors to be forward models, that is, “embodied, whole brain representations” (Barrett, 2017, p. 7) that anticipate upcoming sensory events as well as the best action for dealing with the impending sensory events (Barrett, 2017; Barsalou, 2008). This means that they involve internal activation of encodings of actual perceptual, somatovisceral, and motor activity that promote prediction-error minimization by initiating responses that confirm predictions (active inference). These responses can be unpacked into a large array of lower-level component predictions that eventually initiate defensive-action programs, including proprioceptive, autonomic, and somatovisceral responses associated with defensive activation (Van de Cruys, 2017). We suggest that treatment interventions should affect the way that threat-relevant information is processed at all levels of the “machinery,” including altering defensive-action programs that are intrinsically part of the prior expectations. This emphasizes the necessity to coach individuals toward openness to process aversive information by including components that help the individual to disengage from defensive-action tendencies.
The above description is consistent with exposure-based interventions if conceived in a different way. Although exposure in CBT relies on an extinction rationale implying a “belief change” (e.g., that a feared stimulus is not coming and/or is not as bad as expected; Foa & McLean, 2016), in the current analysis it is not so much the information that a feared stimulus is not coming that makes the change. If defined as an attentive, open, and nondefensive way of processing threat-relevant information, it implies changing the fundamental attitude that characterizes the existence of high trait N/NE individuals in the world. In other words, it is a fundamental disengagement from defensive-response mobilization during processing of the aversive information, thereby changing associated psychophysiological and motor-response programs. Putting it this way reverses the sequence in some way: Defensive-response mobilization is not just something that automatically stops after one realizes that a dreadful stimulus is not coming. It is the disengagement from defensive-action tendencies during processing of the stimulus that makes it less dreadful: It means that the weight of the threat-related prior has been importantly reduced and that more weight is given to processing input in an open, nondefensive way. More detailed sensory processing will then promote updating the prior beliefs. This means that information processing is no longer inspired by a BSTS strategy but by a wait-and-see rationale. This reasoning implies that the threatening aspect and aversive impact of any kind of stimulus can possibly be reduced by this approach, regardless of whether it is a conditioned or unconditioned source of aversiveness. 6
The critical element advanced here—releasing stagnated error reduction by helping individuals to disengage from defensive-action tendencies during processing of aversive information—involves a fundamental change in psychophysiological response set to process potentially threatening information. This analysis has two important consequences. First, it suggests the prediction that any treatment should include this critical element to be effective and that sharpening and reinforcing operationalizations of this element will further improve treatment effectiveness. Second, it allows a common ground to be detected for many treatment strategies that aim to change habitual information processing of negatively valent information. For example, concreteness training (Watkins et al., 2012) aims at changing the abstract processing mode in rumination and perseverative cognition and at promoting concrete processing of difficult, upsetting events by focusing on sensory details, on the specific sequence of events, and on possible steps to take after the event. Memory-specificity training (MEST; Raes, Williams, & Hermans, 2009) is intended to change reduced autobiographical memory specificity by coaching participants to practice retrieving specific memories from cue words delivered across weekly sessions complemented by homework in between. Evidence suggests that MEST can improve autobiographical memory performance and subsequently reduce depressive symptoms (Hitchcock, Werner-Seidler, Blackwell, & Dalgleish, 2017).
In a similar vein, interoceptive-differentiation training has been developed as a treatment strategy to guide patients with MUS to become more sensitive to sensory-perceptual details of somatic sensations as a way to update constantly active symptom-related priors. Initial evidence suggests, for example, that heartbeat-perception training reduces somatization processes (Schaefer, Egloff, Gerlach, & Witthöft, 2014). In some way, the idea of making room for more detailed sensory processing without defensive-response mobilization is also captured by the concepts of “acceptance” and “self-compassion” (Forman & Herbert, 2009; Gilbert, 2009) and is consistent with a general overarching account of therapeutic change in psychotherapy (Lane, Ryan, Nadel, & Greenberg, 2015).
Finally, psychopharmacological treatment with serotonergic drugs such as selective serotonin-reuptake inhibitors (SSRIs) can also reduce behavioral manifestations of trait N/NE in healthy individuals (Ilieva, 2015). This is not surprising given serotonin’s intricate involvement in aversive affective processing and control, which is also revealed by manipulations such as tryptophan-depletion studies in humans (e.g., Ruhé, Mason, & Schene, 2007). However, the role of serotonin is complex, as shown by both positive and negative covariances between serotonin and aversion and their influence on neural plasticity and learning rates. This complexity has led researchers to use computational models to understand the regulatory effects of serotonin, suggesting that it is a signal associated with predictions and prediction errors for future aversive outcomes (Dayan & Huys, 2009; Iigaya, Fonseca, Murakami, Mainen, & Dayan, 2018). Here serotonin is particularly implicated in the coding of loss-related prediction errors, thereby inhibiting overreactions to negative outcomes and modulating behavioral choice selection to handle risk after negative events (Moran et al., 2018). Correspondingly, SSRIs are suggested to boost behavioral treatments by affecting the learning rate and increasing underlying neural plasticity to better deal with (potential) loss and aversion (Iigaya et al., 2018) or, in other words, by releasing stagnated error processing.
Summary and Future Directions
Evidence shows that the variation in psychopathology can be parsimoniously modeled using a limited number of hierarchically arranged dimensions. N/NE is a dimensional trait that appears as a large general vulnerability factor for psychopathology, particularly within the internalization spectrum (Conway et al., 2019). An important question is how to conceive of this trait. Any mechanistic interpretation should be (a) as parsimonious as possible and (b) able to explain a large variety of phenomenally distinct features of psychopathology. For this exercise we turned to a predictive-processing perspective that conceives of the brain as an active prediction-testing organ that tries to make sense of the stimulation it receives. Phenomenal reality emerges from two counterflowing streams of information: expectations (predictions or priors) and prediction errors (input). Prediction errors are propagated through the hierarchical processing architecture of the brain in a prediction-error-minimization process to eventually settle on a generative model that best explains the input. Both priors and prediction errors should be seen as neural distributions with a variance (precision) that determines the relative weight in the eventual generative model.
From this perspective, we describe trait N/NE as reflecting a stagnated error-reduction process, which, we suggest, results from a generalized BSTS strategy to process information that is associated with aversiveness. It is characterized by strong threat-related priors prompting active inference and low-detail sensory-perceptual and meaning analyses of the input facilitating rapid categorization of threat-related stimuli at the expense of updating threat-related priors. Although the resulting generative model may be adaptive in the short term, there is no room to develop a more adaptive generative model of the input in the long term. We argue that this generalized BSTS strategy underlies different cognitive and affective features and risk factors of psychopathology that are typically investigated in separate research lines.
In addition to more classic BSTS exemplars such as attentional and interpretational biases, these research lines include maladaptive perseverative cognition, reduced autobiographical memory, compromised fear learning, and poor symptom processing in somatization. Depending on contextual variables and individual concerns, some high trait N/NE individuals will be characterized more or less by these behavioral features. On the basis of this analysis, we suggest that a critical element in all treatment approaches is some kind of exposure, defined as an attentive, open, and nondefensive way of processing threat-relevant information. We also suggest that further and more powerful operationalizations of this critical element when processing specific concerns and fears may be a fruitful way to decrease trait N/NE and its consequences.
The above account obviously implies major challenges for research to test and validate. Because predictive processing is a (Bayesian) computational framework, it has stimulated interest in (and calls for) computational psychiatry/psychopathology and psychosomatics (Petzschner, 2017; Petzschner, Weber, Gard, & Stephan, 2017). Thus, one way to validate its claims is to flesh out a computational version of the predictive-processing model above involving a clear mechanistic description of the critical variables and their interactions, to run simulations, and to compare the results with evidence from real life. We recently started to do this by developing a computational model that accounts for bodily symptoms that maintain a strong, weak, or absent relationship with bodily input (Pezzulo et al., 2019; Pezzulo, Maisto, Barca, & Van den Bergh, 2020). This should also be done for other phenomena related to N/NE and subsequently tested with real data. Another approach is to experimentally test specific predictions made by the current account. This may require developing new paradigms that manipulate both categorical prior expectations and actual input to investigate the relative impact of priors on the eventual perception and subsequent cognitive processing (for examples related to interoception and symptom perception, see Petersen, Schroijen, Mölders, Zenker, & Van den Bergh, 2014; Van Den Houte et al., 2017; Zacharioudakis, Vlemincx, & Van den Bergh, 2020). These are only a few examples that may provide a way forward, but a huge challenge remains ahead. However, as Barrett (2017) states, “At the beginning, new paradigms raise more questions than they answer” (p. 14).