
Abstract
Previous research has shown that stimulus–response associations comprise associations between the stimulus and the task (a classification task in particular) and the stimulus and the action performed as a response. These associations, contributing to the phenomenon of priming, affect behaviour after a delay of hundreds of trials and they are resistant against overwriting. Here, we investigate their longevity, testing their effects in short-term (seconds after priming) and long-term (24 hr and 1 week after priming) memory. Three experiments demonstrated that both stimulus–classification (S-C) and stimulus–action (S-A) associations show long-term memory effects. The results also show that retrieval of these associations can be modulated by the amount of engagement on the same task between encoding and retrieval, that is, how often participants performed this task between prime and probe sessions. Finally, results show that differences in processing time during encoding are linked to the amount of conflict caused during retrieval of S-C, but not S-A associations. These findings add new information to the existing model of priming as a memory system and pose questions about the interactions of priming and top-down control processes.
Keywords
Introduction
A large proportion of our everyday actions rely on behavioural automatisms that allow us to execute actions without a need for elaborate cognitive processing. Such behavioural automatisms are essential to alleviate the cognitive load of the multitude of stimuli we are exposed to every moment. According to theories of associative learning, responding to stimuli in our environment leads to the automatic creation of representational links between stimuli and the responses made towards them, called stimulus–response (S-R) associations. These S-R associations have been widely studied and are known to contribute to the phenomenon of repetition priming (Henson, Eckstein, Waszak, Frings, & Horner, 2014). That is, for instance, in speeded classification tasks, participants are faster to respond to items consistently with the same response they previously performed on them than to respond with a different one (e.g., Dennis & Schmidt, 2003; Dobbins, Schnyer, Verfaellie, & Schacter, 2004; Logan, 1988, 1990). Such findings indicate that S-R associations are formed when we respond to stimuli and these associations are automatically retrieved when stimuli recur, improving performance when the same response is required (e.g., Dennis & Schmidt, 2003) and impairing it when another response is required (interference; Horner & Henson, 2011, 2012; Wendt & Kiesel, 2008).
S-R associations were initially thought to be rather short-lived direct links between a stimulus representation and a response representation. However, recent studies have shown that they can affect behaviour after a delay of hundreds of trials (Moutsopoulou & Waszak, 2013), they are resistant against overwriting (Moutsopoulou, Yang, Desantis, & Waszak, 2015), and they carry representations of both task-related and action-related information (Horner & Henson, 2009; Koch & Allport, 2006; Moutsopoulou & Waszak, 2012; Moutsopoulou et al., 2015; Waszak & Hommel, 2007; Waszak, Hommel, & Allport, 2003; Wylie & Allport, 2000). For instance, Moutsopoulou et al. (2015) used an item-specific priming paradigm to investigate the components of S-R associations more closely. Participants’ task was to classify everyday objects either according to their size or according to whether they contained a mechanism by pressing a left or right key in accordance with a preceding task cue (e.g., “S + L” for “small vs. large,” indicating that the left key should be pressed if the object was small). Crucially, between the prime instance(s) of a stimulus when S-R associations were established and between the item-specific probe instance(s) of a stimulus when S-R associations were tested several trials later, the action participants had to perform on a specific stimulus (left vs. right key press) and the task to be performed on the stimulus (size vs. mechanism classification) could independently repeat or switch. Moutsopoulou et al. (2015) found that both switches in action mapping and switches in task (classification) mapping were associated with prolonged reaction times (RTs) and increased error rates in comparison with repetitions. But the effects of switches in action mapping and switches in classification mapping between the prime and probe instances of a stimulus did not interact. This finding suggested two independent components of S-R associations: stimulus–action (S-A) associations between stimuli and motor outputs, and stimulus–classification (S-C) associations between stimuli and their task-specific classifications (e.g., small).
What is more, S-C associations are more rapidly formed and are much stronger than S-A associations, challenging the traditional view that S-R associations, at least from the beginning of their encoding, directly link the stimulus to only a unitary response representation (Henson et al., 2014; Hsu & Waszak, 2012; Moutsopoulou et al., 2015). Given this evidence on the complexity of S-R associations, the role of memory processes involved warrants further investigation. It is therefore interesting to investigate, how long these S-A and S-C associations are maintained in memory for and whether there are differences between S-A and S-C associations regarding the longevity of their effects.
Three experiments investigated the longevity of S-R associations. We adapted the paradigm of Moutsopoulou et al. (2015), previously used to investigate these two component processes of S-R associations (paradigm originally adapted from Hsu & Waszak, 2012). Participants were shown images of recognisable objects and were asked to perform one of two semantic classification tasks: Indicate whether in real life the object displayed was smaller or larger than a reference box, or indicate whether the object was mechanical or not. To respond, participants had to press a left or right key as fast as possible. During prime trials, the same item was overall presented twice within the same classification task and requiring the same response (same S-A and S-C mapping at encoding). During probes, the action and classification task for each specific object image could independently repeat or switch. Please note that switches do not refer to task switches in consecutive trials (like in the task-switching paradigm; see Kiesel et al., 2010, for an overview), but to switches of S-A and S-C mappings for specific images/items. Participants were not explicitly told of these possible switches. There were four possible item-specific switch conditions: a repetition of both action and classification, an action repetition with a classification switch, an action switch with a classification repetition, or a switch in both action and classification mapping between the prime and probe trials of a specific stimulus (see Figure 1). These conditions comprised an orthogonal manipulation of classification and action mapping between prime and probe. During the probe presentation of the stimulus (retrieval), which occurred two to seven trials after the item-specific prime presentations, RTs and errors were measured to assess whether there was a difference between repeating and switching the classification task and/or the action. With this orthogonal manipulation of category and action switching, we could investigate the effects of classification independent from action switching/repeating (by comparing the classification switch with the classification repetition conditions collapsing across action switching and action repetition conditions, henceforth called the classification or S-C effect). When this was significant and positive, participants were slower to respond when a classification switch was required than when the classification repeated and the action or S-A effect independent from classification switching/repeating, as well as the possible interaction of the classification and action effects which would indicate a retrieval of a bound representation of the two associations (e.g., an S-C-A association).
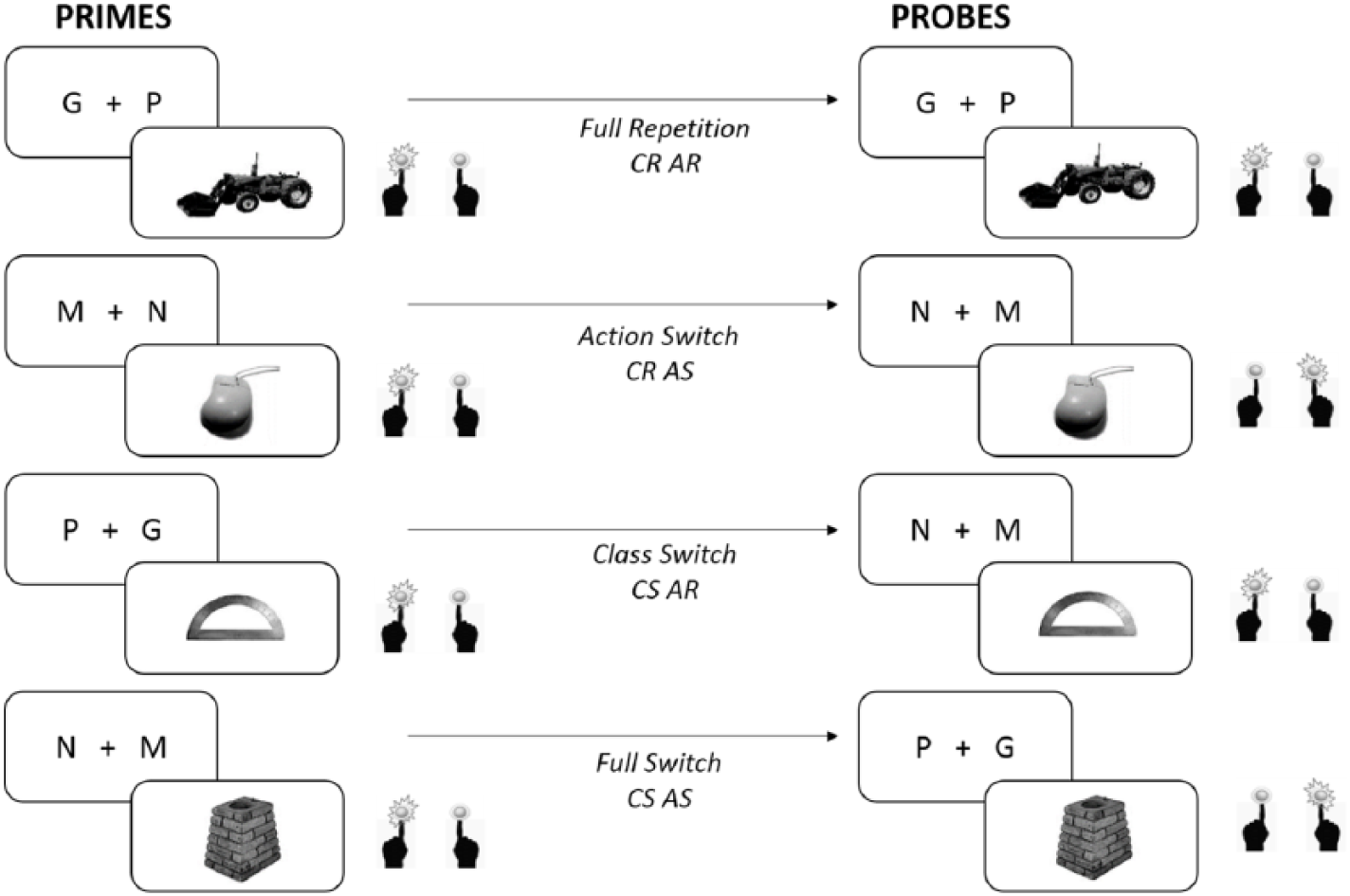
Demonstration of the switch conditions between primes and probes. Classification and action were orthogonally switched/repeated between prime and probe presentations item specifically. CR: classification repeat, CS: classification switch, AR: action repeat, AS: action switch.
Importantly, to investigate how long S-A and S-C associations were maintained in memory for, the probe presentation of the stimuli occurred in one of three probe phases: within the priming session (lag from second prime to probe: two to seven trials), 24 hr after priming, or 1 week after priming. Note that participants were not aware during the first session that stimuli could be probed with different S-R mappings nor that the second and third sessions would be on the same experiment. We hypothesised that S-C associations would be maintained and show traces in long-term memory, whereas S-A associations would be short-lived and only affect behaviour in Session 1.
In the first experiment, we tested S-C and S-A associations within these three probe sessions. The next two experiments were conducted to replicate the results of Experiment 1 and to further investigate the effects observed in Session 3. In particular, we manipulated how much practice participants had with the same task between sessions, to investigate whether more engagement with the task results in larger long-term memory effects 1 week after priming. This was done by comparing the effects produced when participants performed an irrelevant task between Sessions 2 and 3 (Experiment 2), to those when they performed one session of the same task between Sessions 2 and 3 (Experiment 1) and performing two sessions of the same task (Experiment 3).
Finally, memory function, and particularly working memory, has been investigated within the context of individual differences. Research has shown that individuals’ working memory capacity varies widely (e.g., Unsworth & Engle, 2007; Vogel, McCollough, & Machizawa, 2005). However, individual differences in the ability to maintain S-R associations have not been investigated. Yet, priming studies often show large variability in their results. In addition, S-R associations are investigated mostly in terms of facilitation/interference during presentation of a primed stimulus requiring the same or a different response. In Moutsopoulou et al. (2015), we showed that at least as far as the current paradigm is concerned, the priming effects observed might be due to interference/conflict rather than facilitation. In that study, stimuli were primed 4 times and probed 4 times as well. The results showed that RTs were stable after the first presentation of the stimulus across the rest of the primes and the probe trials when the action and classification were repeated (which was equivalent to eight presentations of the stimulus with the same task and action mapping). Conversely, RTs increased when there was a switch in classification task and/or action in the probe. This result showed that facilitation occurs from Prime 1 to Prime 2 but then RTs remain stable when full repetitions occur. Such conflict could depend on processes related to executive function (inhibition and shifting), and the efficiency of these processes has been shown to vary between individuals (Miyake & Friedman, 2012). To that end, in addition to the analyses concerning the longevity of S-C and S-A associations, we investigated whether there were individual differences in the item-specific S-A and S-C priming effects by testing whether differences in the level of learning during priming are associated with differences in the amount of conflict observed during retrieval. We, therefore, investigated whether the amount of encoding (as measured by the speeding up of responses from Prime 1 to Prime 2) would be indicative of the amount of conflict (as assessed by S-A and S-C effects) participants would show during S-R retrieval in the probe.
Experiment 1
Methods
Participants
A total of 20 right-handed participants were tested (14 were female, mean age = 26.7). They were paid at the rate of 10 euros per hour for their participation. They all gave informed consent for participating.
Stimuli
The stimuli were pictures of real, recognisable objects, taken from an online library (Brady, Konkle, Alvarez, & Oliva, 2008). A total of 576 pictures were used. Each belonged in one of two categories (small or large in real life in comparison with a box that was shown to the participants where small would fit in the 27 cm × 36.5 cm × 12 cm box and large wouldn’t). Each stimulus belonged additionally to another one of two categories in terms of its mechanicity (mechanical or non-mechanical). Thus, there were four categories of 144 stimuli each: small mechanical, small non-mechanical, large mechanical, and large non-mechanical. Each stimulus was presented 3 times in total: twice as a prime in Session 1 (with the same classification task and action mapping) and then once as a probe (either in Session 1, Session 2, or Session 3). Out of the 576 stimuli (each primed twice in Session 1), 192 were probed in Session 1, 192 in Session 2, and 192 in Session 3. Stimuli for Probe Sessions 1, 2, and 3 were chosen randomly with the constraint that each probe session (probes in Sessions 1, 2, and 3) included equal numbers of stimuli from each of the four categories (48 stimuli from each of the four categories).
Concerning the selection of stimuli, in Session 1, prime stimuli were the same across participants (all stimuli were presented to all participants in Session 1). However, which classification task and action mapping stimuli would be presented with was random (with the constraint of maintaining equal numbers of left and right responses in each of the cue conditions and for every stimulus category). Which of the stimuli would be probed in Session 1, 2, or 3 was also random with the constraints of an equal number of stimuli from each category being probed in each session and within each stimulus category, maintaining an equal number of left and right responses. This manipulation also meant that each task was used the same number of times.
All images (256 × 256 pixels) were presented in colour against a white background, centrally on a CRT monitor.
Design
The experiment consisted of three sessions Session 1 consisted of 1,344 trials in total (96 blocks of 14 trials each). The 14 trials of each block consisted of 12 prime trials (six stimuli presented twice each) and two probe trials (two out of the six primed stimuli were probed within the block). All primes within a block were presented in random order as were the probes with only two constraints: (1) that probes were presented after their corresponding primes and (2) that probes were presented with a distance of two to seven trials from the second prime presentation of their corresponding stimulus (the same distance as in previous studies; cf. Moutsopoulou et al., 2015). The four primes that were not probed within their block were later probed either in Session 2 or Session 3. Participants conducted three blocks of practice, which consisted of stimuli not presented in the main experiment, before beginning Session 1. Sessions 2 and 3 consisted of 192 trials each. In Sessions 2 and 3, all items presented were probes. To maintain the same structure as in Session 1, stimuli were presented in blocks of 14 trials.
As in Moutsopoulou et al. (2015), each stimulus was preceded by a cue consisting of two letters presented on the left and right of a fixation cross. When the letters presented were M and N, they indicated that the mechanism task should be performed (M = mechanical, N = non-mechanical). The letters G and P stood for grand (the French word for large) and petit (the French word for small), and indicated that the size classification task should be performed. The sides that the letters were presented on indicated the side of the response (if M was on the left and N on the right of the fixation, participants had to press the left button if they wanted to classify the object as mechanical or the right button if they wanted to classify it as non-mechanical). For Prime 1, the sides of the letters were randomised across trials, while maintaining an equal number of trials with each type of cue. Prime 2 trials were identical to Prime 1 in terms of the cue (in Prime 2, Prime 1 stimuli were repeated with exactly the same cue). Given the counterbalancing of probe trials in terms of switching/repeating the classification and/or the action (explained below), an equal number of trials for each cue was maintained also for probe trials. This method ensured that each cue was presented an equal number of times within the experiment. However, note that we did not control the order of the cues. Therefore, the number of task switch and task repeat trials from N – 1 to N was not strictly identical (we have previously investigated whether task-switching proper has an effect on the retrieval of item-specific S-C and S-A associations, and we have shown that our effects are independent of task-switching proper; see Pfeuffer, Moutsopoulou, Pfister, Waszak, & Kiesel, 2017).
The action and classification that participants had to perform upon a specific stimulus remained the same for the two prime instances. From prime to probe, S-A and S-C mapping independently repeated or switched so that four switch conditions emerged: action repetition–classification repetition, action switch–classification repetition, action repetition–classification switch, and action switch–classification switch. Each of the switch conditions occurred equally often (48 times) for the probes of each session. See Figure 1 for switch conditions and Figure 2 for a demonstration of the three prime–probe lags.
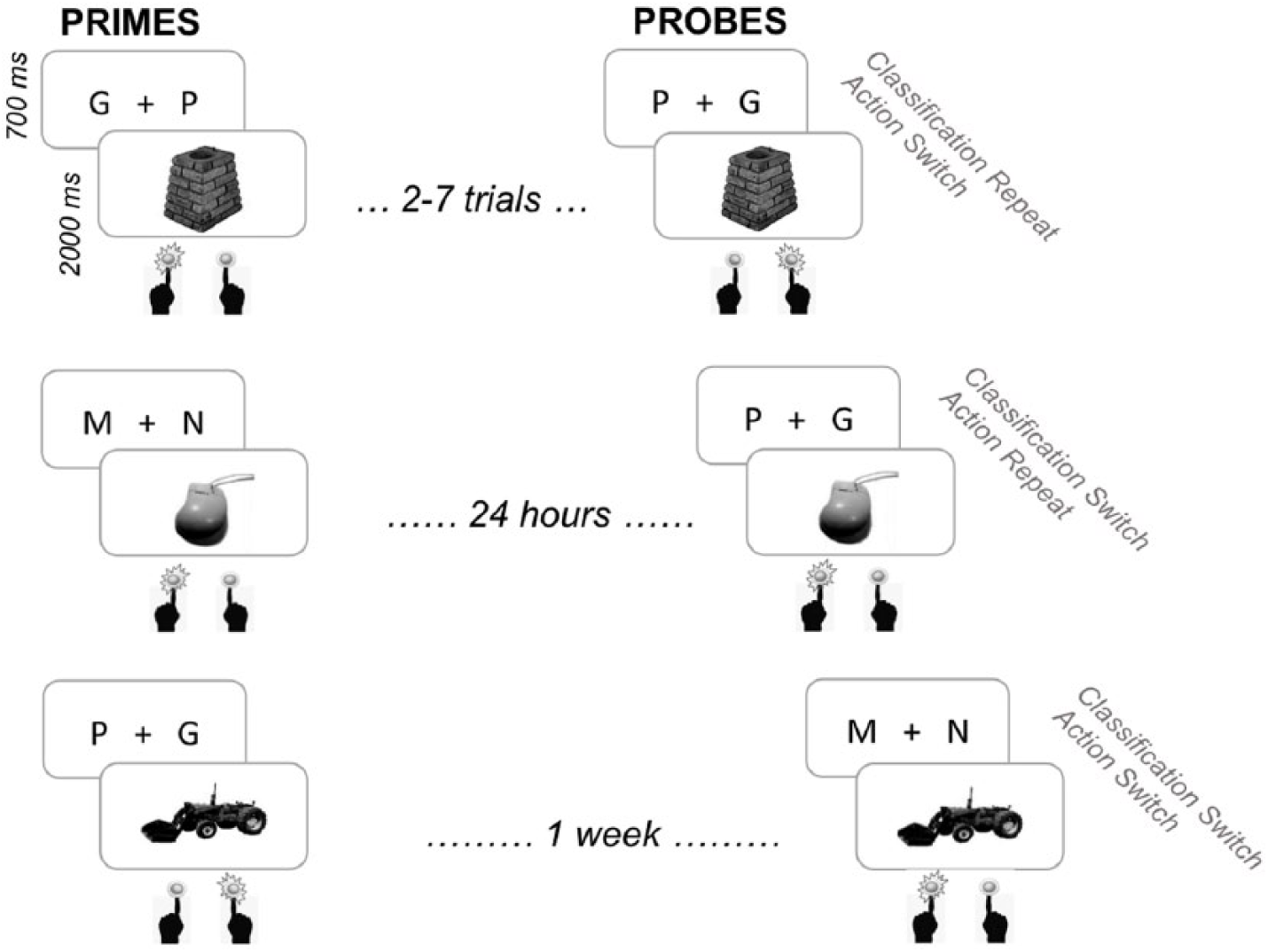
Structure of the lag manipulation. Items were primed in Session 1 (primed twice with the same classification and action mapping with a lag between 1 and 13 trials). Session 1 was identical in all three experiments. Within that session, one third of items was also probed (two to seven trials after their second prime presentation). In Session 2 (24 hr after priming), another third of items was probed. Session 2 was identical in Experiments 1 and 3. The final third of items was probed 1 week after priming. This session was identical in all three experiments (Session 3 in Experiments 1 and 2 and Session 4 in Experiment 3 because another interim session was presented in this experiment). Here, the example of probing in Session 1 displays an action switch trial, the example in Session 2 displays an example of a classification switch, and the example in Session 3 displays an example of a full switch. Note that there were also probe trials in which both classification and action associations were repeated (not displayed here; see Figure 1).
Procedure
Participants sat in a quiet room about 45 cm away from the screen. They were first instructed on how to perform the task. They were told that they would be presented with images of recognisable objects preceded by a cue. This cue indicated which task (size or mechanism classification) they would have to perform on the upcoming image and also indicated the sides of the responses. Participants were not told that objects would be repeated but were only instructed to focus on each cue and on classifying each object. The response keys were “A” and “L” on a QWERTY keyboard. Participants were instructed to always press the left key (“A”) with the left index finger and the right key (“L”) with the right index finger. They were told to be as fast and as accurate as possible.
The cue was presented for 700 ms, followed by the stimulus which was presented for 2,000 ms or until a response was made. The response was followed by feedback (“Correct!” printed in green indicated a correct response, “Incorrect” printed in red indicated an error and “pas de réponse”/“no response” when participants did not respond within 2,000 ms). Feedback was presented for 500 ms and was followed by the cue of the next trial. At the end of each block, participants saw a screen indicating how many trials out of 14 they responded to correctly and how many blocks they had completed out of the total number of blocks for that session. At this point, they had the opportunity to take a short break. To continue with the next block, they pressed the space bar.
Participants were told from the outset that the experiment would be three sessions long, but they were not told that the sessions would be related in any way. Therefore, participants had no explicit reason to maintain the presented items in memory.
Data processing
For the analysis of RTs, all trials associated to an error were removed. That is, when an error was made in one of the three presentations of an item or no response was made, all trials where this item was presented were removed from the analysis. Therefore, although the average error rate was 9.2% (standard deviation [SD] = 4.1%), a total of 23.3% (SD = 9%) of trials were removed due to errors. This way, associative learning was investigated without any interference from errors in previous presentations of an item. Another 1.2% of the remaining trials were removed for being above or below 3 SDs from the mean RT (means calculated for each subject and each condition). For error rate analyses, items with erroneous responses in the prime presentations were removed. Therefore, the percent errors calculated were based on the errors committed in the probe out of all probe trials that were not associated to an error in the prime. Out of all probe trials, 16.97% were removed due to errors made in the prime across subjects. All reported t tests were two-tailed, and for the analyses of variance (ANOVAs), Greenhouse–Geisser corrections were applied where necessary.
Results
Primes
Analysis of RT showed that participants became significantly faster at responding from Prime 1 to Prime 2, t(19) = 13.20, p < .001, d = 2.95. See Table 1 for RTs in each condition of the primes and probes.
Mean reaction times in milliseconds (and standard errors) in primes and each condition of the probes, for Experiments 1, 2, and 3.

Similarly, participants made significantly more errors in the first presentation of the primes than the second, t(19) = 12.40, p < .001, d = 2.77, showing a significant improvement in performance from Prime 1 to Prime 2. See Table 2 for error rates in each condition of the primes and probes.
Mean percent errors (and their standard errors) in primes and each condition of the probes, for Experiments 1, 2, and 3.

Percent errors in the probe were calculated on trials were errors were made, after having excluded all trials associated to errors in the prime.
Probes
Probe RT analysis
A repeated-measures ANOVA was conducted with the factors of session (Session 1, Session 2, and Session 3), classification (repeat, switch), and action (repeat, switch). Significant main effects of classification, F(1, 19) = 51.73, p < .001,
Given that both classification and action interacted with session, suggesting that their effects differed in size across sessions, the classification and action effects were investigated in each session separately. Significant effects of both classification and action were found in Session 1—classification: F(1, 19) = 33.40, p < .001,
To investigate whether the classification and action effects were significantly different between sessions, further ANOVAs were conducted to compare the effects between each pair of sessions.
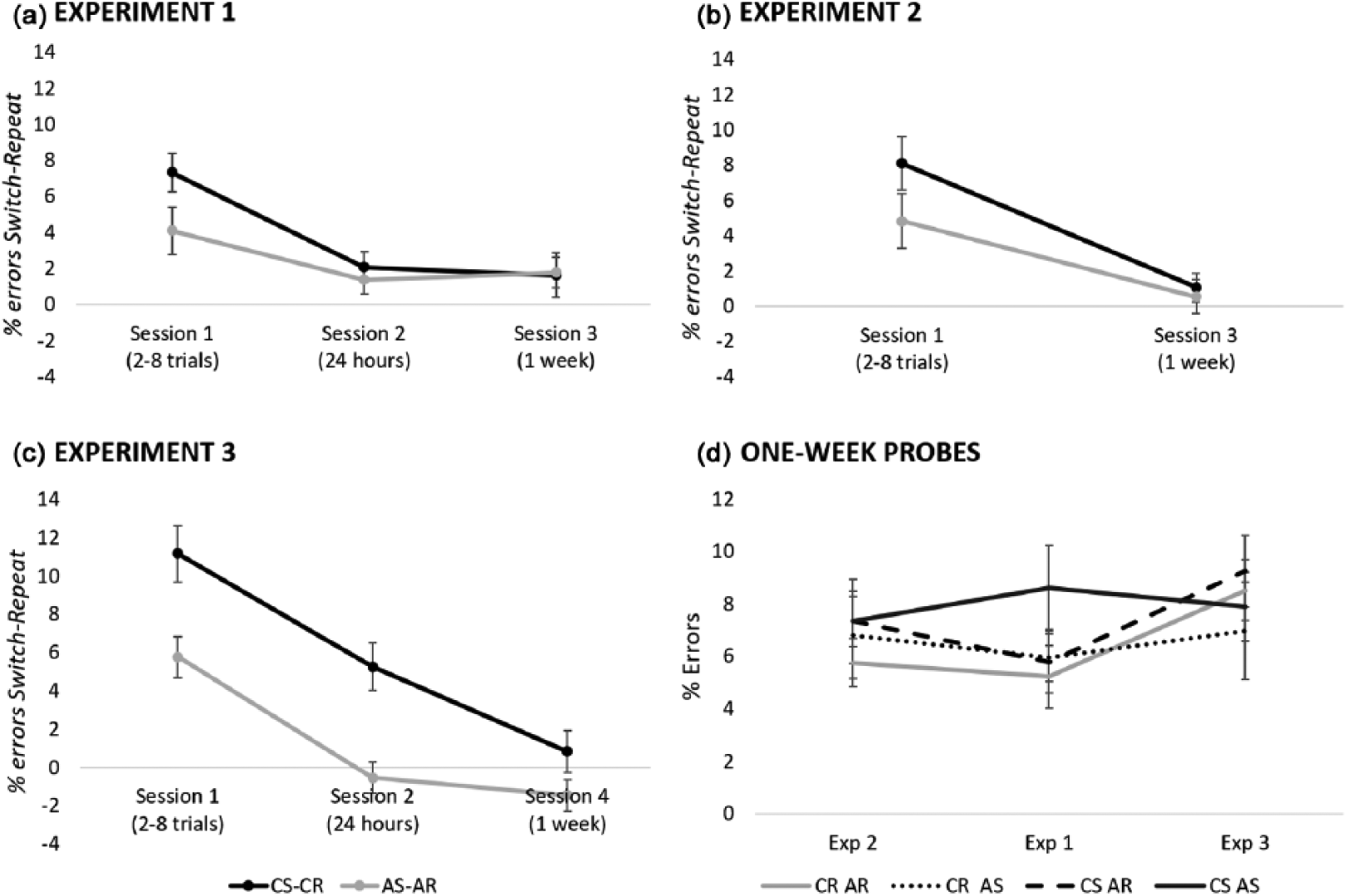
The classification effect (RTclassification switch – RT classification repetition) was significant in both Sessions 1 and 2, yet it was significantly stronger in Session 1, t(19) = 4.36, p < .001, d = 0.97. Although there was a significant classification effect in Session 2 but not in Session 3, the two effects did not significantly differ, t = 0.80, p = .433, d = 0.18. See Figure 2 for the classification and action effects across sessions.
The action effect

Probe RT switch costs (RT switch – RT repeat) for switches in classification (black) and action mapping (grey) from prime to probe in (a) Experiment 1, with RT switch costs displayed separately for stimuli probed in Sessions 1, 2, and 3; (b) Experiment 2, with RT switch costs displayed separately for stimuli probed in Sessions 1 and 3; (c) Experiment 3, with RT switch costs displayed separately for stimuli probed in Sessions 1, 2, and 4; (d) probe RTs of the Session 3/4 (1 week after priming) are displayed separately for the three experiments and four switch conditions (CR: classification repeat; CS: classification switch; AR: action repeat; AS: action switch). Error bars represent standard errors.
Probe error rate analysis
The error rates were very low as the main focus of these experiments was to investigate effects on RT. However, for completion, we performed the same analyses on the error data as well. A repeated-measures ANOVA was conducted on the percentage of errors committed in the probe, with the factors of session (Session 1, Session 2, Session 3), classification (repeat, switch), and action (repeat, switch). Main effects of session, F(2, 38) = 15.02, p < .001,
To investigate the Classification × Session interaction further, the classification effect was tested in each session. A significant difference between classification switching and repeating was found only in Session 1, t(19) = 6.95, p < .001, d = 1.55, and Session 2, t(19) = 2.46, p < .05, d = 0.55, while the classification effect in Session 3 did not reach significance, t = 1.30, p = .209, d = 0.29. See Table 2 for mean error rates in each condition and Figure 4a for a depiction of the classification and action effects in each lag condition.
Mean percent errors made in the probes. (a)-(c) switch effects (switch minus repeat) for switches in classification (black) and action mapping (grey) from prime to probe in (a) Experiment 1 with switch effects displayed separately for stimuli probed in Sessions 1, 2, and 3; (b) Experiment 2, with switch effects displayed separately for stimuli probed in Sessions 1 and 3; (c) Experiment 3, with switch effects displayed separately for stimuli probed in Session 1, 2, and 4; and (d) probe percent errors in Session 3/4 (1 week after priming) are displayed separately for the three experiments and four switch conditions (CR: classification repeat; CS: classification switch; AR: action repeat; AS: action switch). Error bars represent standard errors.
Overall, error data differed from the RT results in three respects: Participants made fewer errors as sessions progressed while they did not become faster. In addition, errors did not reveal an effect of S-A in Session 3 as it was observed in RTs. The lack of this effect in error rates may be due to the overall small percentage of errors and therefore lack of power for this analysis.
Discussion
Experiment 1 replicated our previous findings suggesting that S-C and S-A associations were retrieved independently (Moutsopoulou & Waszak, 2012; Moutsopoulou et al., 2015; Pfeuffer et al., 2017), as the observed S-A and S-C effects did not interact. In addition, the results showed that regarding the longevity of S-R components, both S-A and S-C associations show effects in long-term memory. The S-C effect was present 24 hr after priming had occurred and without any explicit instruction for memorisation, replicating previous findings (Pfeuffer et al., 2017). Given the number of items that were primed in Session 1 (576 in total), it is remarkable that such effects would be observed so long after priming each item only twice. What was even more remarkable was the effect of S-A in Session 3 which occurred 1 week after priming. However, the pattern of the S-A effect across time, that is, the presence of an effect in Sessions 1 and 3, but not in Session 2, was also surprising. One explanation could be that the absence of an effect 24 hr after priming may be due to consolidation processes, which would transfer information into longer term memory systems. However, if we assume this effect to be due to consolidation, we would expect most of the consolidation to take place during sleep the night after priming occurred (for a review on the relationship between memory consolidation and sleep, see Walker & Stickgold, 2004). In this case, we would still observe an effect 24 hr after priming.
Another possible hypothesis is the following: participants did not know at the end of Session 1 that Session 2 would be on the same task and therefore, certain information may not have been maintained in memory. However, by the end of Session 2 (which was on the same task) they could assume that session 3 would also involve performing the same task (given that Sessions 1 and 2 were the same, it was likely the third session would be the same). Therefore, performing Session 2 may have activated memory processes which aided retrieval in Session 3. To assess whether this was the case, we conducted a second experiment, identical to Experiment 1, only now Session 2 was replaced by an irrelevant filler task (filling in questionnaires). We expected that this could eliminate the S-A effect 1 week after priming.
Experiment 2
Methods
Participants
A total of 20 participants were tested (five males). All were right-handed (mean age = 26.7). They were paid as in Experiment 1 and gave written consent prior to their participation.
Stimuli
This experiment was almost identical to Experiment 1. However, during Session 2, participants were given a completely different, filler task to do, that is, they filled in some irrelevant-to-the-task questionnaires. To maintain consistency with Experiment 1, the same number of stimuli was primed in Session 1 (although 192 of them were never probed because Session 2 was replaced by a filler task). Given the difference of tasks between Sessions 1 and 2, we assume participants did not anticipate to perform a task similar to Session 1 in the third session which took place as previously, 1 week after initial priming.
Data processing
For the analyses of RTs, errors were removed for all trials associated to an error as previously. The average error rate was 9.06% (SD = 4.8%); a total of 20.89% (SD = 9.3%) of trials were removed due to errors. A further 1.2% of the remaining trials were removed for being above or below 3 SDs from the mean RT (means calculated for each subject and each condition). Concerning the analyses of errors, these were performed on proper errors performed in the probe after removing the trials associated to errors in the prime, as in Experiment 1. Out of all probe trials, 15.39% were removed due to errors made in the prime across subjects. All reported t tests were two-tailed.
Results
Primes
Analysis of RTs showed that participants became significantly faster at responding from Prime 1 to Prime 2, t(19) = 15.2, p < .001, d = 3.41.
Similarly, analysis of error rates showed that participants made significantly more errors in the first presentation of the primes than the second, t(19) = 9.72, p < .001, d = 2.17, showing a significant improvement in performance.
Probes
All probe analyses in this experiment concerned only Sessions 1 and 3, given that Session 2 was irrelevant to the analyses.
RT analysis
A repeated-measures ANOVA was conducted with the factors of session (Session 1, Session 3), classification (repeat, switch), and action (repeat, switch). Significant main effects of session, F(1, 19) = 8.59, p = .009,
Given that both classification and action interacted with session, suggesting that their effects differed in size across sessions, the classification and action effects were investigated in each session separately. Significant effects of both classification and action were found in Session 1—classification: F(1, 19) = 54.18, p < .001,
Error rate analysis
A repeated-measures ANOVA was conducted on percent errors with the factors of session (Session 1, Session 3), classification (repeat, switch), and action (repeat, switch). The main effects of session, F(1, 19) = 13.79, p < .001,
To investigate the interactions between classification and session, and session and action, the classification and action effects were tested in each session. Paired-samples t tests showed that there were significant classification and action effects in Session 1: classification: t(19) = 5.36, p < .001, d = 1.20; action: t(19) = 3.14, p = .005, d = 0.70. However, no significant effects were observed in Session 3: classification: t(19) = 1.28, p = .215, d = 0.29; action: t(19) = 0.55, p = .590, d = 0.12. Overall, the error data analyses mirrored the results observed in the RT analyses for this experiment.
Discussion
This experiment replicated the independent S-C and S-A effects observed in Session 1 of Experiment 1. Importantly, as expected, Experiment 2 showed that in the absence of an interim session, which would non-explicitly indicate some relevance to maintaining associations, there was no effect of S-A retrieval in Session 3 1 week after priming. However, not observing the effect does not necessarily mean that this was due to the lack of the interim session. Therefore, we conducted another experiment in which, based on the same hypothesis, we would replicate the S-A retrieval effect 1 week after initial priming observed in Experiment 1. If S-A associations in Experiment 1 were somehow made available to memory due to the relevance of Session 2, then adding an extra session of the same experiment prior to the 1-week testing phase should result in an S-A effect as in Session 3 of Experiment 1, and this S-A effect might be even stronger in Experiment 3 as compared with Experiment 1 due to the increase of engagement in the same task.
Experiment 3
This experiment was identical to Experiment 1; however, here we added another session, which occurred 48 hr after priming. This session was of the same structure as Session 2 and Session 3 of Experiment 1 (192 trials long, divided into blocks of 14 trials each). However, the stimuli presented in this session (Session 3 in this experiment) were items that were not primed in Session 1.
Methods
Participants
A total of 20 participants were tested (seven males). All were right-handed (mean age = 27.7 years).
Stimuli
The only difference in this experiment compared with Experiment 1 was the addition of a session which took place 48 hours after priming. Here, 192 new items were presented which were selected from the same online library (Brady et al., 2008). There were equal numbers of stimuli from each of the four categories (small mechanical, large mechanical, small non-mechanical, and large non-mechanical).
Data processing
For the analyses of RTs, errors were removed for all trials associated to an error as previously. The average error rate was 10.8% (SD = 5.2%); a total of 26.9% (SD = 10.17%) of trials were removed due to errors. A further 1.2% of the remaining trials were removed for being above or below 3 SDs from the mean RT (means calculated for each subject and each condition). Analyses on error rates were performed on proper errors performed in the probe after having removed trials that were associated to errors in the prime. Out of all probe trials, 18.54% were removed due to errors made in the prime across subjects. As for Experiment 1, all reported t tests are two-tailed, and for the ANOVAs, Greenhouse–Geisser corrections were applied where necessary.
Results
Primes
As in the previous experiments, participants became significantly faster at responding from Prime 1 to Prime 2, t(19) = 12.03, p < .001, d = 2.69.
Participants also made significantly more errors in the first presentation of the primes than the second, t(19) = 15.52, p < .001, d = 3.47, showing a significant improvement in performance.
Probes
In this experiment, we compared RT and error rates in Sessions 1 and 2 (as in Experiment 1), and in Session 4 which corresponded to Session 3 in Experiments 1 and 2 as it also occurred 1 week after Session 1 (Session 3 in this experiment was a session that we did not analyse and occurred 48 hr after Session 1).
RT analysis
A repeated-measures ANOVA was conducted with the factors of session (Session 1, Session 2, Session 4), classification (repeat, switch), and action (repeat, switch). Significant main effects of session, F(2, 38) = 19.01, p < .001,
Given that both classification and action interacted with session, suggesting that their effects differed in size across sessions, the classification and action effects were investigated in each session separately. We additionally investigated possible interactions between classification and action to follow up on the nearly significant Classification × Action interaction reported above. Even though the triple interaction with session was not significant, it was likely, given the pattern of results, that a nearly significant classification by action interaction would be observed in each session. Therefore, to compare the results with the previous experiments in terms of whether the classification and action effects interacted or not in each session, we performed two-factor ANOVAs in each session. Significant effects of both classification and action were found in Session 1—classification: F(1, 19) = 63.69, p < .001,
Error rate analysis
A repeated-measures ANOVA was conducted on percent errors committed in the probe, with the factors of session (Session 1, Session 2, Session 4), classification (repeat, switch), and action (repeat, switch). Main effects of session, F(2, 38) = 14.39, p < .001,
To investigate the significant interactions further, the classification and action effects were tested in each session. Paired-samples t tests showed that there was a significant classification effect in Session 1, t(19) = 7.53, p < .001, d = 1.68, and Session 2, t(19) = 4.23, p < .001, d = 0.95, but not in Session 4, t(19) = 0.75, p = .461, d = 0.16. The action effect was significant in Session 1, t(19) = 5.36, p < .001, d = 1.20, was not significant in Session 2, t(19) = 0.66, p = .517, d = 0.15, and it approached but did not reach significance in Session 4, t(19) = 1.79, p = .089, d = 0.40. See Table 2 for mean error rates in each condition and Figure 4b for the classification and action effects across sessions.
Overall, there were some differences in this experiment when comparing the error rate with the RT results. The error rates revealed an overall effect of S-A, which was not found in the RT results. Other than that, the results were similar between the two analyses except in Session 4. RT analyses revealed an interaction between classification and action in this session, whereas error rates showed no significant effects. This difference may be due to a lack of power concerning error data in the fourth session.
Discussion
This experiment replicated the findings for Sessions 1 and 2 of Experiment 1: Independent effects of S-C and S-A retrieval were observed in Session 1, whereas S-C retrieval was observed in Session 2. However, Session 4 showed a new pattern of results: S-C and S-A effects interacted and the pattern observed indicated that participants were slower to perform the task when the S-C mapping was switched and the S-A mapping was repeated. Although this pattern is not the same as the S-A effect observed in Experiment 1, it still indicates that there are traces of associations that can affect behaviour even 1 week after priming. The reason why here we observed an interaction may have been because participants had more practice in doing the task and were faster to respond or because more practice somehow facilitated the binding of S-C and S-A information.
Analyses across experiments
RT performance in Session 1 did not differ between experiments—Prime 1: F(2, 57) = 1.72, p = .19,
Modulation of long-term retrieval by task practice
To address this question, we conducted analyses on the effects in the 1-week session, comparing performance between the three experiments. In Experiment 1, participants had one session of the same task between encoding and retrieval. In Experiment 2, they had no relevant sessions (instead, they had one irrelevant experimental session). In Experiment 3, they had two sessions of the same task before being probed 1 week later.
A repeated-measures ANOVA on the data of the 1-week session with the within-subjects factors of classification (switch, repeat) and action (switch, repeat), and Experiment (1, 2, 3) as a between-subjects factor, yielded a main effect of Experiment, F(2, 57) = 4.06, p = .022,
Further ANOVAs comparing effects within pairs of the three experiments showed that the action effect was significantly stronger in Experiment 1 compared both with Experiment 2, F(1, 38) = 7.19, p = .011
Do individual differences in prime RT predict effects in the probe?
The findings reported so far indicate that S-C and S-A associations are not only automatically created, but they have both short-term as well as long-term memory effects (up to 1 week after initial priming). The following analyses address possible individual differences. Does the speed of response in the prime, or the steepness of encoding from one prime to another, predict the amount of interference observed in the probe? Does this prediction apply to both short-term and long-term memory retrieval? Note that 10 correlation tests were performed (see below). Applying a Bonferroni correction, any comparisons with p < .005 are significant.
To answer this question, we performed some analyses across experiments. Given that Session 1 was identical across the three studies, the data of 60 subjects were collapsed to investigate whether individual differences in probe performance and RTs at encoding (i.e., the primes) could be informative in predicting short-term retrieval (i.e., performance in Session 1 probes). In particular, we investigated whether performance in the first prime and the steepness of encoding (as expressed by the difference of RT in Prime 1 and Prime 2) were indicative of the size of the classification and action effects during retrieval. Correlation tests revealed that performance in Prime 1 correlated positively with the classification effect in the probe in Session 1, r(59) = .52, p < .001, whereas it did not correlate with the action effect, r(59) = .16, p = .210 (Figure 5 displays correlation scatterplots for all the significant effects reported). Therefore, the faster the participants in Prime 1 (the shorter their RT), the smaller the classification effect they showed in the short-term probes (the smaller the difference between classification switch and classification repetition). The same effects were observed when looking at the difference between Prime 1 and Prime 2. This difference correlated positively with the classification effect in the probes in Session 1, r(59) = .36, p = .004, but did not correlate with the action effect, r(59) = .04, p = .736.

Scatterplots of significant correlations observed related to performance (RT of correct responses) in Prime 1 (Plots a, b, and c) and related to the amount of improvement participants showed in terms of RT between Prime 1 and Prime 2 (Plots d and e). Plots a and d show correlations of the stimulus–classification effect observed in Session 1. Plots b, c, and e show correlations with the amount of forgetting (how much interference by associations reduced) between Sessions 1 and 2 (Plot b), and between Sessions 1 and 3 (Plots c and e).
To investigate whether performance in the prime was also indicative of longer term retrieval, prime performance was correlated with the classification effect observed in Session 2 in Experiments 1 and 3. Absolute performance in Prime 1 did not correlate with the classification effect in Session 2, r(39) = .15, p = .346, and the correlation with the priming difference score (Prime 1 – Prime 2) was not significant (not below the .005 alpha level defined by the Bonferroni correction), r(39) = .33, p = .036.
The difference in size of the classification effect between Session 1 and Session 2 is also informative as it indicates the amount of “forgetting” for each participant from one day to the next. To investigate whether performance in the prime can be indicative of the amount of “forgetting” between Sessions 1 and 2, correlation tests were performed showing that performance in Prime 1 positively correlated with the size of the difference between the classification effect in Sessions 1 and 2 (the higher the difference between Sessions 1 and 2, the more the association was “forgotten” between the two sessions), r(39) = .45, p = .004. However, a correlation was not observed between the classification difference and the Prime 1 – Prime 2 score, r(39) = .24, p = .129.
Performance in Prime 1 also correlated with the amount of forgetting of S-C associations from Session 1 to the session 1 week after priming (Session 3 in Experiments 1 and 2 and Session 4 in Experiment 3), r(59) = .47, p < .001. The learning steepness between Prime 1 and Prime 2 was also informative in predicting the difference in size of the classification effect between Sessions 1 and 3/4, r(59) = .37, p = .004.
Discussion
The analysis of the effects in the 1-week session, across the three experiments, showed that the retrieval of associations as long as 1 week after priming is modulated by the amount of practice participants had with the task overall across that week. This analysis indicated not only that item-specific across-task priming effects have a very long lifespan but also that their retrieval may be modulated by the amount of engagement with the same task. This finding may be indicating a top-down influence on the retrieval of S-R associations: The more a task is performed, the more information encoded during the task are maintained and retrieved.
The lack of a correlation with the pure classification effect in Session 2, but a significant correlation with the difference of Session 1 – Session 2, shows that performance in the prime is not informative about whether one will retrieve the association one day later, but it is informative about the amount of forgetting in the long term.
General discussion
Three experiments studied the longevity of S-R associations and their component processes, as well as whether individual performance during encoding can predict the size of item-specific classification and action effects observed in the probe. The main findings can be summarised in four aspects. First, both classification (S-C) and action associations (S-A) show effects a few seconds after priming, confirming that both associations are maintained in short-term memory. These effects were additive. Second, S-C associations were present even 24 hr after priming showing long-term memory storage of these associations even with only two prime presentations. This was not the case for S-A associations. Third, traces of associations were detected even 1 week after priming. This effect depended on how much participants engaged in the same task during the week preceding probe testing. This suggests that practice with the same task can either help maintain associations in memory or at least make them more easily detectable behaviourally. Similarly, S-C and S-A associations may be integrated into a single representation in the long term or a speeding up of RTs by extra practice causes detection of a bound representation rather than independent effects. Finally, the speed of responding during encoding, as well as the steepness of RTs from Prime 1 to Prime 2, is informative about how much interference a participant will show in the short term due to automatic retrieval of task information and how much he or she will forget in the longer term. Below, we discuss each point in turn.
The short-term S-C and S-A effects we observed replicate previous findings showing that both classification and action information are automatically associated with a stimulus without explicit instruction, within one to two prime presentations, and can be independently retrieved (Hsu & Waszak, 2012; Moutsopoulou & Waszak, 2012; Moutsopoulou et al., 2015; Pfeuffer et al., 2017). The presence of S-C associations 24 hr after priming demonstrates long-term storage of S-R links, but only regarding information on the semantic classification of the visual stimuli. This effect has been previously observed (Pfeuffer et al., 2017). The effects observed 1 week after priming, however, have not been previously reported.
Although the pattern of results in the 1-week session (Session 3/4) is not clear due to the different effects observed in each experiment, there was a modulation of retrieval 1 week after testing depending on how much experience participants had with the same task between the initial prime session and the probe session 1 week later, demonstrating a context effect on the long-term maintenance of S-R associations. Which information was exactly maintained is not as clear, however. One possible explanation for the observed pattern is that S-A associations were boosted by performing the same task between prime and probe (i.e., action information maintenance may be more susceptible to top-down control), whereas increased engagement in the same classification task may lead to integrating classification and action information to improve efficiency. However, the interaction observed in Experiment 3 (Session 4) may also be due to the significantly faster overall RTs. Generally, the main finding of Session 3/4 was that S-R effects can be maintained for periods much longer than previously thought. Long-term priming traditionally referred to retrieval after some hundreds of trials (cf. Schacter, Dobbins, & Schnyer, 2004; Waszak et al., 2003). The presence of effects after 1 week as well as the modulations by the amount of engagement in the same task suggests that S-R priming effects must be integrated into long-term memory models, taking into account the different types of associations that may be involved. Furthermore, studies have shown that memory consolidation processes can differ for different types of memories and in particular between procedural and declarative memories (Plihal & Born, 1997; Stickgold, 2005). It is therefore likely that consolidation of the various types of S-R components differs as well.
Further to the question of what information is carried within each type of association, previous studies have identified a third type of association built during a priming event. Horner and Henson (2009) used a paradigm in which not only the classification and action were manipulated between the prime and probe presentations of a stimulus, but they additionally manipulated the decision level. In their paradigm, the classification task was indicated by a cue, which posed a question (e.g., “smaller than a shoebox?”) to which participants responded with a Yes or No (decision level) and indicated their decision by pressing a left or right key (action level). That is, the referent for comparison changed between study and test phase to allow for a separate assessment of classification, action, and decision. Using this paradigm, Horner and Henson (2009) demonstrated independent effects of classification, action, and decision on RTs. In contrast to Horner and Henson’s studies, in our paradigm, we did not separate out this level of association. Participants could themselves choose whether to frame the task as, for instance, “smaller than?” or “larger than?” Therefore, the level of decision in this paradigm (given that there was no separate Yes/No question posed) would be part of the classification process, and thus what Horner and Henson (2009) defined as decision-level associations would likely be part of the S-C association here.
Finally, the observed differences in retrieval correlated with performance in the prime/encoding phase. These differences were observed both in relation to absolute speed in responding to a new stimulus, as well as in relation to the amount of speeding up of responses between Prime Presentations 1 and 2. We found that the faster the participants were in the prime, the smaller the S-C effect they showed in Probe Session 1 (additionally, the faster they were in the prime, the smaller the differences between the classification effect in Sessions 1 and 2 and Sessions 1 and 3/4). This was not the case for S-A effects, showing that longer processing times affect S-C associations but not the strength of the association between the stimulus and the action. Similar correlations were observed when investigating the relationship of the amount of speeding up from Prime 1 to Prime 2 and the retrieval effects of the S-A and S-C associations. Again, a relationship was observed for classification retrieval and not for action. The larger the difference between Prime 1 and Prime 2 RTs, the larger the classification effect in the probe in Session 1. Thus, the amount of retrieval in the short-term of S-C associations was increased both with longer processing time in the prime and also with steeper RT increases from Prime 1 to Prime 2 during encoding. This finding suggests that the longer the time taken to respond, the more we encode with repeated priming, and the more likely we are to encode relationships between stimuli and semantic tasks, but not between stimuli and motor codes. It is therefore likely that S-C associations are susceptible to attentional and working memory capacity limitations, whereas S-A associations are not. This difference between S-C and S-A may be due to the fact that S-C associations in this paradigm involved pulling on declarative semantic knowledge about the objects. However, S-A associations were rather procedural.
Associative memory traces created through priming have not been widely investigated in the long term. Although significant progress has been made using classic recall and familiarity paradigms that investigate recognition memory of declarative information as well as procedural memory with classic paradigms like the Serial Reaction Time Task (SRTT; for comprehensive reviews, see Marshall & Born, 2007; Walker & Stickgold, 2004), associative priming has not been investigated in the same way. It is therefore tempting to speculate, based on the results of the current experiments, about whether the types of associations manipulated here share some of the memory properties of declarative and procedural memories in terms of their consolidation.
S-C associations in this paradigm required at least partial identification of objects, pulling on long-term preexisting declarative semantic knowledge about objects, their size, and their function. Conversely, S-A associations were more arbitrary (e.g., associating the image of an apple with a left key press). It is therefore likely that S-A associations did not benefit in the same way as S-C associations from the 24-hr separation between priming and Session 2 probe testing, explaining the observation of an S-C effect in Session 2 and no effect of S-A. Alternatively, the mere strength of S-A associations may not have been enough to maintain a memory trace. To exclude this possibility, further studies would have to investigate whether strengthening S-A associations by introducing more prime repetitions makes them more likely to survive a 24-hr delay. Nonetheless, this finding shows a possible difference in the consolidation of S-C and S-A. This idea goes along with many studies to date, which have shown that procedural and declarative memories differ in terms of consolidation processes (e.g., Plihal & Born, 1997; Tucker et al., 2006).
The reappearance of an S-A effect in Session 3 of Experiment 1 and the interaction observed in the final session of Experiment 3 is an interesting but confusing finding. When it comes to memory processing, it has been long suggested that short-term and long-term memory systems may function in parallel rather than sequentially (Müller & Pilzecker, 1900; see also McGaugh, 2000). It is therefore likely that what was measured in Sessions 2 and 3 are not the same sequential processes. Another possibility is that motor-related associations take longer to consolidate. It has been suggested that learning motor skills kicks off neural representations of internal models which are not stable but change over time and transfer, for example, from cerebellar regions to other parts of the cortex. This transfer may result in the memory trace taking variant amounts of time to become stable depending on the particular processes involved in the task used (Kassardjian et al., 2005; Krakauer & Shadmehr, 2006; Shadmehr & Holcomb, 1997).
Concerning the interaction of S-C and S-A associations, as far as we know, there are no studies to-date that have investigated possible binding of such associative memories with long-term consolidation. One possibility is that information, which co-occur over a long period of time, become bound in an effort to support efficient processing. This theory is supported by one study, which suggested that declarative and procedural memory systems may interact during consolidation (Brown & Robertson, 2007). Further research is necessary to determine whether this is indeed the case for associative memories such as those studied here. The effect of general context engagement on consolidation has not been studied either, as far as we know. Overall, although the current study did not investigate consolidation processes in a controlled way as we did not manipulate sleep conditions, it does open up new interesting questions in memory research for the investigation of the consolidation and reconsolidation of associative priming effects as well as the effect of context engagement on consolidation of S-C and S-A associations.
To summarise, we investigated the longevity of S-R associations at three temporal intervals: a few seconds, 1 day, and 1 week after encoding. We found that S-R associations show effects of retrieval even 1 week after priming. We also show that, as previous research suggests, there are independent associations between stimuli and task-specific classifications, and between stimuli and motor codes, and these associations show different patterns of retrieval across long-term memory intervals. Finally, we observed that differences in processing time during encoding are linked to the size of priming effects occurring during retrieval of S-C associations. Overall, our results suggest that further elaborations are necessary on priming memory, the processes that compose it, and what the individual memory characteristics of S-R components are in terms of longevity, durability, and susceptibility to top-down processes.
Footnotes
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
This research was supported by the Agence Nationale de la Recherche (SRA ANR-13-FRAL-0007-01, K.M. and F.W.) and by the Deutsche Forschungsgemeinschaft (KI1388/5-1, A.K.).