
Abstract
Using a human Pavlovian-to-instrumental transfer (PIT) task, Alarcón and Bonardi showed that the selective elevation of instrumental responding produced by excitatory transfer cues was reduced when these cues were presented with a conditioned inhibitor (CI), relative to a control cue that was simply preexposed. However, previous research has shown that preexposed cues might also acquire inhibitor-like properties. This study aimed to contrast the inhibitory properties of CIs and preexposed cues, using novel stimuli as controls, in summation and PIT tests. Participants were trained to perform two actions, each reinforced with a distinct outcome (O1 or O2). Two images were trained as CIs, each signalling the absence of one of the outcomes, by presenting them with a cue that was otherwise followed by that outcome (e.g., A→O1, AI→no O1). In contrast, the preexposed cues were simply presented in the absence of the outcomes. In the summation test, participants rated the likelihood of the outcomes in the presence of two independently trained excitatory cues, each presented with a CI, a preexposed cue, or a novel stimulus. Similarly, in the PIT test, participants performed both actions in the presence and absence of these compounds. In the summation test, the CIs and the preexposed cues reduced participants’ expectations of the outcomes more than the novel stimuli. However, in the PIT test, only the CIs reduced the selective elevation of responding produced by the transfer cues. These results might reflect distinct properties of stimuli trained as CIs and those simply preexposed.
Keywords
A conditioned inhibitor (CI) signals the absence of a significant event. For example, if a conditioned stimulus (CS) A is always paired with an unconditioned stimulus (US) except when accompanied by another cue I, that is, A→US, AI→no US, this unexpected omission of the US drives I to acquire inhibitory properties opposite to those of A. These are often evaluated in summation or retardation tests (Rescorla, 1969). In the summation test, the CI is presented in compound with a separately trained signal for the US. Inhibition is evident if the CI suppresses this cue’s ability to predict the US more than a control stimulus. In the retardation test, the inhibitor and a suitable control stimulus are both separately paired with a new US and the rate of acquisition is compared; learning an excitatory relationship should be slower if the stimulus is an inhibitor.
Alarcón and Bonardi (2016) recently demonstrated conditioned inhibition in a different manner, using a human Pavlovian-to-instrumental transfer (PIT) task (for reviews on PIT, see Cartoni, Balleine, & Baldassarre, 2016; Holmes, Marchand, & Coutureau, 2010). Participants were trained to perform two responses (R1, R2), each reinforced by a distinct outcome (pictures of food or drinks O1, O2; that is, R1→O1; R2→O2). Two neutral images that were later used as transfer cues, T1 and T2, were paired with the same outcomes (i.e., T1→O1; T2→O2), and in a subsequent PIT test participants performed R1 and R2 while T1 and T2 were presented with one of two neutral control stimuli, C1 and C2 (T1C1/T2C2). Participants performed R1 more than R2 during T1C1, but the reverse during T2C2—a selective elevation of instrumental responding known as outcome-specific PIT. It has been argued that PIT results from the associations between both R1 and T1 with O1: presentation of T1 activates the O1 representation, which elicits performance of R1 (cf. Trapold & Overmier, 1972). This contrasts with the general form of PIT, in which a CS invigorates instrumental responding regardless of whether the CS and the instrumental responses are trained with the same or different outcomes, as long as they are both of the same motivational valence (cf. Rescorla & Solomon, 1967).
In addition, Alarcón and Bonardi (2016) paired a stimulus S1 with O1 unless accompanied by an inhibitor l1 (S1→O1; S1l1→no O), making l1 an inhibitor for O1. In the PIT test, the selective elevation of responding during T1C1 and T2C2 was eliminated when T1 and T2 were presented with inhibitor l1 (T1l1/T2l1). Pavlovian inhibitors are commonly thought to act by suppressing activation of the outcome representation, and so the authors attributed this effect to l1’s inhibitory properties interfering with the T1→O1→R1 chain that mediates PIT. This conclusion was supported by the results of a summation test (cf. Rescorla, 1969), in which participants’ expectation of O1 and O2 during T1 and T2, respectively, was significantly weaker during T1l1 and T2l1 than during T1C1 and T2C2.
In that study, the control stimuli C1 and C2 were preexposed in compound in the absence of any outcome, meaning they were treated identically to I except for never being presented with an excitatory CS—so they should not have become inhibitory. However, some have argued that, because the context in which the cue is presented also elicits expectancy of the US, this is sufficient for the stimulus to develop what is termed differential inhibition (see Miller, Hallam, Hong, & Dufore, 1991). For instance, Karazinov and Boakes (2004) reported that a CI and a preexposed stimulus both reduced expectation of the US relative to a novel cue—although the CI did this more effectively than the preexposed cue. However, in further experiments, the authors tried to enhance the “inhibitory” properties of the preexposed cue by increasing the excitatory strength of the context during training. Despite these manipulations, the degree to which the preexposed cues reduced participants’ expectations of the US did not change—not what would be expected if differential inhibition were responsible. Nonetheless, these findings raise the possibility that the control cues in our PIT study might also appear inhibitory relative to a novel control stimulus. This study explored this possibility.
Alarcón and Bonardi (2016) also found that the effect of the CI was not outcome-specific: although I only signalled the absence of O1, in the summation test, it reduced expectation of O2 during T2 as effectively as expectation of O1 during T1 and also suppressed the PIT effect produced by T1 and T2 to a similar extent. This suggests I predicted the absence of both outcomes, even though it had only been trained as a signal for omission of O1. Although some previous studies have reported evidence consistent with these results (e.g., LoLordo, 1967; Nieto, 1984; Pearce, Montgomery, & Dickinson, 1981), there are also several studies showing that an inhibitor is more effective in reducing responses to a cue that signals the same outcome as that inhibited by the inhibitor (e.g., Delamater, Sosa, & LoLordo, 2003). Although the factors determining whether the inhibitors will act in a general or an outcome-specific manner is unclear, in the case of Alarcón and Bonardi (2016) it may have been because only one inhibitor was trained; perhaps if an inhibitor had also been trained for O2, the inhibitors would act more selectively. This possibility was also explored.
In this study, we used an outcome-specific PIT design to assess the effect of two inhibitors, each signalling the absence of a distinct outcome, in summation and PIT tests. Initially, participants received instrumental training in which R1 and R2 produced outcomes O1 and O2 (Table 1). After this they were trained that images S1 and S2 (in compound with filler cues A and B) produced O1 and O2 unless accompanied by l1 and l2; thus, l1 and l2 predicted the absence of O1 and O2, respectively (S1A→O1, S1I1-, S2B→O2, S2I2-). Two transfer stimuli, T1 and T2, were also paired with O1 and O2, and the critical control cues C1 and C2 were presented in compound without reinforcement, the same number of times as l1 and l2; l1, l2, C1, and C2 were also presented in isolation.
Experimental design.
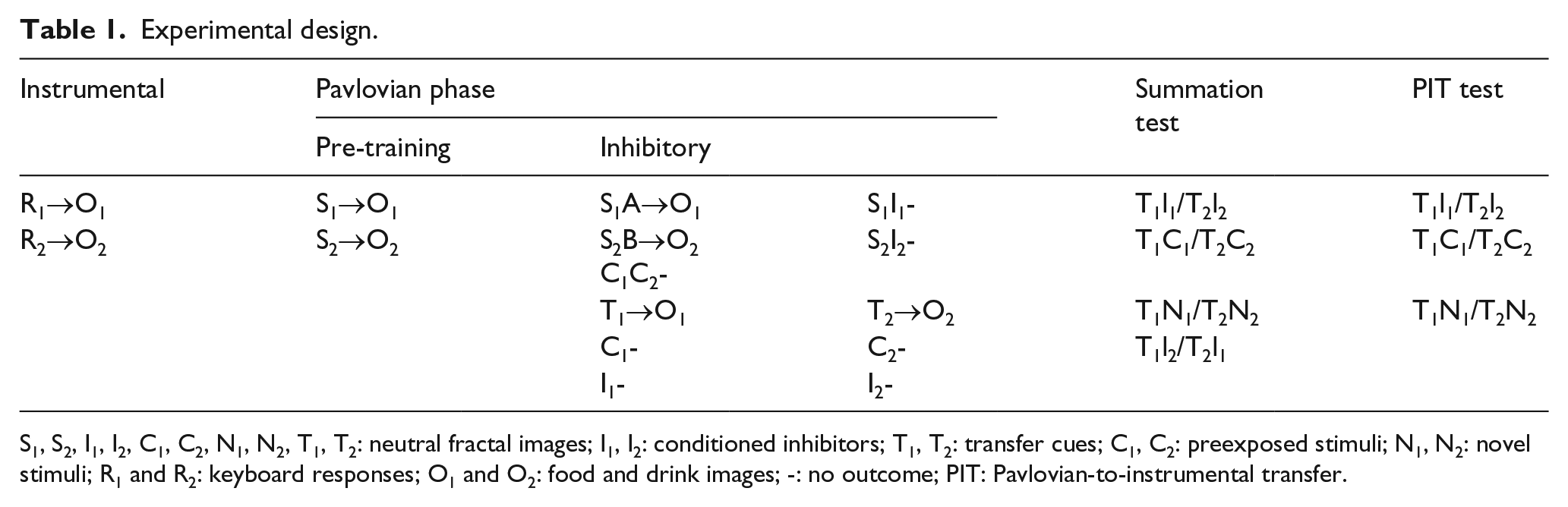
S1, S2, I1, I2, C1, C2, N1, N2, T1, T2: neutral fractal images; I1, I2: conditioned inhibitors; T1, T2: transfer cues; C1, C2: preexposed stimuli; N1, N2: novel stimuli; R1 and R2: keyboard responses; O1 and O2: food and drink images; -: no outcome; PIT: Pavlovian-to-instrumental transfer.
In the subsequent summation test, we measured participants’ expectations of the outcomes by presenting CSs signalling those outcomes together with the inhibitors or with control stimuli that were either novel or preexposed during training. Thus, T1 and T2 were presented with the inhibitors for the outcomes they predicted (T1l1 and T2l2, respectively), the preexposed cues C1 and C2 (T1C1 and T2C2), and two novel cues N1 and N2 (T1N1 and T2N2). This novel control condition allowed us to evaluate the extent to which the preexposed cues C1 and C2 had become differential inhibitors and acquired inhibitory properties similar to those shown by l1 and l2. We expected a greater reduction of participants’ expectations to be produced by the inhibitors than the novel control cue; the question was whether the preexposed control cue would also demonstrate inhibitory properties, reducing expectation of the outcome more than the novel stimulus. In addition, T1 and T2 were also presented with inhibitors for the alternative outcomes (T1l2 and T2l1), to establish whether training inhibitors for both outcomes would render them outcome-specific. If inhibition is outcome-specific, lower expectation of the outcomes should be evident when the CS and the inhibitor were both trained with the same outcome.
In the PIT test which followed, we compared the effect of the inhibitors and preexposed control cues on outcome-specific PIT, by presenting the CSs in compound with the CIs, the preexposed stimuli, or the novel control cues (T1l1 and T2l2, T1C1 and T2C2, and T1N1 and T2N2). We predicted a reduction of the selective elevation of instrumental performance produced by the CSs, that is, outcome-specific PIT, when these cues were presented with the inhibitors relative to the novel control cues; the question was again whether the preexposed control cues also displayed inhibitory properties, reducing outcome-specific PIT more than the novel stimuli.
Method
Participants
The participants were 27 students from the University of Nottingham, aged 18–28 (4 males, 23 females). One participant was excluded for only performing one of the two responses in the instrumental phase. Students from the School of Psychology received course credit and the rest received an inconvenience allowance of £4. The number of participants was chosen based on the number of counterbalancing conditions and on previous studies conducted by the authors (Alarcón & Bonardi, 2016).
Apparatus and materials
The task was programmed in PsychoPy (Peirce, 2007) on a standard computer with a 20-inch screen. Twelve neutral fractal images were used as CSs, and images of foods and drinks (4 of each) as outcomes. All images were 8 cm × 8 cm (see Alarcón, Bonardi, & Delamater, 2018). The neutral images were presented immediately to the left or right of the screen centre, and the outcomes were presented centrally. The components of each neutral stimulus compound were presented an equal number of times on the left and right. A fixation cross (10 mm × 10 mm) was located at the centre of the screen in the instrumental phase and PIT test and was replaced by a fixation dot (3 mm × 3 mm) in the Pavlovian phase. R1 and R2 were pressing the keys “z” and “m.” For half of the participants, “z” was reinforced with food and “m” with drink, and the reverse for the remaining participants. In the summation test, participants used the mouse to click on the rating scale, and the “space” bar to advance through the phases. Instructions were presented on the screen before each phase.
Procedure
Participants received a brief explanation of the task and then completed a consent form before they were guided into a quiet room.
Instrumental training
Participants were instructed to learn the relationships between pressing the “z” and “m” keys and different rewards (images of foods or drinks) and to obtain as many rewards as they could. Each response was reinforced according to a variable ratio (VR) 5 schedule, such that an outcome was presented after an average of five responses. Each outcome was presented in the centre of the screen, either until 0.8 s had elapsed or until another key was pressed, at which point it was replaced by the fixation cross. This phase ended when participants had received 100 outcomes in total.
Pavlovian phase
Participants were informed that different images would appear on the screen, some of them followed by the rewards and others not, and that they would have to answer a series of questions about these relationships by pressing numbers on the keyboard. In each trial, the text “Which reward will appear now?” was presented at the top of the screen, one CS image (or compound) below it, and the text “1) Food 5) Drink 9) Nothing” at the bottom. When participants pressed one of the numbers on the keyboard (1, 5, or 9), these stimuli were removed from the screen and the correct outcome was presented; in addition, depending on the participant’s response, the text “Correct!” in green font, or “Oops! That was wrong” in red font, was presented at the top or bottom of the screen, respectively. The intertrial interval (ITI) was 2 s, in which only the fixation dot was present.
The initial, pre-training stage was divided into two blocks, each comprising four trials of S1→O1 and four trials of S2→O2, to establish S1 and S2 as signals for their respective outcomes. The inhibitory stage that followed was divided into two blocks, each comprising four trials of S1A→O1 and S2B→O2, six trials each of T1→O1, T2→O2, S1I1-, S2I2-, and C1C2-, and three trials of I1-, I2-, C1, and C2 (Table 1). The filler cues A and B were added to S1 and S2 on reinforced trials to ensure that participants could not simply learn that when two cues were presented no outcome followed, without needing to pay attention to the individual cues in each compound. The order of the trial types was semi-random within each block. Participants could, if they wished, take a break between these blocks.
Summation test
Participants were informed that images would appear on the screen, after which they must rate the likelihood of an outcome’s appearance using a rating scale. On each trial the text “In a scale from 1 to 100, how likely is it that this image will be followed by” together with the word “FOOD” or “DRINK” was presented at the top of the screen. For half of the trials, the questions ended with the word “FOOD” and the compounds consisted of the transfer cue signalling food with either the inhibitor trained with food, for example, T1I1, the inhibitor trained with drinks, for example, T1I2, one of the preexposed control cues, for example, T1C1, or one of the novel cues, T1N1. For the other half, the question ended with the word “DRINK,” and the stimuli presented were corresponding compounds containing the transfer cue signalling drink, for example, T2I2, T2I1, T2C2, and T2N2. The compounds were presented, and below them a rating scale with “0” on the left and “100” on the right. The test consisted of two blocks, each comprising two trials of each compound (Table 1); the order of questions was randomised within each block.
Instrumental retraining
This was designed to serve as a reminder of the specific R-O associations learned at the beginning of the experiment, and was identical to the initial instrumental phase, except participants earned only 50 outcomes in total.
PIT test
Participants were instructed to press either “z” or “m” as much as they wanted in order to obtain the rewards, but no outcomes were presented. In each trial, the fixation dot was present for 2 s (pre-CS baseline period), followed by a 2-s CS presentation (CS period). The test was divided into three blocks, each comprising two presentations of each stimulus compound, presented in a semi-random order.
Data treatment
The data were analysed using analysis of variance (ANOVA). Significant two-way interactions were explored with simple main effects analysis using the pooled error term. Partial eta-squared
Results
The Pavlovian and instrumental phases were completed uneventfully. The mean number of R1 responses and O1 presentations were 269.9 (SEM = 14.4) and 51.8 (SEM = 2.9), respectively, and for R2 and O2 251.2 (SEM = 13.4) and 54.4 (SEM = 2.9), respectively. The mean proportion of correct responses in the inhibitory training (0 = no correct answers, 1 = all correct answers), for blocks 1 and 2, respectively, were 0.58–0.58 (SEM = 0.05–0.06) for S1A/S2B; 0.55–0.76 (SEM = 0.04–0.05) for S1I1/S2I2; 0.82–0.92 (SEM = 0.05–0.04) for C1C2; 0.63–0.87 (SEM = 0.03–0.03) for T1/T2; 0.69–0.87 (SEM = 0.05–0.05) for I1/I2; and 0.65–0.9 (SEM = 0.05–0.05) for C1/C2. An ANOVA with Block (1, 2) and Cue (S1A/S2B, S1I1/S2I2, C1C2, T1/T2, I1/I2, C1/C2) showed a significant main effect of Block, F(1, 25) = 61.18, MSe = .03, p < .001, significant main effect of Cue, F(5, 125) = 10.06, MSe = .05, p < .001, and a significant Block × Cue interaction, F(5, 125) = 4.73, MSe = .024, p = .001. The analysis of the interaction confirmed more correct responses in Block 2 relative to Block 1 for compounds S1I1/S2I2 (p < .001), C1C2 (p = .001), T1/T2 (p < .001), I1/I2 (p < .001), C1/C2 (p < .001), but not for S1A/S2B (p = .86). In view of the fact that performance on S1A/S2B trials did not increase over blocks, we further explored the incorrect responses to these compounds and found that in 83% of the cases participants chose the option “nothing” (rather than the alternative outcome). This was likely caused by S1 and S2 also being paired with the absence of the outcomes when presented with the inhibitors, rendering them potentially ambiguous cues.
The rating scores in the summation test to T1 and T2, according to whether they were paired with the corresponding CI (T1l1 and T2l2), the preexposed control cue (T1C1 and T2C2), the novel control cue (T1N1 and T2N2), or the alternative inhibitor (T1l2 and T2l1), are plotted in Figure 1. An ANOVA with Outcome (O1, O2) and Cue (Inhibitor, Preexposed, Novel, Alternative) showed a significant main effect of Cue, F(3, 75) = 10.9, MSe = 607.6, p < .001,
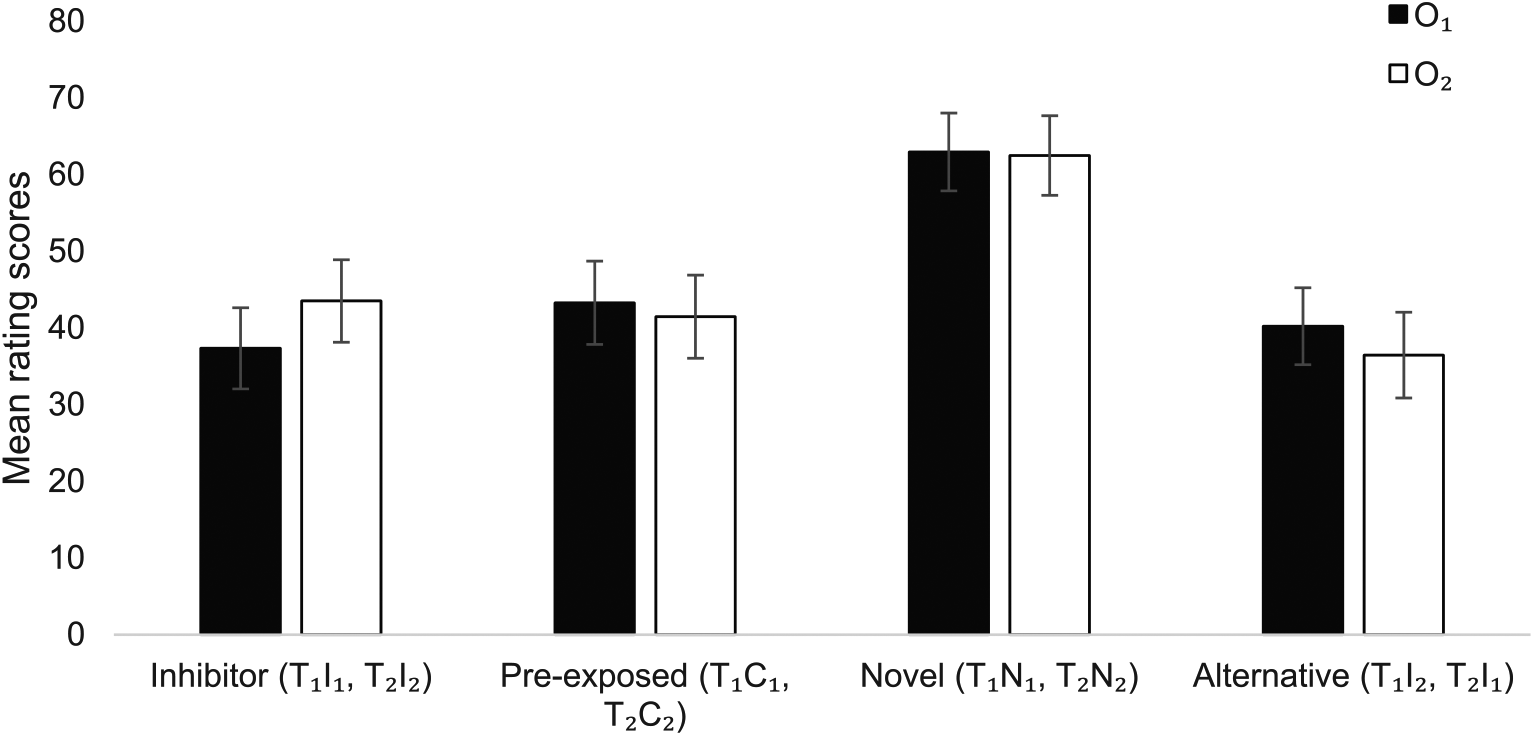
Mean rating scores (participants’ expectancies) ± SEM to the questions about O1 and O2 for the transfer cues in compound with the inhibitors (T1l1, T2l2), the preexposed control cues (T1C1, T2C2), the novel control cues (T1N1, T2N2), and the alternative inhibitors (T1l2, T2l1) in the summation test.
The results of the PIT test are presented in Figure 2, which shows that, as expected, the transfer cues (T1 and T2) elicited more Same than Different responses when presented with the novel control stimuli (T1N1 and T2N2). This outcome-specific PIT effect appeared equally substantial in the preexposed stimulus compounds T1C1 and T2C2, but was reduced with the inhibitory compounds (T1I1 and T2I2). ANOVA with Outcome (Same, Different) and Compound (Preexposed, Inhibitory, Novel) as factors showed a significant main effect of Outcome, F(1, 25) = 9.7, MSe = 15,389.1, p < .01,
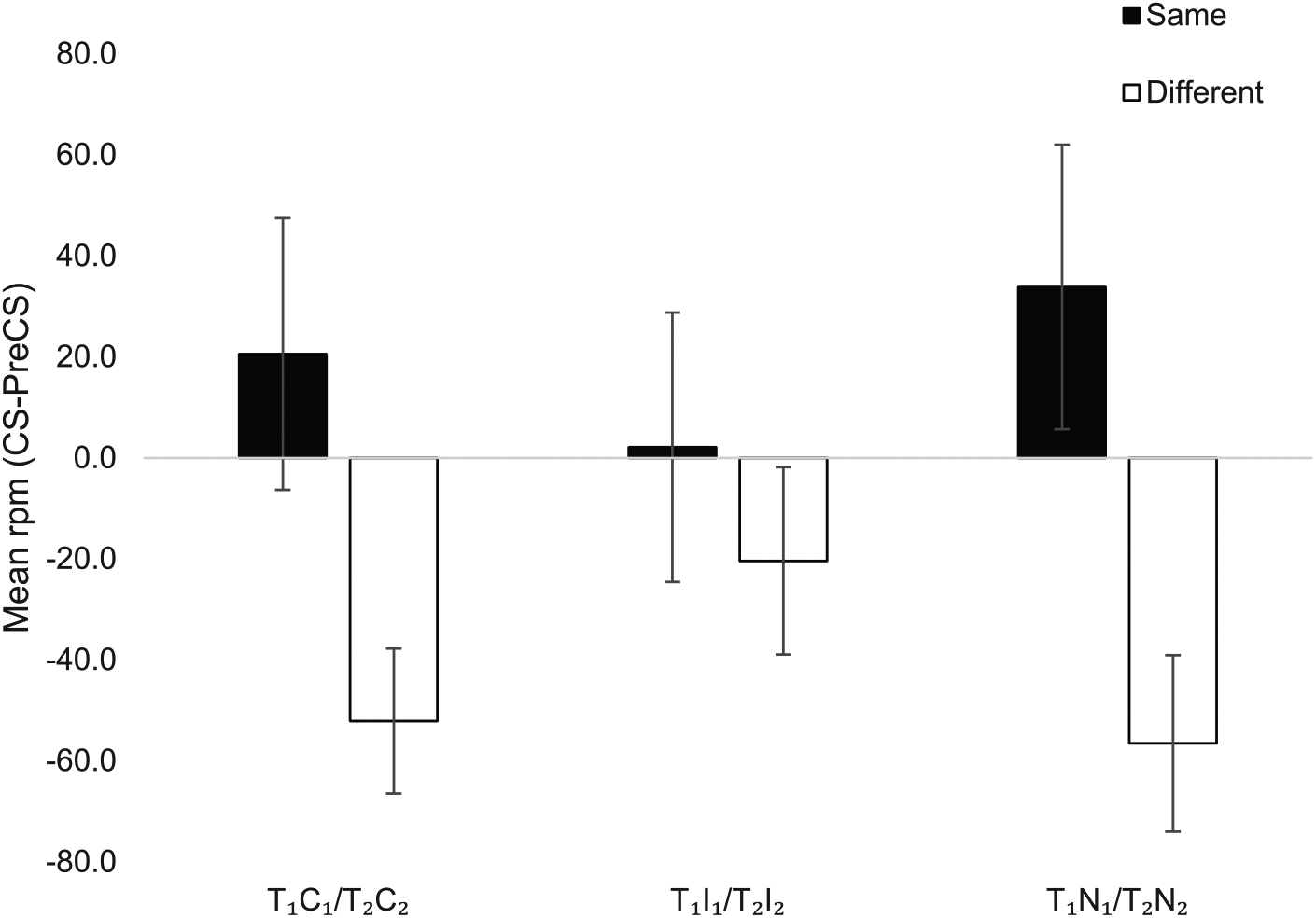
Same and different responding to the transfer cues in compound with the preexposed control stimuli (T1C1/T2C2), the inhibitors (T1I1/T2I2), and novel control cues (T1N1/T2N2) in the PIT test.
Discussion
This experiment aimed to establish whether, in our procedure, preexposed stimuli would behave like CIs in summation and PIT tests. When compounded with the novel control stimuli N1 and N2, the transfer cues T1 and T2 elicited expectation of their respective outcomes; moreover, this effect was significantly reduced when they were compounded with the inhibitors l1 and l2, and also the preexposed control stimuli C1 and C2. This indicates that preexposure of C1 and C2 in training had made them as inhibitory as “true” Pavlovian inhibitors. Karazinov and Boakes (2004) also found that preexposed cues were more inhibitory than novel cues in a summation test, but—in contrast to our findings—less so than Pavlovian inhibitors, a difference we also observed in previous work (Alarcón & Bonardi, 2016). Karazinov and Boakes speculated that the difference between the Pavlovian and differential inhibitors stemmed from differences in how they were trained. While the Pavlovian CI was trained both as a single cue and in a compound, the preexposed differential CI was only trained alone. Seeing the preexposed cue in a compound for the first time at test might have produced uncertainty in the participants, making their ratings to this compound more conservative than to the other cues (the ratings to this compound were closer to the centre of a rating scale marked as “Don’t know”). However, in both Alarcón and Bonardi (2016) and the experiment reported here the Pavlovian CIs and the preexposed stimuli were presented the same number of times as single cues and compounds across the task, making it unlikely this factor was responsible. One potentially critical difference between our two studies is that Alarcón and Bonardi (2016)—which was primarily aimed to explore the effect of Pavlovian inhibition on PIT—excluded participants who had not learned the Pavlovian inhibitory discrimination in training. It is possible that considering only the participants that showed inhibition in the summation test might have increased the differences between the preexposed control and the inhibitor in the PIT test; however, we reanalysed the data of the present experiment using the same response criterion as Alarcón and Bonardi (2016), but still found no significant differences between these cues.
In summary, although the summation technique indicates that preexposure produces conditioned inhibition, it is unclear whether or not this is as strong as that produced by Pavlovian training. Moreover—in view of the failure of Karazinov and Boakes to manipulate the degree of differential inhibition as theory would predict—it is also worth considering alternative explanations. For example, perhaps the preexposed cue is becoming associated with the idea that “nothing” is going to happen, and expectation of this neutral event interferes with expectation of the outcome, reducing response ratings. This could have a similar effect in the summation test as the active expectation of outcome omission produced by the Pavlovian inhibitor (cf. Alarcón & Bonardi, 2016).
Consistent with this suggestion, although the results of the summation test suggested the preexposed cues acquired inhibitory properties, the PIT test did not. Here, the selective elevation of instrumental responding produced by T1 and T2—outcome-specific PIT—was eliminated only by the CIs, and by neither the preexposed or novel cues, which did not differ (cf. Alarcón & Bonardi, 2016). These results suggest that—in contrast to the results of the summation test—the preexposed cues were not inhibitory. This discrepancy between summation and PIT tests may be due to their differential sensitivity in detecting, perhaps subtle, differences in inhibition. Moreover, our sample size may not have been large enough to detect small differences between the inhibitors and the preexposed controls in the summation test, and between the preexposed controls and the novel cues in the PIT test. For instance, a larger sample size might have revealed that the preexposed controls were inhibitory relative to the novel cues, but less so than the Pavlovian inhibitors, that is, inhibitors < preexposed < novel. Alternatively, it may be that the nature of the inhibitory properties endowed by Pavlovian inhibition training and nonreinforced preexposure is qualitatively different and thus manifests differently depending on how it is tested. This would suggest that simple preexposure does not result in true inhibition, summation tests are sensitive to factors other than pure Pavlovian inhibition, and the PIT test is a purer measure of this property. These alternative explanations make it necessary to continue studying the effect of conditioned inhibition on PIT.
Another feature of Alarcón and Bonardi’s (2016) results was that the inhibitor for O1 reduced summation and PIT effects just as much for T2 as for T1, suggesting that its inhibition was not outcome-specific. We suggested that this might be because only one CI-no US relationship was trained, and so in this study we trained two inhibitors, one for each of the outcomes. However, this had no effect on the result: in the summation test, there was no difference in expectation of the outcomes during the inhibitory compounds, regardless of whether the inhibitor predicted omission of the outcome predicted by the excitor with which it was paired, or the alternative outcome. In fact, there is already some controversy over whether CIs are specific to the sensory properties of the outcomes whose omission they signal, or whether they act via a solely motivational mechanism. Initial studies suggested that CIs trained with a specific US, such as a shock to the left eye, would inhibit responding equally to any CS signalling shock, regardless of the eye to which it was delivered (Nieto, 1984; Pearce et al., 1981; see LoLordo & Fairless, 1985, for a review). In contrast, more recent research, using PIT measures of inhibition, suggests a different story. Quail, Laurent, and Balleine (2017; see also Laurent, Wong, & Balleine, 2015) used a human task to train two inhibitors, one for each of two outcomes (S1→O1, S2→O2, S1 l1→nothing, S2 l2→nothing). The effects of l1 and l2 were evaluated in a PIT test, both when presented alone, when paired with the CSs with which they were trained (S1l1, S2l2)—and with the alternative CSs (S1l2, S2l1). If l1, for example, were a specific inhibitor for O1, it should reduce the PIT effect produced by S1—which predicts the precise outcome whose omission l1 signalled—but not that produced by S2, which predicted the alternative outcome. This is what the authors observed.
Unfortunately, there were many differences between this study that demonstrated outcome-specific inhibition and our experiments which did not. One is that we evaluated learning in each trial of the Pavlovian conditioning phase by asking participants to press a key to predict the outcome to be delivered. Because of this, participants had to choose the option “Nothing” for the CIs and the control cues. It is possible that this consequence, being common to both inhibitors, hindered the development of outcome-specific inhibition. Quail et al. (2017), in contrast, evaluated learning at the end of the Pavlovian conditioning phase, and participants could answer if the inhibitor was associated with the absence of a particular outcome, for example, I1 followed by no food, or with the general absence of the outcomes, for example, I1 followed by no foods or drinks, which might have encouraged a discrimination between the CIs. In addition, Quail et al. separated their participants based on these answers (Experiment 2), finding that the “specific learners,” who identified specific inhibitor→no outcome relationships, showed high levels of outcome-specificity in the PIT test, whereas the “general learners,” those who identified the inhibitors as signalling the absence of any outcome, did not show any signs of such specificity. It is possible that we might show a similar effect by separating the participants similarly; however, our measures of Pavlovian learning do not allow us to divide our participants into specific or general learners.
Another difference is that Quail et al. did not evaluate their inhibitors’ properties against separately trained test excitors, and only used a PIT test (Rescorla, 1969). Conversely, our first experiment that demonstrated non-outcome-specific inhibition in the PIT test only trained one inhibitor, while in the present experiment we did not evaluate the alternative compounds in the PIT test. Our evidence for non-outcome-specific inhibition came only from a summation test, which Quail et al. did not include. Thus, there is no real conflict of results here—but a key need to bridge the gap between these various studies in order to establish whether or not inhibition is outcome-specific in these kinds of tasks.
Another point worth mentioning is the use of a brief instrumental retraining before the PIT test, the purpose of which was to serve as a reminder of the instrumental associations learned at the beginning of the task (see Alarcón & Delamater, 2019, for the same procedure in rodent research). It might be argued that this is similar to an “overtraining” procedure, resulting in habitual responding rather than goal-directed behaviour, which is less dependent on the outcome representation. However, we do not consider this training to be sufficient to produce habits. There is one example in which habits have been achieved by overtraining instrumental responses in humans, but it required several separate sessions of instrumental conditioning (Tricomi, Balleine, & O’Doherty, 2009), and recently a series of failed attempts to reproduce those findings have been published (de Wit et al., 2018). Related to overtraining, Garofalo and Robbins (2017) provided evidence that extended training in humans enhances the outcome-specific PIT effect and it would be interesting to assess if the ability of the CIs to reduce selective PIT remains after instrumental overtraining.
A final consideration is that, although it was designed to be similar to the tasks used in animal research, we did not obtain a direct measure of Pavlovian conditioning but rather asked participants about explicit expectations of the outcomes, as in contingency learning tasks (e.g., Shanks, 2007). Some researchers have argued that the processes involved in this type of task depend on the generation of propositions about the relationships between the events, rather than the traditional associative view, and there have been recent efforts to explain outcome-specific PIT with a propositional account (e.g., Hogarth et al., 2014; Seabrooke, Hogarth, & Mitchell, 2016). However, the outcome-specific PIT effect is consistently found in both animal and human studies, suggesting that the measure of Pavlovian conditioning might not be critical. For instance, the inhibition of PIT results found by Quail et al. (2017) was remarkably similar to previous results found in rodents by the same group of researchers (Laurent et al., 2015). Nevertheless, adding direct measures of Pavlovian conditioning might provide further information about the underlying processes in this type of task.
In summary, we have demonstrated that simple preexposure can make a stimulus behave like a CI in a summation test, relative to a novel control; however, this was not evident in a PIT test of inhibition. Of several possible interpretations, one possibility is that summation tests are sensitive to effects other than Pavlovian inhibition and that the PIT test is a better measure of a stimulus’ inhibitory properties. We also failed to produce evidence of outcome-specific inhibition in a summation test, despite training two inhibitors, one for each outcome. Although this ostensibly contradicts findings from other studies (e.g., Quail et al., 2017), a more likely explanation is that this discrepancy arises from procedural differences. More generally, we believe that these findings expand our understanding of the mechanisms underlying PIT and that this task might be helpful to study the effect of conditioned inhibition in maladaptive behaviour, such as drug-seeking behaviours and overeating.
Footnotes
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: The elaboration of this manuscript was supported by a postdoctoral grant awarded by Conicyt (Fondecyt # 3170166) to D.E.A.