
Abstract
Traditional associative learning theories predict that training with feature negative (A+/AB-) contingencies leads to the feature B acquiring negative associative strength and becoming a conditioned inhibitor (i.e., prevention learning). However, feature negative training can sometimes result in negative occasion setting, where B modulates the effect of A. Other studies suggest that participants learn about configurations of cues rather than their individual elements. In this study, we administered simultaneous feature negative training to participants in an allergist causal learning task and tested whether evidence for these three types of learning (prevention, modulation, configural) could be captured via self-report in the absence of any procedural manipulation. Across two experiments, we show that only a small subset of participants endorse the prevention option, suggesting that traditional associative models that predict conditioned inhibition do not completely capture how humans learn about negative contingencies. We also show that the degree of transfer in a summation test corresponds to the implied causal structure underlying conditioned inhibition, occasion-setting, and configural learning, and that participants are only partially sensitive to explicit hints about causal structure. We conclude that feature negative training is an ambiguous causal scenario that reveals individual differences in the representation of inhibitory associations, potentially explaining the modest group-level inhibitory effects often found in humans.
Keywords
The ability to detect contingent relationships in the world is critical to make accurate predictions about the future. The contingency between a given cue and an outcome may be positive (the cue is associated with the outcome), or it may be negative (the cue is associated with the absence of the outcome). Although it is clearly important to learn about positive (causal) relationships, it is just as important to learn about negative (preventive) ones. Theoretical accounts of learning often characterise prevention learning as arising from the detection of negative associations between cues and outcomes (e.g., Cheng, 1997; Cheng & Novick, 1992; Rescorla & Wagner, 1972; Ward & Jenkins, 1965). Thus, at first glance, prevention learning can be seen as simply the opposite to causal learning.
This view of causation and prevention as diametrically opposed is formalised in traditional associative learning models such as the Rescorla-Wagner model (Rescorla & Wagner, 1972). These models, which were originally developed to explain Pavlovian conditioning in animals, have been cited as theoretical accounts of how humans learn causal and preventive relationships between cues and outcomes (Dickinson et al., 1984). The Rescorla-Wagner model conceptualises predictive relationships between cues and outcomes in terms of a single dimension of associative strength. In associative terms, “excitors” (causal cues) have a positive associative strength and activate the representation of an outcome, while “inhibitors” (preventive cues) have negative associative strength and inhibit the representation of the outcome. In such models, associative strength changes on each trial in proportion to the degree and direction of prediction error generated by the cues present on that trial. Negative prediction error (expectation of an outcome that does not occur) results in a decrease in associative strength for any cues present, and may result in a cue acquiring net negative associative strength (i.e., becoming an inhibitor).
A procedure that generates negative prediction error is feature negative training (A+/AB-, also known as conditioned inhibition training). First demonstrated by Pavlov (1927), this procedure consists of pairing a target (A) with an outcome (+), as well as pairing a compound of the target and feature (AB) with no outcome (-). An intuitive interpretation of these contingencies is that since the target (A) predicts the outcome, the addition of the feature (B) must therefore prevent the outcome from occurring. This intuition is captured in the Rescorla-Wagner model’s calculation of prediction error, which leads to the feature accruing negative associative strength and becoming a conditioned inhibitor.
A summation test is typically used to show that the feature B has acquired inhibitory properties (Rescorla, 1969). This test consists of comparing conditioned responding to a compound consisting of the feature (B) and a separately trained target (e.g., C+) to a control compound where the same target (C) is combined with a neutral control cue (e.g., D, a cue paired with no outcome). If responding is lower to the test compound (CB) than to the control compound (CD), then the feature (B) is deemed inhibitory. Conditioned inhibition has been reliably demonstrated using the summation test in a variety of conditioning preparations and animal species (see Savastano et al., 1999; Sosa & Ramírez, 2019; Swartzentruber, 1995 for reviews). Thus, there is good evidence in animals for the idea that feature negative training produces inhibitory learning about the feature (i.e., prevention learning) and that inhibitors control behaviour opposite to excitation (Rescorla, 1969). Note that henceforth, we will refer to the degree to which a negative feature suppresses responding to an excitor in a summation test as transfer of inhibitory learning (i.e., an empirical phenomenon), reserving the term conditioned inhibition for “true” preventive learning where the negative feature passes the summation test and possesses the properties described by Rescorla (1969).
However in humans, evidence for transfer of inhibitory learning at the group level is much weaker (e.g., Karazinov & Boakes, 2007; Lee & Livesey, 2012; Williams, 1995; Wilkinson et al., 1989). This suggests that inhibitory learning in humans may be more complex than predicted by associative models that posit a single dimension of associative strength (Rescorla & Wagner, 1972). In particular, there might be individual differences in what participants learn about the negative feature, with only a subset of participants learning in the manner predicted by the Rescorla-Wagner model. Glautier and Brudan (2019) have recently provided evidence in support of this idea. They presented participants with feature negative training where a context served as the negative feature, and found that participants varied in the degree to which they showed transfer of inhibitory learning in a summation test. Critically, classifying participants into “inhibitors” and “non-inhibitors” predicted how participants responded to conditioning of a different negative feature in a new task. There is growing recognition of the existence of meaningful individual differences in associative learning in humans (e.g., Byrom & Murphy, 2014, 2016; Lee et al., 2018; Stegmann et al., 2019) and even in animals (Jean-Richard-dit-Bressel et al., 2019). In cases where the individual differences are qualitative, aggregate data may be misleading (e.g., see Lee et al., 2018). Glautier and Brudan’s (2019) study provides initial evidence that there might be stable individual differences in feature negative learning.
Individual differences are especially likely to arise in feature negative designs because there are several alternative ways to solve the training task. Feature negative training can sometimes lead to negative occasion setting (see Bonardi et al., 2017; Fraser & Holland, 2019, for reviews). In contrast to learning that the feature B prevents or inhibits the outcome itself, negative occasion setting involves learning that the feature B modulates (sets the occasion for) the target’s (A’s) relationship with the outcome. In other words, A by itself causes the outcome but when B is also present, A does not cause the outcome. A popular view of occasion setting is that it constitutes evidence of hierarchical control of associative behaviour, where the occasion setter is associated not with the outcome but with an association between another cue and the outcome (Bonardi et al., 2017; Fraser & Holland, 2019; see Konorski, 1948; Rescorla, 1969 for an alternative threshold view). Indeed, Glautier and Brudan (2019) attributed the difference between their inhibitor and non-inhibitor subgroups to a tendency to learn first-order (inhibition) or second-order (occasion-setting) associations. In contrast to conditioned inhibition, the defining feature of occasion-setters is that they possess second-order (modulatory) links that are independent of any direct associations with the outcome (e.g., Holland, 1984, 1989).
Interestingly, negative occasion setting is typically found with serial (i.e., B → A → no outcome) presentation of the AB compound, while conditioned inhibition is typically found with simultaneous presentation (AB → no outcome) in animals (e.g., Holland, 1984, 1989; Holland & Lamarre, 1984; Holland & Morell, 1996) and humans (Baeyens et al., 2004). Negative occasion setting (from serial procedures) is distinguished from conditioned inhibition (in simultaneous procedures) by multiple functional properties. One of the key features of occasion setters is that they show limited transfer in a summation test. They tend not to transfer (pass the summation test) when presented serially with other targets, but may sometimes transfer to specific targets that have themselves been part of an occasion-setting discrimination (see Holland, 1989; Lamarre & Holland, 1985; Wilson & Pearce, 1990). Although the bulk of the occasion-setting literature comes from studies with non-human animals, there is some evidence that humans also show transfer to another target that had previously been modulated (trained as an occasion-setter), but not to a simple target paired with the outcome in a serial design (Experiment 4, Baeyens et al., 2004). Given how little research there is on occasion-setting in humans, it is unknown whether participants can learn that the feature is modulatory when the target and feature are presented simultaneously.
Finally, there is a third way (in addition to conditioned inhibition and occasion setting) by which participants may solve the feature negative discrimination. Configural theories such as Pearce’s (1987, 1994) state that compounds are represented as unique configurations rather than as the sum of their elements, and that associative links form between this configural representation and the outcome. In contrast, elemental theories (e.g., Rescorla & Wagner, 1972) posit separate associative links for each element in a presented compound (e.g., for AB, A and B have separate associations with the outcome). In order for configural theories to predict responding to novel configurations of stimuli, they must incorporate some generalisation mechanism. For example, Pearce’s (1987, 1994) model assumes that associative strength generalises between configurations based on the proportion of common elements they share.
Evidence from human causal learning studies suggests that participants do learn about stimuli in this configural way. However, interestingly, they do not always generalise between configurations to the extent predicted by Pearce (1987, 1994). Shanks, Charles, et al., (1998) showed that reversal learning (i.e., conditioning) of the negative feature B following feature negative training (i.e., A+/AB-, then B+) left performance to the original feature negative contingencies (A and AB) largely unaffected. In other words, learning that B was causal did not inflate ratings to the AB compound in a subsequent test, contrary to the predictions of elemental theories of learning (e.g., Rescorla & Wagner, 1972) but also, to a lesser degree, contrary to Pearce’s configural theory which predicts some degree of generalisation between B+ and AB- configurations. Shanks et al. interpreted their findings of intact feature negative performance as evidence for configural learning, but noted that participants were more configural than predicted by Pearce’s generalisation mechanism (see also Shanks, Darby, et al., 1998; Williams, 1995).
This reluctance to generalise suggests a memorisation strategy where participants attempted to remember the associations between cue configurations and outcomes during training with little attempt to infer the effects of individual cues (see also Williams et al., 1994). One limitation of this study is that Shanks, Charles, et al. (1998) did not include a summation test and thus it is unclear what participants learned about the feature B, and how they would transfer that learning to another excitor. In the absence of this test, an equally compatible conclusion from their results is that participants learned that B was a negative occasion-setter. Note, however, that the only study of negative occasion setting in humans showed that unlike animal studies, reversal training of the feature completely disrupted subsequent performance to the original feature negative contingencies (Experiment 3, Baeyens et al., 2004). Thus in the human literature, it is unknown whether a separate “configural” strategy exists that can be differentiated from negative occasion setting.
In summary, separate lines of evidence in both humans and non-human animals suggest that feature negative training, under different conditions, can lead to different types of learning about the feature. The overall goal of the study was to test whether participants learn different causal structures in a feature negative discrimination corresponding to conditioned inhibition, occasion-setting, and configural learning. These three types of learning differ in their underlying causal (or associative) structure (see Figure 1), which has implications for how much the feature should transfer in a summation test. Conventionally, conditioned inhibitors are assumed to directly inhibit activation of the outcome through an inhibitory associative link with the outcome. If participants have learned that B is inhibitory/preventive, they should show strong transfer of the inhibitory properties of B to a new target since B directly inhibits the representation of the outcome. Note that an alternative view proposed earlier by Konorski (1967) is that inhibition involves an association between the negative feature B and a “no-outcome” unit, which itself is in an antagonistic relationship to the “outcome” unit. We decided against including Konorski’s view of inhibition as it does not refer to causal relationships between environmental events and is more difficult to communicate than the simpler preventive option.
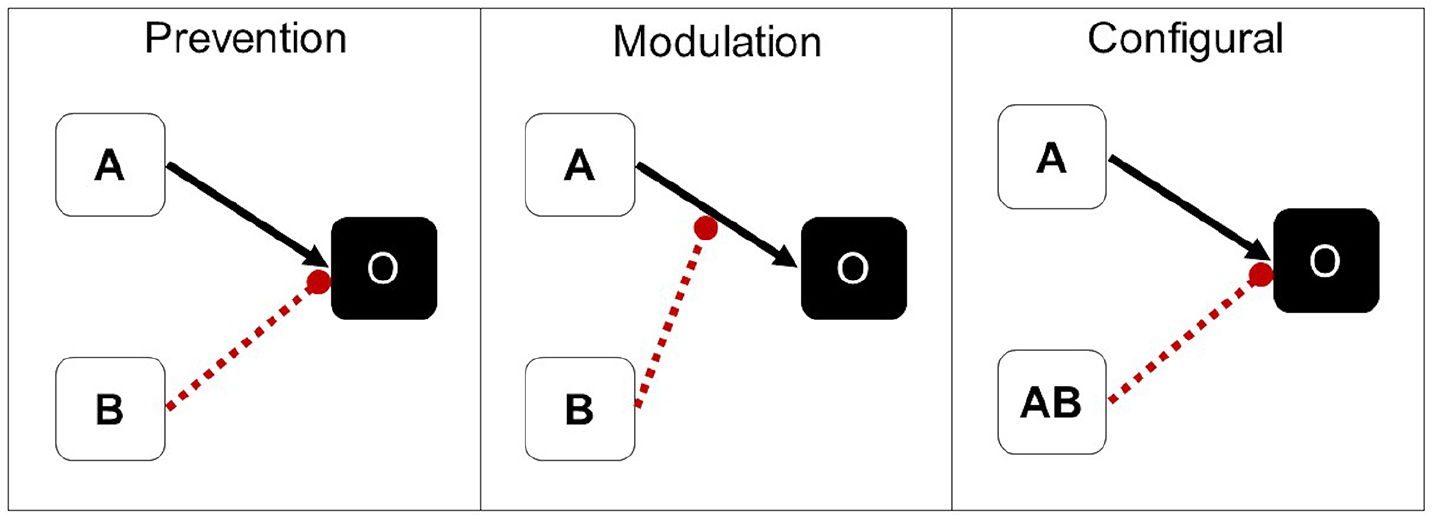
Associative/causal structures underlying conditioned inhibition (prevention), occasion-setting (modulation) and configural learning arising from training with feature negative (A+AB-) contingencies. A is the target and B is the negative feature. Solid black arrows indicate excitatory/causal associations, rounded-head, red dotted arrows indicate inhibitory/preventive associations.
In contrast, Ross and Holland (1981) suggested that occasion-setters act on the A-outcome association, modulating the effect of the target. If participants have learned that B is modulatory, they may or may not assume that the modulatory effects of B will extend to a novel target that had not previously appeared with B, producing an intermediate level of transfer. Finally, configural theories posit associative links between each unique configuration of cues and the outcome. For feature negative contingencies, this means that B is represented as part of the AB configuration that acquires a direct inhibitory link with the outcome (see Figure 1), leaving the status of B ambiguous. The results of Shanks and colleagues (Shanks, Charles, et al., 1998; Shanks, Darby, et al, 1998) suggest that participants memorise configurations of cues and their respective outcomes and do not generalise between configurations. Under this view, participants should be agnostic for all novel compounds, and therefore respond at baseline, showing the least amount of transfer.
There were three specific aims for the current study. The first aim was to test what proportion of participants learn about a negative feature in the manner predicted by the Rescorla-Wagner model (Rescorla & Wagner, 1972), by reporting a prevention causal structure (corresponding to conditioned inhibition). The second aim was to test whether there are individual differences in participants’ self-reported causal structure for the negative feature, even in the absence of any procedural manipulation. Specifically, we wanted to know whether subsets of participants reported a modulation (corresponding to occasion-setting) or configural causal structure using the typical feature negative procedure where elements of the compound are presented simultaneously. The last aim was to test whether the self-report measures of causal structure were consistent with the degree of transfer in a summation test.
Critically, in our study, we used explicit self-report measures to capture individual differences rather than data-driven methods (e.g., Glautier & Brudan, 2019). We have successfully used self-report in our previous work on generalisation to identify subgroups of participants who used different learning strategies, and found that these measures were consistent with other behavioural measures (Lee et al., 2018; Lovibond, Lee, & Hayes (2020); Wong & Lovibond, 2017). Although conditioned inhibition and occasion-setting have been distinguished in terms of their underlying associative structure in previous work (see Fraser & Holland, 2019), there has been little attempt to connect these effects to explicit causal structures in humans. The causal learning literature shows that participants interpret associations differently according to the causal model implied by the cover story (Waldmann et al., 2006; Waldmann & Holyoak, 1992), but there have been few attempts to investigate whether different causal models (structures) arise from associative learning procedures with more neutral cover stories. Such a connection would further strengthen the view that associative learning theories can account for how humans learn about causal relationships (Dickinson et al., 1984) and help clarify the role of higher-order cognitive processes in human associative learning (Lee, Lovibond, & Hayes, 2019; Lee, Lovibond, Hayes, & Navarro, 2019; McLaren et al., 2018; Mitchell, De Houwer, & Lovibond, 2009).
This study is novel in being the first to attempt to measure individual differences in feature negative learning via explicit self-report and to obtain occasion setting (and conditioned inhibition) in the absence of any procedural manipulation of the training conditions (e.g., serial presentation). In both experiments, we use the allergist task where participants learn which foods (cues) are causing allergic reactions (the outcome) in a fictitious patient, Mr X. Our choice of the allergist task was motivated by the fact that it reliably shows other cue competition effects like blocking. Although in real life, food allergies are not often offset by eating other (inhibitory) foods, we note that if anything, this works against us finding any evidence of inhibitory learning. Thus, the allergist task can be seen as a neutral cover story with respect to causal structure. Experiments 1 and 2 measured the causal structures that participants inferred following simultaneous feature negative training. Experiment 2 also attempted to manipulate which causal structure participants inferred by providing verbal hints prior to training.
Experiment 1
The aim of Experiment 1 was to test whether individual differences exist for feature negative learning, and whether these differences can be captured by self-report. The top row of Table 1 shows the design of Experiment 1. The training phase consisted of 6 trial types. A+ and AB- were the feature negative contingencies; A was the target causal cue and B was the negative feature, presented in a simultaneous compound. C+ was the transfer causal cue used in the summation test. DE- and F- provided control cues for the summation test; F- was trained as an unambiguous non-causal cue, while D was trained as a non-causal cue that appeared in combination with another cue. Note that although F is less ambiguous than D in its predictive power, D is arguably a more conservative control as it also appeared in a compound. In contrast, F may suffer generalisation decrement at test due to it never appearing in a compound prior to test. Note that D and E are equivalent and thus we only used one cue from the DE compound as a control in the summation test. Finally, GH+ was a filler compound included to prevent participants from learning that all compounds predicted no outcome.
Design of Experiments 1 and 2.

Following training, participants completed an outcome prediction test where they continued to make the same predictions but in the absence of feedback. CB was the critical summation test compound as it was composed of the feature B combined with the transfer causal cue C (Rescorla, 1969). Therefore, ratings for CB indexed how much participants transferred their learning about B to a new target (C). We included 3 different control compounds (CD, CF, CI) to compare against CB. The control cues consisted of the non-causal cues D (trained in compound) and F (trained alone), as well as a novel cue I that participants saw for the first time at test. We included these three different control compounds as previous human studies (e.g., Karazinov & Boakes, 2004; Shanks, Darby, et al, 1998) had used them and we wished to explore whether there were systematic differences between them.
We assessed causal structure in two ways. The first was via open-ended self-report and the second was with a three-alternative forced-choice (3AFC) question with options describing a preventive, modulatory, and configural causal structure for B. We also included a standard causal ratings test (e.g., Dickinson et al., 1984) to assess what participants learned about the causal status of each individual cue on a bipolar cause-prevent scale. Note, however, that our main hypothesis involved testing whether causal structure predicted differences in the transfer of inhibitory learning in the outcome prediction test.
Method
Participants
100 Mechanical Turk workers (27 female, M age = 34.3, SD age = 9.6) participated in this experiment in exchange for payment (12 min at US$10/hr). To be eligible, workers had to have an approval rate > 95% and have completed 500 Human Intelligence Tasks (HITs).
Apparatus and stimuli
The experiment was programmed using the jspsych library (de Leeuw, 2015) with stimuli and outcome pictures created as separate files (see osf.io/esfm3/). Each stimulus consisted of a picture of a food along with its verbal label on a white background and was 300 pixels wide and 400 pixels high. The experimental cues (A-I, 9 in total) were selected randomly from a total pool of 16 food pictures. The outcomes (allergic reaction and no reaction) were displayed as image files (609 x 380 pixels).
Procedure
All experiments were approved by the University of New South Wales Human Research Ethics Advisory Panel and participants provided written online consent. Table 1 shows the sequence of phases in Experiment 1.
Training phase
Participants were told that an allergist was trying to work out which foods cause allergic reactions in “Mr X.” On each trial, participants were first presented with the text “Mr X eats the following meal”: at the top of the screen, along with the contents of a meal that Mr X had eaten (pictures of either one or two foods with their verbal labels) below the text. Then, 500 ms after presentation of the cues, the prompt “Please rate the likelihood that Mr X will show an allergic reaction after eating this meal” appeared along with a visual analogue scale ranging from “Definitely NO ALLERGIC REACTION” to “Definitely ALLERGIC REACTION.” Participants made their prediction by moving the slider to a point on the scale. Once participants clicked on the scale (i.e., made any rating) a “Continue” button appeared below the scale. Participants had unlimited time to complete their rating in this, and all subsequent phases. After clicking “Continue,” the prediction scale and prompt disappeared and was replaced by feedback. This feedback consisted of text (“Allergic Reaction!” or “No allergic reaction”). If the outcome was an allergic reaction, a sad face accompanied the text. After 2 s, the stimuli and feedback disappeared and the participant entered the 1-s blank inter-trial-interval (ITI) period.
Table 1 shows the six trial types presented in the training phase. Each of these six trial types (A+, AB-, C+, DE-, F-, GH+) was presented twice in each block, with the left-to-right presentation of the compound cues counterbalanced within a block (e.g., AB and BA were presented in randomised order). Training consisted of three blocks in total, meaning that each trial type was presented six times. Trials were randomised such that the same trial type could not be presented on successive trials.
Outcome prediction test
After training was completed, participants were told that they would now be presented with more meals, and that they should continue to make the same predictions about the occurrence of an allergic reaction. However, in this phase, they were told that they would not receive any feedback. The trial types presented in the outcome prediction test are presented in Table 1, and consist of stimuli that were familiar (i.e., presented in training, A, AB, C, DE, D, E, F) as well as novel compounds for the summation test (CB, CD, CF, CI). Each trial type was presented twice at test, with the left-to-right presentation of compound cues again counterbalanced. Ratings in the training and outcome prediction test could range between 0 and 100.
Open-ended questions
Following the outcome prediction test, participants were asked two open-ended questions designed to elicit explanations for their ordinal pattern of ratings for the summation stimuli. The order of the questions (CB vs. CD and CB vs. CF) was randomised. On each trial, participants were presented with the three relevant cues at the top of the screen (C, B, and either D or F from left to right), and told that two of the meals that Mr. X ate were C+B and C+D/F. They were told that compared to CD/CF, they rated the likelihood of allergic reaction as LOWER/HIGHER/SIMILAR for CB. The displayed text was determined by their actual ratings in the outcome prediction test. The threshold for displaying “similar” was a ratings difference < 5 for CB and CD/CF. Participants were asked to explain why they rated the meals in this way by typing their response into a field box.
Causal ratings test
Participants were then asked to judge whether each food caused or prevented an allergic reaction. Participants were presented with the cues (A, B, C, D, E, F, and I) and asked “Please rate to what extent this food tended to prevent or cause an allergic reaction” on a visual analogue scale. The scale ranged from “Strongly PREVENTED ALLERGIC REACTION” to “Strongly CAUSED ALLERGIC REACTION,” with “No effect” labelled in the middle of the scale. The ratings were made in a similar way to the previous test phase with no feedback. The causal ratings were transformed to range between -100 and +100.
Causal structure assessment
In this phase, we asked participants specifically what they learned about cue B (the negative feature) using a 3AFC question. Participants had to select which of the following options best described what they thought about the food representing cue B. The options for the three different causal structures (configural, modulation, prevention) were presented in randomised order. The prevention option was phrased “It prevented allergic reactions in general,” the modulation option was phrased “It prevented allergic reactions caused by specific foods,” and the configural option was phrased “It is hard to know the exact role of individual foods such as this one. I concentrated on remembering which combinations of foods caused an allergic reaction and worked from there.”
Finally, the last question asked participants whether they wrote anything down during the training phase.
Results
Exclusion criteria
Participants were excluded from analysis if they failed to meet the training criterion. The training criterion was an average rating > 75 for the stimuli that predicted the outcome (A+, C+, GH+) and an average rating < 25 for the stimuli that predicted no outcome (AB-, DE-, F-) in the last block of training (i.e., last two presentations of each trial type). Participants were also excluded if they reported writing things down during training, as the instructions clearly stated for them not to. Of the 100 participants, 18 reported writing during training (and all 18 also failed the training criterion). After exclusions, 72 participants remained.
Causal structure subgroups
To examine the effect of inferred causal structure, we divided participants into subgroups based on their response to the 3AFC question. We did not use the open-ended responses to classify participants as many of the responses were not clearly interpretable with respect to causal structure (e.g., re-descriptions of the training contingencies). Note, however, that the open-ended responses did not reveal any alternative causal structures not covered by our existing forced choice options. Using the 3AFC question, there were 25 participants in the configural subgroup, 23 in the modulation subgroup, and 24 in the prevention subgroup.
Data analysis
Our primary measure of transfer was the ratings to the CB compound at test. In line with a traditional summation test (Rescorla, 1969), we also computed difference scores in outcome prediction ratings between CB (the transfer compound) and each of the three control compounds (CD, CF, and CI, see Supplementary Materials for these analyses). Note that we regarded CD as the most conservative control compound and therefore the best control for assessing the degree of inhibitory transfer.
To test the effect of causal structure, we planned 3 pairwise comparisons to examine the differences between subgroups (configural vs. modulation, configural vs. prevention, modulation vs. prevention). Since these contrasts were non-orthogonal, we applied Holm-Bonferroni correction to control the family-wise type I error rate at .05. For ease of interpretation, we report uncorrected p-values and note whether each contrast is significant after Holm-Bonferroni correction. The data were analysed in R (R Core Team, 2020) using the “afex” (Singmann et al., 2018) and “emmeans” (Lenth, 2019) packages.
Training
Figure 2 shows mean outcome predictions from the training phase for each subgroup. It is clear from Figure 2 that participants showed differential responding to stimuli predicting the outcome versus those that predict no outcome. From the second presentation, there are high overall predictive ratings of the outcome for the A+, C+, and GH+ trials, and low overall predictive ratings of the outcome for the AB-, DE-, and F-trials. We tested for differences in causal structure subgroups on terminal performance by calculating the difference between the average of the predictive (A+, C+, GH+) trial types and the average of the non-predictive (AB-, DE-, F-) trial types on the last two trials of training. There was no effect of causal structure subgroup for this difference score, F < 1.
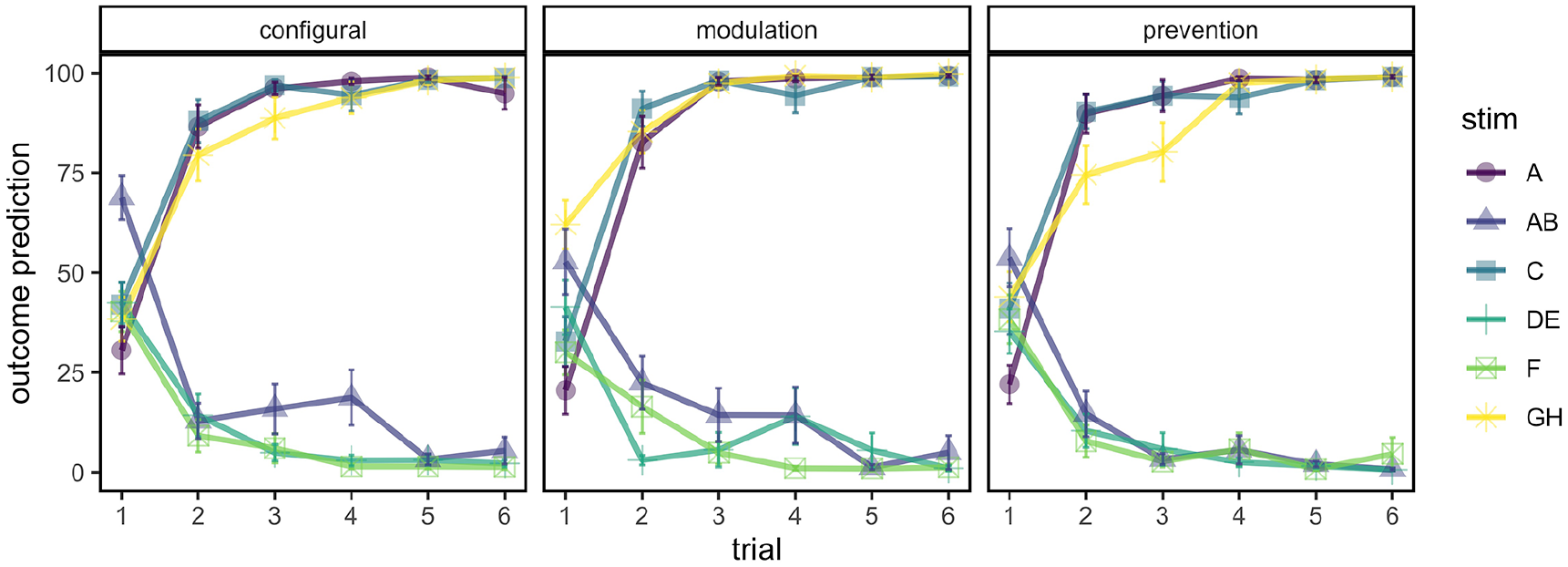
Mean outcome prediction ratings (+/– 1 SE of the mean) for each causal structure subgroup in the training phase in Experiment 1.
Outcome prediction test
For analysis of the outcome prediction test data, each participant’s ratings were averaged across the two test trials for each test stimulus, producing a single rating for each participant for each test stimulus. Figure 3 shows mean prediction ratings for the novel summation compounds split by participants’ reported causal structure (prevention, modulation, or configural). For brevity, we report only the analyses for the summation compounds (see Supplementary Materials for analyses for the other test stimuli).

Mean outcome predictions (+/– 1 SE of the mean) for the inhibition (CB) and control compounds (CD, CF, CI) in Experiment 1 split by causal structure subgroup.
As previously stated, our primary test of the effect of inferred causal structure involved outcome predictions for the CB compound. Examining Figure 3, participants who inferred a preventive causal structure gave the lowest outcome predictions for CB, participants who reported a configural strategy gave the highest outcome predictions for CB, and the participants who inferred a modulation causal structure gave ratings in between. In partial support of this observation, planned pairwise comparisons showed a significant difference in ratings to CB between the configural and prevention group, F(1, 69) = 12.2, p = .001, ηp2 = .15, but no significant difference between the configural and modulation groups, F(1, 69) = 1.79, p = .185, ηp2 = .03, nor the modulation and prevention groups, F(1, 69) = 3.4, p = .071, ηp2 = .06. Thus, there was some evidence that causal structure predicted the degree of transfer of learning about the feature B. In contrast, there were no significant pairwise comparisons between causal structure subgroups for any of the control compounds (CD, CF, or CI, see Supplementary Materials).
Causal ratings test
Figure 4 shows mean causal ratings for each test cue split by inferred causal structure. These results differ from the outcome prediction test in that all test cues were presented individually, and participants were asked to provide ratings on the prevent-cause scale, rather than simply predicting the likelihood of the outcome. The figure shows that participants in all causal structure subgroups gave ratings at ceiling for the causal cues A and C. It also shows that participants gave negative (preventive) causal ratings for the feature B, but also to some extent for the non-causal cues (D, E, and F) and the novel cue I. There were no significant differences between causal structure subgroups when examining A, C, D, E, F, or I (see Supplementary Materials). However for the feature B, causal ratings were significantly lower for the prevention subgroup than for the configural subgroup, F(1, 69) = 10.4, p = .002, ηp2 = .13. There was again no significant difference between configural and modulation, F(1, 69) = 3.77, p = .056, ηp2 = .05, and no significant difference between modulation and prevention subgroups, F(1, 69) = 1.51, p = .223, ηp2 = .02. Thus, the results from the causal ratings test mirrored the results from the outcome prediction test, with the clearest difference in causal structure being between the configural and prevention subgroups for the feature B.
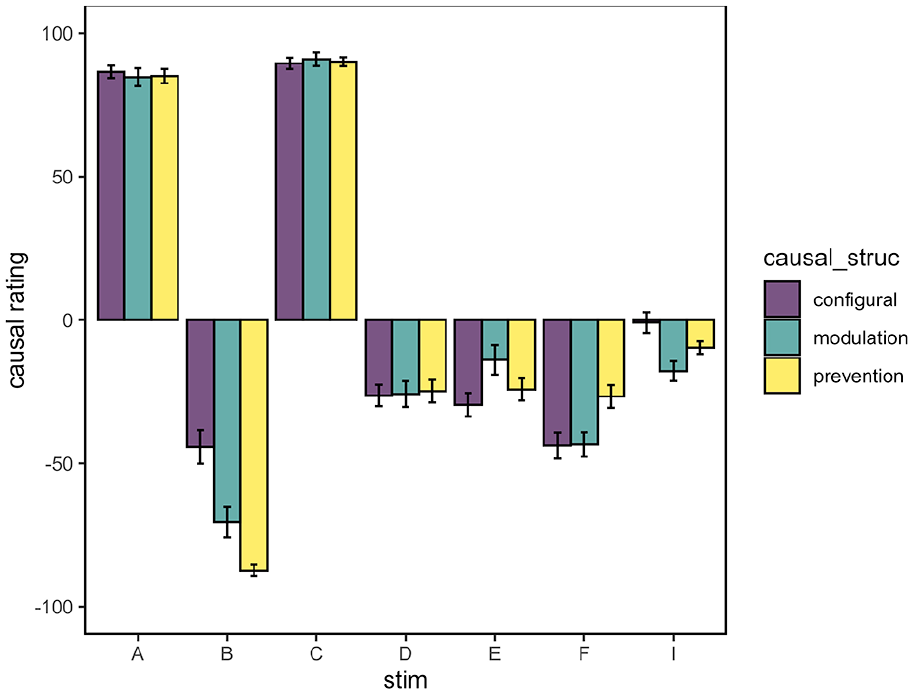
Mean causal ratings (+/– 1 SE of the mean) for individual test cues in Experiment 1. Positive ratings indicate causation, negative ratings indicate prevention, zero indicates no effect.
Discussion
In summary, participants learned the feature negative discrimination and showed evidence of transfer of inhibitory learning, producing lower outcome prediction ratings when the feature B was combined with a new target (C), compared to when the same target was combined with a non-causal control cue trained in compound (CD). The novel finding was that only a third of the participants actually reported a causal structure in line with conditioned inhibition. Participants were evenly split over the other two causal structures, demonstrating substantial individual differences in inferred causal structure in standard simultaneous feature negative training. Participants’ self-reported causal structure predicted the degree of transfer of inhibitory learning of the feature to the new target C, with the configural subgroup showing the highest predictive ratings (least transfer), the prevention subgroup showing the lowest predictive ratings (most transfer), and the modulation subgroup falling in between.
The differences between causal structure subgroups for CB on the outcome prediction test were mirrored in the causal ratings test for cue B. The ordinal pattern of ratings for B and the significant subgroup comparisons (prevention < configural) for this cue were consistent with ratings to CB on the outcome prediction test. Although typically included in human causal learning studies, the main limitation of the standard causal rating test for our purposes is that it assumes a prevention structure whereby inhibition is opposite to excitation, and hence may not differentiate between modulatory and preventive causal structures. Participants who inferred a modulation structure may have given similarly negative ratings to the prevention subgroup for the inhibitor B if they interpreted the question as asking about the effect of B on A. It is interesting to note that although cues D, E, and F were only ever shown to lead to no outcome (i.e., zero associative strength according to the Rescorla-Wagner model; Rescorla & Wagner, 1972), participants on the whole rated these cues as negative on the cause-prevent scale. It is possible that some participants may have interpreted the scale in a similar fashion to the outcome prediction scale, using the left (prevention) side of the scale for stimuli that predicted no reaction rather than prevention of the reaction.
In summary, Experiment 1 provided initial evidence of individual differences in how participants interpret the causal scenario following feature negative training, and showed that these self-reported causal structures predicted the degree of transfer of inhibitory learning. An important follow-up question is whether these individual differences are stable or amenable to manipulation. Experiment 2 sought to test this possibility.
Experiment 2
The primary aim of Experiment 2 was to test whether verbal hints could influence the inferred causal structure adopted by participants. If feature negative training is an ambiguous scenario where participants learn different causal structures, then participants may be sensitive to explicit hints about how the cues exert their effects. Alternatively, there may be stable individual differences that lead participants to interpret the contingencies using their preferred structure irrespective of any hints. Experiment 2 used a between-subjects design with 3 hint groups (configural hint, modulation hint, prevention hint). We used the same 3AFC question as in Experiment 1 to assess to what extent the hint manipulation influenced the causal structure participants inferred from the feature negative contingencies.
Method
Participants
Three hundred Mechanical Turk workers (105 female, M age = 34.6, SD age = 10.4) took part in Experiment 2. The eligibility criteria were the same for Experiment 1 with the addition that participants could not have participated in Experiment 1 before. Due to a technical issue, the data file for one participant was not saved, leaving 299 data files.
Procedure
The method was identical to Experiment 1 except for the following changes. Participants were randomly allocated to one of three hint groups (configural hint, modulation hint, prevention hint). The hint was presented prior to the training phase, with the following wording:
Prevention Hint
“The doctor suspects:
There is at least one food that
and also
there may be another food that
Modulation Hint
“The doctor suspects:
There is at least one food that
and also
there may be another food that
Configural Hint
“The doctor suspects:
There are
B and I were added as test cues to the outcome prediction test, and the open-ended question was changed with the aim of eliciting clearer responses. Instead of prompting participants to explain why they had rated CB in a particular way compared to CD and CF, the open-ended question in Experiment 2 asked participants to report what they had learned about B. We also changed the instructions for the causal ratings test slightly to help differentiate the scale from the outcome prediction scale used in the previous phase.
Results
Exclusion criteria
The same exclusion criteria were used as in Experiment 1. Eighty-seven participants failed the training criterion, and 49 reported writing things down during training (42 of whom also failed the training criterion). After exclusions, 205 participants remained.
Causal structure subgroups
Table 2 shows the division of participants into hint group and reported causal structure after exclusions. The open-ended answers were more informative than in Experiment 1, and aligned moderately well with the answers to the forced-choice question (see Supplementary Materials), but for consistency we again used the forced-choice question to classify participants into causal structure subgroups for analysis. Compared to Experiment 1, where the division of participants into causal structure subgroups was much more even, the majority of participants in Experiment 2 (55.1%) reported a modulatory causal structure.
Number of participants reporting each causal structure by hint group in Experiment 2.

Effectiveness of hints
To test the effectiveness of the hints on reported causal structures we conducted log-linear contrasts on the frequencies displayed in Table 2. Comparing the cell frequencies where the reported causal structure matched the hint (i.e., the diagonal entries) against the other cells produced a significant difference, χ2(1, N = 205) = 6.5, p = .011. The effect of each hint was examined by testing a contrast comparing the proportion of participants in each hint group who reported the matching causal structure relative to the proportion who reported that structure in the other two hint groups. This contrast was significant for the prevention hint, χ2(1, N = 205) = 8.1, p = .004, but not for the configural, χ2(1, N = 205) = 3.1, p = .078, or modulation hints, χ2(1, N = 205) = .193, p = .660. Thus, there was statistical evidence that the hints influenced participants’ causal structures overall, but when examining the hints individually, only the prevention hint produced a significant increase in the proportion of participants adopting that causal structure.
Data analysis
Since the hints appeared to influence the causal structures participants adopted, we did not include the interaction between hint and causal structure in our analysis of the test data. Instead, we analysed the effects of hint group and causal structure separately using the same pairwise contrasts as in Experiment 1.
Training
Figure 5 shows the training data for each causal structure subgroup averaged over hint groups (see Supplementary Materials for the breakdown by hint group). The acquisition curves look very similar in all cells despite the greater amount of variability for the configural subgroup due to the smaller n. Using the same measure for terminal performance as Experiment 1, there was a non-significant effect for hint group, F < 1, marginally non-significant effect for causal structure subgroup, F(2, 196) = 2.89, p = .058, ηp2 = .029, and a non-significant interaction, F(4, 196) = 1.84, p = .123, ηp2 = .036.

Mean outcome prediction ratings (+/– 1 SE of the mean) for each causal structure subgroup in the training phase in Experiment 2.
Outcome prediction test
Figure 6 shows the mean ratings to the critical test compounds split by causal structure (a) and by hint group (b). Similar to Experiment 1, there appear to be no differences between hint groups or causal structures for the control compounds (CD, CF, and CI), but there do appear to be differences for the test compound CB. Figure 6a shows a similar pattern between causal structure subgroups to Experiment 1, with the lowest ratings to CB in the prevention subgroup, the highest ratings in the configural subgroup, and the modulation subgroup in between. The ordinal pattern of ratings to CB is similar when examining the hint groups (Figure 6b), except that the modulation hint group gave similarly low ratings to the prevention subgroup.
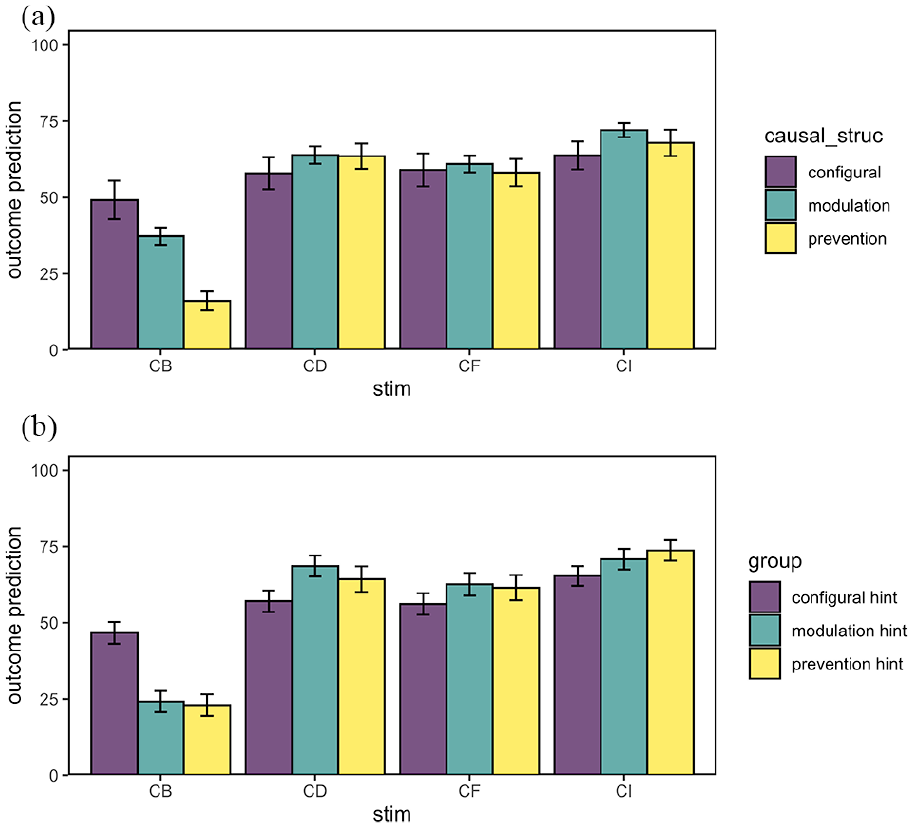
Mean outcome predictions (+/– 1 SE of the mean) for inhibition (CB) and control compounds (CD, CF, CI) in Experiment 2 split by (a) causal structure subgroup and (b) hint group.
The tests of causal structure differences were much stronger in Experiment 2 than in Experiment 1 (Figure 6a). For absolute ratings to the CB compound, the prevention subgroup gave significantly lower predictive ratings compared to the configural subgroup, F(1, 204) = 25.2, p < .001, ηp2 = .11, and also compared to the modulation subgroup, F(1, 204) = 21.7, p < .001, ηp2 = .10, but the difference between configural and modulation subgroups was marginally non-significant, F(1, 204) = 3.76, p = .054, ηp2 = .02. For the CB-CD difference score, clear statistical differences were found in Experiment 2 between all three causal structure subgroups (see Supplementary Materials for full details).
Examining the hint groups, the prevention hint group rated CB significantly lower than the configural subgroup, F(1, 204) = 22.5, p < .001, ηp2 = .10, and the modulation hint group also rated CB lower than the configural hint group, F(1, 204) = 21.1, p < .001, ηp2 = .09, but there was no difference between the prevention and modulation subgroups, F < 1 (see Figure 6b). This pattern of results for absolute ratings to CB was mirrored in the results examining the CB-CD difference scores (see Supplementary Materials for full details). Thus, it appears that the hints were partially effective in influencing the degree of transfer of inhibitory learning in the summation test.
Causal ratings test
Figure 7 shows the results from the causal ratings test split by causal structure (a) and by hint group (b). Examining the causal structure subgroups, Cue B was rated significantly lower (i.e., more preventive) in the modulation subgroup relative to the configural subgroup, F(1, 202) = 39.7, p < .001, ηp2 = .16, and also lower in the prevention subgroup relative to the configural subgroup, F(1, 202) = 40.0, p < .001, ηp2 = .17 (see Figure 7a). There was no significant difference comparing the modulation to prevention subgroup, F < 1. The pattern of subgroup causal ratings to B was similar to the pattern of ratings to CB in the outcome prediction test, except that there was no difference between causal ratings to B in the modulation and prevention subgroups. In contrast, the majority of pairwise comparisons between hint groups and causal structures were non-significant (see Supplementary Materials for full details). For the feature B, there were also no significant pairwise comparisons between hint groups, Fs <= 3.36. All 3 hint groups rated B as strongly preventive on the scale (Figure 7b).
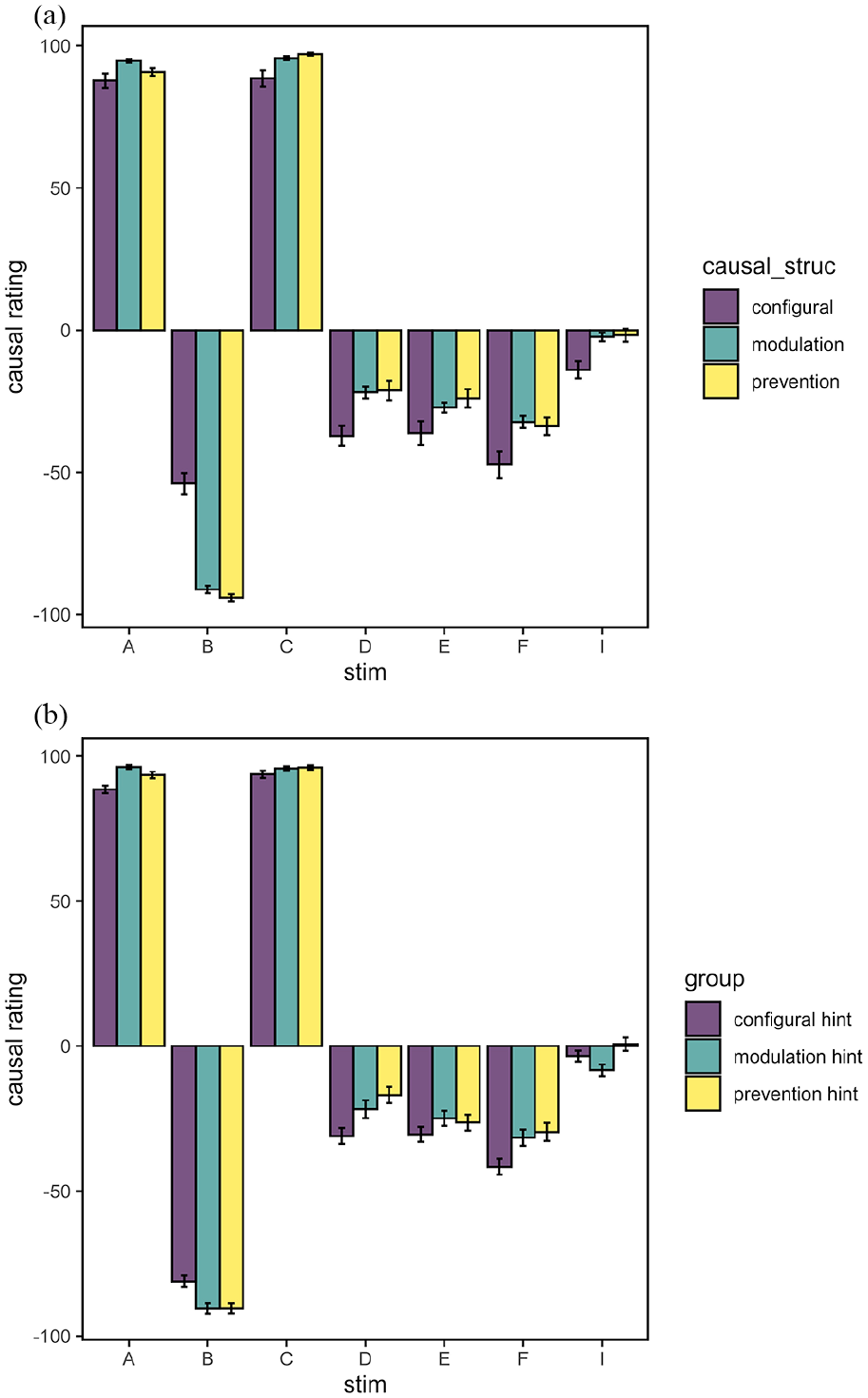
Mean causal ratings (+/– 1 SE of the mean) for individual test cues in Experiment 1 split by (a) causal structure subgroup and (b) hint group. Positive ratings indicate causation, negative ratings indicate prevention, zero indicates no effect.
Discussion
In Experiment 2, the majority of participants reported a modulation causal structure, again demonstrating that only a small subset of participants learn that a negative feature is preventive in the manner described by Rescorla and Wagner (1972). In comparison to Experiment 1, Experiment 2 provided stronger statistical evidence that self-reported causal structure predicted the degree of transfer of inhibitory learning. Participants who reported a preventive causal structure showed the strongest transfer of inhibitory learning, participants who inferred a configural strategy showed the least transfer, and participants who inferred a modulatory causal structure showed an intermediate amount of transfer. Although one of the pairwise comparisons in the outcome prediction test fell short of significance (configural vs. modulation), Experiment 2 replicated the key qualitative pattern of transfer results in Experiment 1, and the configural and modulation subgroups differed significantly on the causal ratings test.
Experiment 2 also extended Experiment 1 by showing that causal structure could be manipulated to some extent via verbal hints given prior to feature negative training. Participants given the prevention and modulation hint showed strong transfer, while participants given the configural hint showed weaker transfer. However, a substantial number of participants also showed resistance to the hint, reporting different causal structures to that presented in the hint. Our analyses revealed that although the hints were effective overall, the prevention hint was the only one that was effective in increasing the proportion of participants choosing the corresponding causal structure, explaining why the same pattern of results was not found comparing hint group and self-reported causal structure. One potential reason for this result may be that the hints differed in their degree of clarity, with the prevention hint offering the clearest indication of the correct underlying causal structure. Most importantly however, the ordinal pattern of transfer between causal structure subgroups was consistent with Experiment 1.
General discussion
Across two experiments, we showed considerable individual differences in the way participants learn the underlying causal structure of feature negative (A+AB-) contingencies. We found that a subset of participants reported learning that B prevented the outcome from occurring, another subset reported that B modulated the causal effect of A, and another subset were agnostic about the effects of B alone as it was never presented outside of the AB compound. The proportion of participants reporting each causal structure was approximately equal in Experiment 1, and slightly biased towards the modulation subgroup in Experiment 2. Thus, the structures we chose seemed adequate in covering the range of possible causal structures that participants learned (the open-ended responses in Experiment 2 confirmed this conclusion; see Supplementary Materials). Notably, only a modest proportion of participants in each experiment reported a preventive causal structure, suggesting that traditional associative models (e.g., Rescorla & Wagner, 1972) do not adequately capture what the majority of participants learn during feature negative training. These experiments demonstrate, along with Glautier and Brudan (2019), that feature negative training with the typical simultaneous procedure leads to a variety of learned causal structures in humans.
We also showed that self-reported causal structure was consistent with the degree to which participants transferred their learning about the feature (B) to a novel target (CB). Across both studies, participants who reported a preventive causal structure showed the greatest transfer of learning about the feature to a compound containing a new target (CB), the configural subgroup showed the least transfer, and the modulation subgroup showed an intermediate amount of transfer. Although we only assessed self-reported causal structure for B and not the control cues, participants also gave causal ratings to the feature B on the prevent-cause scale in broad alignment with their predictive ratings to CB, lending some support to the idea that our findings were specific to the negative feature B. However, for future work, it would be useful to assess participants’ inferred causal structure for the control cues (e.g., D) to ensure that participants’ reported knowledge is specific to B. Finally, Experiment 2 showed that explicit hints about the potential effect of the feature B were only partially effective in determining the causal structure that participants adopted, and there were less clear distinctions in terms of transfer in the summation test compared to self-reported causal structure. Even with the effect of the hint, the proportion of participants reporting a prevention causal structure was still only about a third (64/205), and only 33 participants (16%) reported a prevention causal structure in the absence of the prevention hint. The results from Experiment 2 suggest that there are strong pre-existing individual differences that determine how participants interpret the underlying causal structure of feature negative contingencies, but they are also somewhat sensitive to explicit hints about the effect of the feature.
Differences between causal structures
Thus far, we have argued that the differences in causal structure captured via self-report are qualitative. However, our modulation and prevention subgroups could instead be seen as differing quantitatively in the extent to which they are willing to generalise the properties they have learned about the negative feature B. To explore these potential quantitative differences between causal structure subgroups, we calculated a generalisation score for each participant (the difference between ratings for CB and AB on the outcome prediction test), and examined the relationship between this generalisation score and participants’ causal ratings for the negative feature B. Figure 8 shows a contour plot for Experiment 2 with the generalisation score on the x axis and the participants’ causal ratings for B on the y axis. The contour plot shows regions where the data cluster. Each contour connects regions that have similar density, and the areas in the middle of the contours have the highest density.

Density plot with individual data points overlaid from each causal structure subgroup in Experiment 2. The x-axis represents the generalisation score (difference in outcome prediction ratings between CB and AB) and the y-axis is the causal rating given to B in the causal ratings test (positive values indicate causation and negative values indicate prevention).
From Figure 8, it does appear that the main difference between modulation and prevention subgroups is the degree to which participants vary in their level of generalisation. On the x-axis, the prevention subgroup are clustered around 0, implying that the majority of participants gave very similar (low) ratings to both AB and CB, whereas the modulation subgroup’s scores vary across the full range from 0 to 100. The prevention subgroup distribution overlaps substantially with one end of the modulation subgroup, raising the possibility that even the participants who chose the prevention option could be considered to be modulators with a high degree of generalisation. The wide variation in generalisation scores within the modulatory group makes sense, as it would be ambiguous for these participants whether B will have the same effect on a novel target C as the original target A. This ambiguity then allows the expression of individual differences in willingness to generalise. Both subgroups are clustered at the negative end on the y-axis implying similar (preventive) learning about B’s effects, although as noted previously, the causal rating scale is unable to differentiate between modulatory and preventive causal structures.
In contrast to the other two subgroups, the configural subgroup are much more dispersed on both dimensions than the modulation and prevention subgroups. This pattern might be expected if participants have only learned about the AB configuration predicting no outcome, and are therefore agnostic about the effects of the negative feature B alone. This may lead participants to respond either conservatively (e.g., responding at 50, the midpoint of the scale) or responding randomly, which would result in a large amount of variability. It is also possible that in our efforts to capture the agnosticism about the causal effects of B in the configural participants, we inadvertently captured participants who were simply less confident about their predictions (and who responded somewhat randomly and therefore higher in their causal ratings for B). Thus quantitatively, the differences between our subgroups can be described as differences in variability in responding across two dimensions (causal learning and generalisation).
Implications for occasion-setting and conditioned inhibition
These experiments demonstrate that some participants endorse a causal structure consistent with occasion setting even with simultaneous presentation of the AB compound. Participants who chose the modulation option showed less transfer, on average, than the prevention subgroup. If participants who reported a modulatory causal structure have learned something akin to occasion setting, our experiments demonstrate that occasion setters can pass the typical summation test, but do so to a lesser extent than inhibitors. This means that in human studies, passing a summation test does not necessarily imply that participants have learned that the feature is inhibitory or preventive in the general sense described by Rescorla (1969).
Our results suggest that only a small proportion of participants (the preventors) learn “true” inhibition in the way described by Rescorla (1969) and the Rescorla-Wagner model (Rescorla & Wagner, 1972) given the standard single set of feature negative contingencies. However, further studies with different cover stories, cues, and outcomes are needed to verify the generality of this finding. At this stage, whether the prevention subgroup can be considered analogous to inhibition, or whether they are simply modulators who are willing to generalise (as the analysis in Figure 8 suggests), is unclear. Thus, if participants learn elementally, modulation may be the default outcome for feature negative training, contradicting traditional associative models such as the Rescorla-Wagner model, which predict that the negative feature acquires a direct inhibitory link with the outcome. While lending some support to the validity of associative models, our results also demonstrate that prevention learning is more complex and varied in humans than in animals, potentially explaining why conditioned inhibition effects as assessed in a summation test are less robust in humans (Karazinov & Boakes, 2007; Lee & Livesey, 2012; Williams, 1995; Wilkinson et al., 1989). An interesting avenue for future research would be to test whether classic manipulations known to increase the likelihood of occasion-setting in animals (e.g., serial vs. simultaneous presentation) result in a larger proportion of participants reporting a modulatory causal structure.
Implications for configural learning
An area of continued debate in associative learning is whether stimuli enter into associations with outcomes as configurations (Pearce, 1994) or elements (e.g., Harris & Livesey, 2010; McLaren & Mackintosh, 2000; Rescorla & Wagner, 1972), or whether both types of representation are needed (Rescorla, 1973). Our study suggests that a separate configural strategy can be differentiated from occasion-setting whereby participants learn about configurations of stimuli and are reluctant to make any strong assumptions about novel configurations they have not observed. Our results are thus consistent with studies demonstrating a failure to generalise between configurations of stimuli (Shanks, Charles, et al., 1998; Shanks, Darby, et al., 1998; Williams et al., 1994). Melchers et al. (2008) have proposed that whether participants learn elementally or configurally depends on properties of the experimental task. In other words, the degree to which participants learn configurally is somewhat flexible. This idea is also instantiated in Wagner’s (2003) Replaced Elements Theory. The r parameter determines the proportion of stimulus elements that is replaced when the stimulus is presented with another cue (or context) and therefore controls whether a stimulus is processed more (high r) or less (low r) configurally. Rather than configural learning being a feature of the task or the accompanying stimulus, our results suggest that certain participants have a predisposition towards learning configurally. This idea is consistent with studies demonstrating individual differences in attention to global or local stimulus features, which may account for why some participants appear to learn elementally and others configurally (Byrom & Murphy, 2014, 2016)
Concluding remarks
The presence of reliable individual differences in our task suggest that greater efforts should be made in human associative learning tasks to identify learning strategies that differ in important qualitative ways (e.g., Glautier & Brudan, 2019; Shanks & Darby, 1998). We have already shown that group-level trends can be misleading when there are subgroups of participants who learn and generalise in different ways (Lee et al., 2018). Similarly here, concluding in favour of an effect (conditioned inhibition) due to functional characteristics (passing the summation test) would contradict the explicit report and the transfer behaviour of a large proportion of participants. Our results complement recent calls for further investigation into the generality of classic associative learning effects (Maes et al., 2016; Urcelay, 2017) and greater recognition of individual differences in theory and analysis (Lee et al., 2018; Lee, Mills, Hayes, & Livesey, 2020). Acknowledging that humans can solve associative learning tasks by learning qualitatively different content will facilitate better connections with the animal conditioning literature, and provide a better understanding of the diverse ways in which humans learn about causal and preventive relationships in the world.
Supplemental Material
QJE-STD-20-015r2_supplemental – Supplemental material for Individual differences in causal structures inferred during feature negative learning
Supplemental material, QJE-STD-20-015r2_supplemental for Individual differences in causal structures inferred during feature negative learning by Jessica C Lee and Peter F Lovibond in Quarterly Journal of Experimental Psychology
Footnotes
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This study was funded by an Australian Research Council Discovery Grant (DP190103738) awarded to Peter Lovibond.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.