
Abstract
Fear extinction is not permanent but is instead more vulnerable than the original fear memory, as traditionally shown by the return of fear phenomena. Because of this, techniques to mitigate the return of fear are needed in the clinical treatment of related psychological conditions. One promising strategy is the occasional reinforced extinction treatment, introducing a gradual and sparse number of conditioned stimulus–unconditioned stimulus (CS-US) pairings within the extinction treatment. We present the results of three experiments in which we used a threat conditioning procedure in humans. Our main aim was to evaluate whether occasional reinforced extinction could reduce two different forms of relapse: spontaneous recovery (Experiments 1 and 2) and reinstatement (Experiment 3). Contrary to our predictions and previous literature, the results indicate that an occasional reinforcement treatment did not mitigate relapse compared with standard extinction. From a theoretical standpoint, these results are more consistent with the idea that extinction entails the acquisition of new knowledge than with the idea that there are conditions in which extinction leads to a weakening of the original fear memory. These findings also question the generality of the potential benefits of using occasional reinforced extinction in clinical settings.
Keywords
Introduction
The repeated presentation of a feared cue in the absence of aversive consequences leads to the extinction of the fearful response to the cue (Pavlov, 1927). The process underlying extinction could be key to the success of exposure therapy in which repeated, systematic, controlled exposure to a feared stimulus results in the reduction of the anxious response to that stimulus (Graham & Milad, 2011; Urcelay, 2012). Unfortunately, the effects of extinction are not permanent, and several factors can lead to a recovery of the initial fear response or relapse. For example, the mere passage of time after the extinction treatment may lead to a relapse of the initial fear response (i.e., the spontaneous recovery effect; Pavlov, 1927). Another factor that has been related to relapse is the experience of a stressful situation after extinction, even if that situation is unrelated to the initially feared cue (i.e., the reinstatement effect; Rescorla & Heath, 1975). These forms of relapse have been evidenced in non-human animal experiments and human conditioning studies, using different experimental paradigms and, interestingly, after exposure therapy in clinical settings (Craske et al., 2012). Estimates of return of fear after therapy range from 19% to 62%, according to the review by Craske and Mystkowski (2006). These standard forms of relapse also speak in favour of the idea that extinction could be used as a laboratory model to improve the effectiveness of exposure therapy (Milad & Quirk, 2012).
Thus, the current challenge is not so much to achieve fear reduction but to prevent a relapse of the fear response (Dunsmoor et al., 2015; Vervliet et al., 2013), a serious difficulty for cognitive-behavioural therapy of anxiety disorders. This vulnerability of extinction thus poses a challenge for achieving successful exposure therapy. A deeper understanding of the extinction mechanism itself will help overcome this challenge as it would shed light not only on the origins of such vulnerability but also on how this could be neutralised (Craske et al., 2018; Vervliet et al., 2013). The general objective of the present work was to evaluate the effectiveness of occasional reinforced extinction (ORE) to reduce two forms of relapse (spontaneous recovery and reinstatement) in a human conditioning preparation using threat expectancy ratings, a measure associated with the cognitive component of the fear response (Boddez et al., 2013; Lipp, 2006). Craske et al. (2014, 2018) reviewed the various methods used to address this challenge and argued that the occasional inclusion of reinforced trials during the extinction treatment is one possible way of reducing relapse effects, suggesting that ORE could be a viable and general strategy for enhancing inhibitory learning and its retrieval, with potential translational value in the clinical domain. Consistent with this idea, in a case study, Krompinger et al. (2019) suggested that Obsessive-Compulsive Disorder patients can benefit from confronting negative outcomes while in treatment by allowing them to “confirm their fears.” This could represent a learning opportunity; that is, encountering negative consequences does not mean they are failing, which may help prevent relapse. Moreover, experiencing the unconditioned stimulus (US) during treatment could allow them to embrace their full range of emotions, which can be therapeutically beneficial.
Interestingly, Gershman et al. (2013) found that ORE could have preventive effects on spontaneous recovery and reinstatement. These authors included three extinction groups in an animal conditioning study: gradual ORE, reverse ORE, and standard extinction. Experiment 1 showed that applying an extinction treatment with a gradual decrease in the number of reinforced trials (gradual ORE group) could reduce the spontaneous recovery of fear when the conditioned stimulus (CS) was presented 1 month later. In Experiment 2, this relapse-preventive effect of ORE was extended to reinstatement. After having experienced isolated presentations of the US before the test, Rats in the gradual ORE group showed less fear to the CS than those in the other two groups. Interestingly, the reverse ORE group included the same small number of reinforced trials, but these were distributed differently so that only in the gradual ORE group did the number of reinforced trials gradually decrease across the extinction treatment.
In Gershman et al.’s (2013) account of their results, relapse prevention critically depends on the specific distribution of reinforced trials during extinction in the sense that only a gradual decrease of reinforced trials after acquisition will have this preventive effect. This account assumes that the onset of a standard extinction training produces significant prediction errors (i.e., discrepancies between the predicted outcome—the US—and the experienced outcome—no US). In reinforcement learning models such as Rescorla and Wagner (1972), prediction errors serve as a learning signal, driving the modification of these predictions, namely, reducing the strength of the original fear memory. More recent models (e.g., Gershman et al., 2010; Redish et al., 2007) propose that the accumulation of these prediction errors may also serve as a segmentation signal, indicating a novel state in the environment and, thus, demanding the formation of a new memory. In this case, a new competing inhibitory memory is formed, allowing the original fear memory to persist primarily unmodified. The original association would no longer be updated, and a new context-dependent inhibitory association would be created (see also Bouton, 1993, 2002).
On the contrary, should these prediction errors be small or infrequent but still large enough to drive learning, the original fear memory will be modified, as no segmentation signal would be present. In this case, the extinction treatment will now result in the formation of a modified memory associated with safety. Thus, the expression of this inhibitory memory will not depend on contextual cues, and the original fear memory will not relapse. Gershman et al. (2013) suggested that a gradual ORE treatment (but not the reverse ORE) would have precisely this effect, as this will involve a gradual decrease in the number of reinforced trials. In other words, according to their proposal, gradual ORE represents a condition in which prediction errors are sufficient to drive the modification of the original fear memory but not large enough to serve as a segmentation signal. The presence of sparse, gradually diminishing CS-US trials in gradual ORE ensures that the accumulation of prediction errors will not be as persistent as reverse ORE or a standard extinction condition. Note that the presence of these reinforced trials brings extinction closer to the acquisition training. Under these conditions, prediction errors will not serve as a segmentation signal (see Gershman et al., 2017, for computational details concerning this assertion).
A relevant question, therefore, is whether this same preventive effect produced by gradual ORE would extend to human participants in a threat conditioning preparation. Previous attempts have yielded mixed results regarding these preventive effects on spontaneous recovery and reinstatement. Fortunately, those preventive effects could be demonstrated in a study with human participants, albeit on a different form of relapse, namely, the rapid reacquisition of conditioning after extinction training (see Ricker & Bouton, 1996, for an animal study on this phenomenon). Morís et al. (2017) used an aversive sound as the US and expectancy ratings to measure contingency learning. This study showed that the occasional presentation of CS-US pairings (i.e., reinforced trials) during the extinction phase diminished later reacquisition of the conditioned response (CR) compared with a standard extinction treatment. In the ORE group of their Experiment 3, after the initial acquisition phase, the number of reinforced trials gradually diminished across extinction, with five of the first eight extinction trials being reinforced, three of the second eight trials, one of the third eight trials, and none of the last 32 trials. Across the experimental series, the authors showed that (a) expectancy ratings at the end of extinction were equivalent in the ORE and the standard extinction groups, and that (b) the rate of reacquisition was slower in the ORE than in the standard group when the CS and the US were consistently paired again in the final phase. In other words, the rapid reacquisition was alleviated in a seemingly paradoxical way, that is, by intermixing a low number of CS-US pairings (i.e., reinforced) with standard CS-alone (i.e., non-reinforced) trials. This same pattern of results had previously been demonstrated in animal conditioning studies. Bouton et al. (2004) and Woods and Bouton (2007) found, in appetitive classical and instrumental conditioning, respectively, that including some pairings between the CS and the US as part of the extinction treatment slowed down the rate of reacquisition of a previously extinguished response. This specific preventive effect of ORE on the rapid reacquisition of conditioning could provide support for a different account of how ORE exerts its effects.
The explanation for the ORE effect offered by Bouton and collaborators (Bouton, 1993, 2002; Ricker & Bouton, 1996) assumes that during extinction, the memory of the original CS-US excitatory association is preserved and a new inhibitory association between the CS and US representations is created. Moreover, this new inhibitory memory is context dependent. While the original excitatory memory will be activated with each presentation of the CS, the inhibitory memory will only be activated to the extent that the relevant contextual cues (i.e., those experienced during extinction) are present. As the overlap between those cues and the contextual cues of a given trial reduces, so does the expression of the inhibitory association, leading to an increased CR.
Furthermore, and consistent with Capaldi’s (1967, 1994) sequential learning theory, Ricker and Bouton (1996) suggested that reacquisition is controlled by a “trial-signalling” mechanism, whereby the individual learns that a specific type of trial—either reinforced or non-reinforced—reliably signals the type of upcoming trial. As a result, during acquisition, the subject learns that the CS is paired with the US and also that reinforced trials follow other reinforced trials, whereas during standard extinction, individuals learn that non-reinforced trials follow other non-reinforced trials. Consequently, the preceding trial (reinforced or non-reinforced) becomes part of the context that controls responding. In the case of the standard extinction treatment, a reinforced trial is a very distinctive feature of the acquisition context, which is not present in the extinction context. Instead, when an ORE treatment is applied, reinforced trials are also part of the extinction context. Thus, during reacquisition training, reinforced trials in the ORE group, but not in the standard extinction group, can also facilitate the retrieval of the memory of non-reinforced trials (i.e., the inhibitory association learned during extinction training), slowing down the reacquisition of the CS-US contingency. According to this account, ORE is expected to alleviate rapid reacquisition but not other return of fear phenomena, such as spontaneous recovery or reinstatement.
Thus, an evaluation of the scope of the preventive effect of gradual ORE will also serve to disentangle the predictions derived from these two accounts of ORE effects. A relevant experimental strategy would be to include a CS-US trial as the first test trial. Both accounts make different predictions about how threat expectancies should be affected as a result of experiencing this first reinforced trial. According to Bouton’s (2004) account, the inhibitory memory will only be activated provided that the context is sufficiently similar to the extinction context. After a retention interval, the first presentation of the CS on the test (in a spontaneous recovery design) or after isolated US presentations (in a reinstatement design) must constitute a context change that is identical in the ORE and standard extinction groups. Thus, on the first test trial, similar levels of relapse should be expected in both groups. However, once the presence of the US has accompanied the first presentation of the CS (i.e., a reinforced first test trial), the predicted results differ between the two groups. Only in the ORE group will the extinction and test contexts overlap, as reinforced trials were also part of the former, promoting the retrieval of the extinction memory. Therefore, on the second test trial, threat expectancies should be lower in the ORE group than in the standard extinction group.
According to Gershman et al. (2013), during extinction training in the ORE group, the original CS-US memory will be weakened, becoming eventually extinct. Thus, the first presentation of the CS on the test should produce no sign of relapse in this ORE group (i.e., no spontaneous recovery or reinstatement). However, after experiencing the first reinforced test trial, some level of CR should be expected in the ORE group. And, of course, this recovery would be expected across test trials in the standard extinction group.
Nonetheless, previous attempts to evaluate the effectiveness of ORE in preventing both spontaneous recovery and reinstatement, or even rapid reacquisition, have yielded results that are far from conclusive. Shiban et al. (2015) evaluated the preventive effects of gradual ORE on the reinstatement of the fear response using a virtual reality conditioning paradigm. The authors used a 2-day protocol in which reinstatement was measured 24 hr after the extinction phase had been completed. Thus, some spontaneous recovery could also have affected the results obtained. In their experiment, performance in the gradual ORE group was compared with that of a standard extinction group. The results revealed that gradual ORE prevented the reinstatement effect only when measured by startle responses but not threat expectancies or skin conductance responses (SCR).
Culver et al. (2018) also evaluated the preventive effect of ORE, though not in its gradual version (i.e., aversive stimulation was also included during the last block of trials of the extinction treatment). Specifically, they evaluated the preventive effect of ORE on spontaneous recovery, rapid reacquisition, and a final retest phase 1 week after acquisition and extinction training had been completed. In this case, rapid reacquisition could also have been affected by the spontaneous recovery of the fear response. The results showed that, compared with the ORE group, the standard extinction group showed an increase in the CR from the last trial of extinction to the first test trial, measured as SCR and expectancy ratings. However, these results must be interpreted with caution because asymptotic extinction was not reached. The rate of conditioning reacquisition was slower in the ORE group compared with a standard extinction group and was measured by the SCR rather than valence ratings or threat expectations. In their final retest phase, significant group differences were only found for the SCR measure, which were highly driven by the low response shown by the ORE group at the end of reacquisition.
Finally, Thompson et al. (2018) assessed the effect of ORE compared with standard extinction training, including also unpaired presentations of the US (i.e., independent of CS presentations). These authors evaluated the effect of ORE on spontaneous recovery (using a 10-min interval between extinction and test), reinstatement, and rapid reacquisition. The results showed that ORE and unpaired US presentations during extinction eliminated spontaneous recovery compared with standard extinction, as measured by SCR. The ORE treatment did not differ from standard extinction regarding rapid reacquisition, although the latter was reduced by the unpaired extinction treatment, as measured by SCR. None of the treatments were effective in attenuating reinstatement, although it is important to consider that these manipulations were not wholly effective as no relapse effect was obtained in the control group. No differences between groups were found regarding CS valence ratings. It should be noted that in their design, and similar to Culver et al. (2018), all return of fear phenomena were evaluated sequentially, one after the other, so it was not possible to measure the effects of the various extinction treatments on each phenomenon independently, due to possible carryover effects and the corresponding confound.
The set of studies described (Culver et al., 2018; Shiban et al., 2015; Thompson et al., 2018) could not demonstrate preventive effects on the return of the cognitive component of fear, at least when measured by threat expectancy or CS valence ratings. Regarding skin conductance results, the evidence is mixed. However, even if other components of the fear response are positively affected by ORE (i.e., preventive effects are observed), the fact that threat expectancies are not may eventually cause the return of fear in those other components and, thus, a more generalised return of fear (Boddez et al., 2013). It should be noted that, in all these studies, the various relapse effects were not measured independently from each other but in a sequential way, which might have obscured any ORE preventive effect.
The design of the present experimental series differs from the designs used in these previous evaluations of ORE effects with human participants. In all previous studies, the number of extinction trials used was lower than in our experiments. Thus, it is not clear whether fear extinction in the ORE group was at asymptote using those procedures. Specifically, each CS was presented seven times more frequently during extinction than during acquisition in our series. This 1:7 ratio was reduced to 1:4 in Thompson et al. (2018), to 1:3 in Culver et al. (2018), and 4:5 in Shiban et al. (2015). Culver et al. (2018) and Shiban et al. (2015) found, even at the end of extinction training, a difference between CRs to CS+ and CS–. Conceptually, an important prerequisite is that extinction must effectively be established before assessing any form of a return of fear phenomenon.
Therefore, in our study, the extinction phase, as in the study by Morís et al. (2017) and following Gershman et al. (2013), included a gradually decreasing number of reinforced trials in the ORE group. Unlike previous attempts, (a) the number of trials during extinction training was sufficient to ensure that asymptotic expectancy ratings were equivalent in both the ORE and the standard extinction groups; (b) we focused on threat expectancy ratings (i.e., a cognitive component involved in the multidimensional fear response; given the interconnectedness of the different components of the fear response, this may be regarded as a starting point for a complete assessment of the preventive value of ORE); and (c) the possible preventive effects of ORE on spontaneous recovery and reinstatement were assessed independently from each other; thus, assessing one of these effects does not condition or interfere with the assessment of the other.
Overview of the experiments
Specifically, we followed Morís et al.’s (2017) Experiment 3 protocol, which used a gradual ORE treatment, as described previously. The procedure included three phases. The first phase included a threat conditioning task with a loud noise serving as the US (see Lonsdorf et al., 2017, for evidence of the effectiveness of this US in threat conditioning preparations). The second phase included an extinction task, with two independent groups of participants: a gradual ORE group and a standard extinction control group. Finally, a third phase included specific tests to measure the return of fear phenomena (see the various “Method” sections of the experimental series). In all cases, this test phase comprised a series of trials rather than a single test trial. The first test trial served to assess the potential preventive effect of ORE on the relapse phenomena considered. Note that Gershman et al.’s (2013) account predicts no relapse effect in the ORE group on this first test trial, whereas Bouton’s accounts predict such relapse. The remaining test trials were included to evaluate Bouton et al.’s predictions concerning the operation of the trial-signalling mechanism. It is important to note that the objective of these additional trials was not to test if ORE slows down rapid reacquisition but to test the predictions made by Gershman et al. (2013), Bouton et al. (2004) and Woods et al. (2007). Experiment 3 of Morís et al. (2017) has already provided evidence of the effectiveness of ORE in slowing rapid reacquisition in a direct test of this relapse effect with a very similar task. In the current series of experiments, introducing another relapse effect before this reacquisition test allows us to test the predictions already mentioned. However, this procedure prevents a clear interpretation of the results of the reacquisition test regarding the rapid reacquisition effect, due to the confound previously discussed.
For example, in Experiments 1 and 2, after presenting the first reinforced trial, the remaining test trials were also reinforced trials. In the ORE group, if the first test trial is reinforced, the mechanism will lead to the retrieval of the memory of non-reinforced trials, and such retrieval may affect what is being learned during the remainder of the test phase. Thus, it is expected that reacquisition will be slower in the ORE group than a standard extinction group. On the contrary, in Experiment 3, the remaining test trials were non-reinforced trials, and this time, the memory retrieved due to the signalling mechanism may also interfere with re-extinction in the ORE group. Note that during extinction training, participants in this group experienced a reinforced trial after a series of non-reinforced trials. Thus, the non-reinforced trials could retrieve the memory of reinforced trials, which could hinder re-extinction in the ORE group.
Experiment 1
The main objective of this experiment was to test whether an ORE treatment could prevent spontaneous recovery in a human threat conditioning preparation after a 24-hr interval between extinction and test. More specifically, we aimed to evaluate the possible differential effect of ORE and standard extinction training on relapse prevention after experiencing this retention interval. In this sense, we are not interested in spontaneous recovery itself but in this differential recovery effect produced by ORE and standard extinction. For convenience, we will refer to this differential effect as spontaneous recovery. In addition, the final test phase included a series of reinforced CS-US trials to assess the hypothesis derived from the trial-signalling mechanism proposed by Ricker and Bouton (1996).
Method
Participants and apparatus
In total, 118 undergraduate psychology students from the Universities of Oviedo and Málaga completed the task (97 females). None of the participants had any previous experience with this task. Their participation was voluntary, and they received course credit in exchange. Visual stimuli were presented on a 21-inch monitor with a 1,920 × 1,080 pixels resolution, while auditory stimuli were presented using individual headphones. The experimental programme was written using E-Prime 2.0 (Psychology Software Tools, Pittsburgh, PA, USA). This study was approved by the Ethics Committee of the University of Málaga (CEUMA-2013-0006-H) and was conducted in accordance with the Code of Ethics of the World Medical Association (Declaration of Helsinki). As with the rest of the experimental series, the sample size included the maximum possible number of participants that could be recruited from the convenience samples of students available at the time of each experiment. After the planned rejection of participants (see “Pre-analysis participant rejection” section), this sample provided an 80% probability of detecting an effect of size d = 0.55 for the pre-planned t-test of the first Cue A trial of the test phase.
Design and procedure
The design of the experiment is summarised in Table 1. The experiment included three phases: acquisition, extinction, and test. During the acquisition phase, participants were exposed to eight reinforced trials in which a stimulus (Cue A) was followed by the presentation of an aversive stimulus (US; aversive sound), and eight unreinforced trials in which another stimulus (Cue B) was never followed by the US. We used a differential conditioning procedure to ensure minimal discrimination training during the task. This way we could obtain differential responses to both trial types as evidence of participants’ understanding of the contingencies programmed. However, in our experimental series, the focus of interest was not differential conditioning. As described in the “Introduction” section, our target experimental hypotheses are exclusively concerned with expectancy ratings of Cue A throughout the different phases of the experiments. The trials were presented in a pseudorandom order. Out of each group of four trials, two of each type were presented in a randomly selected order. This pseudorandom order was used across the different phases of the experiment.
Summary of the experimental designs.
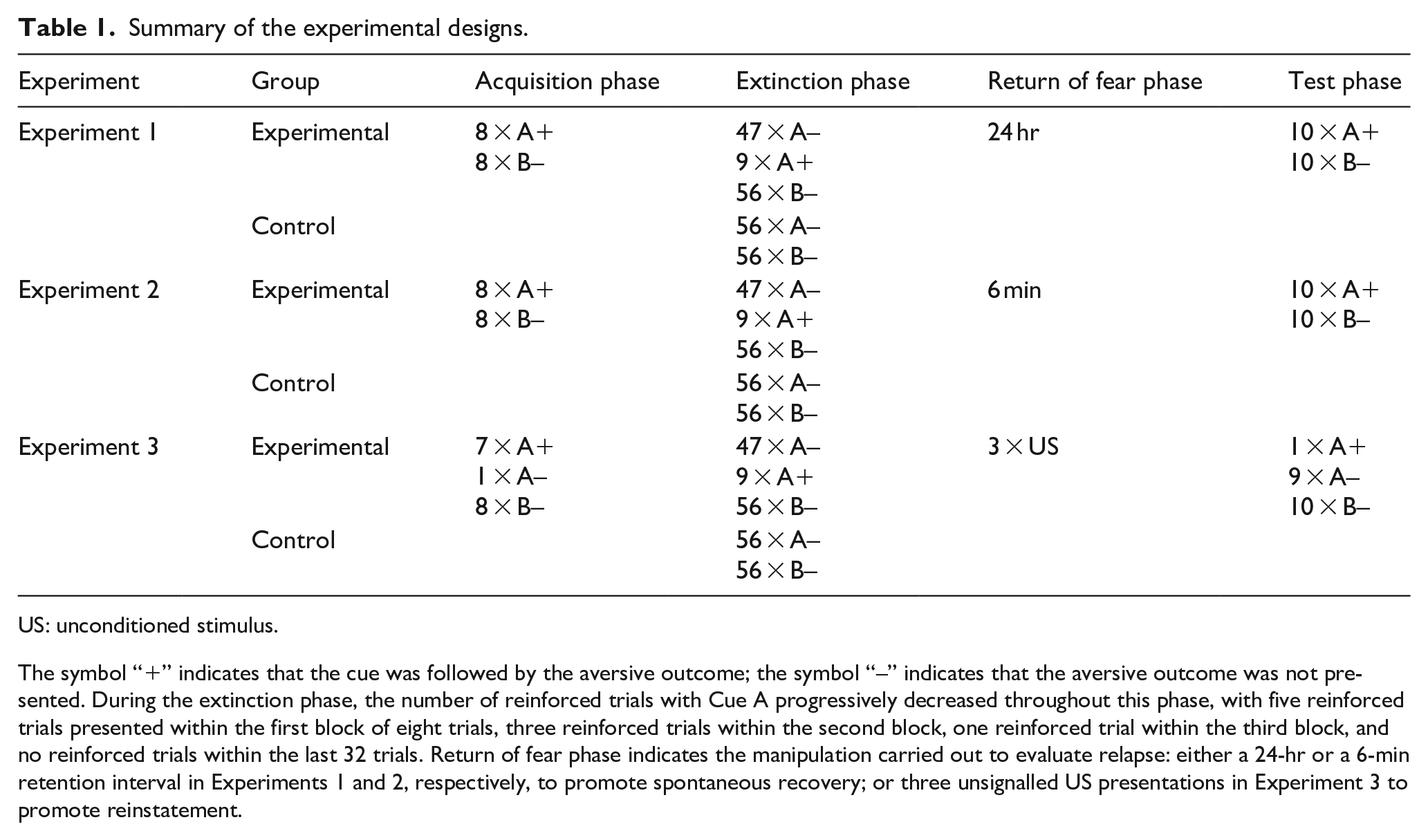
US: unconditioned stimulus.
The symbol “+” indicates that the cue was followed by the aversive outcome; the symbol “–” indicates that the aversive outcome was not presented. During the extinction phase, the number of reinforced trials with Cue A progressively decreased throughout this phase, with five reinforced trials presented within the first block of eight trials, three reinforced trials within the second block, one reinforced trial within the third block, and no reinforced trials within the last 32 trials. Return of fear phase indicates the manipulation carried out to evaluate relapse: either a 24-hr or a 6-min retention interval in Experiments 1 and 2, respectively, to promote spontaneous recovery; or three unsignalled US presentations in Experiment 3 to promote reinstatement.
The experimental manipulation took place in the extinction phase. Participants were randomly assigned to one of two groups: the experimental group (ORE) or the control group (standard extinction). Participants in the ORE group were exposed to 47 non-reinforced trials and nine reinforced trials of Cue A (see Table 1 for more details). Participants in the standard extinction group were exposed to an identical sequence of trials, except that Cue A was never followed by the US. In addition, in both groups, there were 56 non-reinforced trials of Cue B.
In the test phase, the initial trial for all participants was a reinforced trial with Cue A (the target cue for which the tested predictions are concerned; see the “Introduction” section for details). After that, a series of nine reinforced trials of Cue A and 10 non-reinforced trials of Cue B were randomly presented.
After the test phase, participants were asked to complete two rating scales regarding the discomfort and anxiety caused by the aversive sound. First, they were asked to indicate how much discomfort they felt when listening to the aversive sound throughout the task on a scale from 1 (extremely low) to 9 (extremely high). After this, they had to indicate how much anxiety the sound caused them, on a scale from 1 (extremely low) to 9 (extremely high). See Supplementary Materials for further analyses regarding these ratings across the experimental series.
A red circle with a diameter of 359 pixels (RGB values of 192, 80, and 77), a blue square with a size of 359 × 359 pixels (RGB values of 85, 142, and 213), and a yellow triangle with a base and height of 359 pixels (RGB values of 255, 242, and 0) were used as Cues A and B, selecting two of them, counterbalanced across participants. Although only two cues were used here, A and B, the counterbalancing procedure included three different geometrical figures as cues. As in other experiments from our laboratory (see, for example, Morís et al., 2017) that included three cues, we decided not to alter the counterbalancing routine used here as if anything using three cues would make it more likely that any observed effects were independent of the specific geometrical figures used as cues A and B. All visual stimuli were presented against a light grey coloured background (RGB values of 128, 128, and 128) in the centre of the screen. The aversive outcome was a 90 ± 5 dB white noise, presented for 500 ms through headphones (Manufacturer: Sennheiser Electronic GmbH & Co.; Model: HD-201). This kind of high-intensity noise is widely used in human aversive conditioning protocols (Lonsdorf et al., 2017).
Participants were first informed about the study aims and were asked to sign an informed consent document. The experiment then began with the following instructions (translated from the Spanish version): Thank you for participating in this experiment! The task you are about to perform is straightforward. During the experiment, you will see a series of figures. A noise will sometimes follow these figures. Your goal is to predict on each occasion whether the noise will be presented. On each trial, you will first see the figure and, below, you will see a rating scale to enter your predictions. To make your prediction, click on any point of the scale. Click on values close to the right extreme (close to the 100 label) if you think that the noise is very likely to follow. If you think that the noise is unlikely to be presented, click on values close to the left extreme (close to zero). When you click on any point of the rating scale, you will see a marker and a label with the specific value you have chosen. If you want to change this value, just click again on a different part of the scale. Once you have chosen the desired value, you can press ENTER to see what happens and check whether the noise is presented. Pay attention to the figures and to the noise to learn how to make good predictions. If you have any queries now or while you are conducting the task, ask the experimenter for help to make sure that you understand everything. Good luck!
Given that in Experiment 1 we were interested in the spontaneous recovery of threat expectancies, the experimental task was completed in 2 days. After concluding the first two phases of the task, participants returned at the same time on the following day to complete the test phase. This 24-hr interval is routinely used in the literature to promote spontaneous recovery in human threat conditioning (Kindt et al., 2009; Myers et al., 2006; Soeter & Kindt, 2012; van Ast et al., 2013). On Day 2, participants were instructed to continue with the same task as in the previous day, asking them to use what they had learned. In addition, they were reminded of the stimuli, the potential relations between those stimuli, and their task (e.g., predicting whether or not the sound would appear depending on the stimulus present on the trial).
Each trial began with a fixation point (a black cross, Courier New Font, size 180) presented on the centre of the screen for a time interval ranging from 2,500 to 4,700 ms (a uniform distribution with 100 ms step). The cue was then presented along with a rating scale on the lower part of the screen. Participants had to click to give their response following the instructions. Their response was an estimation of the expected occurrence of the aversive outcome, using this measure as the threat CR. The scale went from 0 (the noise is unlikely to be presented) to 100 (the noise is very likely to follow). The cue remained on screen for 3,000 ms after the participant had given their expectancy rating, and thus it overlapped with the last 500 ms during which the outcome was presented. At the end of each trial, both the visual and auditory stimuli disappeared, and the fixation cross of the subsequent trial was presented.
Statistical analyses
For all the experiments in the series, statistical analyses were conducted using IBM SPSS (version 23; IBM Corp., 2015) and R (version 4.0.2), with packages BayesFactor (Morey & Rouder, 2018) and pwr (Champely, 2020). For the three phases (acquisition, extinction, and test), we conducted a repeated-measures analysis of variance (ANOVA), including a between-subject factor (Group: ORE vs. standard extinction) and two within-subject factors (Cue: A vs. B, and Trial). In addition, we conducted another repeated-measures ANOVA for the extinction and test phases but only for Cue A data, including a between-subject factor (Group: ORE vs. standard extinction) and a within-subject factor (trial). The final block of extinction was compared between groups with a Bayesian t-test with default prior scales providing a Jeffreys-Zellner-Siow (JZS) Bayes factor. Return of fear was measured using another repeated-measures ANOVA that included a between-subject factor (Group: ORE vs. standard extinction) and a within-subject factor (Phase: extinction phase vs. test phase), but only for target Cue A. The presence of a significant phase effect would indicate a return of fear effect and a prerequisite for further analyses. A pre-planned t-test was used to compare the responses between the two groups on the first trial of the test phase. Significant differences would be expected on this test if the ORE and standard extinction treatments lead to different levels of return of fear. An additional set of analyses was completed, including the discomfort and anxiety judgements as covariates, finding equivalent results to those described in this article (see Supplementary Materials).
In all the repeated-measures ANOVAs, the sphericity was tested, and the degrees of freedom were corrected using the Greenhouse–Geisser correction, if necessary. The effect size statistics reported are partial eta squared
The data and scripts for all the experiments can be found in the Open Science Framework (OSF) repository (https://osf.io/k9u3q).
Results
Pre-analysis participant rejection
Before the analysis, we followed the same rejection criteria as Morís et al. (2017). We excluded the data of those participants whose mean scores for Cues A and B on the last three trials of the acquisition phase were more than 2 SDs away from the group mean. The same criterion was applied for the last three trials of Cue B from the extinction phase. This selection ensured an adequate understanding of the stimulus contingencies before the experimental manipulation while avoiding the effects of any situation unrelated to the effect of interest. Eleven participants did not meet the criteria and were removed from the final sample, which consisted of 107 participants (56 in the ORE group and 51 in the standard extinction group).
Acquisition phase
A repeated-measures ANOVA was used to analyse the data, including a between-subject factor (Group: ORE vs. standard extinction) and two within-subject factors (Cue: A vs. B and Trial: 1–8). The results revealed a significant effect of Cue, F(1, 105) = 4,222.35, p < .001,
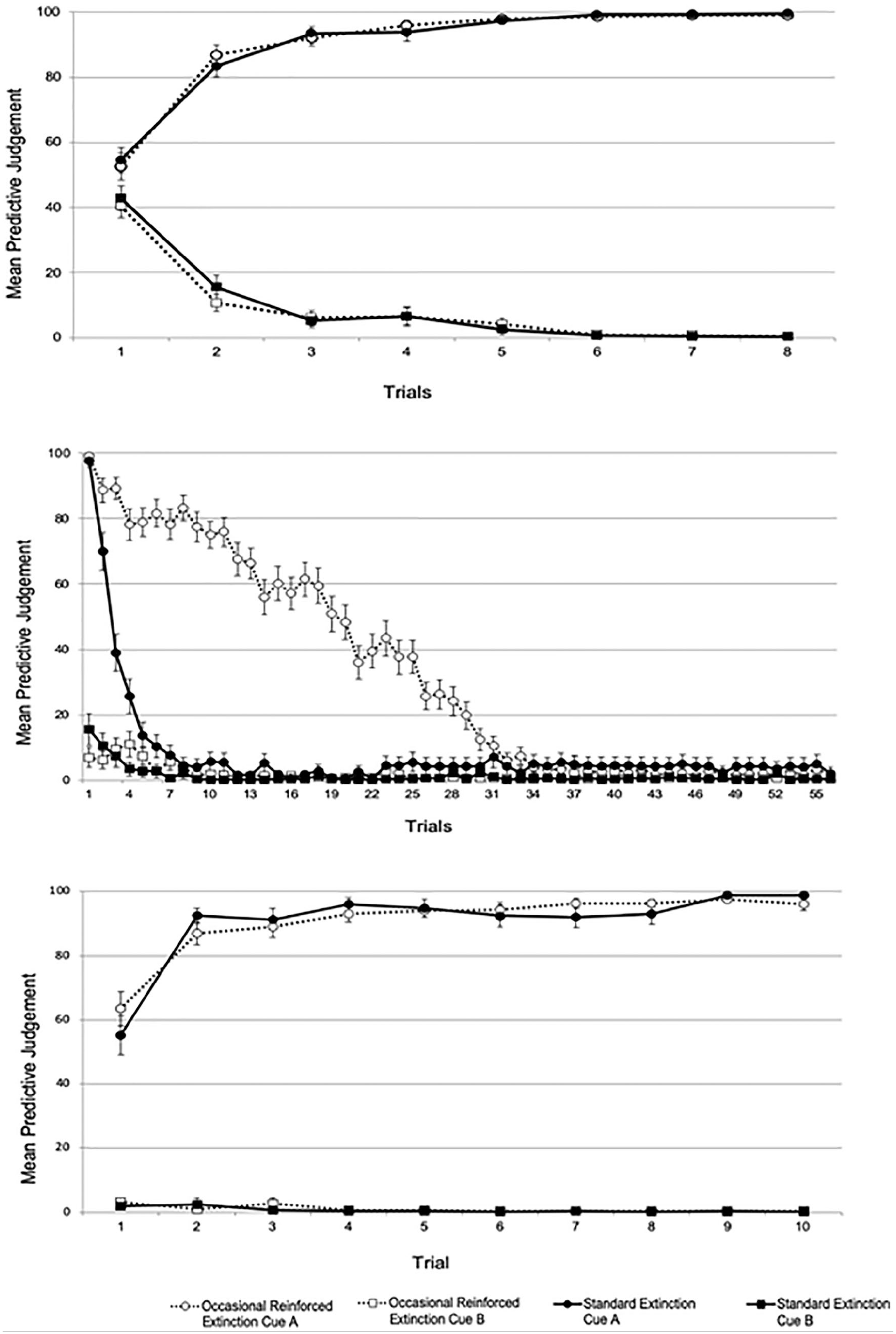
Mean threat expectancies of the acquisition (top), extinction (middle), and test (bottom) phases in Experiment 1.
Extinction phase
An ANOVA was conducted (see Figure 1, middle panel), including a between-subject factor (Group: ORE vs. standard extinction) and two within-subject factors (Cue: A vs. B and Trial: 1–56). This analysis revealed a significant effect of Cue, F(1, 105) = 469.23, p < .001,
Given that the Cue × Trial × Group interaction was significant, we conducted a further repeated-measures ANOVA but only for the data of target Cue A, including a between-subject (Group: ORE vs. standard extinction) and a within-subject factor (Trial: 1–56). We found a main effect of Trial, F(15.56, 1634.2) = 98.93, p < .001,
Test phase
During this phase (see Figure 1, bottom panel), a repeated-measures ANOVA showed a significant effect of Cue, F(1, 105) = 7,516.67, p < .001,
Spontaneous recovery
We analysed the difference between expectancy ratings at the end of the extinction phase (last eight trials for a more reliable measure) and those at the beginning of the test phase (first trial) for Cue A to evaluate the spontaneous recovery effect. For this, we conducted another repeated-measures ANOVA, including a between-subject factor (Group: ORE vs. standard extinction) and a within-subject factor (Phase: extinction phase vs. test phase). We found a main effect of Phase, F(1, 105) = 197.91, p < .001,
Reacquisition
In addition, and following Morís et al. (2017), we conducted another repeated-measures ANOVA but only for the data of target Cue A, including a between-subject (Group: ORE vs. standard extinction) and a within-subject (Trial: 1–10) factor. We found a main effect of Trial, F(3.88, 407.77) = 35.02, p < .001,
US ratings
Participants expressed mild levels of discomfort (M = 4.13, SD = 2.24) and anxiety (M = 3.45, SD = 2.26) when rating the US at the end of the experiment. These ratings indicate that the US was mildly aversive.
Discussion
Expectancy ratings did not differ between the two groups during the acquisition phase. In the extinction phase, we found differences regarding the extinction rate of Cue A, that is, extinction was slower in the ORE than in the standard extinction group. However, at the end of this phase (last block of eight trials), extinction reached asymptotic levels in both groups.
Regarding the first test trial, spontaneous recovery was observed in both groups, in the sense that we detected an increase in threat expectancies relative to the end of the extinction phase. This result is inconsistent with Gershman et al.’s (2013) proposal though consistent with Bouton’s account of the return of fear phenomena.
During the test phase in the ORE and the standard extinction groups, the reacquisition rates did not differ, contrary to the expectations according to Ricker and Bouton’s (1996) trial-signalling mechanism. However, the possible effect of ORE on reacquisition was measured after a 24-hr retention interval. Thus, it is possible that the measure employed to test the prediction derived from the trial-signalling mechanism was insufficiently sensitive and the effect could have been obscured by the spontaneous recovery effect evidenced on the first test trial. A more sensitive test might be obtained if such a retention interval is shortened. This was precisely the objective of Experiment 2.
Experiment 2
In this experiment, the differential effect of the ORE on relapse prevention was measured after a much shorter retention interval of 6 min. It was expected that this would provide a more sensitive measure that would allow us to observe our target differential effect. Again, the final test phase included a series of reinforced CS-US trials to assess the hypothesis derived from the trial-signalling mechanism proposed by Ricker and Bouton (1996).
Method
Participants and apparatus
In total, 173 undergraduate psychology students, 86 from the University of Málaga and 87 from the University of Oviedo, took part in the experiment (140 females). None of the participants had any prior experience of this task. Their participation was voluntary, and they received course credits in exchange for their participation. Visual stimuli were presented on a 21-inch monitor with a 1,920 × 1,080 pixels resolution, while auditory stimuli were presented using individual headphones. The experimental task was written using E-Prime 2.0 (Psychology Software Tools). After the planned rejection of participants (see “Pre-analysis participant rejection” section), this sample provided an 80% probability of detecting an effect of size d = 0.45 for the pre-planned t-test of the first Cue A trial of the test phase.
Design and procedure
The procedure was the same as in Experiment 1, except for two differences. The whole experiment was completed in one session, and the delay between extinction and test was 6 min. Furthermore, to prevent rehearsal of the learned relations, a non-related task, previously used in human learning tasks (Morís et al., 2014), was introduced during the delay. Participants carried out a memory card task in which they had to click on the cards to turn them upside down, trying to pair them correctly to complete the task. If they managed to find a pair, the cards would remain face up; if not, the cards would be face down. When the board was completed, the position of the cards was randomised, and the game started again. This memory task lasted 6 min, after which we evaluated spontaneous recovery in the test phase. Because participants had to concentrate on the card game, which was cognitively demanding, this promoted a temporal context change from the extinction to the test phases to evaluate the spontaneous recovery of the threat expectancies within our two groups.
Results
Pre-analysis participant rejection
The same rejection criteria described in Experiment 1 were used. Therefore, 16 participants did not meet the criteria and were removed from the final sample, which consisted of 157 participants (77 in the ORE group and 80 in the standard extinction group).
Acquisition phase
A repeated-measures ANOVA was used to analyse the data, including a between-subject factor (Group: ORE vs. standard extinction) and two within-subject factors (Cue: A vs. B and Trial: 1–8). This revealed significant effects of Cue, F(1, 155) = 10,974.92, p < .001,
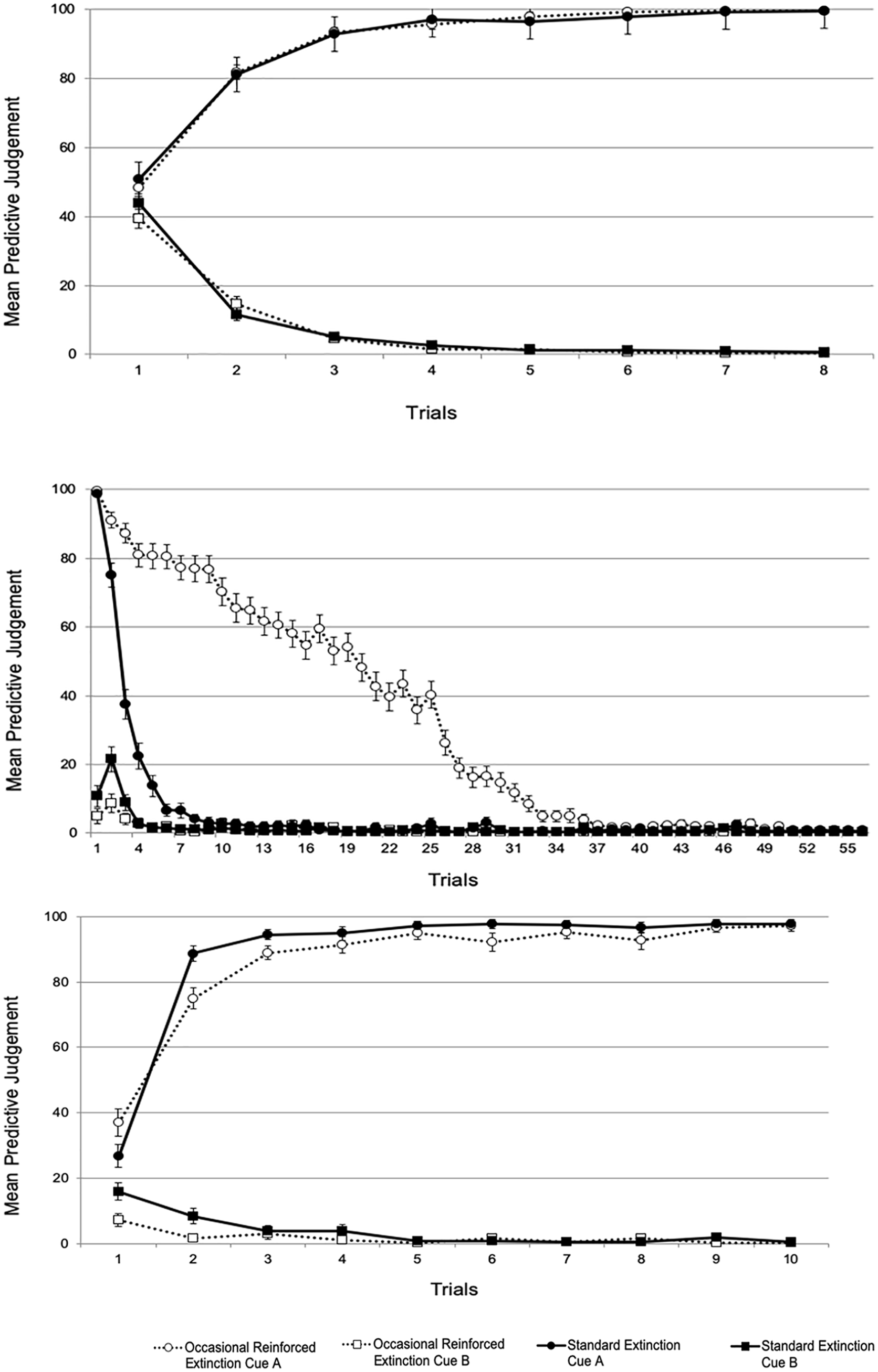
Mean threat expectancies for the acquisition (top), extinction (middle), and test (bottom) phases in Experiment 2.
Extinction phase
A similar ANOVA was used (see Figure 2, middle panel), including a between-subject factor (Group: ORE vs. standard extinction) and two within-subject factors (Cue: A vs. B and Trial: 1–56). This revealed a significant effect of Cue, F(1, 155) = 1,178, p < .001,
Once we had conducted this general analysis, and because the Cue × Trial × Group interaction yielded a significant effect, we carried out a further repeated-measures ANOVA but only on the data for target Cue A, including a between-subject factor (Group: ORE vs. standard extinction) and a within-subject factor (Trial: 1–56). We found an effect of Trial, F(16.53, 2562.72) = 231.65, p < .001,
Test phase
During this phase (see Figure 2, bottom panel), the repeated-measures ANOVA, as before, showed significant effects of Cue, F(1, 155) = 9,255.26, p < .001,
Spontaneous recovery
We analysed the difference between threat expectancies for Cue A at the end of the extinction phase (last eight trials, again, for a more reliable measure) and those at the beginning of the test phase (first trial) to evaluate the spontaneous recovery effect. For this, we carried out another repeated-measures ANOVA, including a between-subject factor (Group: ORE vs. standard extinction) and a within-subject factor (Phase: extinction phase vs. test phase). This analysis revealed a main effect of Phase, F(1, 155) = 129.03, p < .001,
Reacquisition
In addition, and again following Morís et al. (2017), we carried out another repeated-measures ANOVA but only for the data of target Cue A, including a between-subject (Group: ORE vs. standard extinction) and a within-subject (Trial: 1–10) factor. We found a main effect of Trial, F(5.14, 796.21) = 192, p < .001,
US ratings
Participants expressed mild levels of discomfort (M = 5.1, SD = 2.42) and anxiety (M = 4.17, SD = 2.38) when rating the US at the end of the experiment. Again, these ratings indicate the mild aversiveness of the US used.
Discussion
Expectancy ratings during the acquisition phase did not differ between the two groups. In the extinction phase, ratings for Cue A extinguished at a slower pace in the ORE than in the standard extinction group. However, at the end of this phase (last block of eight trials), extinction reached asymptotic levels in both groups.
Regarding the first test trial, spontaneous recovery was observed in both groups, shown by an increase in threat expectancies relative to the end of the extinction phase. This finding is again inconsistent with Gershman et al.’s (2013) proposal.
Unlike in Experiment 1, the rate of reacquisition during the test differed between the ORE and the standard extinction group, consistent with Ricker and Bouton’s (1996) trial-signalling mechanism. The shorter retention interval used here to promote spontaneous recovery offered a more sensitive test to evaluate this hypothesis, and reacquisition was found to proceed more slowly in the ORE than in the standard extinction group.
Experiment 3
The previous results suggest that an ORE treatment could effectively reduce the rate of reacquisition, in spite of being ineffective in producing a relapse prevention effect after a retention interval. In this experiment, we intended to assess the potential effect of this occasional reinforcement treatment on another form of relapse, namely, reinstatement. More specifically, our objective was to evaluate the possible differential effect on relapse prevention of ORE and standard extinction training after experiencing non-signalled presentations of the US after extinction had been completed. In this sense, we are not so much interested in reinstatement itself, but in this differential recovery effect produced by ORE and standard extinction. For convenience, we will refer to this differential effect as reinstatement. In addition, during the test, after presenting a first reinforced trial, the remaining test trials were non-reinforced. This re-extinction allows us to evaluate whether the signalling mechanism (Ricker & Bouton, 1996) also interferes with the reacquisition of extinction in the ORE group.
Method
Participants and apparatus
In total, 84 undergraduate psychology students from the University of Málaga completed the task (72 females). Participants had no previous experience with the task. Their participation was voluntary, and they received course credit in exchange. Visual stimuli were presented on a 21-inch monitor with a 1,920 × 1,080 pixels resolution, while auditory stimuli were presented using individual headphones. The experimental program was written using E-Prime 2.0 (Psychology Software Tools). After the planned rejection of participants (see “Pre-analysis participant rejection” section), this sample provided an 80% probability of detecting an effect of size d = 0.65 for the pre-planned t-test of the first Cue A trial of the test phase.
Design and procedure
The design of this experiment was the same as that used for Experiments 1 and 2, with the following exceptions. As the discomfort judgements of the aversive sound in Experiments 1 and 2 were below (M = 4.13, SD = 2.24) and barely above (M = 5.1, SD = 2.42) the midpoint of the rating scale, respectively, we decided to change the outcome to a 3-s beep of 97 ± 3 dB, which has previously been used in our laboratory (Flores et al., 2018). The acquisition phase was separated from the rest of the phases by a time interval of 24 hr. At the beginning of the second day, participants were instructed to continue with the same task as in the previous day, asking them to use what they had learned. In addition, they were reminded of the stimuli, the potential relations between these stimuli, and their task (e.g., predicting whether or not the sound would appear depending on the stimulus presented on the trial).
The relationships programmed during the acquisition phase were probabilistic, and the probability of reinforcement of Cue A was 87.5%. In Experiments 1 and 2, participants encountered probabilistic relationships in the ORE group but not in the standard extinction group. By including this change, probabilistic relationships were experienced in both groups (see Morís et al., 2017, for a similar strategy).
In addition, as shown in Table 1 (Experiment 3), after the extinction phase, participants were exposed to three 1-s unsignalled presentations of the US to evaluate the reinstatement effect during the test phase. The intertrial interval between these presentations varied randomly from 2,500 to 4,700 ms.
Regarding the test phase, after a first reinforced trial, the rest of this phase included non-reinforced trials (i.e., a re-extinction phase; see Table 1). As described in the “Introduction” section, it is also possible to evaluate the predictions derived from the operation of the trial-signalling mechanism. Ratings regarding the discomfort and anxiety caused by the aversive sound were requested twice (after the acquisition phase and after the test phase). Here, we report only the latter ratings for comparability with previous experiments.
Results
Pre-analysis participant rejection
As in the previous experiments and before the analysis, we followed the same participant selection criteria as Morís et al. (2017). Nine participants did not meet the criteria and were excluded from the final sample, which consisted of 75 participants (38 in the ORE group and 37 in the standard extinction group).
Acquisition phase
A repeated-measures ANOVA was used to analyse the data, including a between-subject factor (Group: ORE vs. standard extinction) and two within-subject factors (Cue: A vs. B and Trial: 1–8). The results revealed a significant effect of Cue, F(1, 73) = 1,767.73, p < .001,
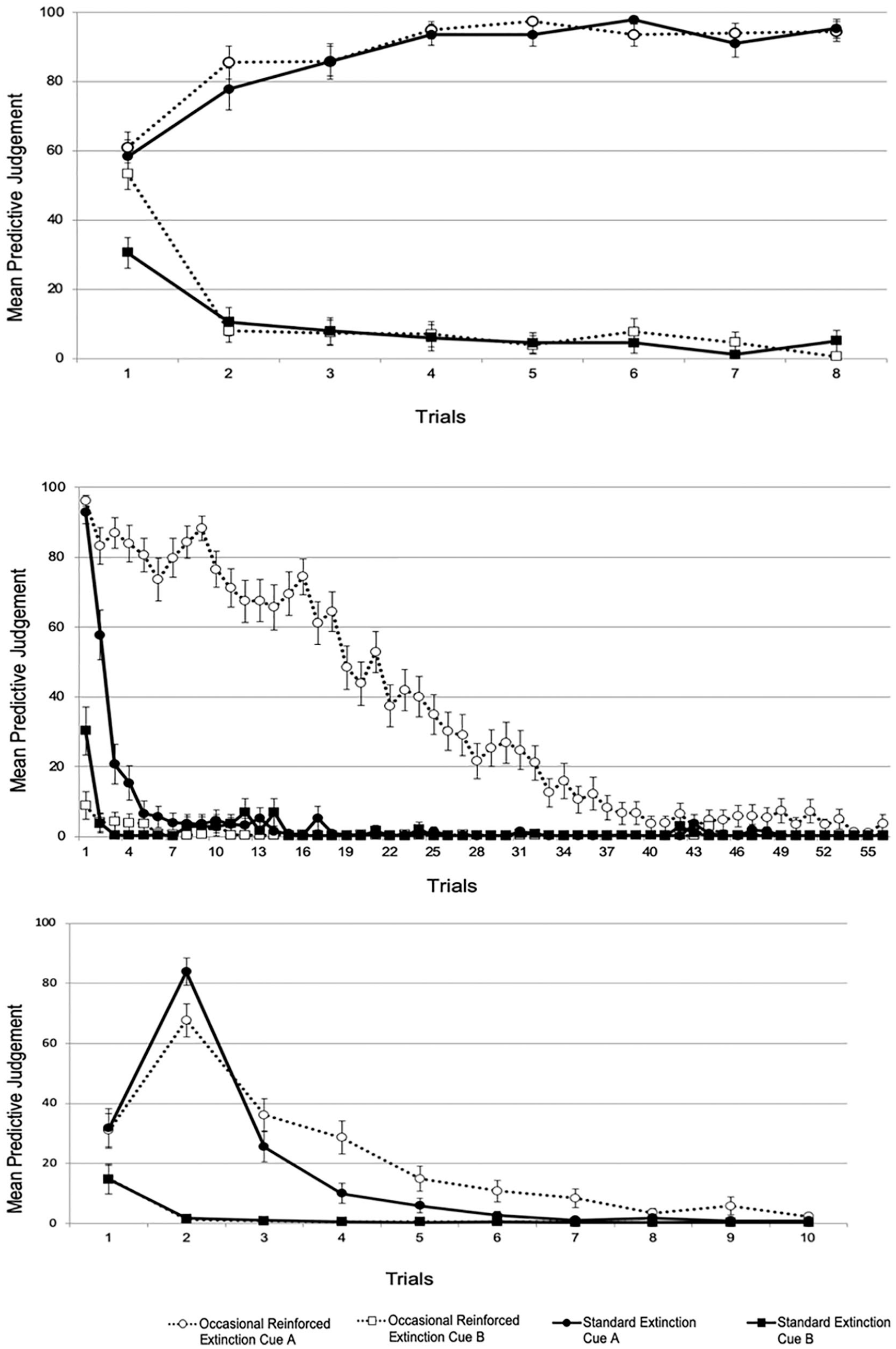
Mean predictive judgements of the acquisition (top), extinction (middle), and test (bottom) phases in Experiment 3.
Extinction phase
A similar ANOVA was used (see Figure 3, middle panel), including a between-subject factor (Group: ORE vs. standard extinction) and two within-subject factors (Cue: A vs. B and Trial: 1–56). This analysis revealed a significant effect of Cue, F(1, 73) = 251.1, p < .001,
As the Cue × Trial × Group interaction yielded a significant effect, we conducted a further repeated-measures ANOVA but only for the data of target Cue A, including a between-subject factor (Group: ORE vs. standard extinction) and a within-subject factor (Trial: 1–56). We found a main effect of Trial, F(15.56, 1136.05) = 80.23, p < .001,
As in the previous experiments, this last interaction showed that extinction proceeded more slowly in the ORE group. At the end of the phase (last eight-trial block), mean expectancy ratings were M = 4.12, SD = 10.08 in the ORE group and M = 0.28, SD = 0.17 in the standard extinction group. The Bayesian t-test did not show clear support for either the null hypothesis or the alternative hypothesis (BF01 = 0.4), because the means of both groups differed by less than 4 points (on a scale of 100 points). Given this observation, and the results of the previous experiments, extinction is assumed to have reached asymptotic levels in both cases.
Test phase
During this phase (see Figure 3, bottom panel), the repeated-measures ANOVA revealed a significant effect of Cue, F(1, 73) = 142.56, p < .001,
Reinstatement
We analysed the difference between expectancy ratings at the end of the extinction phase (last eight trials) and those at the beginning of the test phase (first trial) for Cue A to evaluate the reinstatement effect. For this, we conducted a repeated-measures ANOVA, including a between-subject factor (Group: ORE vs. standard extinction) and a within-subject factor (Phase: extinction phase vs. test phase). We found a main effect of Phase, F(1, 73) = 42.97, p < .001,
Re-extinction
Using a re-extinction test allowed us to examine whether there were differences between groups in terms of how extinction was reacquired after the first reinforced test trial. To this end, we carried out a repeated-measures ANOVA, including a between-subject factor (Group: ORE vs. standard extinction) and a within-subject factor (Trial: 1–10) for target Cue A ratings. This analysis revealed a main effect of Trial, F(3.97, 289.42) = 87.18, p < .001,
US ratings
Participants expressed mild levels of discomfort (M = 5.88, SD = 2.56) and anxiety (M = 4.85, SD = 2.43) when rating the US at the end of the experiment. Again, these ratings indicate the mild aversiveness of the US used.
Discussion
As in the previous experiments, no group differences were found in the acquisition phase. In the extinction phase, we found differences in the extinction of the target Cue A, mainly due to the treatment given to each group (occasional reinforced or standard extinction). Nevertheless, in both groups, at the end of the phase, expectancy ratings converged to asymptote. Finally, on the first trial of the test phase, we observed an equivalent reinstatement of the cognitive component of the fear response in both groups in the sense that threat expectancy ratings in both groups were of a higher magnitude than at the end of the extinction phase. Again, this finding is inconsistent with the account proposed by Gershman et al. (2013). Regarding the re-extinction rate, and in line with the reacquisition results of Experiment 2, we found that the ORE group showed a slower rate of re-extinction (i.e., needed more trials to reach asymptotic levels) than the standard extinction group, consistent with the trial-signalling mechanism proposed by Ricker and Bouton (1996).
Bayesian meta-analysis of the first trial of the test phase
In the three experiments reported, the judgements of the ORE and control groups did not differ significantly on the first trial of the test phase, contrary to the prediction of Gershman et al. (2013). As described in Rouder and Morey (2011), we carried out a Bayesian meta-analysis using the BayesFactor R package (version 0.9.12-4.2, Morey & Rouder, 2018). This analysis integrates the information provided by a series of t-tests, considering the sample size of each of them. This results in a Bayes factor, showing support for the null hypothesis or the alternative hypothesis. In this case, the null hypothesis was a point-null hypothesis,
Finally, we carried out a robustness check of the analysis, using priors with width parameters ranging from 0.01 to 1.5 (Figure 4). In all the cases, the Bayes factors were smaller than 1. When the scale parameter was 0.07 or higher, the Bayes factors were smaller than 1/3, showing adequate support for the null hypothesis.
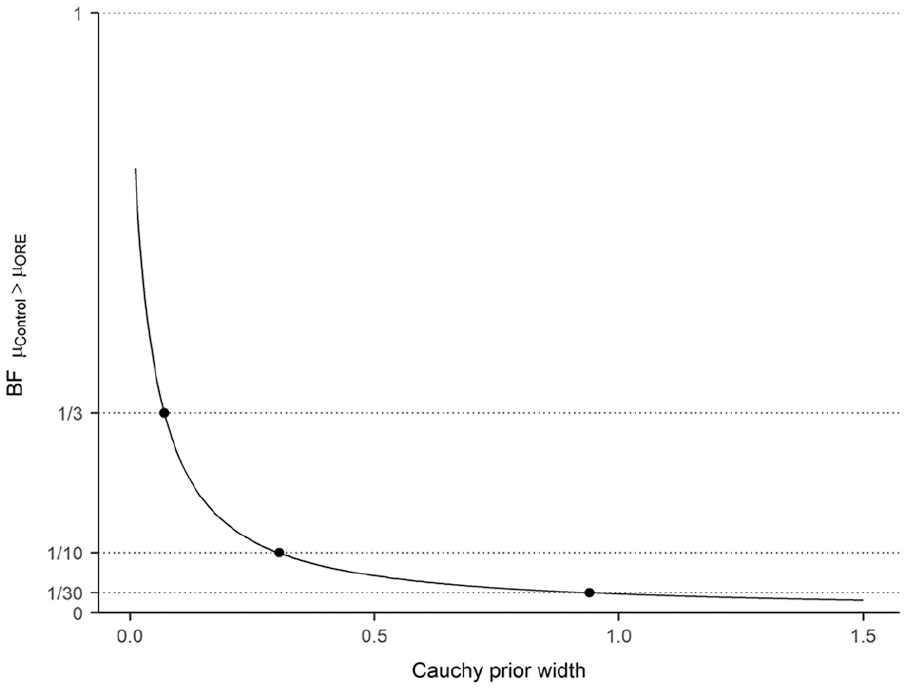
Results of the Bayesian meta-analysis of the results of the first trial of the test phase in the three experiments, depending on the width of the prior used.
General discussion
The occasional inclusion of reinforced trials during extinction has been suggested as an effective strategy for enhancing inhibitory learning and retrieval and thus for reducing relapse effects (see, for example, Craske et al., 2014, 2018). Previous animal conditioning studies have shown that gradually reducing the number of US presentations during extinction training can prevent the spontaneous recovery and reinstatement of fear responses (see Gershman et al., 2013). Bouton et al. (2004) and Woods and Bouton (2007) found that ORE can also slow down the rate at which the later reacquisition of conditioning occurs after extinction training. Morís et al. (2017) extended this preventive effect on rapid reacquisition to a human threat conditioning preparation. Specifically, Morís et al.’s Experiment 3 measured the impact of gradual ORE on threat expectancies, a cognitive component of the fear response. Although there have been previous attempts to assess the preventive effects of ORE on spontaneous recovery and reinstatement, the results obtained are far from conclusive. Thus, the present experimental series tried to extend further this promising result obtained in human threat conditioning to spontaneous recovery and reinstatement effects, as obtained initially by Gershman et al. (2013) in their animal study. Experiments 1 and 2 (after a 24-hr and a 6-min time interval, respectively) assessed the preventive effects of gradual ORE on spontaneous recovery, while Experiment 3 assessed the effect on the reinstatement of the CR. The three experiments compared participants’ threat expectancies in the two extinction treatment groups, the gradual ORE group, and the standard extinction group. The whole experimental series demonstrated similar rates of acquisition of conditioning in the two groups and a slower rate of extinction in the ORE group. However, in all three experiments, extinction reached comparable asymptotic levels in both extinction treatment groups.
Experiments 1 and 2 showed spontaneous recovery during the final test phase in both groups. Note that here spontaneous recovery is understood to have occurred when threat expectancies on the first test trial are of a higher magnitude than those observed at the end of the previous extinction phase. Despite the short temporal delay between extinction training and the test phase used in Experiment 2, the same pattern of results was observed, with a reduced magnitude of the spontaneous recovery effect when compared with the results on the first test trial of Experiment 1. Experiment 3 also showed a significant reinstatement effect in both extinction groups. Reinstatement is assumed to have occurred if threat expectancies on the first test trial are of a higher magnitude than those observed at the end of the previous extinction phase. The Bayesian meta-analysis showed support for the null hypothesis, an absence of differences between the ORE and the control groups on this first trial of the test phase. Taken together, the results from this test trial showed that gradual ORE did not prevent spontaneous recovery or the reinstatement of threat expectancies in mildly aversive conditions. Our results also speak to the effects of ORE on the reacquisition and re-extinction of the threat expectancies during the test phase once participants had experienced the aversive stimulation on the first test trial. Experiments 2 and 3 revealed slower reacquisition of the relationship programmed in the ORE than in the standard extinction group. While in Experiment 2, the ORE treatment slowed down the reacquisition of conditioning, in Experiment 3, it slowed down re-extinction of conditioning. Experiment 1 revealed that the reacquisition of conditioning during the test was equivalent in both groups. The more significant spontaneous recovery experienced in this experiment after a 24-hr retention interval meant that responding was already at a high level at the beginning of the test, making it a less sensitive test for detecting ORE effects on reacquisition. The results found in Experiment 2, after using a much shorter (6-min) retention interval, support the viability of this explanation.
Overall, the set of results reported here have not provided evidence to demonstrate the preventive effects of ORE on the spontaneous recovery or reinstatement of threat expectancies. In all three experiments, threat expectancies recovered spontaneously or were reinstated, as measured on the first test trial. Unlike previous attempts to assess the preventive effects of ORE on return of fear phenomena (Culver et al., 2018; Shiban et al., 2015; Thompson et al., 2018), our procedure was able to establish asymptotic performance at the end of extinction training, both in the ORE and in the standard extinction groups. In addition, our design ensured that the possible preventive effects of ORE on spontaneous recovery and reinstatement were assessed independently from each other so that assessing one did not condition or interfere with the assessment of the other. These conditions may be regarded as necessary requirements for adequately assessing the utility of the ORE strategy for preventing specific relapse phenomena.
The results reported here also have theoretical implications. First, these findings are not consistent with the general idea that gradual ORE weakens the original fear memory, as shown by the relapse phenomena obtained in all three experiments. Contrary to the predictions of Gershman et al.’s (2013) account, the cognitive component of the fear memory (as measured by threat expectancies) did not seem to lose its strength as a consequence of gradual ORE (see also Shiban et al., 2015; Thompson et al., 2018). This result is in contrast with those reported by Gershman et al. (2013) in animal studies. Although it is possible that the preventive effects of ORE do not extend to human participants, this could be an overly premature conclusion. There are several ways in which our procedure can be distinguished from that used by Gershman et al. (2013), all of which could provide an alternative explanation for the discrepant results. For example, while we used mild aversive sounds as US, Gershman et al. used electric shocks. Another procedural difference with more potential theoretical interest is the number of acquisition trials included in the different studies. While Gershman et al. (2013) included three acquisition trials in both experiments, our participants experienced eight trials during the acquisition phase in all three experiments. Gershman et al.’s account assumes that the preventive effects of gradual ORE depend on an effective modification of the original fear memory. This effective modification will only occur if extinction training involves a prediction error (i.e., a discrepancy between the predicted outcome—the US—and the experienced outcome—no US). However, this prediction error should be large enough to drive new learning but not so large that it can be taken as a segmentation signal, and thus, a new context-dependent inhibitory memory is formed during extinction. Any standard associative learning theory would assume that the fear memory formed during acquisition would be gradually strengthened as the number of CS-US pairings is increased across trials. Thus, gradual ORE will entail a more significant prediction error after eight trials than only three acquisition trials. In other words, it would be expected that gradual ORE will entail a more significant prediction error in our procedure than in the procedure of Gershman et al. (2013). It might then be argued that gradual ORE involved the formation of a new context-dependent inhibitory memory in our procedure but not in the animal study of Gershman et al. This might explain why we could not find any preventive effects of gradual ORE on spontaneous recovery and reinstatement. For this reason, the number of acquisition trials may be a boundary condition for ORE to show its preventive effects on relapse phenomena.
Moreover, this could also be regarded as a highly relevant limitation of the potential translational value of ORE in the clinical domain. Anxiety disorders involve strong forms of fear in patients. Thus, prediction errors derived from the gradual exposure to those feared stimuli (as in gradual ORE) would also serve as a segmentation signal, indicating a novel state in the environment and, consequently, demanding the formation of a new context-dependent memory. Future studies should shed light on the number of acquisition trials needed for gradual ORE to have a preventive effect.
On the contrary, the set of results reported here is consistent with Bouton and collaborators’ trial-signalling mechanism. First, we have demonstrated spontaneous recovery and reinstatement of threat expectancies as anticipated by their account. Furthermore, during the test phase of Experiments 2 and 3, we obtained differences between the rate of acquisition of the CS-US contingencies (i.e., either reacquisition or re-extinction) in the gradual ORE and standard extinction groups. Even the lack of differences in the reacquisition rate between the two groups in Experiment 1 could be interpreted as consistent with Ricker and Bouton’s (1996) trial-signalling mechanism. After a 24-hr retention interval between the extinction phase and the test phase, the spontaneous recovery of threat expectancies that were obtained could have limited the sensitivity of the test for revealing group differences in the rate of reacquisition. Reducing this retention interval to 6 min was sufficient to obtain the target difference in the reacquisition rate between the two groups. In other words, measuring the significant spontaneous recovery effect before reacquisition might have weakened the retrievability of the extinction memory in the ORE group, and, therefore, no effect on reacquisition was found. Only when the retention interval was shortened, and spontaneous recovery reduced, could the group difference in reacquisition during the test be detected.
Some limitations of our study should be noted. Our focus of interest was limited to threat expectancy ratings. However, the emotional and physiological activation dimensions of the fear response represent a relevant scenario in which the effect of gradual ORE also needs to be measured. Unlike Shiban et al.’s (2015) procedure in which gradual ORE was also evaluated, increasing the length of extinction training as in the design of the present experiments would be a more effective way of assessing ORE effects on these additional components of the fear response. Future research using these other measures will allow us to determine more precisely the merits of using gradual ORE for preventing the return of fear phenomena. Nevertheless, the potential translational interest of ORE for preventing relapse will depend on the ability to demonstrate its preventive effects on the different components of the fear response, given their interconnectedness.
Concluding comments
In a series of three experiments, we aimed to evaluate whether a gradual ORE treatment could prevent spontaneous recovery and reinstatement effects. Our findings did not provide evidence to suggest that an ORE treatment could be effective in mitigating relapse. From a theoretical standpoint, our results are more consistent with the idea that extinction entails the acquisition of new knowledge (as proposed by Bouton, 2004) than with the idea that there are conditions in which extinction leads to a weakening of the original fear memory (Gershman et al., 2013). This has important implications for the use of this technique. In particular, the potential benefit of using ORE to mitigate return of fear phenomena seems to be more limited than previously anticipated, and a more thorough investigation of this procedure is therefore needed before it can be implemented in clinical settings.
Supplemental Material
sj-docx-1-qjp-10.1177_17470218211043434 – Supplemental material for Limitations of occasional reinforced extinction to alleviate spontaneous recovery and reinstatement effects: Evidence for a trial-signalling mechanism
Supplemental material, sj-docx-1-qjp-10.1177_17470218211043434 for Limitations of occasional reinforced extinction to alleviate spontaneous recovery and reinstatement effects: Evidence for a trial-signalling mechanism by María J Quintero, Amanda Flores, María T Gutiérrez-Huerta, Patricia Molina-Guerrero, Francisco J López and Joaquín Morís in Quarterly Journal of Experimental Psychology
Footnotes
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by grants UMA18-FEDERJA-051 from Junta de Andalucía and UMA FC14-SEJ-332014 from the University of Málaga, and grant PGC2018-096863-B-I00 from the Spanish Ministry of Science, Innovation, and Universities. María José Quintero has been awarded a PhD fellowship from the Spanish Ministry of Science, Innovation, and Universities (Programa de Formación de Profesorado Universitario, FPU18/00917).
Data accessibility statement
Notes
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.