
Abstract
Cal Izard has provided psychology a robust vision of human emotional feelings. He has addressed the full spectrum of emotional-developmental-cognitive complexities entailed in clarifying seemingly impenetrable mysteries: How do we experience emotions and how do they guide cognitive development? Izard’s developmental studies of infant minds integrate the primal evolutionary affective foundations of our nature with the diverse paths of nurture, and are framed in ways that can promote human thriving. His multilayered vision of our emotional nature resonates well with modern cross-species affective neuroscience perspectives.
Cal Izard’s studies of human emotions were initiated long before his scientific findings could be integrated with the weight of modern neuroscientific evidence. His life’s work has provided substantive empirical support to evolutionary perspectives of our emotional nature, and is consistent with cross-species affective neuroscientific perspectives (Panksepp, 1998; Panksepp & Biven, 2012). His work ranged from differential emotions theory and research on facial expressions (Izard, 1971, 1990, 1992, 1994), to the role of emotions in the developmental construction of higher mental functions as well as disruptive emotional disorders (Izard, 2009). The practical side of his work promoted emotionally sensitive education of children, providing solid developmental guideposts for healthy living (Izard, Stark, Trentacosta, & Schultz, 2008; Trentacosta & Izard, 2007) and optimal personality development (Abe & Izard, 1999).
I first became conversant with Izard’s works long after my own affective neuroscience project was well underway (Panksepp, 1982, 1998). My goal was to establish an empirically sound cross-species neuroscientific bottom-up understanding of primary-process mammalian emotional networks, with inroads to the raw affective experiences of animals (Figure 1). My project was resonant with Cal’s, including his quest for practical knowledge that could benefit emotional health and promote new interventions (Izard, 2002; Izard et al., 2008; Panksepp, 2004, 2006; Panksepp, Wright, Döbrössy, Schlaepfer, & Coenen, 2014).
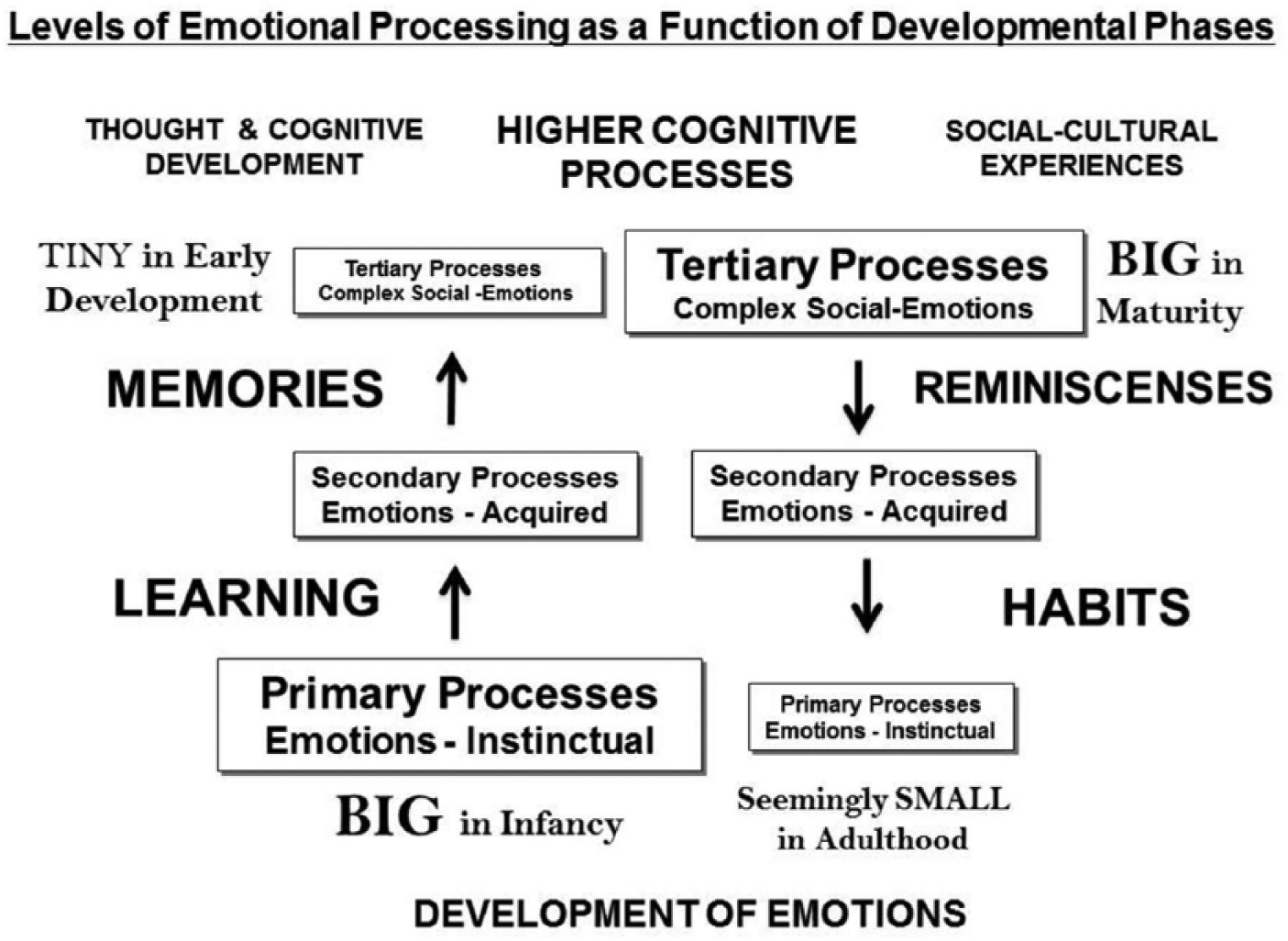
Conceptual diagram summarizing levels of control within babies’ BrainMind (left), where Izardian instinctual primary-process emotional responses are very prominent, and higher mental processes are undeveloped, since they are almost completely programmed by life experiences, operating via neural mechanisms of learning and memory. Developmental landscapes (left) must be contrasted with MindBrain organization in adults (right), where higher mental (tertiary) processes are well developed, which regulate primary processes, and parse raw emotions into greater varieties depending on personal and cultural learning. Although primal emotions are cognitively integrated in adult brains, they continue to influence mental life and sustain consciousness, with a return to more primal, brainstem states during dreaming (Solms & Panksepp, 2012).
Of the many paths to understanding human emotions, my project has been remarkably synergistic with Izard’s (1993, 2009) naturalistic approaches. And both of us favored a multitiered approach to emotions (Figure 1) that respected the ways our fundamental (“basic” or “primary-process”) emotions were foundational for the construction of more complex emotional schemas. We sought consilience with investigators whose approaches could not illuminate what evolution had provided for the construction of the human mental apparatus (e.g., Izard, 2007; Panksepp, 2007). In addition, I faced the persistent challenge of dealing with “never-mind neuro-behaviorism” (Panksepp, 1990) where discussion of emotional feeling in animals has remained taboo to the present day (see LeDoux, 2012).
Izard’s projects, as well as my own, arose from Darwinian convictions: We are mammals, with dramatic brain emotional homologies with other mammals (Izard, 2009; Panksepp, 1982, 1998). We all agree how special humans are in terms of cognitive development, allowing for remarkable upper cerebral abilities, from deep thinking to complex feelings. But those developmental/epigenetic cortical achievements are built upon cross-mammalian subcortical, primary-process, affective homologies. Our vast cortico-cognitive competences are grounded upon ancestral affective foundations, allowing massive diversification, regulation, and repression of subcortical-affective powers of mind (for overviews of top-down influences, see Liotti & Panksepp, 2004; Northoff, 2014). Why we both needed to engage those holding ultraconstructivist views of human nature remains puzzling to both of us (see Izard, 1992, 2007; Panksepp, 1992, 2007); with regard to the construction of higher mental functions (Figure 1, right), we have both been ultraconstructivists.
In current academic psychology, with its continuing modern love affairs with evolutionary psychology and human brain imaging, it is often forgotten that at birth, our massive neocortices resemble tabula rasa—a blank slate, rather than a beehive of evolved modules—where developmental experiences construct diverse higher mental functions. Within the neocortex, even refined vision is learned (Sur & Rubinstein, 2005). In infancy, evolved subcortical functions conspire with life experiences to guide the development of upper mental functions more than any demonstrably evolved neuro-cognitive, modular specializations: for example, we have strong emotional communicative urges, but perhaps no “language instinct.” Still, the primal affective powers of the mind are more genetically endowed than any of our higher cognitive abilities and illuminable by studying brains of other animals. In proposing interest as a basic emotion, Izard (1977) anticipated and appreciated the nature of the long neglected SEEKING system for mediating human enthusiasm (Panksepp, 1992, 1998). Behavioral neuroscientists still prefer to call this massive general-purpose mammalian emotional/motivational network “the brain reward system,” despite clear evidence it mediates foraging enthusiasm rather than sensory pleasures, contributing much to our mammalian LUST, CARE, and PLAY urges, and our artistic/scholarly aspirations and many addictions (Alcaro & Panksepp, 2007, 2011; Ikemoto & Panksepp, 1999).
During the modern era of psychology, with cognitive approaches prevailing in human research, few have embraced the good news, as did Izard (1993, 2009), that work on other mammals offers evidence-based scientific glimmers of the actual neurobiological constitution of primal affective experiences, indeed consciousness itself (Panksepp, 2011a, 2011b; Solms & Panksepp, 2012). Although animal research has a major role to play in understanding primal human affective feelings (a view yet to be embraced by academic psychology), Izard recognized how cross-species neuroscientific approaches could illuminate basic human emotions, with abundant implications for understanding psychiatric problems (Izard, 2009). To appreciate why full integration has not been achieved (Figure 1), it may be useful to share a brief historical retrospective.
A Synopsis of the Scientific History of Our Field (With Minimal Referencing)
In the Expression of Emotions in Man and Animals (1872), Darwin described with grace, depth, and clarity many of the “instinctual” bodily, behavioral, facial, and physiological processes that relate to states of emotional arousal in humans and other animals. In line with Darwin’s insights, modern basic emotion theory in humans arose from such work. Cal Izard and Paul Ekman, both inspired by Silvan Tomkins, analyzed diverse emotional displays in humans which are congruent with bottom-up primary-process cross-species affective neuroscience analyses (Figure 1, left side). Although human cultural learning readily modifies emotional displays (Figure 1, right side), that does not negate the nature of our primary-process emotionality. Of course basic emotion approaches work best on foundational brain/mind issues (Panksepp, 1982, 1998; Panksepp & Biven, 2012). The fact that cross-species primary-process affective neuroscience findings converge with facial-emotional studies of humans, formulated independently of each other, affirms the existence of a diversity of Darwinian primary-process emotional systems across mammalian species. Although William James offered an alternative neocortical read-out account of human emotions, that still captivates psychology (Figure 1, right side), let’s remember that he also noted in his Principles that many instincts are accompanied by affective feelings (Figure 1, left side), leaving open the door for a cross-species, primary-process affective science.
Indeed, the social psychologist, William McDougall, more explicitly envisioned how human motives and personalities were strongly related to primal emotional urges shared by all mammals. His list included anger, disgust, fear, wonder (interest and curiosity), subjection (subordination), elation (dominance), as well as gregarious instincts accompanied by tender emotions (caring for young), the absence/loss of which promotes separation distress. McDougall’s list is remarkably congruent with the primary-process emotional networks revealed by affective neuroscience—primal RAGE, FEAR, SEEKING along with specific social urges (PLAY, LUST, CARE, and PANIC)—discovered with direct brain stimulation (DBS), highlighting that primal emotions arise from subcortical brain networks shared homologously across mammals (Panksepp, 1998), a conclusion mirroring Izard’s work with human infants.
Walter Cannon (1927) offered a robust critique of the highly speculative James–Lange cortico-centric theory of emotional feelings and suggested that emotional feelings arise from fundamental subcortical brain processes, focusing on thalamic contributions (indeed PANIC and PLAY rely heavily on trans-thalamic systems). Cannon’s conjecture, theoretically elaborated by James Papez (1937), led to the recognition of a subcortical locus of emotionality that came to be known as the limbic system—a step in the right direction, that is periodically criticized (LeDoux, 2012).
Concurrently, in Nobel Prize worthy work, physiologist Walter Hess (who received the Nobel Prize for his work in 1949) mapped the autonomic nervous system with DBS. He inadvertently discovered that activation of sympathetic hypothalamic regions yields rage displays in cats. Regrettably, he labeled his seminal discovery “sham rage” so his work would not be marginalized by the powerful Anglo-American behaviorists. He regretted his timidity in autobiographical reflections—namely not elaborating his private conviction that his cats actually experienced anger. Panksepp (1971) demonstrated such circuits generated aversive states that animals escaped when given opportunities—the gold standard for negative internal feelings in nonspeaking animals (Panksepp, 1982, 1998).
Then it was discovered that the “ascending reticular activating system” (ARAS) in the brainstem was critical for consciousness (for summary, see Solms & Panksepp, 2012). While seeking to stimulate those ARAS brain areas in ongoing DBS studies, Olds and Milner (1954) at McGill University, accidentally discovered rewarding brain sites in rats. Concurrently, Neal Miller and colleagues (Delgado, Roberts, & Miller, 1954) at Yale identified punishing sites in nearby hypothalamic sites. The existence of various rewarding and punishing states evoked by DBS of the various brain emotional systems is now recognized as a critical (necessary and sufficient) criterion for the existence of positive and negative affective states in the brains (may we say minds?) of other animals. This is validated by the fact that DBS of homologous brain regions in humans evokes related affective states in humans (Panksepp, 1985).
Independent of the aforementioned advances, Silvan Tomkins, a clinical psychologist interested in human psychiatric problems, postulated eight basic emotions in the early 1960s, namely anguish, disgust, fear, interest, joy, rage, shame, and surprise. Paul Ekman and Cal Izard, both inspired by Tomkins’s clinical insights, advanced basic emotion theory into the empirical realm through the study of adult and infant human facial emotion displays. Ever since, there have ensued heated arguments about “What’s Basic About Basic emotions?” (Ortony & Turner, 1990), commonly carried on independently of the robust cross-species neuroscientific evidence for emotional reactions evoked by DBS in animals.
Concurrently, Panksepp (1982), pursuant to the brain-reward/punishment DBS effects described before, provided evidence for at least four fundamental subcortical emotional systems: SEEKING/EXPECTANCY (anticipatory-enthusiastic, foraging-reward states), RAGE (anger), FEAR (anxiety), and separation-distress/PANIC (sadness/grief). These were supplemented later by evidence for LUST, CARE, and PLAY reward systems, all of which are closely linked to general-purpose SEEKING urges, namely intrinsic emotional enthusiasms/interest-generating foraging circuits of the brain.
At that point, in the mid-1980s, behaviorists were beginning to embrace neuroscience, because funding for behavior-only studies of learning began to evaporate. A host of newly minted “behavioral neuroscientists” emerged—most prominently Mike Davis, Michael Fanselow, Joe LeDoux, Steven Maier and their prominent students (e.g., see Maren, Phan, & Liberzon, 2004; Maren & Quirk, 2013, for overviews). These labs started to craft rigorous understanding of mammalian learning and memory, largely through classical conditioning studies of “fear” (while disregarding affective states). Regrettably, they have uniformly denied that one can study emotional feelings scientifically in animals, while disregarding the DBS data, culminating in LeDoux’s (2012, p. 666) assertion that “We will never know what an animal feels.” Of course, science has no privileged access to any “absolute knowledge”; all positions, including affective sentience in animals, need to be resolved by the weight-of-evidence rather than proclamations of what we can or cannot know. I remain puzzled that neuro-behaviorists have yet to discuss how DBS-induced primal emotional arousals can be uniformly “rewarding” (SEEKING, LUST, CARE, and PLAY) or “punishing” (FEAR, RAGE, and PANIC), without associated feelings. Clearly DBS of those brain regions evoke emotional feeling in humans—replicable evidence that may allow investigators to access animals’ affective/experiential states and neural underpinnings (Panksepp, 1982, 1998).
From 1990 onward and for the ensuing quarter century, the field has massively diversified, with a prevalence of cognitive and constructivist approaches. The emergence of human brain imaging has promoted such trends, with increasing tensions and intellectual crosscurrents, emerging among those working at different evolutionary levels of brain and mind. Those focusing on higher psychological processes understandably prefer constructivist views of emotions (e.g., from Ortony & Turner, 1990 to Barrett, 2006), yielding “either-or” polarizations, which Izard (1992, 2007) and Panksepp (1992, 2007) chose to deconstruct and counter. We both advocated for an evidence-based, middle-road approach, recognizing that both views are needed for integrating bottom-up and top-down (Figure 1) perspectives (see Zachar & Ellis, 2013, for various such polarized perspectives).
A Decline of Primary-Process Emotional Perspectives?
Once I had the pleasure of getting to know Cal personally, I soon realized that he was one of the few human emotion researchers who supported straightforward empirically based views of emotional feelings in other animals which could guide our search for evidence-based constitutional (i.e., causal) understanding of how primal affective states arise in human/mammalian subcortical brain regions, providing evolutionary substrates that guide construction of higher mental processes neuro-developmentally (Figure 1). Although there is increasing agreement on the critical role of animal research in the various neurosciences, there remains abundant skepticism that animal brain research can clarify human emotional feelings (for related recent debates, see Lindquist, Wager, Kober, Bliss-Moreau, & Barrett, 2012). Izard never equivocated about the utility of neuro-evolutionary perspectives in affect science informing us about how affective mentalities arise from neurobiological dynamics. He also integrated constructivist perspectives harmoniously with modern neuroevolutionary understandings of emotions (Izard, 2007, 2009), seeking to avoid sterile academic disagreements.
I remain a fan of Izard’s open-minded view that we psychologists simply must bring neuroscience beyond the correlative vagaries of modern brain imaging, to clarify what emotional feelings really are. To understand the whole package of human emotions, top-down psychological strategies, requiring human research (Figure 1, right), should be integrated with bottom-up neurophenomenological investigations that require human developmental and animal research (Figure 1, left). Integration can be achieved by discussing and perhaps accepting that the rewarding and punishing properties of DBS-induced activations of subcortical emotional networks allow empirical access to animals’ primal affective feelings. This could provide workable models of the constitution of our own raw emotional states, and could inform and greatly impact psychiatric practice (Panksepp et al., 2014)
Top-down constructivism and bottom-up essentialism can work in cooperative ways, as opposed to being in perpetual conflict (Izard, 2007; Panksepp, 2007). A comparable need for integration could even heal the more prolonged clash between behaviorism and mentalism (Panksepp, 1990). Among the great tragedies of modern psychology is the failure of many evolutionary psychologists to integrate cross-species neuroevolutionary perspectives into their thinking, and the general resistance of behavioral neuroscientists to recognize that affective aspects of animal minds can finally be addressed experimentally. If rigorous cross-species brain predictions can credibly be related to human psychological levels of analysis, it could substantially strengthen the neuroscientific foundations of psychology (Panksepp, 2010), hopefully promoting more rapid progress in the development of effective psychiatric medicines and treatments (Panksepp et al., 2014).
Regrettably, many prominent investigators trained in behavioristic traditions remain committed to the century-old view that minds of other animals are scientifically impenetrable (Panksepp, 1990). The overall argument is simple: we can experimentally determine that the various primal emotional circuits of animal brains can sustain rewarding and punishing effects (Panksepp, 1982, 1998). Since we humans have no reward or punishments that we do not experience affectively, it is sensible (and full of novel scientific predictions, especially for psychiatry, e.g., Burgdorf, Panksepp, & Moskal, 2011; Coenen, Schlaepfer, Maedler, & Panksepp, 2011), that other animals have homologous affective experiences. Whether they have thoughts (“awareness-” related knowledge) about such experiences currently remains the impenetrable part. Affect in animals is an empirically workable topic.
Although psychological science alone does not have the power to penetrate into the mysteries of animal affects, functional neuroscience, conducted (hopefully ethically) on our fellow animals, can penetrate the shield of perpetual agnosticism, with direct clinically relevant predictions for our own species (Panksepp et al., 2014). As Cal Izard especially well recognized, the sooner such paths of scientific understanding are pursued, the sooner psychology will become a truly sophisticated psychobiological science. Primary-process affective neuroscience strategies, judiciously pursued with nonspeaking species, can give us testable ideas about how fundamental emotional feelings are actually constituted in human brains. We should emulate Izard’s lead on such integrative syntheses. Izard and I, as well as many others, including neuropsychiatrist Thomas Schlaepfer and neurosurgeon Volker Coenen (Panksepp et al., 2014) agree: There must be a substantial role for cross-species functional neuroscience in understanding some basic human psychological processes, including our raw emotional feelings, especially their neuroanatomical and neurochemical substrates, which may promote insightful translational research. We have recently seen how we can build a useful bridge between animal and human research with the massive enthusiasm about intranasal oxytocin, and its capacity to increase social attention and social warmth and confidence.
Had Cal entered the field of emotion studies a few decades later than he did, I suspect he would also have been tempted to pursue knowledge about our nature that can be obtained realistically only by understanding the emotions of our fellow creatures. Still, Izard’s research has been a blessing for emotion studies. He gave us a solid characterization of emotions in early development, and he integrated that knowledge with emerging neuroscientific understanding of animal emotions that started earnestly in the late 1970s. His seminal developmental work and broad integrative approach to emotions studies has been an exemplar of interdisciplinary scholarship. His contributions to our field are enormous. We now better appreciate how much we share with other creatures, and how the unique aspects of human emotionality arise from our expansive higher mental abilities and cultural values.
Basic emotion approaches to understanding the fundamental affective lives of human infants and other mammals have worked synergistically to give us an understanding of the psychological foundations upon which higher order human mental constructs are built. Among the living species, our higher order human emotions, “energizing” autonoetic consciousness (Tulving, 2002), may be unique. Of course (and obviously), our minds are more complex than those of the other creatures, allowing unique abilities, such as imagination. Nonetheless, our understanding of higher order human emotions can be informed not only by the emotions of human infancy, but also by comparative neuroscience perspectives. As Izard has long advocated, consilience among different approaches is needed to make psychological science more robust than any of the approaches taken alone. Because of Izard’s work, we now have the possibility of blending bottom-up basic emotion approaches with top-down constructivist approaches, and not marginalizing either approach. This can improve psychological science, yielding a more comprehensive vision of the complexities that we must deal with in order to understand the kinds of creatures that we are.