
Abstract
Huntington's disease (HD) is a hereditary neurodegenerative condition, passed down in an autosomal dominant manner, characterized by the gradual decline of motor functions, such as chorea, along with psychiatric symptoms and cognitive deterioration. In HD, the striatum is the main area impacted, although cerebellar atrophy has also been observed independently of striatal degeneration. Consequently, HD patients may exhibit symptoms consistent with cerebellar cognitive affective syndrome (CCAS) due to cerebellar involvement. CCAS manifestations have been noted in various rodent models of motor dysfunctions linked to cerebellar atrophy. Previous research has identified anxiety, memory deficits, and depressive-like behavior in the YAC128 transgenic murine model of HD. This investigation examines the effects of prolonged administration of the small-conductance calcium-activated potassium channel (SK channel) positive modulator chlorzoxazone (CHZ), both alone and in combination with folic acid (FA), on cerebellar Purkinje cell (PC) electrophysiology and morphology, along with locomotor and memory decline and emotional state changes in YAC128 mice. The findings indicate that both CHZ alone and the mixture of CHZ and FA similarly ameliorated cerebellar-specific deficits, encompassing the precision of PC electrophysiological activity and beam walk performance in YAC128 mice. However, solely the mixture of CHZ and FA significantly alleviated depressive-like symptoms and improved recognition memory. These findings imply that a therapeutic approach addressing both motor and cognitive-affective deficits is essential for the holistic management of movement disorders associated with cerebellar atrophy, including HD.
Keywords
Introduction
Huntington's disease (HD) is a hereditary neurodegenerative condition, passed down in an autosomal dominant manner, that typically manifests in individuals aged 35–50 years, leading to mortality within 15–20 years post-onset. A key characteristic of the disease is the progressive deterioration of motor function, with chorea being one of the symptoms, as well as psychiatric disturbances and cognitive decline. 1 Cognitive decline and mood disturbances often precede the more characteristic motor symptoms of Huntington's disease (HD). 2 Among the cognitive impairments, patients exhibit deficits in time perception, processing speed, emotion recognition, and difficulties with non-verbal communication, which can begin 10–15 years prior to a definitive diagnosis. 3 The most debilitating psychiatric disorder associated with HD is depression, which significantly increases the risk of suicide. Additionally, affective symptoms commonly include elevated anxiety, irritability, obsessive-compulsive behaviors, apathy, and, less frequently, psychotic episodes.4,5
The striatal degeneration of GABAergic medium spiny neurons (MSNs) is the main neuropathological marker of HD.1,6,7 However, cerebellar atrophy has also been recognized in HD. 8 Studies indicate that the cerebellum can degenerate early in the course of HD and independently from striatal atrophy.9,10 This points to the possibility that HD's pathological process is not confined to a single area and affects multiple brain regions simultaneously or sequentially. 9 In the progression of HD symptoms, the cerebellum works in conjunction with the basal ganglia and cerebral cortex. The specific pattern of cerebellar damage is closely linked to the disease's symptoms, highlighting a complex interaction between these brain regions. 11 HD is polyglutamine diseases caused by an expansion of CAG repeats, which encode a glutamine stretch in the huntingtin (Htt) protein. There is established correlation between the length of the polyglutamine tract, the age of disease onset, and the severity of symptoms. Cerebellar atrophy and ataxia are frequently noted in juvenile Huntington's disease (JHD), which manifests before the age of 20 and is typically associated with more than 60 CAG repeats. A case of severe cerebellar atrophy in an 18-year-old HD patient with 56 CAG repeats on one allele and 20 on the other has been described. 12 In a 9-year-old JHD patient with 84 CAG repeats, cerebellar atrophy and pronounced gait abnormalities were observed. 13 A histological study of post-mortem cerebellar tissue from a father with adult-onset HD (54 CAG repeats) and his son with JHD (164 CAG repeats) revealed significant midline and vermis atrophy in the juvenile case. Histopathological analysis demonstrated marked atrophy of the cerebellar cortex and prominent gliosis in JHD but not in the adult-onset manifestation of the disease. 14
Studies on mice also demonstrated cerebellar involvement in HD pathogenesis. Investigations using the R6/2 transgenic murine model of HD have revealed decreased levels of calcium-interacting proteins specific to cerebellar Purkinje cells (PCs), such as parvalbumin and calbindin. 15 Additionally, PC loss and decreased PC firing rates were observed in presymptomatic R6/2 mice. 15 Interestingly, in the HdhQ200 knock-in mouse model of HD analogous findings were reported. 16 In our previous studies, the extracellular single-unit recording technique has YAC128 mice when compared to their wild type (WT) controls. 17
Therefore, preceding research has shown a relationship between cerebellar degeneration and motor symptoms in HD.11,18 It is known that the cerebellum's involvement may extend beyond motor functions and may include cognitive and affective symptoms.19,20 This implies that patients with HD might display signs of cerebellar cognitive affective syndrome (CCAS) due to the cerebellar component of their condition. Thus, it is plausible that HD patients could suffer from CCAS, contributing to their cognitive and affective impairments. Manifestations of CCAS have been documented in various mouse models of movement disorders Specifically, in transgenic murine models of spinocerebellar ataxia type 1 (SCA1), the altered anxiety levels and deficits in spatial and fear memory were reported. 21 SCA2 transgenic mice displayed reduction in anxiety, minor impairments in visuospatial memory function, and depressive-like symptoms. 22 Similarly, murine models for episodic ataxia type 2 (EA2) demonstrated disruptions in anxiety levels and recognition memory. 23 SCA3-84Q transgenic mice experienced anxiety, recognition memory decline, and depressive-like symptoms. 24 SCA6 mice exhibited reduction in anxiety and decline in visuospatial memory function. 25 Thus, murine models of motor diseases associated with cerebellar atrophy generally exhibit both motor and cognitive-affective symptoms, providing a valuable framework for studying combined therapeutic approaches targeting both ataxia and CCAS.
Cerebellar PCs constitute a primary dynamic component within the cerebellar cortex. Dysregulation of PC firing is a shared characteristic across various types of movement disorders associated with cerebellar atrophy. 26 Small conductance calcium-activated potassium channels (SK channels) are fundamental in governing the intrinsic activity of cerebellar PCs, highlighting their pivotal importance in neural functionality and signaling. 27 It has been hypothesized that pharmacological positive modulation of SK channels could ameliorate motor symptoms by restoring PC firing. Indeed, SK activators have been shown to rescue PC pacemaking and enhance motor coordination in various models, including SCA1-82Q mice, 28 SCA2-58Q mice, 29 SCA3-84Q mice, 30 EA2 mouse models,31–33 mutant CACNA1A mice, 34 and the YAC128 mice. 17 Moreover, the SK positive modulator riluzole ameliorated SARA scores in human patients with hereditary ataxia without severe side effects. 35 However, SK channel activation is known to inhibit electrophysiological activity in neurons,36,37 which could potentially lead to adverse effects in human patients. In prior studies involving SCA2-58Q and SCA3-84Q mice, we observed that folic acid (FA) demonstrated an ability to counteract the suppression in neuronal firing rate induced by chlorzoxazone (CHZ), a widely recognized SK channel activator, highlighting its potential therapeutic significance.38,39 The present study was conducted with the objective of rigorously assessing the impact of CHZ, FA, and their combination on motor performance, cognitive abilities, and mood disturbances in YAC128 hemizygous transgenic mice, underscoring the importance of understanding these variables in the context of neurodegenerative conditions (Figure 1).
A timeline of the CHZ-FA trial in YAC128 mice. At 10.5 months of age, WT and YAC128 mice received i. p. injections of either vehicle, CHZ, FA, or a combination of CHZ and FA over a six-week period. Subsequently, various behavioral tests were conducted: the BW assay to evaluate motor coordination, the OF test to assess anxiety levels, the NOR test to examine recognition memory, and the TST to measure depressive-like behavior.
Our prior research indicated that the CHZ-FA combination exerted beneficial effects on both motor and cognitive deficits and further improved changes in mood in SCA2-58Q mice 38 and also in SCA3-84Q mice 39 without influencing the firing frequency of cerebellar PCs. Our preceding results have demonstrated that CHZ improves impaired beam walk performance in YAC128 mice. 17 In the current study, we sought to examine the impact of the SK channel activator CHZ, both independently and in combination with FA, on motor performance, cognitive abilities, and mood disturbances in YAC128 mice.
Materials and methods
Mice breeding and genotyping
In the present study, transgenic YAC128 mice and their WT controls, aged 12 months, were utilized,. The YAC128 mice, possessing a FVBN/NJ background, were procured from Jackson Laboratory (Bar Harbor, ME, USA; strain number 004938) and bred according to previously established protocols.17,40 Specifically, heterozygous male YAC128 mice were bred with WT females to generate mixed litters; from these breeding pairs, 64 one-month-old female offspring were selected for genotyping. Genotyping of the offspring was performed using PCR targeting the Htt transgene. Each PCR reaction had a total volume of 25 µL, comprising: 2.5 µL of 10× Taq polymerase buffer, 0.5 µL of 10 mM dNTPs, 1.5 µL of 25 mM MgCl2, 0.125 µL of 20 µM forward and reverse primers, 0.25 µL of Taq polymerase, 2 µL of DNA template, and 18 µL of dH2O. The forward primer sequence was 5′-CCGCTCAGGTTCTGCTTTTA-3′, and the reverse primer sequence was 5′-TGGAAGGACTTGAGGGACTC-3′, yielding a PCR product of 170 bp. The mice were grouped in cages containing three to six animals each and kept in a carefully controlled vivarium environment, maintained at a temperature of 22–24°C, with a regulated 12-h light/dark cycle to ensure consistency and stability essential for their well-being. Food and water were supplied without restriction, ensuring the subjects’ needs were met. All experimental protocols received formal approval from the Bioethics Committee of Peter the Great St Petersburg Polytechnic University, St Petersburg, Russia, and were strictly carried out in compliance with established guidelines and regulations. Additionally, the study rigorously followed the ARRIVE guidelines to maintain ethical and methodological standards.
Drug delivery in mice
The experimental protocol was performed following the procedure outlined in prior descriptions.38,39 The experimental animals underwent genotyping, they were further matched by weight, and allocated into four WT groups and four transgenic YAC128 groups, with each group comprising 7 to 9 subjects. Over 6-week period mice received intraperitoneal (i.p.) injections of 30 mg/kg CHZ, 30 mg/kg FA, or a combination of both at 30 mg/kg CHZ and 30 mg/kg FA dissolved in 5% DMSO in PBS, according to a 5/2 regimen, with injections suspended on the weekly cage-changing days. Control groups received vehicle injections similarly to that described above. Injections were administered to experimental animals aged 10.5 to 12 months.
This study exclusively involved the use of female subjects due to the aggressive behavior exhibited by male YAC128 littermates.41,42 Following genotyping, 64 one-month-old mice (34 WT and 30 YAC128) were divided into eight experimental groups. The experimental groups were as follows: 8 WT Ctrl, 9 WT CHZ, 8 WT FA, and 8 WT CHZ-FA; 8 YAC128 Ctrl, 8 YAC128 CHZ, 7 YAC128 FA, and 7 YAC128 CHZ-FA. A graphical timeline of the experimental design, including therapeutic interventions and structured behavioral tests, is presented in Figure 1. The behavioral assessments were conducted in the light phase, with the mice's body weight did not differ across groups.
Motor activity assessment
The beam walk (BW) test was employed to evaluate motor activity in YAC128 mice, following the procedure outlined in prior descriptions.17,43 A custom-made setup consisting of a 1.5-meter beam suspended 0.5 meters above the floor was utilized. Wood beams of varying diameters (18 mm, 12 mm, and 8 mm) were utilized. Over a period of three consecutive days, mice underwent training to navigate the beams, with each beam involving three dedicated training sessions per day. On the fourth day, the mice underwent three consecutive trials on each beam, systematically advancing from the widest beam to the narrowest. The latency to cross one meter of each beam and the incidence of hind paw slips, referred to as “footslips,” were evaluated for each trial. The mean values from three trials for each beam were computed separately for each group tested.
Behavior tests
Behavioral battery order
To avoid potential confounding effects from prior test exposure, the behavioral tests were performed in a fixed order (Figure 1). Following the motor coordination assessment using the Beam Walk (BW) test, general locomotor activity and anxiety-like behavior were evaluated in a one-day Open Field (OF) test session. The following day, recognition memory was assessed using a two-day Novel Object Recognition (NOR) protocol conducted in the same OF apparatus under identical lighting conditions. Finally, depressive-like behavior was evaluated using the Tail Suspension Test (TST), which was intentionally placed last in the battery to prevent stress from this procedure from influencing performance in the preceding tests.
Open field (OF)
The OF task was utilized to assess anxiety levels in experimental animals. The OF arena consisted of a non-transparent Plexiglas cylindrical chamber with a diameter of 62.4 cm which was divided into three distinct zones: a central zone with a diameter of 28.4 cm, an intermediate zone, and a border zone located 8.5 cm from the chamber's outer wall, illuminated at 950 lux. The mice were placed at the center of the setup, and their behavior was systematically observed and video documented over a 10-min period. The analysis encompassed parameters such as the time spent in the central
Novel object recognition (NOR)
The NOR test was employed to evaluate the critical ability to identify previously encountered information in YAC128 mice, following a technique adapted from previous studies.23,44 The NOR task was conducted after the OF test, with mice already habituated to the apparatus. The OF apparatus was lit with an illumination level of 950 lux. During the initial test day, experimental animals were placed between two identical blue cube objects, and their actions were monitored through video tracking over a duration of 10 min. During the next test day, the mice were positioned between a familiar object (a blue cube) and a novel object (a red torus), and their actions were monitored through video tracking over a 10-min period. The apparatus was sanitized with 70% ethanol solution between trials. The latency to approach the novel object and the frequency of interactions with the novel object were assessed through analysis using Any-maze software. The percentage preference for equal and novel objects was determined by calculating the ratio of the time spent investigating the object of interest to the overall exploration time. Exploration time was measured as the duration during which the animal's head remained within a 2 cm perimeter surrounding the object. Trials were considered successful if mice explored objects for at least 10 s. Consequently, two mice were removed from the WT FA group, and one mouse was excluded from the YAC128 FA group.
Tail suspension test (TST)
The TST was conducted to evaluate behaviors indicative of depressive-like symptoms in YAC128 mice. To prevent tail-climbing behavior during the TST, a smooth plastic cylinder (4.5 cm in length, 1.5 cm in diameter) was placed around the base of each mouse's tail. Mice were positioned 40 cm above the floor within a custom-made white chamber measuring 20 × 80 × 50 cm, partitioned into four smaller compartments, each with dimensions of 20 × 20 × 50 cm. Each trial lasted for 6 min, and the time spent immobile in 0–120 s, 120–240 s, and 240–360 s intervals was assessed manually using Pole Krest software (developed by Dr V.V. Sizov, Institute of Experimental Medicine, Saint Petersburg, Russia, sizoff@list.ru).
Cerebellar slice recordings
In vivo loose-patch recordings of spontaneous PC activity from 12-month-old WT and YAC128 mice were performed as previously described.38,39 Briefly, mice were deeply anesthetized with urethane (2000 mg/kg) and transcardially perfused with ice-cold artificial cerebrospinal fluid (aCSF) containing (in mM): 85 NaCl, 24 NaHCO3, 25 glucose, 2.5 KCl, 0.5 CaCl2, 4 MgCl2, 1 NaH2PO4, and 75 sucrose (equilibrated with carbogen: 95% O2 / 5% CO2). Cerebella were rapidly dissected, and 300 μm thick sagittal slices were prepared using a VT1200S vibratome (Leica). Slices were recovered for 30–40 min at 35°C in standard aCSF containing (in mM): 119 NaCl, 26 NaHCO3, 11 glucose, 2.5 KCl, 2.5 CaCl2, 1.3 MgCl2, and 1 NaH2PO4 (equilibrated with carbogen), then maintained at room temperature until recording. Recordings were conducted within 6–7 h after dissection in a chamber heated to 35°C (TC-324C controller, Warner Instruments). The external recording solution was identical to the recovery aCSF but additionally contained 100 μM picrotoxin (PTX) and 10 μM 6,7-dinitroquinoxaline-2,3-dione (DNQX) to block inhibitory and excitatory synaptic transmission, respectively. Loose-patch recordings (<100 MΩ seal resistance) were obtained from the soma at the axon hillock region of PCs using glass pipettes (1.5–3 MΩ) filled with an internal solution of 140 mM NaCl buffered with 10 mM HEPES (pH 7.3) and held at 0 mV. Spontaneous action potential currents were recorded for 5 min per cell using an Axon Multiclamp 700B amplifier (Molecular Devices). Data analysis was performed using Clampfit 10.2 (Molecular Devices). For pharmacological experiments, 30 μM chlorzoxazone (CHZ), 30 μM folic acid (FA), or both (CHZ-FA) were added directly to the recording solution.
Immunohistochemistry (IHC)
The mice underwent transcardial perfusion using +4°C phosphate-buffered saline (PBS) with a pH of 5.4, continuing the process until the solution ran clear, ensuring thorough vascular flushing, followed by perfusion with 4% paraformaldehyde (PFA). Brains were further extracted and postfixed in 4% PFA throughout the night at +4°C and then dehydrated in 30% sucrose for approximately two days. Cerebellar sagittal slices (50 µm thick) were prepared using an NVSLM1 vibroslice (World Precision Instruments). The cerebellar slices were washed twice in PBS and treated with acetone at −20°C for 30 min to enhance tissue antigenicity, 34 followed by an overnight incubation in 0.5% Triton X-100 in PBS at 4°C. The cerebellar slices were then rinsed in a blocking solution containing 0.5% Triton X-100 with 2% bovine serum albumin (BSA) dissolved in PBS during 1 h at +22°C, followed by a 48-h incubation at +4°C with primary antibodies (mouse monoclonal anti-calbindin-D-28 K antibody, 1:900; Sigma-Aldrich, C9848) in blocking solution. Following four PBS rinses, slices were incubated with secondary antibodies (1:500 Alexa Fluor 594 goat anti-mouse, A11005; Invitrogen) in blocking solution for an additional 48 h at +4°C. Following four final PBS washes, the cerebellar slices were placed onto slides using ProLong Gold antifade reagent (P36934, Invitrogen). The imaging was conducted utilizing a confocal microscope (ThorLabs). Three individual slices from each mouse were examined for analysis, with three mice per experimental group. The thickness of the molecular layer was analyzed using 20× images processed with ImageJ (NIH Image). This parameter was measured 100 µm from the primary fissure, following the procedure outlined in prior descriptions.30,39 The quantity of PCs was determined within a 100 µm region extending from the depth of the primary fissure, using three slices per mouse with a total of nine counts analyzed for each experimental group.
Statistical analysis
All statistical tests were performed using Origin software. One-way ANOVA was used for comparisons between experimental groups, while two-way ANOVA was employed for multiple comparisons. The data are displayed using bar graphs accompanied by SEM bars, with overlaid dot plots for additional visualization. Significance levels are indicated as follows: * p < 0.05; ** p < 0.01; *** p < 0.001; **** p < 0.0001. The figure legends provide detailed information on the number of mice used in each experiment.
Results
CHZ-FA restores the firing variability in YAC128 cerebellar PCs
Disturbances in cerebellar PC spontaneous electrophysiological activity have been observed in different murine models of HD. Thus, a reduction in PC firing frequency was reported in R6/2, 15 HdhQ200, 16 and YAC128 transgenic mice. 17 In the present study, the loose-patch recordings were employed to investigate the PCs’ firing in acute cerebellar slices derived from YAC128 mice and their WT controls of the same age at 3, 6, and 9 months old. For each group, the average firing rate and the variability in firing were assessed. The coefficient of variation of interspike intervals (CV ISI), determined by dividing the standard deviation (SD) of the interspike intervals (ISIs) by their average, served as the measure of firing variability. Our findings revealed no differences in firing rate or variability between YAC128 and WT mice at the age of 3 months (Figure 2B and C).
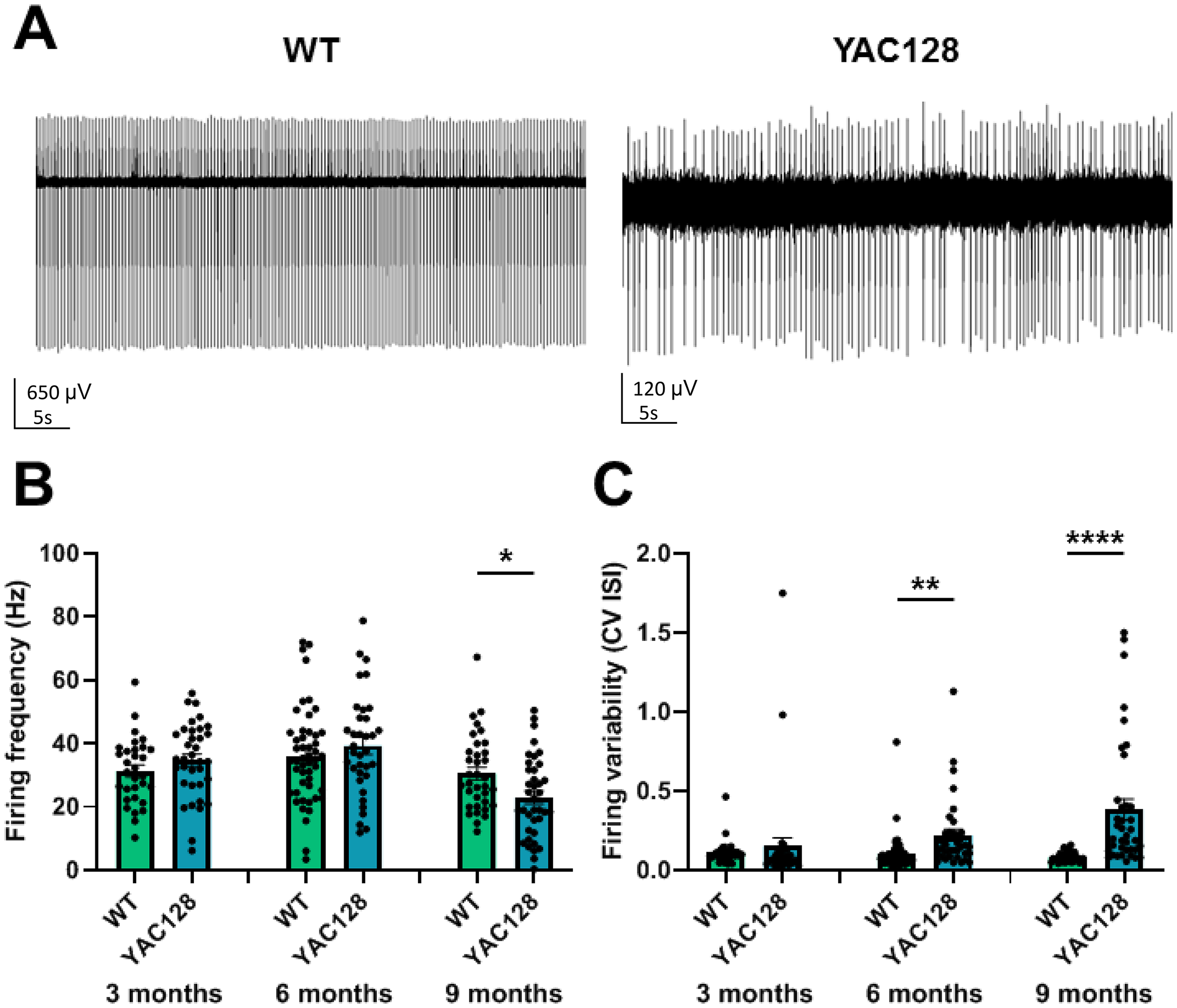
The cerebellar Purkinje cells’ firing is impaired in aged YAC128 mice. (A) Examples of PC firing in WT and YAC128 mice at the age of 9 months are presented as 5 s duration recordings. (B) The firing frequency was assessed in 3-, 6-, and 9-month-old WT and YAC128 PCs. The firing frequency of PCs in 9-month-old YAC128 mice was lower than in age-matched WT PCs. (C) The CV ISI was assessed in 3-, 6-, and 9-month-old WT and YAC128 PCs. In 6- and 9-month-old YAC128 mice PCs fired more irregularly compared to their WT littermates. Statistical significance was evaluated with ANOVA (* p < 0.05; ** p < 0.01; **** p < 0.0001). Number of recorded PCs (n) and number of mice (m): n = 31 and m = 3 for 3-month-old WT mice; n = 35 and m = 3 for 3-month-old YAC128 mice; n = 49 and m = 3 for 6-month-old WT mice; n = 35 and m = 3 for 6-month-old YAC128 mice; n = 35 and m = 3 for 9-month-old WT mice; n = 38 and m = 3 for 9-month-old YAC128 mice.
However, at 9 months, YAC128 mice exhibited a significantly reduced (* p < 0.05) PC firing rate relative to their WT controls (Figure 2A and B). At 9 months, WT mice showed an average firing frequency of 30.56 ± 2.01 Hz (n = 35 PCs, m = 3 mice), compared to 22.97 ± 2.06 Hz (n = 38 PCs, m = 3 mice) for YAC128 mice. The precision of PC firing in YAC128 mice was significantly (** p < 0.01) reduced by the age of 6 months (Figure 2C). At 6 months, the mean CV ISI for WT mice was 0.10 ± 0.02 (n = 49 PCs, m = 3 mice), compared to 0.22 ± 0.04 (n = 35 PCs, m = 3 mice) for their YAC128 littermates. YAC128 mice at 9 months also showed significantly (**** p < 0.0001) less regular PC firing (Figure 2C). At 6 months, the mean CV ISI for WT mice was 0.08 ± 0.01 (n = 35 PCs, m = 3 mice), compared to 0.38 ± 0.06 (n = 38 PCs, m = 3 mice) for their YAC128 littermates. At 3 months, YAC128 mice and WT controls exhibited comparable PC firing precision (p = 0.51).
Overall, the abnormal PC firing variability at 6 months of age and decline in PC firing frequency at 9 months of age in YAC128 mice aligns closely with the age at which motor incoordination begins, as noted in our earlier research. 17 We hypothesize that the anomalies in PC spontaneous electrophysiological activity observed in YAC128 mice contribute to the deficits in motor coordination tasks exhibited by these mice beginning at 6 months of age.
In the subsequent battery of experiments, we investigated the impact of CHZ both independently and in conjunction with FA on cerebellar PC spontaneous electrophysiological activity in mice aged 12 months (Figure 3A). The sets for cerebellar slice electrophysiology were as follows: WT and YAC128 control (Ctrl), WT and YAC128 PCs exposed to 30 μM CHZ, WT and YAC128 PCs exposed to 50 μM FA, and WT and YAC128 PCs exposed to combination of both at 30 μM CHZ and 30 μM FA. The recording bath solution was used to administer drugs. Consistent with previous findings (Figure 2B), a significantly lower average firing frequency was observed in the YAC128 Ctrl group relative to the WT Ctrl group (* p < 0.05; Figure 3B). WT Ctrl PCs exhibited an average firing frequency of 34.24 ± 1.70 Hz (n = 82 PCs, m = 5 mice), whereas YAC128-Ctrl-PCs showed an average of 27.69 ± 1.93 Hz (n = 35 PCs, m = 3 mice). The presence of CHZ in the bath solution significantly lowered the firing rate in both WT CHZ and YAC128 CHZ groups (Figure 3B). WT-CHZ-PCs exhibited an average firing frequency of 17.86 ± 0.93 Hz (n = 49 PCs, m = 3 mice, **** p < 0.0001 vs WT Ctrl group), whereas YAC128-CHZ-PCs showed an average of 19.43 ± 1.38 Hz (n = 42 PCs, m = 3 mice, *** p < 0.001 vs YAC128 Ctrl group). On the other hand, the presence of FA in the bath solution resulted in a higher firing rate in both WT and YAC128 PCs, but the difference was not statistically significant in the WT FA group (Figure 3B). WT-FA-PCs exhibited an average firing frequency of 39.89 ± 3.09 Hz (n = 43 PCs, m = 3 mice, p = 0.08 vs WT Ctrl group), whereas YAC128-FA-PCs showed an average of 44.03 ± 4.58 Hz (n = 33 PCs, m = 3 mice, ** p < 0.01 vs YAC128 Ctrl group). Lastly, adding both CHZ and FA to the bath solution did not affect the mean firing rate in WT or YAC128 control PCs (Figure 3B). WT-CHZ-FA-PCs exhibited an average firing frequency of 32.00 ± 1.42 Hz (n = 66 PCs, m = 4 mice, p = 0.33 vs WT Ctrl group), whereas YAC128-CHZ-FA-PCs showed an average of 23.86 ± 1.44 Hz (n = 30 PCs, m = 3 mice, p = 0.13 vs YAC128 Ctrl group).
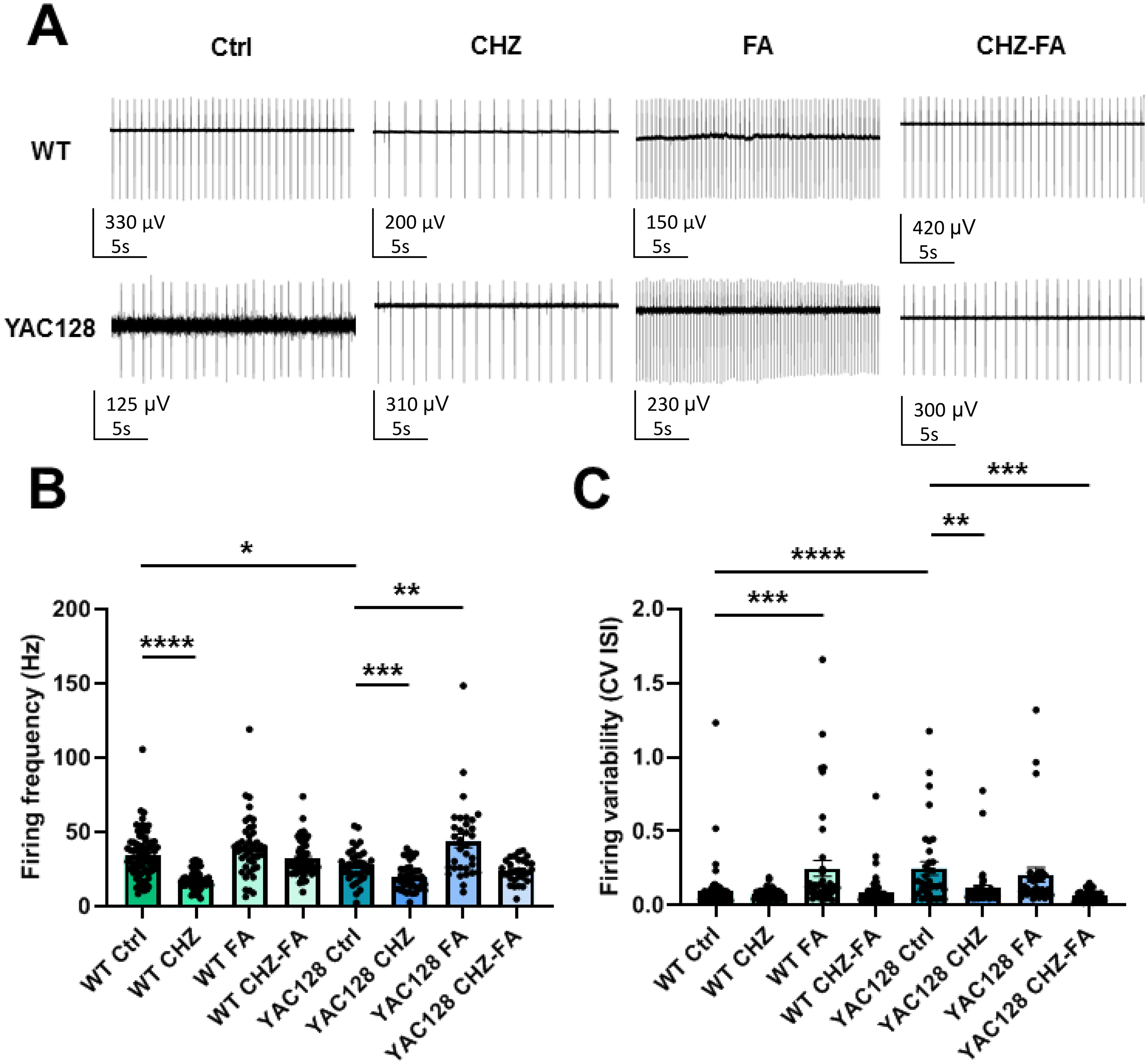
CHZ-FA ameliorates PC firing precision in YAC128 mice. (A) Representative traces of PC firing in 12-month-old WT and YAC128 mice, both in control conditions and following treatment with CHZ, FA, and a combination of CHZ and FA. Recordings 1 s duration are shown. The firing frequency (B) and the CV ISI (C) of cerebellar PCs were assessed in 12-month-old WT and YAC128 mice, both in control conditions and following treatment with CHZ, FA, and a combination of CHZ and FA. Statistical significance was evaluated with ANOVA (* p < 0.05; ** p < 0.01; *** p < 0.001; **** p < 0.0001). Number of recorded PCs (n) and number of mice (m): n = 82 and m = 5 for WT Ctrl group; n = 49 and m = 3 for WT CHZ group; n = 43 and m = 3 for WT FA group; n = 66 and m = 4 for WT CHZ-FA group; n = 35 and m = 3 for YAC128 Ctrl group; n = 42 and m = 3 for YAC128 CHZ group; n = 33 and m = 3 for YAC128 FA group; n = 30 and m = 3 for YAC128 CHZ-FA group.
The results of this research demonstrated that both CHZ alone and the combination of CHZ and FA ameliorated the impaired firing variability of the YAC128 PCs (Figure 3C). The firing precision was altered in the YAC128 Ctrl group contrasted with their WT controls (Figure 3C). WT-Ctrl-PCs exhibited the mean CV ISI of 0.09 ± 0.02 (n = 82 PCs, m = 5 mice), whereas YAC128-Ctrl-PCs showed an average of 0.25 ± 0.05 (n = 35 PCs, m = 3 mice, **** p < 0.0001). The presence of CHZ in the bath solution had no impact on the firing variability of WT PCs, but reduced the firing precision in YAC128 CHZ group (Figure 3C). WT-CHZ-PCs exhibited the mean CV ISI of 0.08 ± 0.01 (n = 49 PCs, m = 3 mice, p = 0.41 vs WT Ctrl group), whereas YAC128-CHZ-PCs showed an average of 0.11 ± 0.02 (n = 42 PCs, m = 3 mice, ** p < 0.01 vs YAC128 Ctrl group). On the other hand, the presence of the FA in the bath solution decreased the precision of spontaneous activity in WT PCs, but had no impact on the CV ISI in YAC128 FA group (Figure 3C). WT-FA-PCs exhibited the mean CV ISI of 0.25 ± 0.05 (n = 43 PCs, m = 3 mice, *** p < 0.001 vs WT Ctrl group), whereas YAC128-FA-PCs showed an average of 0.21 ± 0.05 (n = 33 PCs, m = 3 mice, p = 0.53 vs YAC128 Ctrl group). When CHZ and FA were combined, the firing variability of WT PCs remained unaffected, while it was ameliorated in YAC128 PCs (Figure 3C). WT-CHZ-FA-PCs exhibited the mean CV ISI of 0.09 ± 0.01 (n = 66 PCs, m = 4 mice, p = 0.72 vs WT Ctrl group), whereas YAC128-CHZ-FA-PCs showed an average of 0.06 ± 0.01 (n = 30 PCs, m = 3 mice, *** p < 0.001 vs YAC128 Ctrl group).
Our research demonstrates that at the age of 12 months cerebellar PCs from YAC128 mice exhibit reduced firing frequency and precision when juxtaposed with WT mice of the same age. The presence of CHZ decreases the rate of spontaneous activity in both WT and YAC128 PCs, whereas FA increases it in YAC128 PCs. Notably, CHZ, rather than FA, enhances the variability of spontaneous activity in YAC128 PCs. Furthermore, the combination of CHZ and FA recovers the variability of spontaneous activity in YAC128 PCs without altering their frequency.
Administration of CHZ and FA, but not CHZ-FA combination, increases the number of PCs in YAC128 mice
In our preceding study we have observed the reduction in the arborization of PCs in 12-month-old YAC128 mice, whereas the thickness of molecular layer was equal to WT controls. 17 In this study, we employed anti-calbindin immunostaining (Figure 4A) to assess the PC count (Figure 4B) and the thickness of molecular layer (Figure 4C) in the cerebella of mice. The experimental cohort comprised WT and YAC128 mice at 12 months of age that underwent chronic administration of control solution, CHZ, FA, and a CHZ-FA combination. Following behavioral tests, these mice were utilized for IHC experiments. Three cerebellar slices per mouse were analyzed, with n = 3 mice per each trial group. The obtained results revealed a significant reduction in the number of PCs in YAC128 mice relative to their WT controls (Figure 4A and B). Specifically, the mean PC count positioned within a 100 μm radius of the primary fissure was 36.61 ± 1.74 in WT control mice and 29.67 ± 1.55 in YAC128 control mice (** p < 0.01). Administration of CHZ had no effect on the number of cerebellar PCs in WT mice yet notably elevated it in YAC128 mice. The average PC count was 33.33 ± 1.28 in WT CHZ mice (p = 0.15 vs. WT control) and 34.56 ± 1.17 in YAC128 CHZ mice (* p < 0.05 vs. YAC128 control). Similarly, FA injections did not affect WT mice although it considerably amplified the number of cerebellar PCs in YAC128 mice, with averages of 33.89 ± 2.63 in WT FA mice (p = 0.40 vs. WT control) and 36.33 ± 1.73 in YAC128 FA mice (* p < 0.05 vs. YAC128 control). The number of cerebellar PCs remained unchanged by the mixture of CHZ and FA in both WT and YAC128 mice, with averages of 38.83 ± 3.49 in WT CHZ-FA mice (p = 0.58 vs. WT control) and 32.17 ± 2.04 in YAC128 CHZ-FA mice (p = 0.34 vs. YAC128 control). The thickness of cerebellar molecular layer remained consistent across all experimental subsets (Figure 4C).
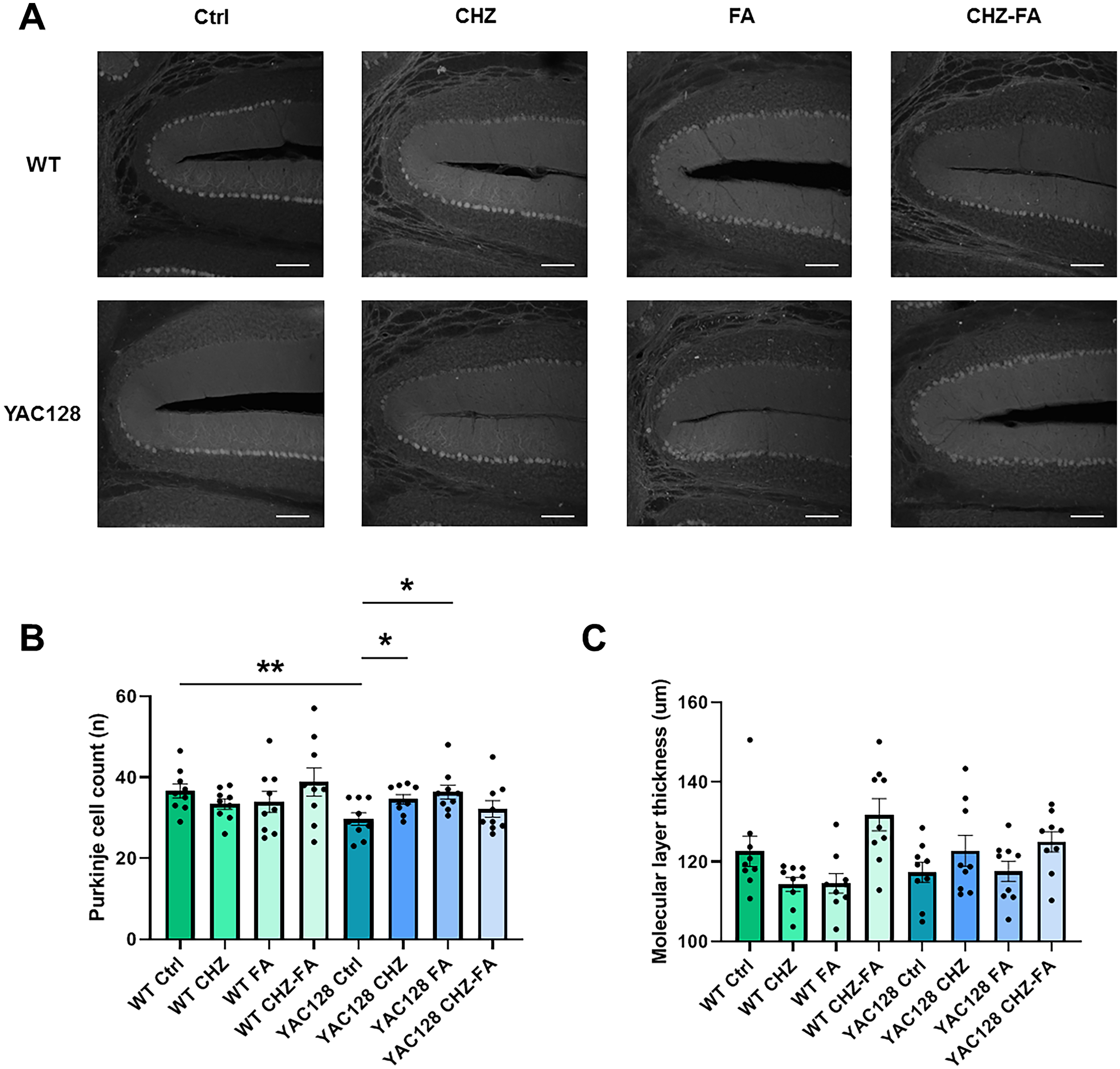
CHZ and FA recover the PC count in YAC128 mice. (A) Immunohistochemical analysis of calbindin in cerebellar tissue sections from 12-month-old WT and YAC128 mice, both in control conditions and following treatment with CHZ, FA, and CHZ-FA. Scale bars, 100 μm. (B) The PC count was analyzed in 12-month-old WT and YAC128 mice, both in control conditions and following treatment with CHZ, FA, and a combination of CHZ and FA. (C) The thickness of the molecular layer was consistent across the cerebella in all experimental groups. Statistical significance was evaluated with ANOVA (* p < 0.05; ** p < 0.01). Number of slices analyzed: n = 9 and number of mice: m = 3 mice per each group.
Our findings align with previously published results, demonstrating a significantly lower number of cerebellar PCs in HD mice measured against WT controls, with no difference in molecular layer thickness.15–17 Longitudinal administration of CHZ and FA, but not CHZ-FA combination enhanced PC counts in YAC128 mice.
CHZ individually and in combination with FA ameliorate BW performance in YAC128 mice
Motor coordination was assessed using the BW test in eight test cohorts at the age of 12 months (Figure 5A). The mean body weight did not differ significantly among the groups (data not shown). The test utilized 18 mm, 12 mm, and 8 mm round wooden beams to evaluate movement activity by measuring latency and the footslip count.17,29,38,45 The BW test has demonstrated that YAC128 control mice exhibited a higher frequency of rear paw slips (Figure 5C, E and G) compared to their WT controls. The chronic administration of CHZ substantially improved the motor activity in YAC128 mice (Figure 5C, E and G) without affecting WT mice (Figure 5). In contrast, FA injections prolonged the traversal time for all tested beams in WT mice, but they did not affect the footslip count or latency in YAC128 mice (Figure 5). The CHZ-FA combination improved motor performance in YAC128 mice on the smallest beam (* p < 0.05 vs. YAC128 control group, Figure 5G). Moreover, this mixture had no effect on the motor performance in WT mice.
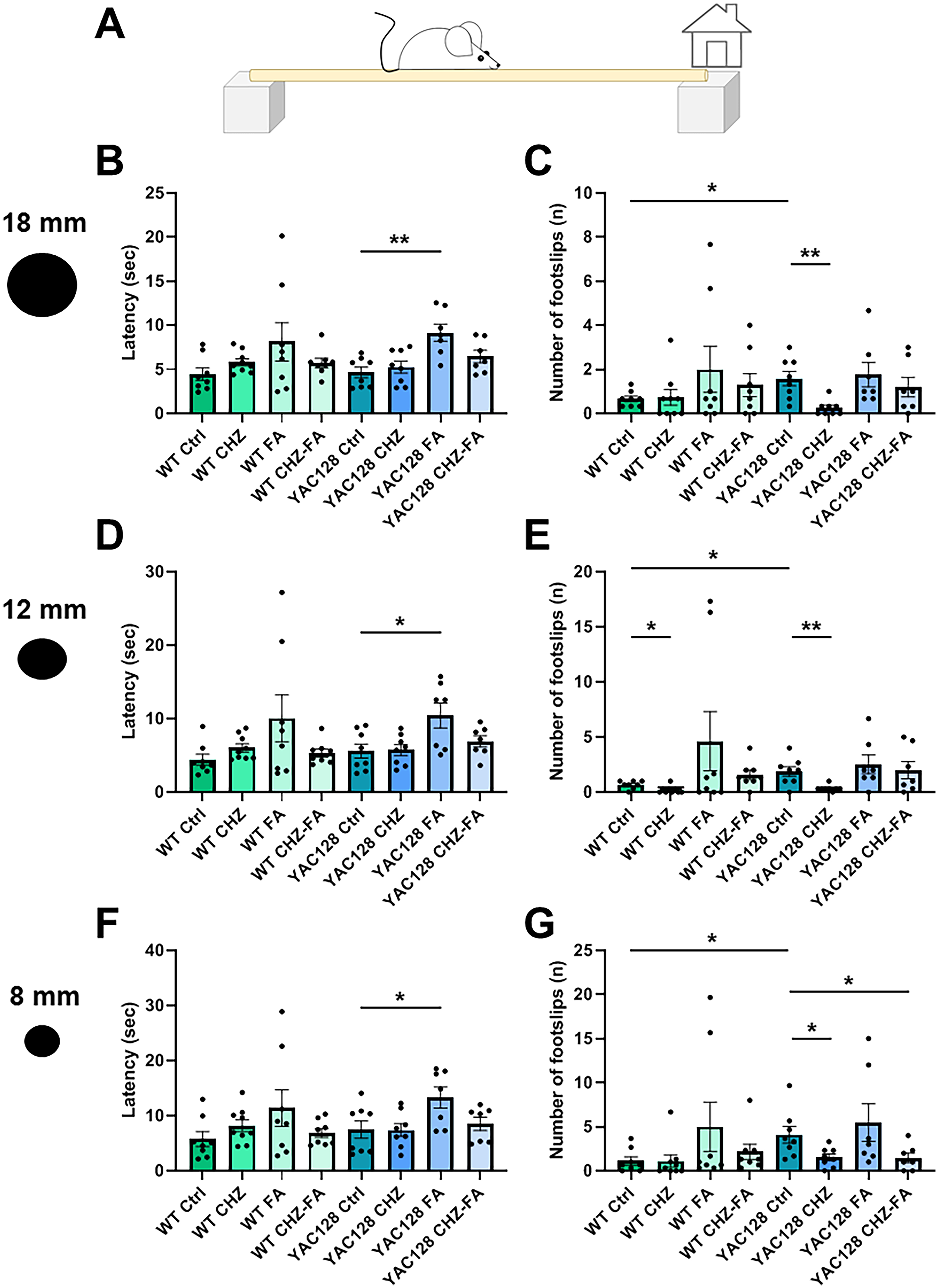
CHZ and CHZ-FA combination improve motor performance in YAC128 mice. (A) Scheme of the BW assay. The mean traversal time for mice crossing a 1-meter section of the beam, with diameters of 18 mm (B), 12 mm (D), and 8 mm (F), was evaluated in 12-month-old WT and YAC128 mice under control conditions and following long-term administration of CHZ, FA, and the CHZ-FA combination. The average count of foot slips during the 1-meter traversal of the 18 mm (C), 12 mm (E), and 8 mm (G) beams was assessed in 12-month-old WT and YAC128 mice across control, CHZ, FA, and CHZ-FA treatment groups. Statistical significance was evaluated with ANOVA (* p < 0.05; ** p < 0.01). The number of mice tested per group (n): n = 8 WT Ctrl mice, n = 9 WT CHZ mice, n = 8 WT FA mice, n = 8 WT CHZ-FA mice; n = 8 YAC128 Ctrl mice, n = 8 YAC128 CHZ mice, n = 7 YAC128 FA mice, n = 7 SCA3 CHZ-FA mice.
CHZ, FA, and CHZ-FA do not affect anxiety in YAC128 mice
We previously reported that YAC128 mice demonstrate anxiolytic behavior in the OF test at the age of 9 months most likely due to their increased locomotor activity. 44 In this study, we investigated the impact of prolonged administrations of CHZ and FA, individually and together, on the anxiety level in 12-month-old YAC128 mice and their WT controls using the OF task (Figure 6). By this age, WT and YAC128 mice exhibited comparable behavior during the OF task (Figure 6). Thus, experimental animals in both control groups allocated the same amount of time in the border (Figure 6B), intermediate (Figure 6C), and central areas (Figure 6D). The cessation of anxiolytic behavior in 12-month-old YAC128 mice, measured against the 9-month-old YAC128 mice, was likely due to a more than 50% reduction in locomotor activity in the older mice (Figure 6E). Thus, in the 10-min OF test, the distance traveled by 9-month-old YAC128 mice was 80.38 ± 18.26 m, 46 compared to 29.21 ± 6.02 m for 12-month-old YAC128 mice (Figure 6E). Noteworthy, the mean overall distance traveled was comparable in the control groups of 12-month-old YAC128 and WT mice (p = 0.46, Figure 6E). Chronic administrations of CHZ, FA, and the mixture of CHZ and FA showed no effect on anxiety levels in either WT or YAC128 mice (Figure 6).

Treatment with CHZ, FA and CHZ-FA did not affect mice behavior in the OF test. (A) Initially, mice were positioned at the center of the OF arena, and their behavior was monitored and analyzed over a 10-min period. The trajectories of 12-month-old WT and YAC128 mice, both in control groups and following long-term administration of CHZ, FA, and the CHZ-FA combination, are presented. The mean time spent in the border (B), intermediate (C), and central (D) zones was quantified for WT and YAC128 mice across the control, CHZ, FA, and CHZ-FA treatment groups. (E) The total distance traveled during the OF test was assessed for WT and YAC128 mice in the control, CHZ, FA, and CHZ-FA groups. Statistical significance was evaluated with ANOVA (n. s.). The number of mice tested per group (n): n = 8 WT Ctrl mice, n = 9 WT CHZ mice, n = 8 WT FA mice, n = 8 WT CHZ-FA mice; n = 8 YAC128 Ctrl mice, n = 8 YAC128 CHZ mice, n = 7 YAC128 FA mice, n = 7 YAC128 CHZ-FA mice.
CHZ-FA ameliorate recognition memory in YAC128 mice
Our past studies have indicated that YAC128 mice display cognitive deterioration in recognition memory using the NOR test. 46 In this study, we investigated the impact of chronic administrations of CHZ and FA, both alone and combined, on object recognition in YAC128 mice and their WT controls using the NOR task (Figure 7). This trial was conducted the day following the OF test, ensuring that the experimental animals had already adapted to the test arena (Figure 1). On the initial day of this trial, a pair of matching objects were positioned in opposing sectors of the OF setup. During the following day, one of the known items was exchanged for an unfamiliar item (Figure 7A). On the start day of the test, all groups displayed alike preferences for the identical items (Figure 7B). On the following day, YAC128 Ctrl mice exhibited notably reduced interest in the new item relative to WT Ctrl mice (* p < 0.05; Figure 7C). Additionally, YAC128 Ctrl mice demonstrated notably reduced frequency (* p < 0.05; Figure 7E) in approaching the new item relative to WT Ctrl mice. Interestingly, we observed no significant differences in the distance traveled among control groups of WT and YAC128 mice throughout the initial (p = 0.12) and following (p = 0.24) days of the NOR test, indicating no motor decline in YAC128 mice at this disease stage in the NOR test. Long-term administration of CHZ-FA significantly (** p < 0.01) reduced the latency to novel object in WT mice (Figure 7D). Otherwise, chronic administrations of CHZ, FA, and the mixture of CHZ and FA had no effect on the performance of WT mice in the NOR task (Figure 7). In YAC128 mice, exposure to CHZ increased the frequency of interactions with the novel object (Figure 7E). FA treatment alone did not mitigate the recognition memory impairment reported in YAC128 mice (Figure 7). Nevertheless, the CHZ-FA combination notably enhanced the recognition memory in YAC128 mice (Figure 7C and E). Specifically, YAC128 mice treated with the CHZ-FA combination demonstrated significantly higher interest in the new item (* p < 0.05; Figure 7C) and notably increased frequency of visits to the new item (* p < 0.05; Figure 7D) relative to YAC128 controls.
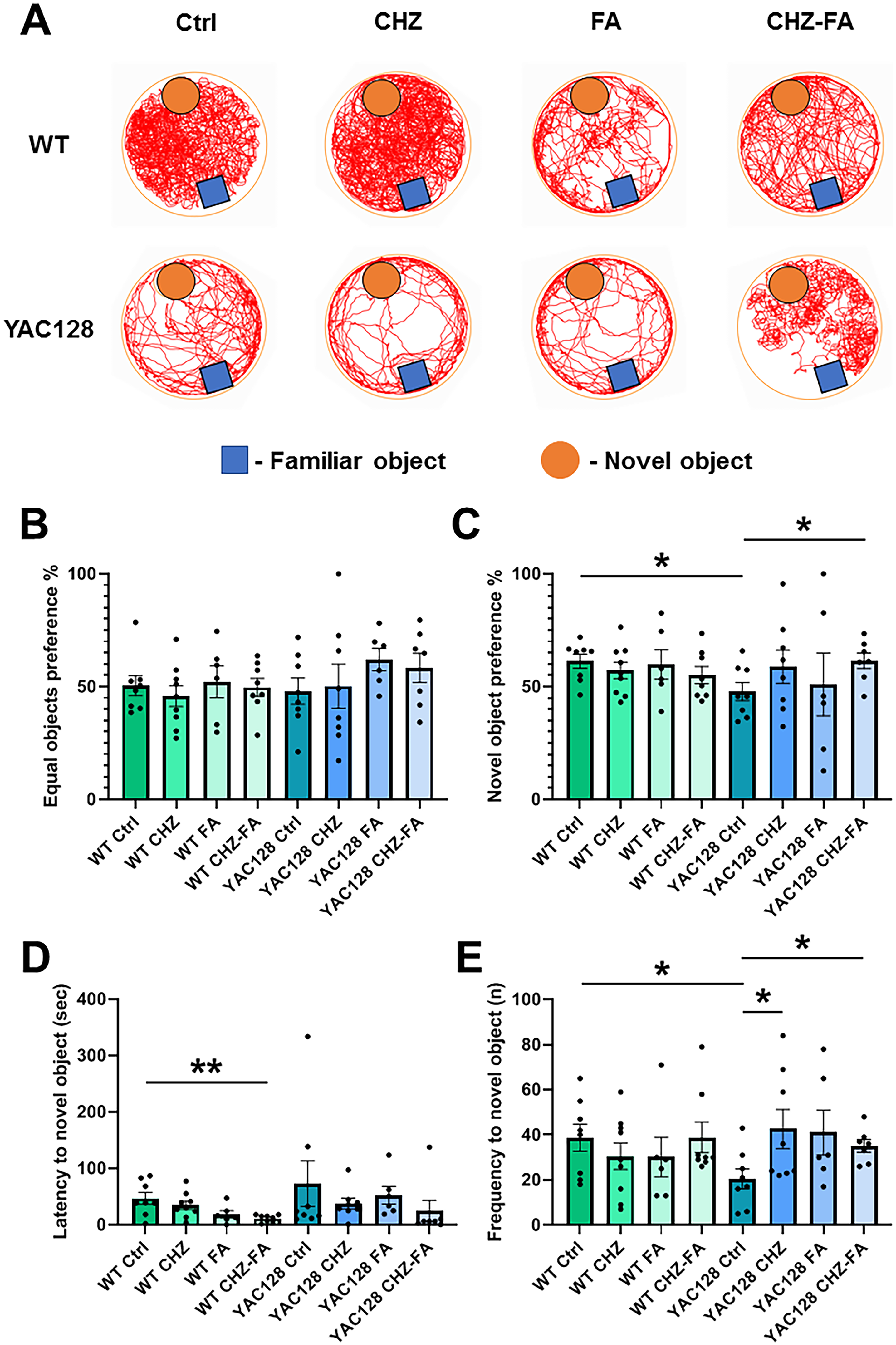
The CHZ-FA combination enhances recognition memory in YAC128 mice. (A) On the initial testing day, mice were positioned in the center of the OF arena between two identical objects. On the subsequent testing day, mice were placed in the center of the OF arena between one familiar object and one novel object. Mouse behavior was monitored for a duration of 10 min. The trajectories of 12-month-old WT and YAC128 mice, both in control groups and following long-term administration of CHZ, FA, and the CHZ-FA combination, are presented. The preference (%) for equal objects (B) was assessed during the first day of testing. On the second testing day, the preference (%) for novel objects (C), latency to approach the novel object (D), and frequency (n) of visits to the novel object (E) were analyzed in WT and YAC128 mice across the control, CHZ, FA, and CHZ-FA treatment groups. Statistical significance was evaluated with ANOVA (* p < 0.05; ** p < 0.01). The number of mice tested per group (n): n = 8 WT Ctrl mice, n = 9 WT CHZ mice, n = 6 WT FA mice, n = 8 WT CHZ-FA mice; n = 8 YAC128 Ctrl mice, n = 8 YAC128 CHZ mice, n = 6 YAC128 FA mice, n = 7 YAC128 CHZ-FA mice.
CHZ-FA combination mitigates depression-related behaviors in YAC128 mice
Preceding investigations have indicated that YAC128 mice exhibit depressive-like symptoms, as evidenced by their performance in the TST, 47 forced swimming test (FST) and sucrose consumption test (SPT). 48 This study aimed to evaluate the impact of CHZ and FA, both alone and combined, on mood fluctuations in YAC128 and WT mice using the TST task (Figure 8). In the TST, mice were secured by their tails using adhesive tape on a holder positioned 40 cm above a horizontal surface. The immobility time was analyzed in three intervals: 0–120 s, 120–240 s, and 240–360 s over a 6-min test period (Figure 8A). YAC128 Ctrl mice exhibited significantly greater immobility time relative to WT controls across all examined intervals (Figure 8). Long-term i.p. injections of CHZ and FA, both alone and combined, had no effect on the performance of WT mice in the TST (Figure 8). Moreover, chronic CHZ and FA exposure separately did not alter TST readings in YAC128 mice (Figure 8). Notably, the mixture of CHZ and FA markedly enhanced TST performance in YAC128 mice, as indicated by reduced immobility time in all analyzed intervals compared to YAC128 Ctrl littermates (Figure 8B, C and D).
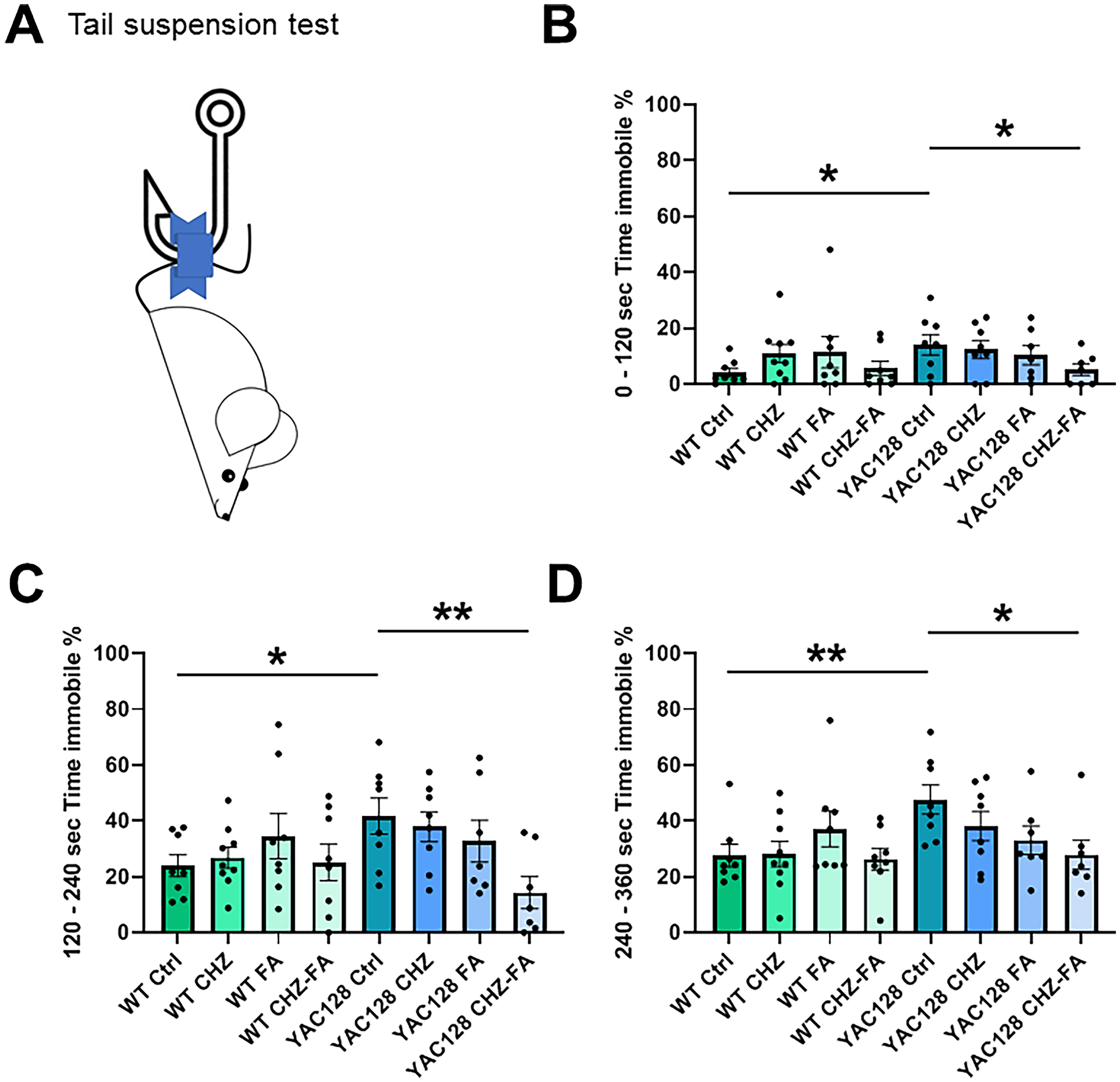
The CHZ-FA combination mitigates depressive-like behavior in YAC128 mice. (A) Experimental design of the TST. The mean duration of immobility was quantified during the intervals of 0–120 s (B), 120–240 s (C), and 240–360 s (D) of the TST in WT and YAC128 mice across control, CHZ, FA, and CHZ-FA treatment groups. Statistical significance was evaluated with ANOVA (* p < 0.05; ** p < 0.01). The number of mice tested per group (n): n = 8 WT Ctrl mice, n = 9 WT CHZ mice, n = 8 WT FA mice, n = 8 WT CHZ-FA mice; n = 8 YAC128 Ctrl mice, n = 8 YAC128 CHZ mice, n = 7 YAC128 FA mice, n = 7 YAC128 CHZ-FA mice.
Discussion
In this investigation, we conducted a comprehensive assessment of locomotor and cognitive-affective functions, involving measures for motor skills, anxiety, memory performance, and emotional well-being, in the YAC128 mouse model following chronic administration of the SK channel positive modulator CHZ, both independently and in combination with FA (Figure 1). Our findings indicate that both CHZ alone and the combination of CHZ and FA exerted comparable beneficial effects on cerebellar-specific deficits, such as PC electrophysiology and movement execution in YAC128 mice. Nevertheless, solely the mixture of CHZ and FA significantly alleviated depressive-like behavior and notably enhanced recognition memory in YAC128 mice, whereas CHZ alone did not. These findings indicate that a combined treatment targeting both locomotor and cognitive-affective manifestations is essential for the effective management of HD.
Comparable effects of CHZ and CHZ-FA on cerebellar impairments in YAC128 mice
The YAC128 murine model utilized in this research exhibits progressive motor decline beginning at 6 months of age, becoming fully manifested by 12 months, as evaluated by the BW test. This decline is characterized by increased durations to cross beams and a higher footslip count relative to WT controls.17,49 Notable disruptions in PC electrophysiological activity commence at 6 months of age in YAC128 mice (Figure 2C), with a notable reduction in cerebellar PC count observed by 12 months (Figure 4B). Previous research has indicated that YAC128 mice display anxious behavior, recognition memory decline, and depressive-like states by 9 months of age.46–48
This study investigated the impact of chronic administrations of CHZ, FA, and the mixture of CHZ and FA on locomotor, and cognitive-affective deficits in YAC128 mice. The dose and treatment duration of CHZ and FA utilized in these studies were chosen based on previous in vivo research with this compound and the well-established cumulative effects of FA.38,39,50,51 Additionally, the effects of these treatments on PC electrophysiology and morphology were examined. CHZ is an activator of SK channels capable of crossing the blood-brain barrier (BBB).52,53 SK channels are known to influence PC pacemaking activity. 27 Activating SK channels through pharmacological means could help ameliorate impaired PC electrophysiology and enhance motor performance in movement disorders. In fact, CHZ has demonstrated the ability to normalize firing variability and pacemaking rate in PCs, which enhances motor activity in various mouse models of movement disorders.17,28,33,34,45 This study also observed that CHZ facilitated the restoration of firing precision in YAC128 cerebellar PCs (Figure 3C) and enhanced performance in the BW test (Figure 5). Furthermore, the number of PCs was ameliorated in YAC128 CHZ mice (Figure 4B).
Nevertheless, SK activators have been shown to reduce PC firing rates in rodents,29,37–39 this effect was also observed in WT and YAC128 PCs in this study (Figure 3B). To mitigate this, FA was employed as a result of its defensive properties against the toxic effects of glutamate and oxidative stress in the cerebellum.38,39,54,55 In WT mice, FA increased PC firing variability (Figure 3C) and worsened movement capability in BW (Figure 5). Furthermore, FA elevated spiking frequency in YAC128 PCs (Figure 3B) and enhanced the number of cerebellar PCs in YAC128 mice (Figure 4B), although it had no effect on the motor activity in YAC128 mice (Figure 5).
The combination of CHZ and FA restored the CHZ-mediated reduction in spiking rate in both WT and YAC128 PCs (Figure 3B) and improved spiking precision in YAC128 PCs (Figure 3C). The mixture of CHZ and FA also restored motor activity in YAC128 mice on the smallest beam (Figure 5G). None of the applied treatments affected the motor performance of WT and YAC128 mice in the OF task, as the distance moved was equivalent across the experimental subsets (Figure 6E).
In conclusion, this study demonstrates that both CHZ alone and in combination with FA exert comparable beneficial effects on the cerebellar PC electrophysiology and motor activity in YAC128 mice, indicating their comparable efficacy in addressing cerebellar impairments.
The mixture of CHZ and FA improves recognition memory in YAC128 mice, unlike CHZ alone
Previous research has indicated that YAC128 mice displayed reduced ability to recognize objects, as evidenced by their performance in the NOR test. 46 In the current study, YAC128 mice demonstrated a reduced interest in new item on day two of testing relative to WT controls (Figure 7C). Additionally, YAC128 mice exhibited lower frequency of interactions with new item (Figure 7E) relative to WT mice. This study investigated the effects of CHZ and FA, both alone and together, on the observed recognition memory decline in YAC128 mice.
Limited data exist regarding the impact of CHZ on cognitive functions. A preceding investigation on murine model of Alzheimer's disease showed that CHZ improved visuospatial function in the Morris water maze (MWM). 56 However, our previous research on SCA2-58Q mice showed that CHZ had no effect on the spatial memory. 38 Moreover, CHZ did not influence NOR performance in SCA3-84Q mice. 39 In this investigation, CHZ restored the frequency of interactions with novel object in YAC128 mice (Figure 7E) in comparison with YAC128 controls. However, CHZ did not influence the interest for the new item on day two of testing in YAC128 mice (Figure 7C).
FA supplementation has been shown to impact memory. Previous studies on rodents have shown impaired visuospatial and recognition memory in folate-deficient mice in comparison with controls. 57 Furthermore, FA intake improved memory in patients with mild cognitive impairment. 58 However, our previous research indicated that FA had no influence on the visuospatial memory capabilities in SCA2-58Q mice as measured by the MWM test, 38 nor did it enhance the ability to recognize objects in SCA3-84Q mice in the NOR task. 39 Additionally, FA did not affect this ability in YAC128 mice as assessed by the NOR task in the current study (Figure 7).
Interestingly, the mixture of CHZ and FA showed a beneficial effect on MWM performance in SCA2-58Q mice in our recent study. 38 In our next study, the CHZ-FA combination increased the interest in new items and decreased the time to approach new item in SCA3-84Q mice. 39 In the present study, the mixture of CHZ and FA also enhanced the interest for a new item (Figure 7C) and increased the number of interactions with a new item (Figure 7E) in YAC128 mice. Therefore, the mixture of CHZ and FA, unlike the CHZ alone, ameliorates the ability to recognize objects in the NOR assay in YAC128 mice.
The CHZ-FA combination exerts beneficial effect on depressive-like symptoms in YAC128 mice
YAC128 mice are widely recognized for exhibiting depressive-like behaviors which was observed in different behavioral tests, including the TST,47,59 FST60,61 and SPT. 48 In the current investigation, YAC128 mice demonstrated significantly longer immobility periods in the TST test compared to WT controls (Figure 8). This study aimed to evaluate the impact of CHZ and FA, both independently and together, on depressive-like symptoms in YAC128 mice.
Further investigation is warranted to elucidate the antidepressant mechanisms of SK positive modulators, such as CHZ. Previous studies in other polyQ expansion disorders involving cerebellar degeneration have reported that CHZ administration improved depressive-like behavior in SCA2-58Q mice, assessed by the SPT, 38 and in SCA3-84Q mice, assessed by the TST. 39 However, in this research, CHZ did not produce any notable effects on the TST performance in YAC128 mice (Figure 8).
Similarly, FA is recognized for its antidepressant properties in both rodents62–65 and humans,66,67 did not influence depressive-like behavior when administered alone in SCA2-58Q 38 or SCA3-84Q 39 mouse models. Here, we also demonstrated that FA administration did not affect the depressive-like behavior of YAC128 mice in TST (Figure 8). Notably, a convergent pattern emerges when examining combination therapy. The mixture CHZ and FA has previously been shown to ameliorate anhedonia in SCA2-58Q mice in SPT 38 and reduce immobility in SCA3-84Q mice TST. 39 In the present study, we observed a similar beneficial trend, where the CHZ + FA combination significantly reduced immobility time across all intervals of the TST in YAC128 mice (Figure 8B-D). Thus, while the specific behavioral outcomes vary across models and tests, the combination of CHZ and FA consistently demonstrates a positive effect on depressive-like symptoms, whereas the individual components show limited efficacy in polyQ disorders with cerebellar neurodegeneration.
Limitations of the study
While this study provides robust evidence for the therapeutic potential of the CHZ + FA combination in a YAC128 mouse model of Huntington's disease, several key limitations must be acknowledged to contextualize the findings. A direct investigation into the molecular pathways underlying the observed synergy—such as oxidative stress markers, BDNF levels, or inflammatory cytokines—was not conducted, leaving the precise mechanism speculative. While CHZ is an approved drug and FA has a well-established safety profile, their combined long-term use specifically for Huntington's disease represents a novel and untested clinical approach that requires future validation. In addition, the study focused on the contribution of SK channels to the regulation of Purkinje cell activity in the cerebellum; the potential side effects of the SK channel modulator CHZ on neuronal activity remain a subject for future investigation.
The study's methodological design introduces certain constraints. The exclusive use of female YAC128 mice, though methodologically justified to avoid male aggression, limits the generalizability of findings across sexes. The assessment of depressive-like behavior relied solely on the TST; a more comprehensive behavioral battery would provide a stronger phenotype. Future studies are needed to dissect the specific contribution of these distributed circuits to the therapeutic outcomes.
Footnotes
Acknowledgments
We appreciate the advice and suggestions from the members of the Laboratory of Molecular Neurodegeneration, and the administrative support provided by Anastasia V. Bolshakova. The authors would like to acknowledge the use of artificial intelligence tools in the preparation of this manuscript. Specifically, AI was utilized for language editing and rephrasing to enhance the clarity and coherence of the text.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by the Ministry of Science and Higher Education of the Russian Federation, (grant number 075-15-2024-548).
Declaration of conflicting interests
The authors declared the following potential conflicts of interest with respect to the research, authorship, and/or publication of this article: IBB is an Editorial Board member of this journal, but was not involved in the peer-review process nor had access to any information regarding its peer review.
Data availability statement
The raw and processed datasets utilized in this study can be obtained from the corresponding author, KSM (kivi5656@gmail.com), with reasonable inquiry.