
Abstract
Understanding and recreating biomechanical forces in in vitro cultures is integral to regenerating cardiac tissue and finding novel therapies for cardiovascular disease. Mechanical forces influence cardiac physiology from a microscopic cellular scale to the global function of the heart. Here, we will review the mechanical forces within the heart as understood clinically and evaluate in vitro device designs that mimic these mechanical forces at a cellular and tissue level. We will further follow the evolution of bioreactors toward recapitulating multiaxial mechanical forces as they are experienced in vivo and understand the current limitations associated with in vitro systems in the context of recreating in vivo cardiac physiology.
Impact Statement
Translating multidimensional mechanical forces within the myocardium, as understood by our clinical and biological knowledge of myocardial mechanics, into in vitro systems is imperative in designing better bioreactors that emulate cardiac function in health and disease. Our review will help readers bridge our bed-to-benchside understanding and implementation of myocardial mechanics through the evolution of cardiac bioreactors.
Introduction
Cardiovascular disease has dominated as the leading cause of morbidity and mortality throughout the world for many decades, with an estimated prevalence of over 620 million people worldwide. 1 Hence, understanding the pathophysiology of cardiovascular (CV) disease, recreating the tissue characteristics in vitro in health and disease, and regenerating cardiac tissue for engraftment are undisputed priorities for the research community. However, as one of the most complex tissues in the body, the myocardium poses unique challenges for recreating its physiology in vitro. The cardiomyocyte is the fundamental cellular unit of the myocardium that imparts contractile function to the heart. Unique to cardiomyocytes is the ability for spontaneous beating. The contractility characteristics, such as intensity and duration of contraction and relaxation, change as cardiomyocytes mature or become diseased. Additionally, cardiomyocytes consume the largest amount of energy in the body in the form of adenosine triphosphate due to their continuous commitment to maintaining cardiac output. This continuous beating in cardiomyocytes is highly regulated through a coordinated flux of various ions including calcium, sodium, and potassium, which not only potentiates contraction and relaxation but also facilitates molecular signaling in neighboring myocardial cells. At an organ level, these movements of ions behave as electrical currents that traverse from their originating cells in the right atrium and propagate toward the ventricles, followed closely by mechanical contraction of the myocardium, thereby generating a phenomenon called excitation–contraction coupling (ECC), which is integral to the optimal function of the heart.
At a cellular level, the composition of the myocardium is diverse and comprises endothelial cells, smooth muscle cells, and fibroblasts, each of which has cellular subtypes based on their molecular signature and function. Cardiomyocytes themselves only make up 70–80% of cellular volume and 25−35% of cell type in the myocardium. 2 Yet, the cardiomyocytes are regulated by the mechanical and molecular forces provided by neighboring myocardial cells that help organize their contractile apparatus and regulate their energy substrates during development and disease. Hence, replicating this cellular mélange in vitro requires a system that can mimic the milieu of molecular factors and mechanical forces. Over the past 20 years, bioreactors have evolved to mimic the myocardial environment, to uncover mechanisms of disease, and to facilitate the regeneration of cardiac tissue. This review highlights the progression of bioreactors as a more physiological in vitro representation in the context of the mechanobiological milieu experienced by cardiomyocytes. For ease of following along with the various terms used in both clinic and research contexts, please refer to Table 1, which is a compendium of common terms used throughout this review article.
Definitions of Common Terms Used in This Review Article

LV, left ventricle.
Left versus right ventricular myocardial forces
Myocardial motion of the left ventricle
Within the heart, the left ventricle (LV) is the workhorse that pumps oxygenated blood to the brain and the other organs. The force generated by the LV with every cardiac beat is equivalent to 5–10 N 3 , cumulatively generated by cardiac fibers (also known as myocardial fibers or myofibers), composed of cylindrically connected longitudinal cardiomyocytes. The myocardial fibers are arranged in three layers in the LV: the outermost epicardium, the midmyocardium, and the innermost endocardium, each possessing a unique orientation of cardiac fibers (Fig. 1A). Cardiac contraction occurs through the lengthening and shortening of these cardiac fibers, described as myocardial strain, 4 of which there are three forms: longitudinal, circumferential, and radial (Fig. 1B). Longitudinal strain describes myocardial shortening along the long axis of the heart (base to apex), while circumferential strain describes shortening along the short axis of the ventricle (Fig. 1B-i–iii).5–7 Lastly, radial strain represents the thickening of the myocardium from the endocardium to the epicardium (Fig. 1B-iv–v).4,8,9 Cumulatively, the three-directional deformations by the myocardium generate a global strain within the myocardium that contributes to the contractility of the heart. In addition to the linear strain generated by the myocardial layers, the myofibers also organize into helices to promote a “twisting” motion of the ventricle. The epicardial fibers form a left-handed helix, whereas the endocardium forms a right-handed helix, as shown in Figure 1C. Such opposing directional presentation of the fibers in the epicardium and endocardium results in clockwise motion at the LV base and counterclockwise motion at the apex (Fig. 1C-i–ii),8,10 which further augments the outward flow of blood from the heart.
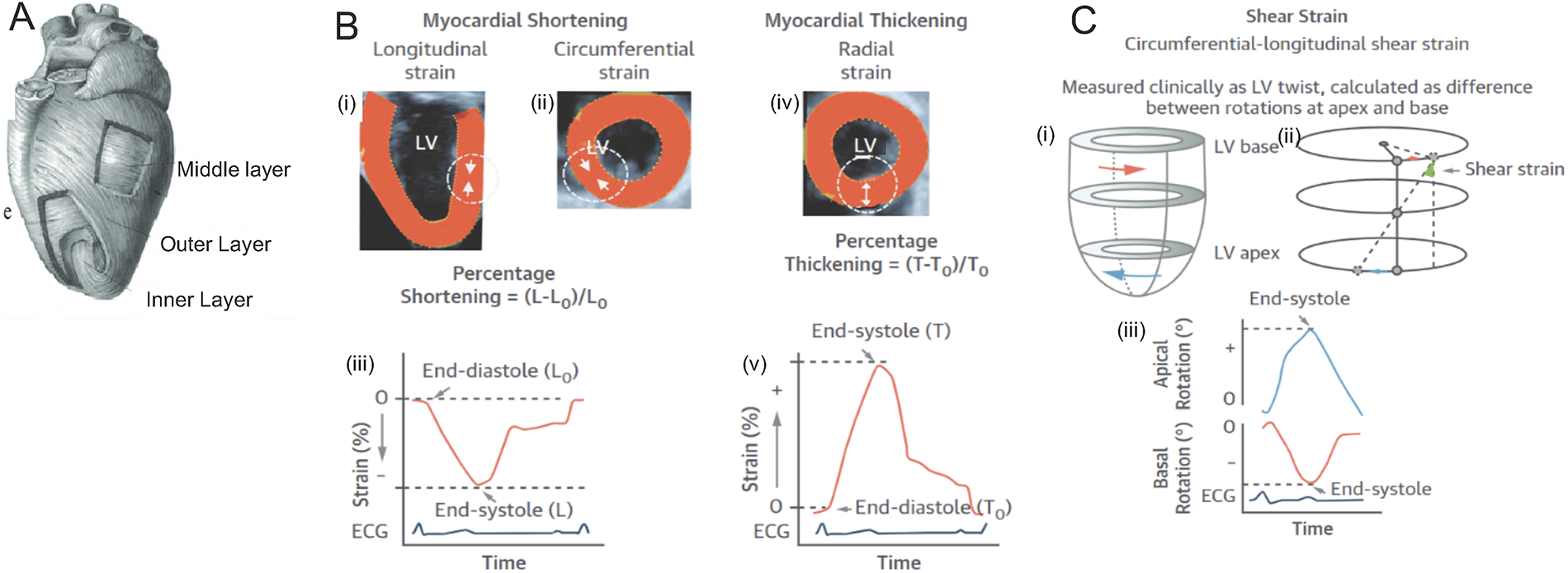
Schematic diagrams of the physiology of the heart.
At a cellular level, individual cardiomyocyte contraction generates stresses of 1 µN/mm2 with a 10% shortening. 11 However, globally, the myocardium generates a global strain of 18–22% and develops a peak stress of 22 mN/mm5. The summative forces generated by the myocardium result from the synchronous contraction of adjacent cardiomyocytes tethered together with adhesion molecules and chemically connected through gap junctions, which allow the flow of calcium and other ions. This combination of connections that organize into myofibers at a macroscopic level augments the longitudinal, circumferential, and radial force within the myocardium that expels blood upward and outward from the heart.
Myocardial motion of the right ventricle
Unlike the three-layered LV, the myocardial architecture of the right ventricle (RV) comprises two layers, the endocardium and epicardium, and lacks the middle layer of transverse fibers. Hence, RV outflow of blood is predominantly due to the longitudinal shortening of the RV free wall, as the RV base moves its annulus toward the RV apex. Therefore, while the LV has been described to work with a twisting motion, the RV motion is described as a slingshot. As the RV directs blood to the pulmonary vasculature, which has lower resistance and shorter destination for blood to travel, the RV does not generate the same forces as the LV. While the maximum RV strain ranges between 21% and 27%, the normal range of ejection fraction of the RV is lower than that of the LV (RV: 43–60%, LV: 50–70%). The anisotropic arrangements between the three myocardial layers—epicardium, midmyocardium, and endocardium—in the LV help generate a greater cardiac output than the RV, which has only two layers.12–17 Mathematical modeling of circular and longitudinal fibers alone, without helical fibers in the LV, could not achieve the cardiac output seen clinically. 18 Both LV and RV, however, share an intraventricular septum that has a three-layered myocardial architecture and synchronizes cardiac output between the ventricles. Studies show that the intraventricular septum contributes as much as 80% of the RV cardiac output. 19 As LV dysfunction due to ischemic heart disease is significantly more common than RV dysfunction, the LV myocardium is commonly used to develop in vitro translational models. However, the RV also has a unique disease pathophysiology, associated with both intrinsic myocardial disease and translated through its association with the LV, which is described in extensive detail in other literature reviews.6,8,20,21
Myocardial strain in disease
Myocardial strain is strongly associated with the heart’s function and reflects myocardial abnormalities with high sensitivity. In many cardiac diseases, abnormalities in cardiac strain precede the more evident decrease in contractility or cardiac output of the heart. Takahashi and their team found an inverse relationship between longitudinal myocardial strain and multiple cardiac risk factors, such as hypertension and dyslipidemia. 22 Furthermore, strain improvement corresponds to treatment responsiveness in multiple cardiac diseases, including amyloidosis 23 and secondary cardiomyopathies. 24 In a clinical setting, strain has been used to determine the timing of initiation of heart failure therapy for patients who are receiving chemotherapy so as to preserve myocardial function during ongoing chemotherapy. 25 Overall, strain imaging through various imaging modalities is quickly becoming an early, reliable marker of cardiac function.
Mechanical forces and anisotropy in health and disease
Cardiac physiology is governed by the laws of physics and fluid dynamics. The ventricles fill with blood during myocardial relaxation, or diastole. The outward force generated by the blood filling the ventricle promotes the myocardial fibers to lengthen. Much like a stretching rubber band, the fibers lengthen to a point where the intrinsic tension within the myocardial fibers forces the myocardial fibers to stop elongating. The intrinsic cardiac electrical stimulus forces cardiac fibers to contract through ECC, further shortening the myocardial fibers and promoting cardiac contractility.
Frank–Starling law
The contraction of the myocardium relies upon the lengthening of the cardiomyocyte during diastole, whereby a greater stretching of the cardiomyocytes generates a more vigorous contraction. At a physiological level, this allows for a beat-by-beat modulation of cardiac output through increased myocardium contractility when the ventricles fill with larger volumes of blood, as with exercise. This is the Frank–Starling relationship that forms the cornerstone of cardiac physiology. Frank–Starling curves exemplifying the myocardium in health versus sickness are distinct, and within the diseased myocardium, they are different for different types of cardiomyopathies (Fig. 2).
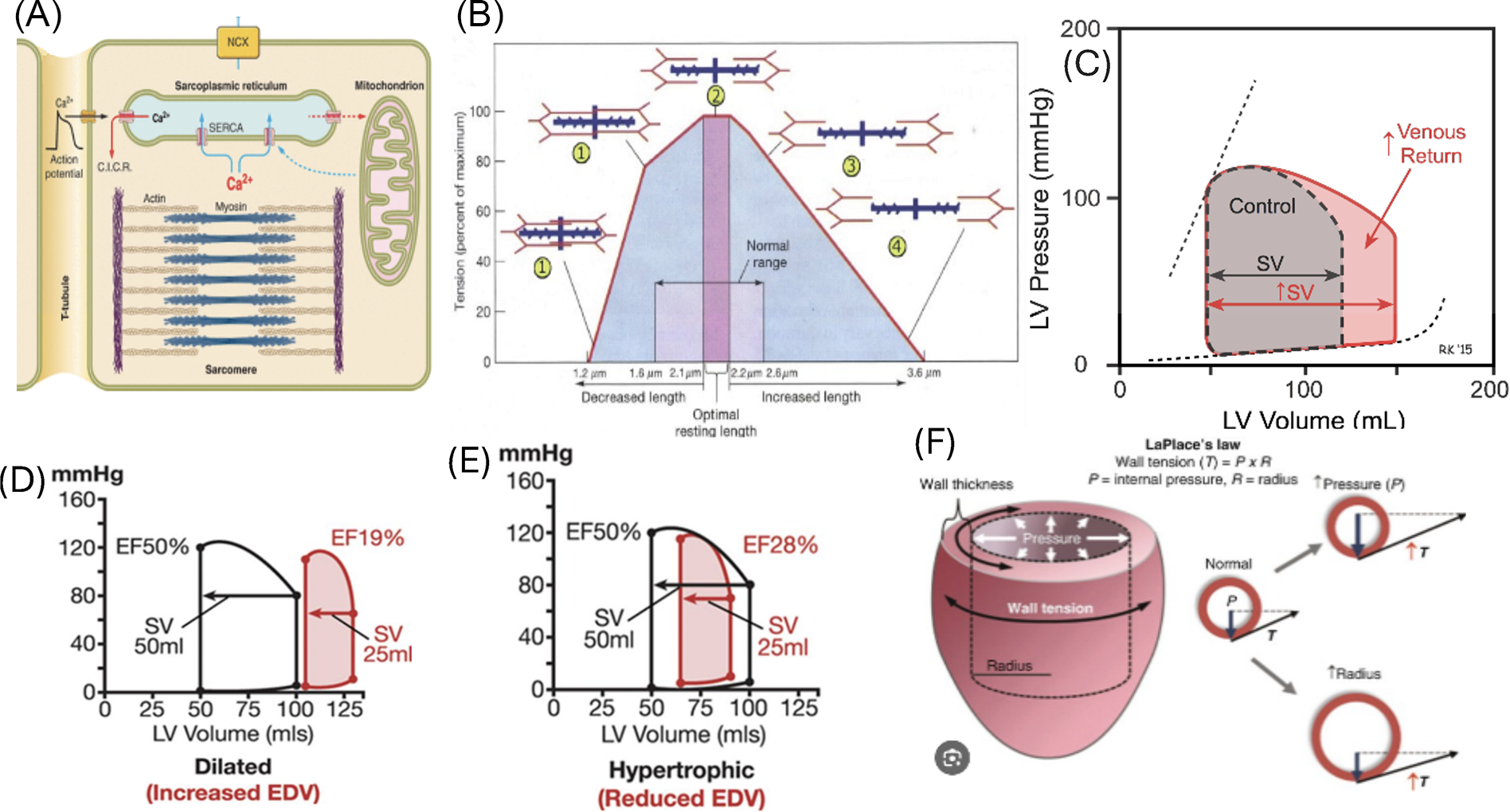
The cellular mechanobiology of the Frank–Starling relationship is driven by intra- and extracellular forces. At a cellular level, mature cardiomyocytes have a tubular morphology with a long-axis to short-axis ratio of 7:1. 26 Cardiomyocytes contract through sarcomeres (Fig. 2A) that are oriented along the longitudinal axis to facilitate maximal cardiac contraction.7,27–29 There is an optimal length between sarcomeres where the tension in the muscle fiber generates the maximum force, as shown in Figure 2B. If the sarcomeres are too close or far apart, the force of contraction decreases. As shown in Figure 2C, a healthy heart has optimal LV volumes to facilitate maximal interaction of proteins within the sarcomeres, corresponding to position “2” of the sarcomere length from Figure 2B. When venous return increases, as it does with exercise, the additional stretching of the sarcomeres further allows increased cardiac output, as the sarcomeres stretch to position “3” in Figure 2B.
In disease, sarcomeres rearrange to maintain maximum cardiac output and optimize the Frank–Starling relationship. In volume overload or dilated cardiomyopathy, where the LV volumes increase, sarcomeres are overstretched, as represented by position “4” in Figure 2B, and rearranged in series along the cardiac fibers. This produces a Frank–Starling curve represented by Figure 2D, where the overstretched sarcomeres in a large ventricle reduce cardiac output.
In response to pressure overload where the LV myocardium hypertrophies reducing ventricular volumes, new sarcomeres are arranged in parallel, increasing wall thickness. In other forms of cardiac hypertrophy, including genetic hypertrophic cardiomyopathies, the overall assembly of the sarcomeres is disorganized due to abnormalities in sarcomeric proteins. This causes the overall contractility to decrease, thereby decreasing cardiac output (Fig. 2E).
Laplace’s law
Laplace’s law is used to estimate myocardial wall stress related to intraventricular pressure, radius of the ventricle, and wall thickness. Wall tension or wall stress is the amount of counterforce or pressure the heart muscle experiences during contraction. Dilated ventricles encounter increased wall pressure due to reduced overlap between sarcomeric proteins that decreases ease of contractility. Similarly, thinner myocardial walls experience increased wall stress due to a decrease in sarcomeric units arranged in parallel to promote contraction. Hence, myocardial wall stress is directly proportional to intraventricular pressures and size, but inversely proportional to myocardial wall thickness. Increasing wall tension further increases oxygen demands on the heart, thereby exacerbating myocardial disease.
The various parameters governing wall tension are influenced by the type of myocardial disease. For example, the integrity and thickness of the myocardial wall rely on the composition of the extracellular matrix (ECM), which can be different in diseased and healthy myocardium, and which in turn influence the tissue’s cellular signaling and tensile properties. 30 The ECM houses a variety of proteins, collagens, proteoglycans, growth factors, and receptors (i.e., integrins), and the composition of these various biomolecules and their interaction with their surrounding cells influence the remodeling of the myocardium in disease.31,32 Noncardiomyocyte cells such as cardiac fibroblasts further influence ECM composition based on circulating signaling factors primarily driven by inflammatory cells that are upregulated in cardiac disease. The complexity of ECM and its multiple interactions with cardiac cells are not explored in this review. However, we address the interactions of these receptors with sarcomeric proteins, such as actin filaments and protein adaptors, involved in mechanotransduction pathways relevant to the bioreactors.33,34 In addition, we examine various genetic and protein markers of cardiomyocytes at different stages of cardiac development.
Markers for in vitro evaluation of cardiomyocyte maturation and function
Achieving a mature cardiomyocyte phenotype in vitro is essential for its inclusion in models mimicking its various physiological functions. Studies over the past two decades have demonstrated that implementing mechanical stretch facilitates cardiomyocyte maturation. Cardiomyocyte maturation can be tracked with the help of well-established cardiac markers such as NKX2.5, GATA binding protein 4 (GATA4), Troponin T, and T-box 5 (TBX5), especially when immature cardiomyocytes derived from induced pluripotent stem cells (iPSCs) are used.35–39 Moreover, multiple studies use these cardiac markers with other endpoints to evaluate the degree of maturation after differentiation. Mature cardiomyocytes differ from neonatal cardiomyocytes, most importantly in their ability to contract. Markers such as sarcomeric length, presence of α-actinin, β-myosin, Z-bands, connexin43 (Cnx43), sarcomere alignment, and cardiomyocyte orientation and shape highly influence the stress induced by mature cardiomyocytes and are used to indicate whether the cardiomyocytes have matured properly.40–49 These various cardiac markers become more relevant and apparent at different stages of cardiac maturation and can be characterized as either early, mid, or late markers. A comprehensive overview of these markers is provided in Table 2.
Biological Markers Associated with Differentiation and Maturation of Cardiomyocytes

Mimicking the myocardial mechanical environment in vitro
In vitro, bioreactors strive to implement the multidimensional forces experienced physiologically by cardiomyocytes with varying success. 50 A steady progression from devices that implement simple static or dynamic uniaxial strain to newer devices that impose more intricate biaxial or multiaxial force profiles has been made.50–53 Noncardiac devices have also implemented these forces with larger success. Torsion or twist type of force experienced in the biomechanical milieu of the LV, has been reported in tendon research. 54 The focus of the next section is to highlight research efforts on biomechanical loading profiles that mimic the complex in vivo multidimensional forces experienced by cardiomyocytes and other cardiac cells in the LV, as mentioned in the first half of this review.
The directionality of forces is essential in defining force profiles as either anisotropic or isotropic, where anisotropic forces have unequal distribution of forces in each direction. Isotropic forces are equally balanced in all directions. 55 Force profiles within bioreactors can be achieved by varying the magnitude and direction of force implemented on the cell substrate or by adding curvature to the substrate.56,57 Biomimetic models of cardiac physiology and/or pathological conditions can be developed with bioreactors that can apply strain in one or multiple directions, with the possibility of varying the strain magnitudes in each direction.
Bioreactor device design
Since the first studies in the 1980s, bioreactors for cardiac cell culture have progressively evolved to represent various degrees of force profile complexity. We would like to refer the reader to a review article that summarizes the principles behind cardiac cell mechanotransduction in parallel with the evolution of stretch bioreactor systems to study mechanobiology. 53 The next sections describe bioreactor systems ranging from devices that apply uniaxial static strain to ones introducing multiaxial dynamic strain profiles, as well as the design principles and study outcomes for each type. These devices are summarized in Figure 3 and Table 3.
Evolution of Mechanotransducive Bioreactors and Their Effect on Cardiovascular Cells
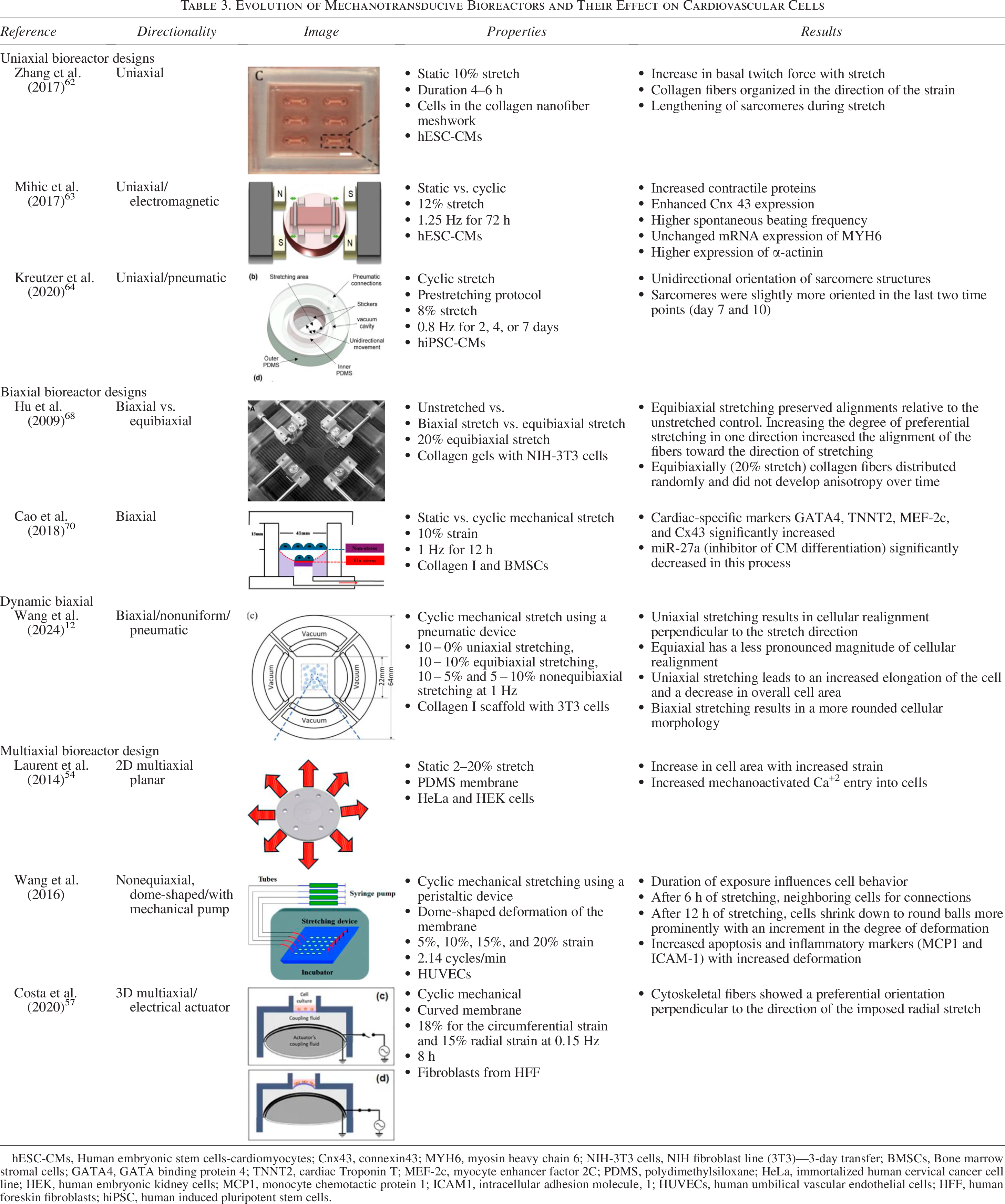
hESC-CMs, Human embryonic stem cells-cardiomyocytes; Cnx43, connexin43; MYH6, myosin heavy chain 6; NIH-3T3 cells, NIH fibroblast line (3T3)—3-day transfer; BMSCs, Bone marrow stromal cells; GATA4, GATA binding protein 4; TNNT2, cardiac Troponin T; MEF-2c, myocyte enhancer factor 2C; PDMS, polydimethylsiloxane; HeLa, immortalized human cervical cancer cell line; HEK, human embryonic kidney cells; MCP1, monocyte chemotactic protein 1; ICAM1, intracellular adhesion molecule, 1; HUVECs, human umbilical vascular endothelial cells; HFF, human foreskin fibroblasts; hiPSC, human induced pluripotent stem cells.

Devices used to simulate the cardiac biomechanical environment by applying various strain profiles. Devices
Uniaxial strain designs
Both static and dynamic strain regimes have been introduced in cardiac in vitro devices. This first section focuses on representative systems that apply strain in one axis only and are thus the simplest in design. Uniaxial strain systems are used to evaluate the effects of mechanical protocols on cardiomyocyte maturation as well as cardiac cell interaction.64–66
As a first example, bioreactor A has been designed to apply static stretch to human embryonic stem cell-derived cardiomyocytes (hESC-CMs) in conjunction with fibroblasts or marrow stromal cells (MSCs).
In the native myocardium, cardiomyocytes coexist with other cell types. One of these cells is the fibroblast, which is mechanoresponsive and contributes to the collagen architecture in the ECM. While fibroblasts remodel collagen, changes in matrix density affect the matrix’s volume, a phenomenon termed contraction. If external forces are applied to the construct, such as static strain, collagen fibers will organize in an oriented fashion and tension will develop within the matrix. 67
In bioreactor A, three-dimensional cell-matrix strips were attached to stainless steel posts on both ends and perfused with medium for cell maintenance. The tissue strips could compact and maintain their length, which provided static tension to the construct. 58 The addition of fibroblasts and MSCs aided in tissue contraction. Static stretch was shown to result in higher cell alignment and sarcomere lengths compared with unstretched controls.
In order to create a more complex force profile and further simulate the in vivo environment, dynamic strain can be applied. Cell-seeded constructs or substrates are stretched in intervals, usually with the use of an electromechanical or pneumatic device. Studies have shown that the use of dynamic strain leads to a superior resemblance of myocardial tissues. For example, the previous study included constructs that were subjected to uniaxial dynamic cyclic stretch, which resulted in higher moduli and expression of cardiac maturation markers, such as ventricular myosin light chain 2 and atrial natriuretic peptide, compared with statically strained samples. 58
Bioreactor B is an example of uniaxial dynamic cyclic stretch on hESC-CMs. 59 In this system, noncontact electromagnetic forces are applied to uniaxially stretch cell-seeded gelatin scaffolds from both ends. Stretch can be applied intermittently, with constructs cycling between stretched and relaxed states. 59 Parameters such as the extent, frequency, pattern, and duration of stretch could be varied. Cyclic stretch led to more successful maturation compared with static conditions in these 3D constructs, as evidenced by cardiac markers such as Cnx43 and α-actinin, as well as higher beating frequencies. 59
Bioreactor C represents an example of a pneumatically controlled device for uniaxial cyclic strain application. 60 It utilizes vacuum to stretch the substrate rather than an electromechanical actuator. Computational simulations determined the bioreactor’s dimensional parameters so that truly uniaxial strain could be applied. When human induced pluripotent stem cells-derived cardiomyocytes were cultured in the pneumatic bioreactor, cyclic strain facilitated proper sarcomere orientation and elongation with longer strain intervals. 60
Biaxial strain designs
Biaxial strain devices, in which cells and substrates can be stretched in two directions, were developed, introducing an additional level of complexity.37,68,69 Studies have shown that aligned cardiomyocytes in a two-dimensional environment can distinguish between forces applied in different directions, namely, parallel or perpendicular to their long axis.27,70 Compared with uniaxial strain, biaxial strain profiles are generally more representative of biological conditions since different global strains are manifested in the myocardium, such as longitudinal, circumferential, radial, and torsional. As a result, cardiac tissues and cardiomyocytes in vivo are subjected to strain in more than one direction. 10 Devices have been designed to apply strain that varies in magnitude and direction (nonequibiaxial), or same across directions (equibiaxial).
One design that has been utilized to study the microenvironment of the heart is the static biaxial model.61,71 It is particularly relevant to understanding how cardiac fibroblasts interact with collagen in the ECM and the subsequent effects on cardiomyocytes. As mentioned above in example A, collagen remodeling is orchestrated by fibroblast cells, and static strain has been employed to study the effects on cellular response and collagen architecture. A bioreactor designed for the application of biaxial strain is 3D. 61 Its design allows for the visualization of three-dimensional collagen architecture and tissue properties in a nondestructive, real-time fashion, in order to study how fibroblasts respond to multiple mechanical stimuli and deposit and remodel collagen. Varying types of static biaxial strain applied on cruciform-shaped, fibroblast-seeded collagen gels resulted in changes in collagen alignment. Nonequibiaxial strains, that is, higher strains in one direction, were shown to increase fiber alignment in the direction of the strain. In addition, collagen fiber alignment was observed to depend on both cell culture time and region of the construct, and static biaxial stretch was found to result in more anisotropic tissue architecture and, thus, more biomimetic, as compared with uniaxial.
Biaxial strain devices have also been used to study the effect of this type of biomechanical stimulus on cell differentiation and function. Results obtained with bioreactor 3E show that cyclic biaxial strain can facilitate mesenchymal stem cell differentiation into cardiomyocyte-like cells. 62 In this bioreactor model, bone marrow-derived mesenchymal stem cells were seeded on collagen I-coated, flexible membranes, which were biaxially strained through a motor-controlled mechanism. The resulting tissue showed more differentiation into cardiomyocytes as compared with static controls, with increasing expression of cardiac-specific markers such as GATA4 and Cnx43 at the mRNA and protein level. 62 Additionally, the presence of miR-27a, a marker for decreased cardiomyocyte differentiation, also decreased in the experimental group. 62
An example of a pneumatically controlled bioreactor is illustrated in Figure 3F. This device has two independent pneumatic channels that allow for nonequibiaxial strain, but it also has the capability of applying uniaxial and equibiaxial strain, as well as live-cell imaging. 12 This low-cost platform is designed to evaluate cellular response to varying strain patterns. Two-dimensional cell orientation and morphology can be monitored live for the experimental duration while varying strain protocols are applied.
Multiaxial designs in 2D and 3D
Finally, designs have been proposed that can apply strain in multiple directions,27,72 either in planar (2D) or a three-dimensional fashion.
The “IsoStretcher”53,55 (system 3G) is a device that allows for the application of planar multiaxial strain on a cell-seeded polydimethylsiloxane (PDMS) membrane. Stretch of equal magnitude was applied in all radial directions, confirming the isotropic nature of the applied strain, while the displacement in the z-direction was negligible. The authors evaluated the mechanosensitivity of Ca2+ entry into HL-1 cells, derived from the atrial cardiomyocyte tumor lineage and exhibiting embryonic-like structural characteristics, while their functional phenotype is comparable with adult atrial myocytes. 73 The results from HL-1 cells stretched to 10% with the IsoStretcher underline the importance of strain-induced Ca2+ entry into the cells that was not observed under static control conditions.
Cardiac cell behavior on a two-dimensional surface may be different than in 3D. 74 To elucidate the effect of multiaxial, isotropic strain in a more tissue-biomimetic manner, the IsoStretcher was used on cardiomyocytes that were embedded in hydrogels of varying stiffnesses, forming a three-dimensional environment. It was found that stiffer hydrogels could better translate mechanical stretch to cells as compared with softer ones and enabled stretch-mediated Ca2+ entry into adult cardiomyocytes.
Multiaxial strains, especially 3D strains, are important to investigate as they highlight the impact of the z-direction. By adding a third dimension, the effect of more complex strain profiles can be evaluated. Additionally, by deforming a substrate and applying force in the z-direction, flexure can occur, which simulates the cardiac fiber layer interactions within the cardiomyocyte environment. 75 To that end, systems have been developed that apply strain in three dimensions by deforming the cell substrate in the x-, y-, and z-directions.
An example thereof is illustrated in Figure 3H. This device is composed of elastomeric membranes that are electrically deformable. 57 It explores the impact of the curvature of the substrate, which results in varying strain profiles in each direction, which is a more biomimetic representation of physiological conditions where cells experience anisotropic and inhomogeneous strain. 57 The unique aspect of this bioreactor is that it is modular, and the cell culture membrane actuation can be dynamically controlled by purely electrical means, without any fluidic system requiring external pumps or motors, as is the case with many existing systems in the literature. The authors utilized this model to study cytoskeletal fiber changes in fibroblasts. Through staining of cytoskeletal fibers, they observed a preferential orientation perpendicular to the direction of the radial strain, with a distinct difference compared with static controls. This can have implications for cocultures with cardiomyocytes, as studies have shown the effect of fibroblasts on the mechanoresponsiveness on cardiomyocyte orientation and shape. 70
Finally, a biomimetic model that aims to replicate both the multidimensional biomechanical and electrical cardiac environment is device in Figure 3.76 This platform is based on PDMS that is patterned with electrically conductive nanogrooves. Device I features silver nanowires embedded within its PDMS matrix, which allows for electrical signal transmission to the cardiomyocytes. Cardiomyocytes cultured on the nanogrooved array device and exposed to multiaxial strain showed increasing maturation markers as opposed to static controls. Specifically, the cardiomyocytes exhibited regular and synchronized beating, as well as longer sarcomeres and increased α-actinin and Cnx43 expression. The authors hypothesized that the presence of multiaxial strains likely results in uniform stress distribution, which can promote better organization on the cellular and ECM levels. 63
Current limitations and future directions
Myocardial strain is an aggregate of multiple anisotropic fiber layers and is augmented from a cellular to an organ level. The added level of complexity as we transition from a cellular to an organ level enhances the challenge of designing in vitro systems that recapitulate the biomechanical milieu in terms of force magnitude and directionality.
We have summarized multiple in vitro models to study the biomechanical environment of cardiac tissues to represent in vivo forces with increasing likeness. The majority of in vitro systems induce strain on one or two axes, mimicking simple unidimensional forces seen in vivo. 76 Adopting 3D biomimetic mechanical environments is necessary to recapitulate the ECM, where forces and cell–cell communication are multidimensional. This requires both biomaterials and bioreactor device efforts. Additionally, the torsional or circumferential forces observed in the heart continue to remain elusive in cardiac bioreactor systems, as current technologies have not been able to replicate these with accuracy. Developmentally, torsion contributes to the structural formation of the heart by providing mechanical support during cardiac looping. 77 Moreover, Cutrì et al. showed that torsion and cardiac abnormalities, such as alterations in contractility, preload, and afterload, are closely related. 78 Lastly, biological factors such as the ECM and endothelial cells also influence cardiac tissue by providing mechanical forces at a cellular level. While 3D-printed scaffolds and decellularized scaffold systems are evolving to replicate these complex systems successfully,79–81 they have not yet been integrated into mechanotransducive bioreactors for CV research. As various cardiovascular cells respond differently to mechanical forces, adapting bioreactors to study complex cardiovascular tissues will be necessary for future growth.
Authors’ Contributions
A.D.: Conceptualization, methodology, investigation, resources, data curation, visualization, and writing—original draft. L.K.: Conceptualization, methodology, validation, writing—original draft, writing—review and editing, visualization, and supervision. A.S.: Conceptualization, methodology, visualization, validation, writing—review and editing, and supervision.
Footnotes
Author Disclosure Statement
The authors have no conflicts or competing interests to disclose.
Funding Information
A.S. is funded by the American Heart Association Career Development Award #852875 and NIH K08 # K08HL161440.