
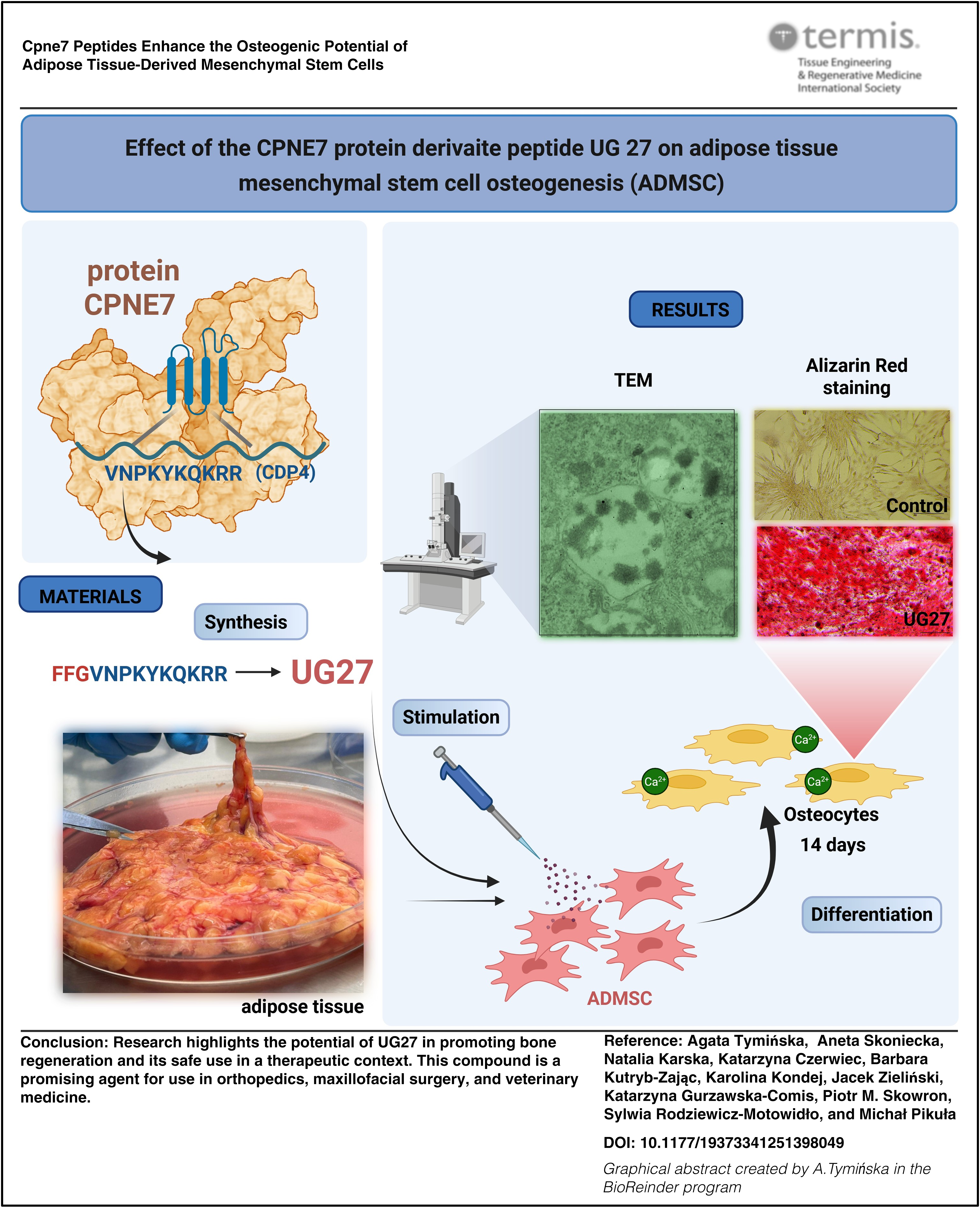
Abstract
Cartilage and osteochondral disorders pose an increasing clinical, economic, and social challenge. With an aging population, there is a need to develop innovative, nonsurgical strategies for treating defects, fractures, and other osteochondral disorders. Bone implants require surgical intervention, carry a risk of complications, are expensive, and do not always provide comfort to the patient. Alternatively, stimulation of bone regeneration using synthetic peptides is a promising and less invasive option in the treatment of trauma, orthopedics, craniofacial surgery, and dentistry. In the present study, we evaluated the biological effects of two peptides: the novel peptide UG27 and CDP4 derived from the protein Cpne7 (Copine 7), on the activity of adipose tissue-derived mesenchymal stem cells. Using labeling, differentiation, and imaging methods, we demonstrated the effect of UG27 on viability, biomineralization, extracellular matrix, and calcium salt growth in osteocytes. The peptides were immunologically safe and stimulated cell migration without showing any cytotoxic effects. The peptide, UG27, has an active connection with the biomaterial and is a promising compound in bone injury therapies.
Impact Statement
This study has significant translational potential, combining adipose-derived stem cells with an osteoinductive peptide featuring a controlled-release linker, leading to the repair of bone defects. The peptide promotes osteogenic differentiation and may aid osteocyte cultivation by stimulating a collagen- and calcium-rich extracellular matrix. This less invasive, personalized approach could accelerate healing compared to traditional bone grafts. These findings could benefit osteoporotic patients, those undergoing long-term steroid therapy, those with bone defects and fractures, and dental patients following tooth extractions.
Keywords
Introduction
Bone tissue has a unique ability to regenerate compared to most other tissues. However, in cases of large defects, trauma, infection, necrosis, or advanced age, the regenerative capacity may be impaired, which is why ongoing research and medical interventions aim to improve bone healing in such situations.
Data show that osteoporosis is a common disease 1 among people aged 50 and older, as many as 12.6% in the United States suffer from hip or spine osteoporosis. The incidence of this disease is increasing in women, while in men it remains largely unchanged.2,3 In 2019, as many as 128.7 million people (78.4 million women and 50.3 million men) suffered from osteoporosis. It is estimated that between 2030 and 2034, the global prevalence of osteoporosis will reach 263.2 million (154.4 million women and 108.8 million men). 3 Pharmacology in osteoporosis aims to reduce the rate of bone loss and prevent fractures. Agents such as bisphosphonates induce osteoclast apoptosis and affect osteoblasts, reducing the risk of skeletal fractures. 4 Several growth factors inducing osteogenesis, bone morphogenetic protein (BMP) and transforming growth factor β (TGF-β), as well as vitamin D, play a role in embryonic skeletal development and bone homeostasis. BMP2 and BMP7 have the best osteoinductive properties and have been approved for clinical use by the Food and Drug Administration. 5
In order to differentiate mesenchymal cells or induce them to secrete extracellular matrix (ECM), as indicated by literature data, short-chain bioactive peptides mapping intercellular signaling or recruiting host cells can be released in a controlled manner from a designed biodegradable biomaterial.6,7 Among the osteogenic peptides are those derived from type I collagen motifs, GFOGER, P15, KOD, DGEA, and BCSP1, which induce osteogenic differentiation in vitro and bone healing in vivo.8,9 Hormones such as parathyroid hormone stimulate osteoblast activity and increase bone density at the fracture site, leading to healing in the absence of unions. 10 Others, that is, osteogenic growth peptide—responsible for osteoprogenitor differentiation, leads to regulation of osteogenic markers, including mineralization. 11 Thrombin peptide 508 enhances proliferative differentiation and osteoblast chemotaxis. 12 Biomineralizing peptides such as dentin osphoprotein with Asp-Ser-Ser motifs strongly bind calcium ions and hydroxyapatite (HA). 13 In our studies, we focused on a peptide from the Cpne protein family, which is dependent on the intracellular Ca2+ growth level, is highly conserved, moves to the plasma membrane, binds phospholipids, and is ubiquitous in various eukaryotes. 14 One of them is Cpne7, a protein (633 amino acids) regulating the formation and differentiation of odontoblasts. 15
Our goal was to investigate the biological activity of adipose-derived stem cells (ADSC) in the UG27 peptide environment and to demonstrate their effect on the differentiation of mesenchymal cells into osteocytes, their effect on calcium ion secretion, and the formation of ECM in the form of collagen. The experiments also examined immunological safety and allergenicity.
Materials and Methods
Peptides
The CDP4 (Copine Derived Protein 4) peptide includes amino acid residues with the sequence VNPKYKQKRR (Val-Asn-Pro-Lys-Tyr-Lys-Gln-Lys-Arg-Arg). On the contrary, the peptide named UG27 has an additional three-amino-acid linker
Amino Acid Sequences of Designed Peptides with a Linker That Is Enzymatically Cleaved by MMP7

The peptide UG27 was synthesized on solid support (SPPS) using the Fmoc methodology, using the Liberty Blue automated microwave synthesizer of CEM Corporation with a constant flow of reagents in Rink Amide ProTide Resin with a deposition rate of 0.59 mmol/g (1.69 g, 1.0 mmol)
The UG27 peptide was purified by high-performance liquid chromatography (Supplementary Fig. S1A) in a reversed-phase system (chromatograph consisting of two pumps [K1001] and a UV detector [K-2001] coupled to a Gilson collector) using a semi-preparative Jupiter® Proteo-90-4-C8 column (Phenomenex) (21.2 × 250 mm). Chromatographic separation was performed in a linear gradient. The peptide mass was confirmed using the MALDI (BRUKER) MS technique (Supplementary Fig. S1B).
Isolation of ADSC
Subcutaneous ADSCs were isolated using the mechanical–enzymatic isolation method using 8 mM collagenase, based on the Zuk method. 17 After enzyme digestion, the tissue was filtered through a 100 µm strainer, and the cells were centrifuged twice for 8 min/362 g. Subsequently, the cells were subjected to erythrocyte lysis, centrifuged in PBS, and plated onto 75 cm3 bottles.
Material, which is medical waste, was obtained during elective surgery in the Department of Plastic Surgery, for which approval was obtained from the Independent Bioethics Committee for Scientific Research of the Medical University of Gdańsk (approval no. NKBBN/672/2019, NKBBN/672-65/2023).
Patients who had undergone chemotherapy or radiotherapy, patients with diabetes, or patients receiving steroid therapy were excluded. Demographic data, age, gender, smoking, diabetes, and body mass index were collected.
Cells were cultured in DMEM LG (Dulbecco’s modified Eagle medium low glucose—with 1 mg/mL glucose, Sigma Aldrich), supplemented with 10% fetal bovine serum (FBS) and antibiotics at 37°C, 5% CO2. The medium was changed every 2–3 days.
Cell viability (XTT test)
The XTT test was used to measure cell viability based on mitochondrial activity (Cell Proliferation Kit II; Sigma Aldrich, Mannheim, Germany) was used to measure cell viability based on mitochondrial activity. Cells (0.005 × 106) were seeded in wells of a 96-well plate in DMEM LG medium with 10% FBS and 1% P/S (penicillin/streptomycin). After 24 h, the medium was replaced with DMEM LG with 2% FBS and antibiotics, and treated with 0.01, 0.1, 1, 10, 25, 50, 100, and 150 μg/mL of UG27 and CDP4 peptides (in four replicates). The cells were incubated with the peptide for 48 and 72 h. In the case of 72-h cultures, after 48 h, the medium was replaced with a new one, adding a specific concentration of the peptide. After this time, XTT reagent was added, and after 4 h of incubation under standard conditions, the absorbance was read at 450 nm OD. The results were compared with the ADSC control without peptide in DMEM LG with 2% FBS (C—control; 100%). The positive control was ADSC cultured in DMEM LG with 10% FBS. The experiment was performed on four patients in four repetitions (n = 16).
Cytotoxicity assay
The cytotoxicity of compounds was measured in the ADSC culture supernatants by testing lactate dehydrogenase (LDH; Clontech, Takara, Japan) activity. The highest concentrations of peptides (50, 100, and 150 μg/mL) were used for the cytotoxicity tests. After 48 h of incubation, the filtrate was collected from above the cells and tested according to the manufacturer’s procedure. The results were compared with the DMEM LG 2% FBS control and the 10% FBS positive control, as well as Triton X-100, representing 100% cytotoxicity. The experiment was performed on four patients in four repetitions (n = 16).
Migration test (scratch test)
The effect of peptides on ADSC migration was analyzed after 24 h using a scratch test (Ibidi, Gräfelfing, Germany) on 3 cm diameter plates. Each of the four wells of the insert was seeded with 0.02 × 10 6 cells per 24 h in medium containing DMEM LG with 10% FBS and P/S. After removing the insert, mitomycin C (5 μg/mL; 2 h) was added to inhibit cell proliferation. The medium was changed to DMEM LG with 2% FBS with the addition of two selected peptide concentrations (10 and 25 μg/mL). After another 24 h, the cells were fixed with 3.7% paraformaldehyde and stained with 0.05% crystal violet. Unbound dye was rinsed with PBS. The effect of the peptide on cell migration was measured using Graphix software by measuring the area of the scratch not occupied by cells. The experiment was performed on five patients in four repetitions (n = 20).
Collagen production
ADSC were seeded into a 96-well plate at a density of 0.003 × 106 per well and cultured in standard DMEM LG medium with 10% FBS and P/S. Cells without peptide served as controls. Collagen synthesis after peptide stimulation was assessed after 14 days of culture. After 48 h, the medium was replaced with a new one, adding a specific concentration of the peptide. Cells were fixed with Bouin’s solution and stained with Direct Red 80 (Sirius Red) for 1 h with gentle mixing. The cells were washed with 0.01 N hydrochloric acid, and then the dye was dissolved in 0.1 N NaCl for 30 min. Absorbance was measured at OD 550 nm. The experiment was performed on four patients in four repetitions (n = 16).
Immunogenicity
The immunogenic potential of the peptides was tested on human peripheral blood cells isolated from “buffy coats” on Ficoll using a concentration gradient. Peripheral blood mononuclear cells were seeded at 1 × 106/mL of RPMI 1640 medium (P/S, 10% FBS)/well. They were stimulated with 10 μg/mL peptide and incubated under standard conditions for 24 h. The negative control was cells without peptide, and the positive control was cells stimulated with lipopolysaccharide (1 μg/mL). After incubation, cells were washed and labeled with antibodies (anti-CD3, CD4, CD8, CD25, CD69, CD71, and HLA-DR). After 30 min/room temperature in the dark, they were analyzed using CytoFLEX (Beckman Coulter, Marseille, France). The experiment was performed on four patients in four repetitions (n = 16).
Basophil activation test
The basophil activation test (BAT) was analyzed using the Beckman Coulter A17116 kit (Marseille, France), 2–3 h after blood collection. Peripheral blood from healthy volunteers (n = 5) was collected into ethylenediaminetetraacetic acid (EDTA) tubes. A total of 100 µL of blood was incubated separately with 20 µL of the UG27 (25 μg/mL) and 20 µL of PBS. Samples were stained using 40 µL of CD3, CD203c, and CRTH2 antibodies mixture. Then, the samples were incubated for 15 min at 37°C in a water bath. After the incubation, lysis and wash procedures were done. The samples were analyzed up to 2 h using CytoFLEX. Before the analysis, quality control and compensation for FITC, PE, and PC7 channels were performed. For each patient sample, negative and positive (anti-FceRI mAb and fMLP [N-formylmethionine-leucyl-phenylalanine]) stimulation controls were prepared. The cutoff values for the analyzed allergen were estimated as 5% of the negative stimulation control sample.
Gating strategy used in flow cytometry analysis of BAT. After erythrocyte lysis, leukocytes (forward scatte/side scatter) and, in the next step, lymphocytes with monocytes (CD3-) were gated. Within the CD3 population, basophils were identified using the markers CRTH2 and CD203c (Supplementary Fig. S6). A gate indicating activated basophils was particularly established for each patient (based on negative and positive controls). The whole procedure was performed according to the manufacturer’s instructions.
Differentiation of ADSC
ADSCs were seeded into 96-well plates at 0.005 × 106 for adipocytes and osteocytes and 0.08 × 106 for chondrocytes, cultured for 14 days in dedicated media (StemPro®—Adipogenesis/Osteogenesis/Chondrogenesis Differentiation Medium with supplement and MesenPRO RS™ Kit medium Life Technologies, USA). The media were changed every 2 days, and cells in MesenPro—a nondifferentiating medium—served as a control for differentiating cells. Cells were fixed with 4% paraformaldehyde and stained with oil red (adipocytes) and alizarin red (osteocytes). Quantitative analysis was performed. In adipocytes, dye was eluted with isopropanol and read at OD 490 nm. Osteocytes were incubated in 10% HCl, placed in a thermoblock at 85°C/10 min, and cooled on ice. To 100 µL of the filtrate, 10% ammonia water was added and measured spectrophotometrically at OD 450 nm. Papain proteinase enzyme (Papain Extraction Reagent, Sigma Aldrich, Mannheim, Germany), EDTA, cysteine HCl, sodium acetate, and PBS were added to the chondrocytes. They were incubated at 65°C/3 h. Sulfated glycosaminoglycans were quantified using a Blyscan assay (Blyscan Sulfated Glycosaminoglycan Assay Kit; Biocolor Ltd., Belfast, United Kingdom) according to the manufacturer’s procedure. Readings were taken at OD 656 nm (Supplementary Fig. S2). The experiment was performed on four patients in four repetitions (n = 16).
Analysis by transmission electron microscopy
ADSC at 0.01 × 106 were seeded into a 48-well plate. They were stimulated with peptide at a concentration of 25 µg/mL, changing the osteogenic medium every 2–3 days. After 48 h, the medium was replaced with a new one, adding a specific concentration of the peptide. Cells without peptide constituted the control. Cells were differentiated for 14 days. Samples were washed in PBS and flooded with 2.5% glutaraldehyde at 4°C/24 h. Sucrose cacodylate buffer was washed with fixative three times for 5 min and counterstained with 1% osmium tetroxide at 4°C/60 min. The samples were dehydrated in increasing concentrations of ethanol and propylene oxide. They were then impregnated with Poly/Bed® 812 (Polyscience, Warrington, PA, USA) epoxy resin and placed in gelatin capsules. They were polymerized for 2 weeks at 37°C. Ultra-thin sections (2–5 nm) were cut into slides. Slides were evaluated in a JOEL type JEM 1200EXII transmission electron microscope and photographed with a Samsung Electronics camera.
Measurement of nucleotide concentrations in cell cultures
Cells were cultured for 14 days in a 96-well plate in StemPro® Osteogenesis Differentiation Medium for control and stimulated with UG27 peptide. After 48 h, the medium was replaced with a new one, adding a specific concentration of the peptide.
Cells were then washed three times with Hanks′ balanced salt solution at 37°C to remove the high-protein medium. About 150 µL of 0.4 M HClO4 was added to the wells. The plate was immediately frozen at −80°C for at least 24 h. It was thawed on ice and refrozen twice at −80°C. After thawing, 100 µL HClO4 was transferred to the tubes. Centrifugation at 18,400 g, 10 min, 4°C, and the pellet was poured into 100 µL of 0.5 M NaOH. The supernatant was neutralized (to a pH of about 6) with 3 M K3PO4 for 15 min on ice and then centrifuged. The supernatant was frozen until the samples were analyzed. After thawing, it was centrifuged at 18,400 g, 10 min, 4°C, and analyzed using reverse-phase high-performance liquid chromatography. 18 The remaining protein in the well was flooded with 100 µL of 0.5 M NaOH and combined with the protein from the supernatant. Determination of protein levels was performed using the Bradford method. The experiment was performed on three patients in four repetitions (n = 12).
Statistical analysis
Data represent the mean ± standard error of measurement of four independent experiments (four replicates each). Comparisons between groups were made using the Mann–Whitney U test. Statistical significance was determined assuming a two-tailed distribution. Statistically significant results are denoted by *p < 0.05. To compare results within the same group at a single time point and within groups across time points, we performed analysis of variance (ANOVA) with Tukey’s standard deviation (HSD), analysis of differences between categories with 95% confidence intervals.
Statistical analysis was performed using Statistica 13.3, and for the graphical presentation of the results, the program GraphPad Prism 5.0 was used.
Results
Confirmation of ADSC phenotype according to International Society for Cell and Gene Therapy
Mesenchymal stem cells showed positive surface markers CD105, CD90, CD73, and CD13 and negative markers CD45, CD31, and CD19 (Supplementary Fig. S3). They could differentiate into adipocytes, osteocytes, and chondrocytes (Supplementary Fig. S2) and were characterized by adhesion to the substrate. The phenotype of ADSC was confirmed according to the guidelines of the International Society for Cell and Gene Therapy.19,20
Cell viability (XTT test) and cytotoxicity assay
The XTT test was used to determine cell viability after peptide stimulation at 48 and 72 h. The toxicity of the tested peptides was assessed using the LDH test at the highest selected concentrations. Statistically significant differences compared to the control group (cells cultured in medium with 2% serum), p < 0.05.
Analysis of cell viability after induction with CDP4 peptide showed an increase in mitochondrial activity compared to the control group without stimulation by 40% (p = 0.001) at a concentration of 10 µg/mL and by 43% (p < 0.001) at a concentration of 25 µg/mL after 48 h of culture. Increased mitochondrial activity was also observed at a concentration of 1 µg/mL after 72 h of culture with this peptide. The results were not statistically significant (Fig. 1).
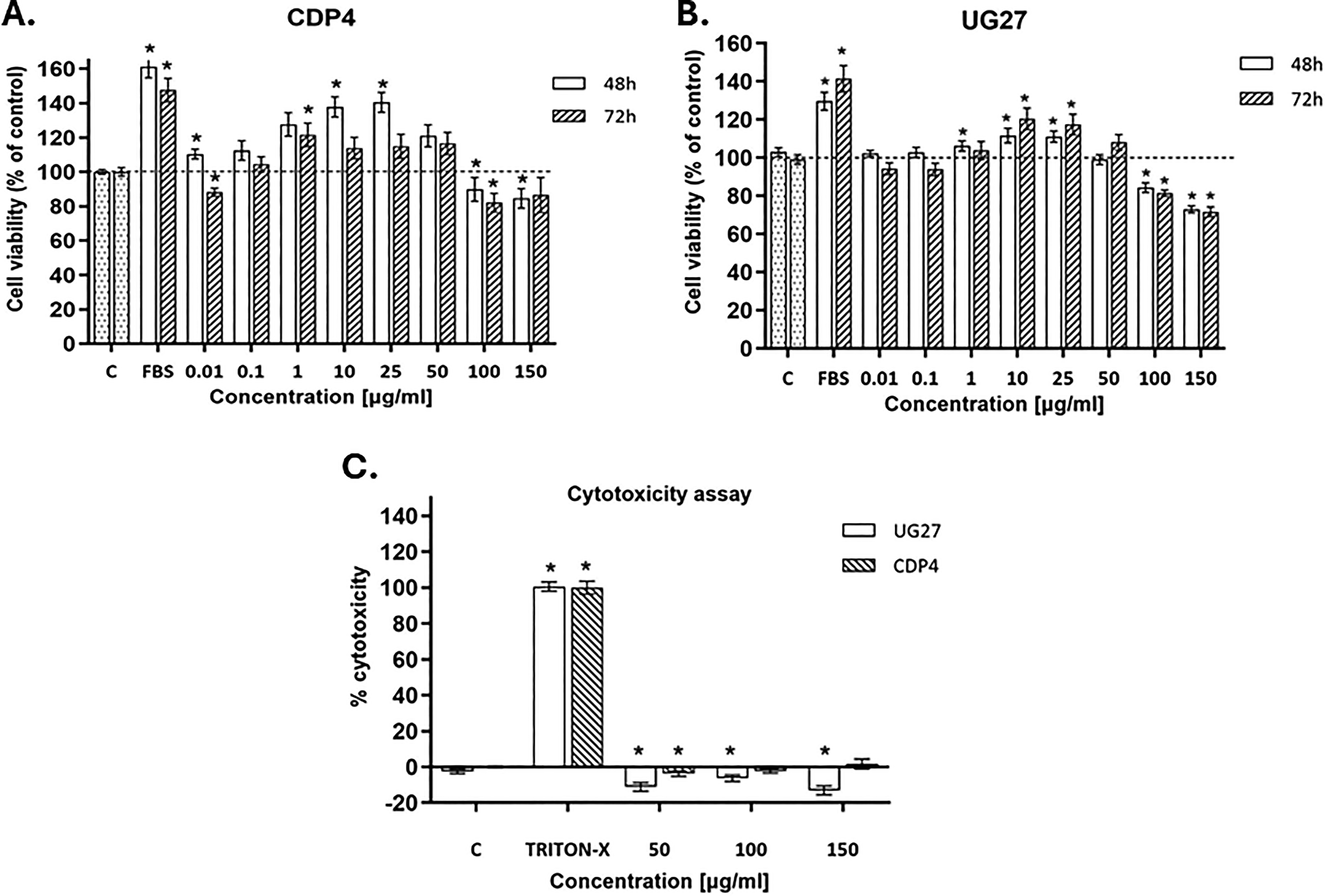
Analysis of adipose-derived stem cells (ADSC) viability and cytotoxicity against the tested peptides.
XTT analysis of cells after induction with the UG27 peptide showed a 10% (p = 0.001) increase in viability after 48 h and a 20% (p = 0.001) increase after 3 days of culture compared to the control group (at a concentration of 10 µg/mL). At a concentration of 25 µg/mL, however, there was a 9% increase in viability after 48 h (p < 0.001) and an 18% increase after 72 h (p = 0.005). ANOVA statistical tests were performed with Tukey’s correction, and analysis of significant differences between categories with 95% confidence intervals after 48 and 72 h in the XTT study is presented in Supplementary Table S1A and B.
The method of assessing the cytotoxicity of compounds and peptides based on the colorimetric LDH test measuring the activity of LDH released into the environment, that is, the culture medium, after cell membrane damage, did not show cytotoxicity in the experiment with peptides. In the case of the UG27 peptide, a slight protective effect was observed, statistically significant at a concentration of 50 µg/mL 10% (p < 0.00004), 100 µg/mL 5% (p = 0.007), and 150 µg/mL 12% (p = 0.001). In cells treated with Triton X-100 (positive control), a statistically significant (p = 0.001) increase in dehydrogenase activity was observed. For peptide cytotoxicity assays on ADSC cells, statistical tests are presented in Supplementary Table S1C.
Migration test and synthesis of collagen
The ability of the cells to migrate under the influence of the tested peptides was checked by the scratch test (Fig. 2A). Collagen synthesis was assessed using Direct Red staining (Fig. 2B).
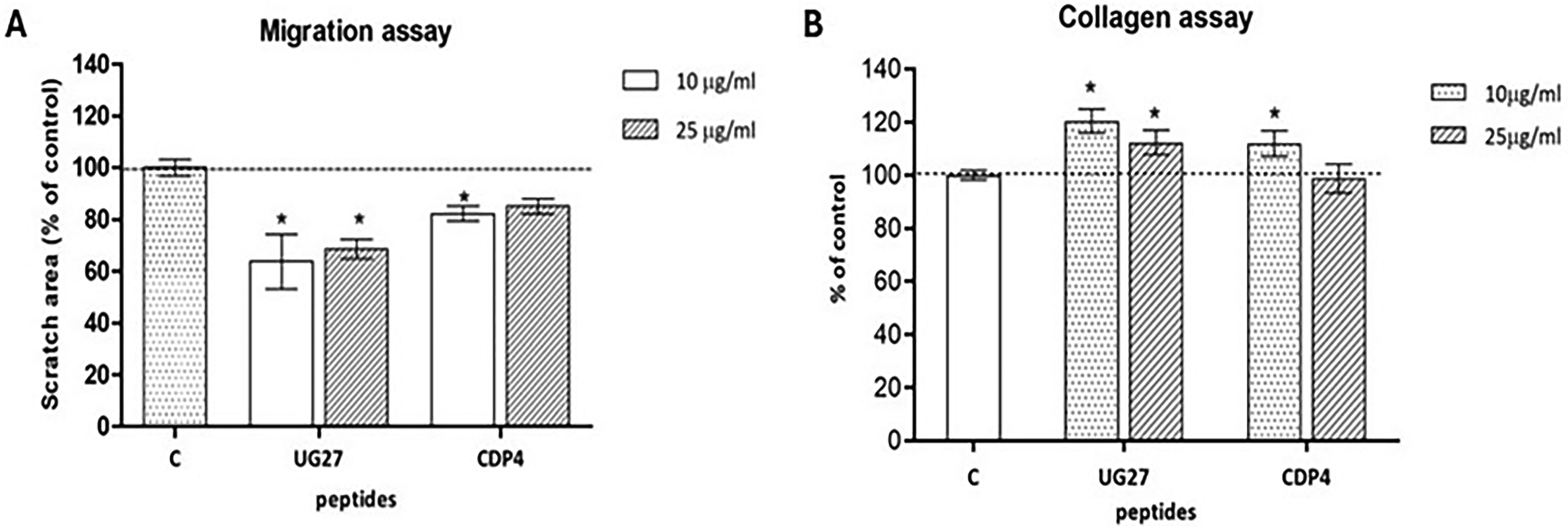
Analysis of ADSC migration and collagen synthesis in the presence of the tested peptides.
The UG27 peptide affected ADSC migration at concentrations of 10 and 25 µg/mL, by about 32–38% compared to the control, the result reached statistical significance (corresponding to concentrations of p < 0.001 and p = 0.012). For the CDP4 peptide, the migration effect was weaker at 18% for a 10 µg/mL concentration, achieving statistical significance (p = 0.042). At a 25 µg/mL concentration of CDP4, cells migrated more slowly, with no statistically significant differences (Supplementary Fig. S4, Supplementary Table S1F).
The peptide UG27 at a concentration of 10 µg/mL had a significant effect on collagen synthesis in ADSC, with the result showing a 20% increase in the synthesis of this protein (p < 0.001). Furthermore, after the application of UG27 at a dose of 25 µg/mL (p = 0.012) and CDP4 at a dose of 10 µg/mL (p = 0.042), statistically significant differences were observed in relation to the control group (p < 0.05), reaching up to 15% (Supplementary Fig. S5, Supplementary Table S1E).
Differentiation of ADSC
UG27 at a concentration of 25 µg/mL increased calcium salt concentration (statistically significant difference compared to the control group, p = 0.049) by as much as 24% compared to the control group without the peptide. This effect was not observed at a dose of 10 µg/mL UG27 and CDP4 peptide at both concentrations (Fig. 3, Supplementary Table S1D). UG 27 at a lower concentration corresponded to the control value without peptide, while Osteo C and CDP4 at concentrations of 10 and 25 µg/mL corresponded to below the control value, 90% and 92%, respectively.
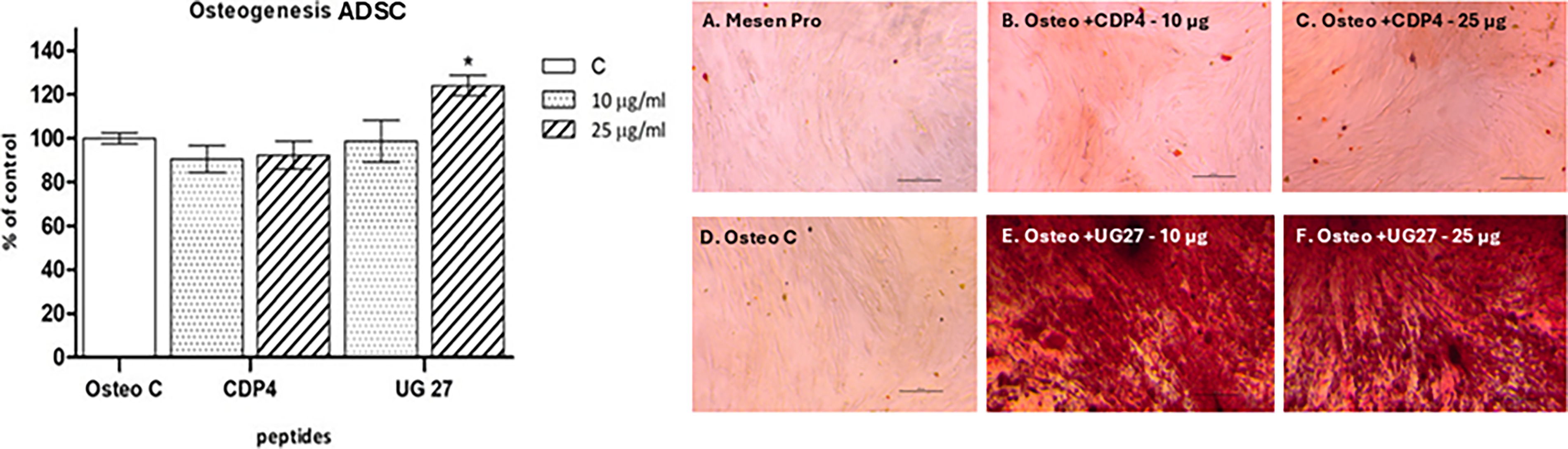
Effects of peptides on osteogenic differentiation of ADSC. Effect of peptides: CDP4 and UG27 at 10 and 25 µg/mL on the differentiation of ADSC toward osteocytes. Quantitative measurement of calcium salts produced by ADSC in MesenPro nondifferentiating medium and OsteoStemPro medium. The graph shows the mean ± standard error of measurement of four independent experiments (in four replicates, n = 16). *Statistically significant differences compared to Osteo C, Mann–Whitney U test, p < 0,05. Alizarin red staining of calcium deposits after differentiation of ADSC toward osteogenesis, after stimulation with CDP peptide derivatives.
Analysis by transmission electron microscopy
Photographs of microscopic preparations made using a transmission electron microscope show the structure of osteoblasts differentiated in a 14-day culture without peptide (Fig. 4A, B) and in the same medium treated with UG27 peptide at a concentration of 25 µg/mL (Fig. 4C–G). The ultrastructure of ADSCs after UG27 stimulation indicates the presence of internal vesicles, likely autophagosomes with amorphous calcium phosphate (ACP). Figure 4C shows long connecting mitochondria, while Figure 4D shows extracellular vesicles just below the cell membrane surface.
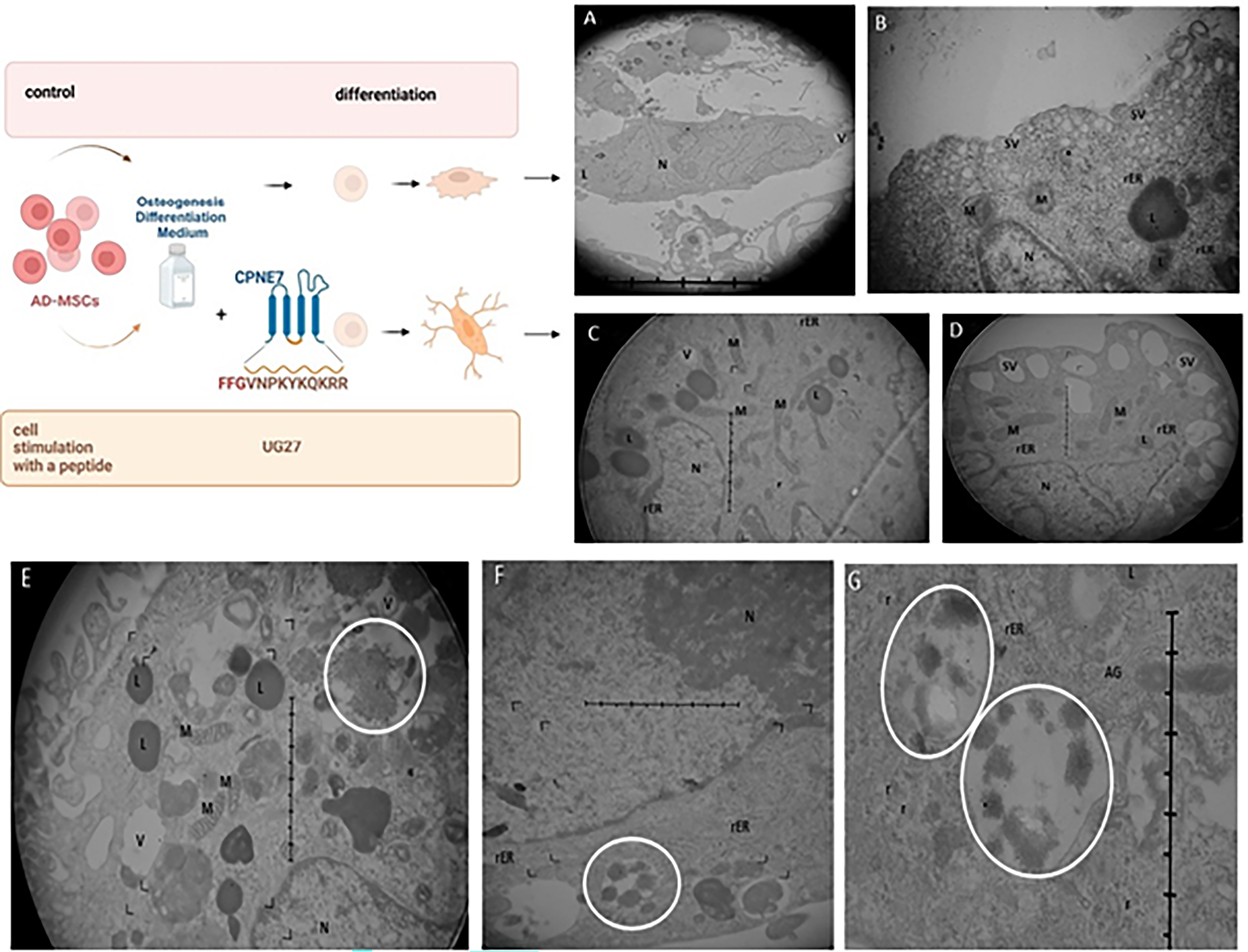
Ultrastructure of ADSC after 14 days of culture in osteogenic medium
Immunogenicity assay and basophil activation assay
The early activation antigens CD25/CD69 and late activation antigens CD71/HLA-DR did not show elevated levels of activated human Th and Tc lymphocytes against Cpne7 protein derivatives at low peptide concentrations (10 μg/mL), statistical significance was observed for the positive control C+ (p = 0.030).
The BAT was performed using the Allergenicity Kit, Cellular Analysis of Allergy. For each patient sample, negative (C−) and two positive (C+), anti-IGE, and fMLP controls were prepared (Fig. 5). In total, 0.1 × 106 cells were evaluated; the number of activated basophils above 5% indicated cell activation under the influence of the peptide. The BAT showed increased values of cell activation for one patient against both peptides at two concentrations up to 10% for UG27, for CDP4 at a concentration of 25 µg/mL 17.4%. Other patients did not reveal the effect of basophil degranulation, and the values fluctuated around the threshold value. The results did not show statistical significance.
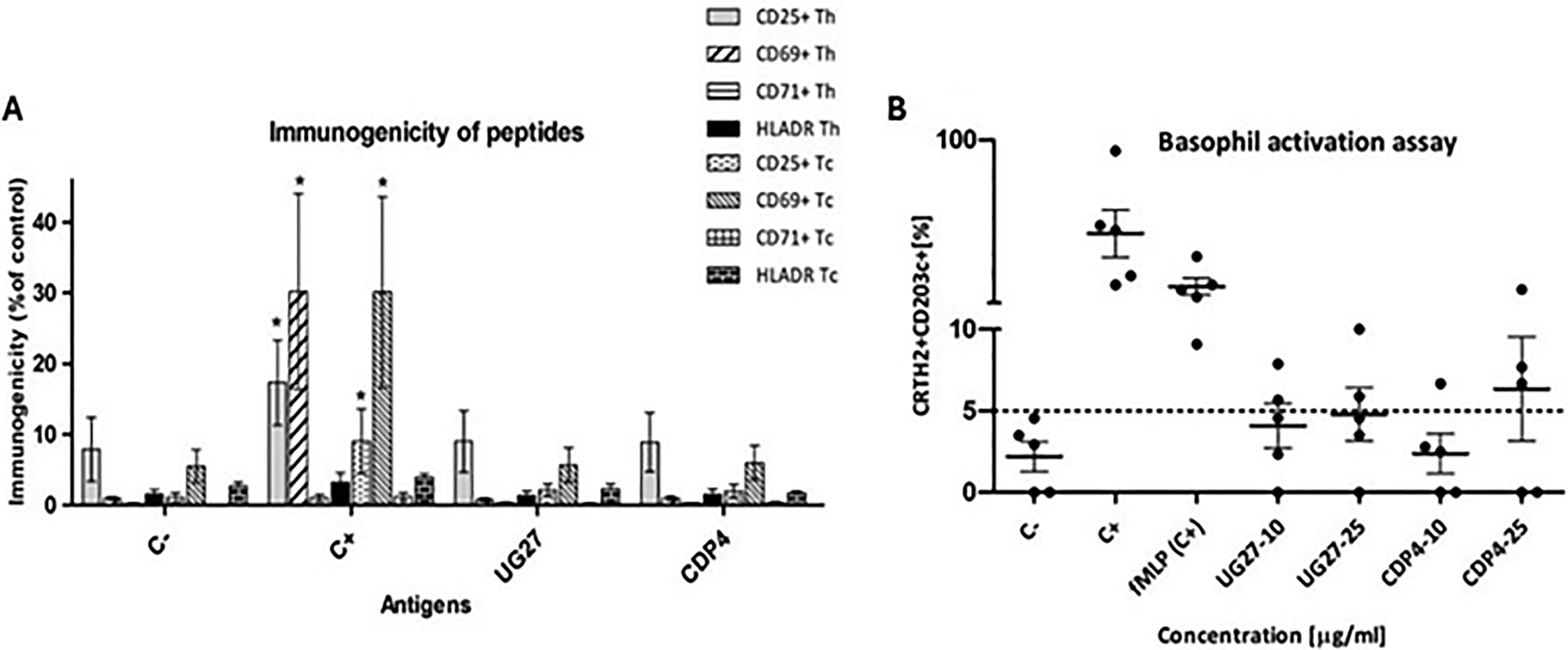
Immunological safety and allergenic potential against the tested peptides.
Analysis of intracellular nucleotide concentration after treatment with the UG27 peptide and in the control
Adenosine triphosphate (ATP) is the main energy carrier, and measurements of intracellular adenosine nucleotide concentrations provide information about the metabolic state of the cell. ATP is used in endoenergetic processes in every cellular function, including maintaining calcium ion homeostasis on both sides of cell membranes and synthesizing compounds necessary for cell differentiation and growth, such as fatty acids, glycogen, proteins, cholesterol, and nucleic acids. 21 The breakdown of the high-energy bond between phosphate groups (hydrolysis) produces ADP (adenosine diphosphate), from which ATP can be resynthesized.
Our results showed that ATP concentration and total adenine nucleotide levels are reduced in cells treated with osteoblast differentiation medium and UG27 peptide compared to osteoblast medium alone, with statistically significant differences relative to the control (p = 0.019) (Fig. 6A, B). At the same time, the ATP/ADP and ATP/AMP (adenosine triphosphate/adenosine monophosphate) ratios were maintained (Fig. 6C, D). 21 As a result of breaking the high-energy bond between the phosphate groups (hydrolysis), ADP is formed, from which ATP can again be synthesized.
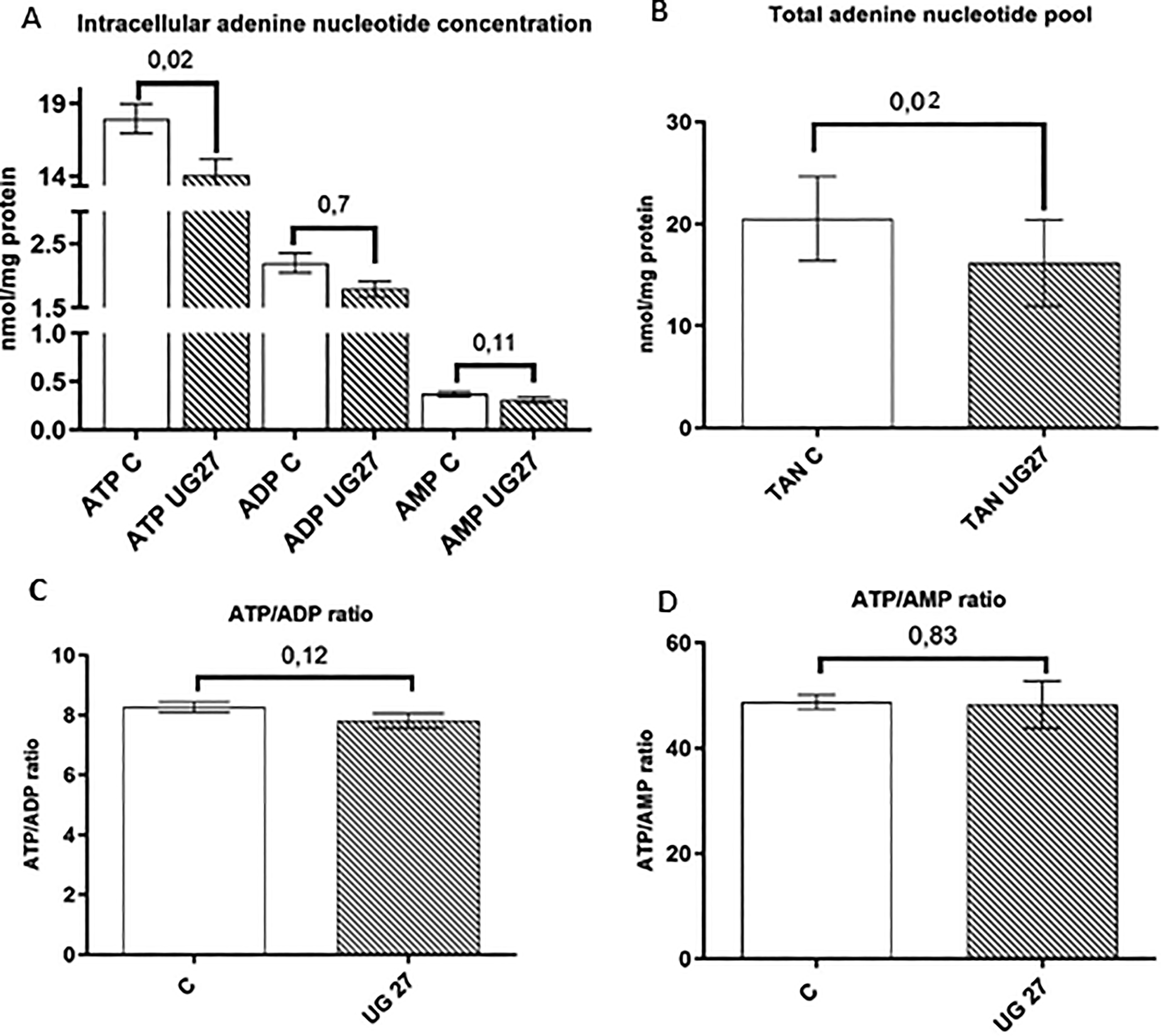
Effect of peptides on the metabolomic potential of nucleotides.
Then we analyzed the effects of UG27 peptide in the presence of the osteoblast differentiation medium on the intracellular concentration of oxidized nicotinamide adenine dinucleotide (NAD+) and its reduced form (NADH). NAD+ is a key substrate for metabolic enzymes, including (1) sirtuins, which are a family of deacetylases that play an essential role in the regulation of energy metabolism and mitochondrial function, 22 (2) poly(ADP-ribose) polymerase, 23 and (3) cyclic ADP ribose synthases, such as CD38 and CD157. 24 The NAD+/NADH redox cycle is integral to energy production through glycolysis, the tricarboxylic acid cycle, and mitochondrial oxidative phosphorylation. Our study revealed a tendency toward a lower concentration in cells after stimulation with the UG27 peptide without affecting NADH, NAD+/NADH, and ATP/NAD+ (Fig. 7A–C).
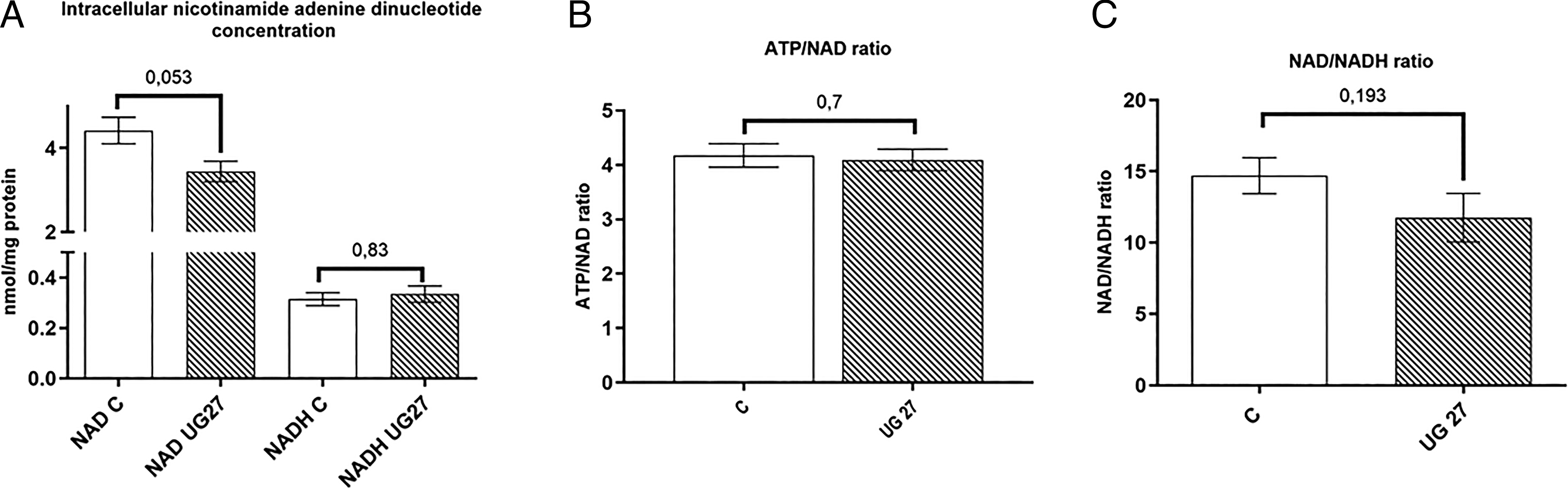
Effect of peptides on the metabolomic potential of nucleotides.
Discussion
The healing process of bone fractures and bone loss is crucial for restoring normal functioning and mobility in humans. The goal of orthopedic and dental therapy has become the delivery of mediators and mesenchymal cells that regulate osteocyte differentiation in biocompatible biomaterials to damaged tissues.
Previous studies have demonstrated the osteogenic potential of the Cpne7 protein, similar to that of the most potent bone morphogenetic factor BMP-2. 25 In dental studies on human dentin cells, Cpne7 increased dentin sialophosphoprotein and osteocalcin, leading to dentin mineralization within 14 days.15,25 This protein is valuable in bone tissue engineering, and its domain 340–350—CDP4 (Val-Asn-Pro-Lys-Tyr-Lys-Gln-Lys-Arg-Arg)—may be critical for osteogenic cell differentiation in vitro and in vivo.25,26 Therefore, in our study, we focus on the UG27 peptide, a derivative of CDP4, designed so that the FFG amino acid sequence, to which PEN sequences sensitive to the action of metalloproteinase VII can be attached, can release the peptide into the environment.
Based on our team’s research on platelet-derived growth factor (PDGF), 27 angiopoietin growth factor, 28 UG28, 29 and other compounds, 30 two concentrations of the peptide (10 and 25 µg/mL) were selected, with the best ratios in terms of viability (Fig. 1A, B), migration test (Fig. 2A), and collagen synthesis (Fig. 2B), as the right concentration and their effective delivery improve bone healing. As Liu showed, BMP-2 must be delivered at a dose 20 times higher than BMP-2/7 through a collagen–HA scaffold to achieve similar results.31,32 We also confirmed, similarly to Lee D, 25 the lack of cytotoxicity of peptides and even demonstrated a slight pro-protective effect (Fig. 1C). Peptides typically activate internal signaling pathways, which initiate a cascade of intracellular reactions leading to cytokine release. Therefore, we evaluated the immunological safety of the UG27 and CDP4 peptides, which did not show activation of Th and Tc lymphocytes (Fig. 5A). Minimal basophil activation was observed in vitro in one patient (Fig. 5B). However, it should be mentioned that synthetic peptides can induce basophil activation in allergic individuals. 33 The absence of an inflammatory response was also confirmed in vivo in mice by Lee D in studies with CDP4 on collagen.
Lee YS et al. showed in in vivo studies on the teeth of beagle dogs that Cpne7-DP (344–353 [KYKQKRRSYK] of the Cpne7 protein) penetrates odontoblasts, increases sialoprotein synthesis, and promotes periapical tissue regeneration.26,31 As suggested by Lee D, CDP4 penetrates the cell through electrostatic interactions between positively charged Lys and Arg residues and negatively charged phospholipids of the cell membrane. 25 We are inclined to believe that similar mechanisms of osteogenic activation occur in our studies on ADSC, which is also confirmed by Byun on mesenchymal stem cells (MSCs) after the addition of a peptide (transcription cofactor), which increases RUNX2 transcriptional activity in the early stages of osteogenesis. 34
Calcium is a signal pathway transmitter that promotes osteogenesis, MSC differentiation into osteoblasts, and ECM. 35 The calcification process begins with the formation of ACP due to the aggregation of Ca2+ and inorganic phosphate (Pi), which transform into more stable crystalline phases within the cell. 36 Alizarin red staining of differentiated ADSC after UG27 stimulation (Fig. 3A, B) showed higher calcium salt values in our tests than in the same tests conducted by Lee. 25 This may indicate the activation of Smad-dependent signaling pathways, which are key to TGF-β and BMP.37,38 Gug noted that the Cpne7-DP molecule penetrates dentinal tubules and reactivates autophagy in inflammatory odontoblasts, leading to dentinal matrix secretion and physiological remineralization. 39 Pei D 2018 identified mitophagy as a mechanism for ACP delivery in osteogenic MSC lines during ECM biomineralization. 40 Accumulated ACP triggers mitophagy, initiating ACP transport between organelles. 36
Transmission electron microscopy (TEM) images of ADSC treated with UG27 showed closed vesicle structures, likely lysosomal granules (Fig. 4E–G), as in the experiments by Pei et al.36,40 After the vesicles are released, the crystals pierce the vesicle membrane, and the storage of ACP and HA crystals in the ECM is induced due to the presence of free ion complexes. 41 ACP granules are transported to autolysosomes and released outside through exocytosis, ensuring mineralization of the ECM and phase transformation of ACP into mineral structures containing apatite. 40 Also noteworthy is the lower number of mitochondria, which may confirm Pei’s observations. 40 Wan indicates that mitophagy aims to regulate mitochondrial dynamics by removing dysfunctional mitochondria from the mitochondrial network in order to maintain mitochondrial network homeostasis. 42 The activation of mitochondrial dynamics through mitochondriogenesis, fusion (Fig. 4C), and mitophagy is responsible for the osteogenic differentiation of MSCs seeded on collagen membranes. The fused mitochondria form a mutual mitochondrial network in response to high-energy demand, 43 which is essential for changing the metabolic needs of cells and adjusting the number of mitochondria to specific stages of cell development, such as cell differentiation.43,44
The involvement of lysosomes in ACP transport in the cytosol appears to be specific to the delivery of compounds for ECM mineralization. 45 Such observations were also confirmed by Boonrungsiman’s team, who demonstrated in their experiments that calcium phosphate was present in both the mitochondrial granules of osteoblasts and in membrane vesicles (Fig. 8), which transported this material to the ECM. 46 Collagen synthesis is of great importance for the formation and deposition of mineralized nodules in the spaces between collagen fibers in the ECM. We demonstrated enhanced collagen synthesis by stimulating ADSC with UG27 compared to the original CDP4, resulting in better HA deposition and ECM mineralization.
Stem cell differentiation is accompanied by changes in ATP production, shifting from glycolysis to oxidative phosphorylation to meet the increased energy demands of collagen synthesis, substance transport, and osteogenic gene activation.47–50 This metabolic change is crucial for supporting the osteogenic differentiation of stem cells. In this study, ADSC treated with the UG27 peptide in osteoblast differentiation medium showed a 20% lower ATP concentration and total adenine nucleotide pool with a slight decrease in intracellular NAD+ (Fig. 7C). This suggests that the UG27 peptide affects the energy metabolism of cells during differentiation. 51 This slight decrease in nucleotide levels may indicate faster osteoblast cell differentiation and thus greater energy consumption, as confirmed by differentiation analysis (Fig. 3A) and alizarin red staining (Fig. 3B). On the contrary, ATP depletion after stimulation with the UG27 peptide during osteogenesis 52 may be the result of changes in mitochondrial dynamics, including fusion and mitophagy processes, which are considered key in biomineralization. 42 However, parameters assessing cell bioenergetics, including unchanged ATP/ADP ratios, ATP/AMP, and NAD+/NADH, showed metabolic compensation for these changes in ATP. Nucleotide ratios are indicators of the energy status of cells and are tightly regulated during differentiation processes. 53 This suggests that cells can adapt their bioenergetics to maintain homeostasis during differentiation.
Conclusion
Our study showed that CDP4-based UG27 stimulates ADSC to proliferate and differentiate into osteocytes. It improves collagen synthesis in ADSC and affects their migration. Application of the peptide leads to collagen biomineralization in the ECM, probably through mitophagy, followed by autophagy and apatite formation. These findings highlight the potential of UG27 in promoting bone regeneration and its safe use in a therapeutic context. This compound is a promising topical or direct agent in orthopedics, maxillofacial surgery, and veterinary medicine.
Future studies will have to focus on the molecular mechanisms of mitophagy by UG27; the expression of OSX, OPN, ALP, or COL1 genes in osteogenesis; and the release rate of the compound from the biomaterial.
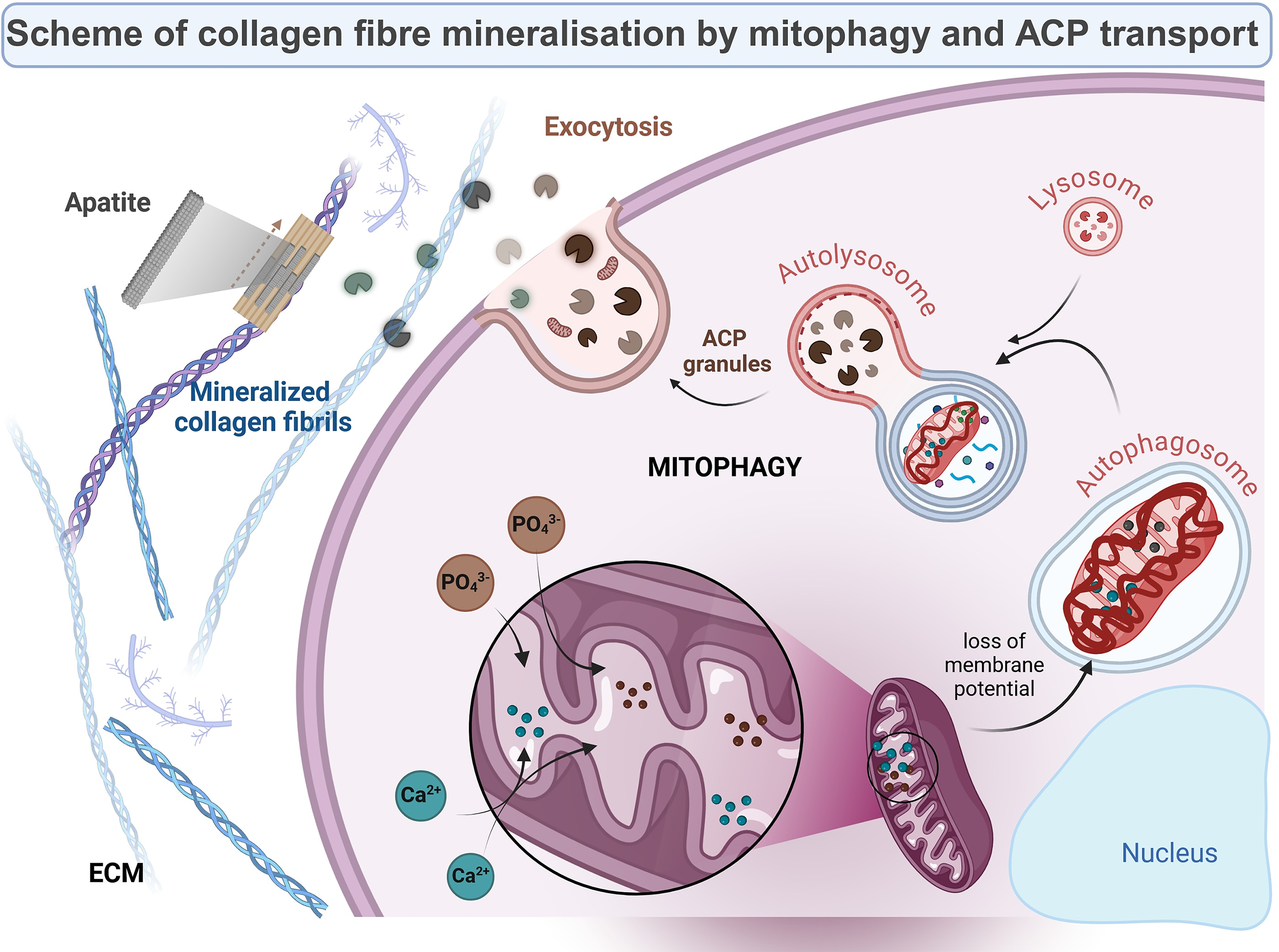
Proposed mechanism of action of the UG27 peptide on ADSC during osteogenesis.
Authors’ Contributions
A.T.: Data curation, methodology, visualization, conceptualization, writing—original draft, and writing—review and editing. A.S.: Methodology and visualization. K.N. and K.C.: Methodology. B.K.Z.: Data curation, methodology, and writing–review and editing. K.K. and J.Z.: Provided material for testing. K.G.-C.: Formal analysis and writing—review and editing. P.M.S.: Conceptualization and funding acquisition. S.R.-M.: Conceptualization, funding acquisition, supervision, and writing—review and editing. M.P.: Funding acquisition, conceptualization, supervision, and writing–review and editing.
Footnotes
Acknowledgments
The authors also like to express their gratitude to A. Olewniak-Adamowska, T. Wolszczak, and A. Guzman for their invaluable assistance in preparing the slides for TEM of the Department of Histology headed by Prof. M. Żmijewski of the Medical University of Gdansk, for granting access to the equipment needed.
Funding Information
This work was supported by the National Center for Research and Development, Poland (Grant TECHMATSTRATEG2/410747/11/2019, acronym BIONANOVA) and Medical University of Gdańsk (Excellence Initiative-Research University program, IDUB, No. 71-01422 granted to M.P.) and statutory funds of the Medical University of Gdańsk (grant number 01-10025).
Data Availability
Data will be made available upon request.
Disclosure Statement
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this article.
Supplemental Material
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.