
Abstract
Over the past three decades, cell sheet technology has evolved from its original scaffold-free approach developed for epithelial cells into a versatile platform applicable to mesenchymal stromal cells (MSC). Despite numerous experimental achievements and convincing preclinical outcomes, MSC-based cell sheets have yet to reach the stage of a clinically reproducible product. This gap reflects not so much the technological limitations of the method as an incomplete understanding of its biological nature. MSC-based cell sheets represent more than a vehicle for cell delivery; they are self-organizing systems governed by intrinsic biophysical and morphogenetic principles. Their structural maturation involves (1) cellular condensation, (2) extracellular matrix deposition, and (3) contractile remodeling—processes that mirror the early phases of granulation tissue formation. Viewing the cell sheet as an in vitro model of connective tissue regeneration opens new research avenues extending beyond its therapeutic applications. This review summarizes the key milestones in the development of MSC-derived cell sheet technology, identifies major challenges and conceptual inconsistencies, and discusses the potential of studying these constructs as autonomous biological systems. The integration of mechanobiology, spatial omics technologies, and tissue engineering approaches may help reconceptualize MSC-based cell sheets both as tools for translational therapy and as a fundamental model for studying self-organizing regenerative processes.
Impact Statement
This review briefly describes predisposing factors that support a redefining view on mesenchymal stromal cell sheets as self-organizing systems rather than routine tissue-engineered delivery vehicles. By integrating insights from mechanobiology, spatial omics, and tissue engineering, it highlights the potential of cell sheets as in vitro models of connective tissue formation, regeneration, and as an experimental platform for investigation of fundamental principles of self-organization in biological systems.
Keywords
Introduction
Cell sheet (CS) engineering has emerged in the early 1990s in T. Okano’s works describing spontaneously assembling cell-bearing constructs devoid of scaffolds.1,2 Methodological milestone that enabled CS detachment, preserving intercellular junctions and extracellular matrix (ECM), was introduction of thermoresponsive polymer coatings for cell culture. This sharply distinguished new technique from conventional treatment of cells by proteolytic enzymes to obtain suspension. Initially, CS were developed for epithelial 3 or corneal endothelium cells 4 for treatment of mucosal or ocular lesions5–7 and CS-based product prototypes have rapidly demonstrated advantages. The latter included high cell density, integration within host tissue, lack of scaffolds of chemical origin, and tunable biodegradability. 8 In the 2000s, CS technology has been adapted for a new cell type—namely mesenchymal stromal cells (MSC) expanding their application to regeneration or replacement of skeletal tissues such as bone or cartilage. Not being limited to mentioned tissue, MSC sheets have also been tested for healing of myocardium, skin, and periodontium. 9
By that time, MSC had already been in the spotlight, and for a couple of decades, they were considered a universal cell therapy platform due to a combination of high efficacy and low immunogenicity that was deemed the basis of clinical safety.10,11 MSC at this point have reached initial clinical and commercial success in applications ranging from tissue engineering to cell-based products (e.g., for treatment of graft-versus-host disease in organ recipients12,13).
Addressing up-to-date reviews on MSC products one may be surprised that from several dozens of prospective cell therapies only a small portion has advanced to the market with majority staggering or terminated in clinical trials.14–16 Indeed, most of these therapies relied on MSC delivery in suspension associated with major limitations: rapid cell death, dissemination from injection site(s), and minimal engraftment.17,18
In this context, CS possessed an exclusive advantage—namely, delivery of a significant amount of organized viable MSC with intact intercellular junctions and ECM. Furthermore, local application provided strong adherence to damaged tissue and was expected to enhance the efficacy of therapies using MSC as the main effector, similarly to active substance in pharmacology. Indeed, once drug’s active substance fails to act (like in case of injected MSC death due to anoikis) one may hardly expect a significant deal of clinical relief once delivery hinders activity.
Despite three decades of extensive trials, MSC-based CS are yet to reach market authorization or conditional approval in any leading nation’s regulatory framework (US Food and Drug Administration, European Medical Agency, UK Medicines and Healthcare products Regulatory Agency, or Japanese Pharmaceuticals and Medical Devices Agency). By 2020 a total of 45 clinical trials using CS of different origins were active or completed accounting 427 patients who have been fully or partially treated in these trials, and still the first MSC sheet product is yet to be marketed. 19 Epithelial—or myoblast-based CS products have received authorization as cellular- or tissue-based medical devices,6,20 and their use remains confined to Japan, where technology was developed and implemented 20 reflecting the complexity of cell technologies translation.20–22
The reasons behind discrepancy of convincing preclinical data and lack of clinical implementation are beyond technical issues and include conceptually overlooked caveats. The present review summarizes the current research effort in the area of MSC-derived CS and discusses the potential to reinterpret their significance as a model of tissue self-organization and therapeutical mode of action, which may suggest new routes in research and advance of CS bioengineering.
Evolution of the Technology and Directions of Application
MSC sheets emerged as a natural extension of scaffold-free tissue engineering principles. Unlike epithelial sheets characterized by tight intercellular junctions, in MSC sheets cells are bound by ECM components (fibronectin, type I and III collagens, laminins), providing cellular adhesion and mechanical integrity. 9 Potential designations for MSC sheets span over a range of tissues—from dense skeletal and muscular structures to parenchymal organs.23,24 With certain degree of relativity, we divide our path to understanding MSC sheets as an object into the following stages based on stipulated CS mode of action.
MSC sheets as delivery tools and tissue-replacing constructs
Implementation of CS allowed delivery of large numbers (up to 107) of viable MSC held together by biocompatible ECM accumulated in vitro. 9 Many groups have considered MSC sheets as a feasible self-organizing construct and put a load of effort using them as “plus-constructs” that filled or replaced damaged tissue. Overall, at this period the general paradigm for MSC therapy was focused on their putative integration to the organ followed by differentiation and even reprogramming25–27 to noncanonical types (beyond bone, cartilage, and adipose tissue). 28 The majority of researchers adopted similar view in MSC engineering, putting trust in CS integration. However, certain authors indicated importance of soluble factors produced by MSC25,29,30 which later became the next basis for understanding their regenerative mode of action.
At this period, the field was driven toward application of MSC sheets for replacement of bone, cartilage, heart, and hollow organs’ mucosal or mural cells. For these designations works published between 2000 and 2015 often pinpointed their role as a tissue-replacing element.
In most reports, transplantation of MSC sheets into bone defects has been shown to accelerate osteogenesis and fracture consolidation by producing and preserving ECM or enhancing expression of osteogenic factors.31–33 Concordantly similar approach has been applied to cartilage repair, where improved regeneration of the subchondral zone and reduced inflammation have been observed after CS transplantation. 34 In some of these works additive scaffold technologies or biomaterials were used (especially for critical bone defects) to facilitate stability, but general involvement of transplanted MSC to the formation of tissue regeneration and eventual incorporation into regrown structure was adopted in MSC sheet engineering for skeletal tissues. 35
Compelling clinical results have been obtained in cases of severe periodontitis36–38 where autologous periodontal ligament MSC sheets restored alveolar bone and cementum without signs of inflammation or rejection. Subsequent multicenter clinical trial confirmed method’s safety and efficacy, with novel tissue persisting >4 years of follow-up, suggesting sheets integration within recipient tissue. 38
Alternative field for MSC sheet was heart repair after myocardial infarction (MI) or other blood supply catastrophes. In these designations MSC sheets were successfully used either as constructs directly attachable to ventricular wall39,40 or as injectable fragments.41,42 Overall, in a porcine model of MI both approaches have shown signs of engraftment and even markers of MSC transdifferentiation to endothelial or smooth muscle cells. 41 This work was of seminal importance as clearly demonstrating superiority of CS (even being fragmented to a certain extent) over injections of dissociated MSC to alleviate heart failure and myocardium loss. To date works in heart disease models point importance of engraftment for therapeutic effect being defined by CS features distinguishing them from suspension.43,44 However, none of these successes has undergone a clinical translation. Only autologous myoblast sheets have been transplanted to severe heart failure patients with limited positive effect20,45 and iPSC-derived cardiomyocyte sheets demonstrated long-term safety in three recipients by 2023. 46
Potential of MSC sheets for mucosa and skin repair has relied on them serving not only as a biomimetic dressing occluding the wound surface but also as modulators of reparative processes via paracrine secretion.47,48 Interestingly, many studies on skin lesion treatment by superficial CS application have encountered short retain of construct and minimal to none engraftment. Stipulated beneficial effects of suggested paracrine mode of action gained more attention after demonstration that delivery of soluble factors derived from MSC mimic and reproduce majority of living MSC effects.49,50
MSC sheets as paracrine regulators of tissue healing: Functionalization and treatment of parenchymal organs
The apparent divergence in translational outcomes between MSC-based sheets and ones comprised of terminally differentiated cells (e.g., epithelial, endothelial, or myoblast sheets) underscores a pivotal misconception regarding their respective modes of action. Sheets of differentiated cells predominantly function as a structural “biological patch”—akin to a “missing puzzle piece” or “spare part” that integrates to restore tissue architecture and its function.20,46 Thus, their efficacy is largely predicated on preexisting mature phenotype and direct structural contribution.
In stark contrast, MSC sheets are not primarily replacement units, but rather orchestrators of healing and its architectural outcome. They deliver a dense, self-organized community of signaling-competent stromal cells whose primary function is to modulate the host microenvironment, guide connective tissue formation, and steer the endogenous repair program—acting more as a “site manager” than a “building block.”29,47,51,52 This profound difference in biological roles explains their impressive preclinical effects (driven by paracrine, immunomodulatory, and matrix-remodeling activities) coupled with inconsistent or unpredictable clinical translation. The failure to achieve market authorization does not stem from the MSC sheet technology per se, but from implying a “patch” paradigm to a different biological entity. To harness the true potential of MSC sheets, the field might shift focus from forcing sheets engraftment to understanding and leveraging their intrinsic capacity as self-organizing, bioactive systems capable of triggering and guiding regeneration.
Understanding how MSC sheets induce regeneration has occurred concordantly with novel insights53,54 to this lineage not being “stem,” but rather “signaling” type as referred by A. Caplan in his work on “MSC” term and their role in tissue homeostasis and repair. 29 At the same time numerous groups demonstrated increase of anti-inflammatory, angiogenic, neurotrophic, development-related factors in MSC sheets 55 compared with conventional confluent culture.
This novel concept was illustrated in models of ischemia: after MI in mice, application of MSC sheets to the epicardium increased ejection fraction and vascular density despite lacking contact with myocardium and not engrafting to ventricular wall.56,57 The deemed mechanism was MSC paracrine activity and diffusion of vascular endothelial growth factor (VEGF), hepatocyte growth factor (HGF), and insulin-like growth factor-1 (IGF-1) that improved microcirculation within the ischemic zone. Same effects have been reported by our group in peripheral artery disease, where subcutaneously transplanted MSC sheets were superior to dispersed cells as stimulators of angiogenesis.58,59
Similar principle of CS regenerative action was applicable to skin repair and regeneration which became the basis for a mechanism we have termed “trigger effect.” We have shown that after surface application to a skin lesion adipose-derived MSC sheets retained for 3–5 days and this short-term paracrine activity was sufficient to promote connective tissue formation, stimulate blood vessel maturation, and prevent fibrosis. 47 Interestingly, short-term impact by MSC sheet resulted in scar-free healing of dermis by day 21 after transplantation. This pointed that factors produced by CS for several days have modified further healing process outcome via mentioned trigger effect. From a mechanistic point of view repertoire of molecules upregulated in CS (vs. monolayer) was enriched by growth factors and cytokines inducing vascular stabilization: HGF, platelet-derived growth factor (PDGF), angiopoietins, but not VEGF 165 known to be a major activator of endothelial sprouting. Basing on this we have speculated that MSC sheets act not only as a biomimetic dressing for the wound, but also as a source of factors directing healing toward epimorphic reconstruction. 47 Collectively, clinical and experimental evidence supports mucosal and cutaneous tissues as optimal targets for MSC sheet application combining the benefits of localized delivery, structural support, and strong paracrine impact.
Significant effort was put into the modification of MSC paracrine activity by its functionalization—enrichment of secreted factors with molecules that drive tissue repair and break pathological circuits. The majority of studies used viral vectors to transduce gene(s) of interest to MSC either prior to assembly or a ready-to-use CS. Our group has shown that adipose tissue MSC sheets transduced by a baculovirus bearing VEGF165 gene markedly enhanced angiogenesis and restored muscle perfusion. 60 Improved recovery after MI by MSC sheets treated by the same vector system was demonstrated by our colleagues with reduced necrosis size and functional characteristics close to a non-injured heart. 61
Once another pleiotropic growth factor was delivered (namely, HGF), functionalized CS induced a wide array of beneficial effects in limb ischemia model: improved vascular and muscular regeneration and reduction of necrosis. 62 Recovery of neural trophic, on par with vascular by importance, has become an attractive task for modified MSC sheets following GDNF gene delivery by baculoviral system 63 or via CRISPR-based activator of GDNF, NGF, and BDNF, demonstrating functionalized MSC sheets superiority 64 in a model of sciatic nerve injury. These works are yet to undergo clinical translation, while they added strength to a view on MSC sheets regulating tissue repair via secretion but not tissue replacement.
Unlike surface tissues and mucosae, delivery of MSC sheets to parenchymal organs remains relatively scarce. This reflected technical and physiological difficulties of integrating sheets into tissue architecture and achieving sustained functional contribution due to high specialization of parenchyma.
Nevertheless, transplantation of MSC sheets to kidney surface in a chronic fibrosis model reduced fibrotic marker expression, restored glomerular architecture, and attenuated nephropathy progression. 65 Similarly, renal application of MSC sheets conferred structural protection and reduced tissue fibrosis after acute damage. 66
Another visceral organ suitable for CS superficial transplantation is liver and MSC sheets in mice with chronic fibrosis induced regression of established fibrotic changes (collagen I/III deposition, hepatic stellate cell activation) and overall improved hepatocyte proliferation and liver function. 67 Despite encouraging findings in animals clinical application of MSC sheets in parenchymal organs is virtually absent. This supports the conclusion that, at present, the technology is primarily applicable to tissues with accessible surfaces—such as mucosal, epithelial, and superficial structures—and requires substantial refinement before being reliably extended to parenchymal regeneration.
MSC sheets reproduce intrinsic programs of stromal cells during development and wound healing—a novel view on subject
CS formation is considered a merely technical step, but recent findings suggest a more complex overview of this process driven by an intrinsic ability of connective tissue cells for self-organization. As MSC reach high confluence, they actively produce and deposit ECM components (fibronectin, collagen type I, laminins) to create a 3D network providing adhesion and mechanical integrity.9,47 Under these conditions, cells engage in novel mechanical and biochemical interactions, redistributing intracellular tension to form a stable, organized structure retaining after detachment.
Interesting feature of this process is stereotypical reproducibility regardless of MSC source—bone marrow, adipose tissue, umbilical cord, or dental pulp. While these tissues differ in terms of density and structural features, resulting CS exhibit similar morphology. They comprise a moderately stratified multilayer construct 20–50 µm thick, containing interwoven collagen and fibronectin fibers with dense and uneven ECM deposition. 2 Thus, despite established tissue-specificity strongly supported by numerous factors68,69 (transcriptional program, developmental origin, molecular patterns—e.g., HOX-identity 70 ) a stereotypic morphogenetic program putatively exists and is autonomously implemented by MSC in vitro, resulting in similar morphology of CS.
CS formation as a reminiscence of stromal organization via condensation in development
A peculiar feature of MSC sheets is spontaneous formation of stable regions with increased cell and ECM density. This pattern in vitro results in structural anisotropy resembling condensed state of mesenchyme during ontogenesis. This phenomenon was described and investigated in detail by our group demonstrating that during sheet maturation MSC undergo self-organization into condensed domains. 51 In our further studies the morphogenetic events within MSC sheets were shown to be closely linked to ECM deposition pattern and dynamics. 71 The structure and orientation of collagen and fibronectin fibers determine the rate of maturation and the mechanical stability of the construct. Spatial heterogeneity emphasizes that a CS is not a static construct but a dynamic system governed by intrinsic mechanoregulatory principles and cytoskeletal tension. Indeed, once mechanosensing in MSC was blocked by ROCK-inhibitor during assembly, previously observed condensation was fully abolished. 51
Generally, CS may be regarded as a self-organizing, tissue-like system reproducible in vitro. This supports existence of an autonomous program that MSC implement once regulatory context of tissue is absent and cultural expansion allows them to self-govern the structure in vitro. Whether we can expand this to in vivo physiological processes remains highly enigmatic, but in our studies the transcriptomic link between ECM, condensation and embryogenesis has been concordant with lineage commitment observed in CS.51,52,71 Once sheet is formed, cells readily undergo differentiation into two of three lineages, namely, bone and cartilage, but not fat.
Indicated bias toward osteo- and chondrogenic differentiation may reflect a deeper biological program recapitulated within CS. In embryogenesis, bone and cartilage formation are tightly linked to preceding mesenchyme condensation—a process of spatial self-organization governed by cell adhesion and mechanotransduction.72,73 Autonomous emergence of condensed domains within MSC sheets in vitro 51 may therefore not only ensure structural integrity but also create a microenvironment that selectively supports skeletogenic developmental pathways. It explains the efficacy of MSC sheets in bone and cartilage regeneration, where their role may expand beyond direct replacement to being an organizational center that introduces morphogenetic signals invoked by condensed mesenchyme. Indeed, during epimorphic bone repair after fracture in adult 74 condensed MSC are not always integrated to growing callus but provide regulatory pivots sometimes compared with ossification points.
This predisposition requires further validation, as it has been primarily documented for sheets derived from adipose tissue-derived MSCs, and rigorous comparative studies between MSC of different tissue origins (e.g., bone marrow, umbilical cord) are virtually absent. Interestingly, we have established there is at least one exception among MSC populations that failed to form cohesive sheets. Namely, endometrial MSC fail to provide typical multilayered anisotropic configuration75,76 which indicates that capacity for self-organization to a condensed structure is a variable and potentially tissue-specific trait. Thus, the restricted differentiation potential is not a definitive limitation but an under-explored feature that underscores the necessity to study the MSC sheet as a dynamic biological system with properties dictated by cells’ origin and acquired, self-organized state.
Here we may raise a fundamental question: what, in fact, is a CS? One should acknowledge that it goes beyond layers of stromal cells held together by ECM but stands closer to a self-organizing structure with its own internal dynamics and regulatory rules. Let’s summarize the point that keeps this concept together: regardless of source MSC in vitro consistently form a similar morphological pattern of a dense, mildly stratified construct containing an organized functional ECM network. Reproducibility of this observation suggests existence of an intrinsic biological program likely reflecting connective tissue during in vivo development.
CS formation is providing environment-dependent clues for early stages of wound healing
The weak part of the postulate above is lack of context—culture conditions and medium composition reproduce the tissue to a minimal extent and obviously provide sufficient but not physiological settings for cell propagation and activity.
Being seeded as suspension to serum-containing medium, MSC are likely to sense this environment as a damage site with shattered intercellular contacts filled by wound fluid. Indeed, plasma proteins, cytokines, and fibrin-related components create a microenvironment reminiscent of early wound healing. Consequently, despite establishment of intercellular contacts in ECM at later stages of CS formation, initial behavior of stromal cells may be similar to their involvement to early healing programs. In this context, morphogenetic events in MSC sheet may reflect formation of granulation tissue—a dense primary cell–matrix complex existing at the start of tissue repair. 77
Granulation tissue (GT) represents a stereotypic stage of regeneration involving active angiogenesis, cell proliferation, and formation of a provisional ECM. Balanced growth and remodeling of GT promotes regenerative outcomes, whereas its distortion or overgrowth leads to fibrosis.78,79 Its molecular and morphological dynamics are well characterized, including the physicochemical properties of matrix, 80 proteoglycan patterns, 81 programmed cell death during transition to scar, 82 and mechanisms underlying hypertrophic scarring in impaired remodeling. 83
Overall, MSC sheets can be considered a feasible and robust in vitro model of the early/organizing GT, enabling controlled investigation of key aspects of tissue repair aside from systemic signals. Numerous parallels between CS and GT structure are reinforced at several levels:
Cellular composition and phenotype: Similar to GT, in vivo MSC sheet is composed predominantly of activated, ECM-synthesizing fibroblast-like cells. Furthermore, during sheet maturation (∼7 to 12 days of culture), a subset of cells acquires myofibroblast phenotype, with increased expression of α-smooth muscle actin (α-SMA) and enhanced contractility—a hallmark feature of organizing GT cells responsible for contraction and remodeling;51,84 Provisional ECM dynamics: The MSC sheet recapitulates the sequence of ECM synthesis and remodeling characteristic of GT. In the early stages of sheet formation, active deposition of fibronectin and type III collagen creates a provisional matrix network analogous to early GT.71,80 As maturation proceeds, a shift to deposition of a more stable type I collagen is observed resembling in vivo process of ECM maturation and compaction; Spatial self-organization (patterning): A hallmark of GT is its heterogeneity and formation of condensed cell–matrix domains. This phenomenon is vividly reproduced in MSC sheets via spontaneous formation of condensed domains interspersed with less dense areas.51,85 This topological feature is not an artifact but is governed by the same mechanisms (ROCK-dependent cellular tension) that drive condensation in development and, presumably, in GT organization; Secretory profile and regulatory function: The MSC sheet in vitro demonstrates a significantly enhanced secretion of a broad spectrum of factors regulating angiogenesis (VEGF, HGF, angiopoietins), inflammation, and fibroblast proliferation compared with monolayer culture.47,55 This paracrine “burst” directly corresponds to GT’s biological function as a local biochemical hub orchestrating healing via control of cell recruitment, neoangiogenesis, and inflammation control; Commitment potential: As discussed above microenvironment of condensed sheet predisposes MSC toward osteo- and chondrogenic differentiation pathways (section “CS formation as a reminiscence of stromal organization via condensation in development”), aligning with GT role as a precursor to regenerating skeletal tissues. Concurrently, the adipogenic pathway is suppressed, which is also consistent with in vivo data where adipogenesis in active bone repair zone is considered pathological.
General overview of this section is provided in Figure 1 with a focus on MSC sheet engineering. Therefore, an MSC sheet can be viewed not only as a vehicle for cell delivery but as a reduced yet biologically relevant model of the early phase of connective tissue repair—an isolated system in which key stages of regeneration (matrixogenesis, condensation, acquisition of a contractile phenotype, secretory activity) can be observed and manipulated independently of vascular and immune influences. This transforms CS technology from a utilitarian transplantation method into an experimental platform to studying the fundamental principles of connective tissue self-organization and regeneration.
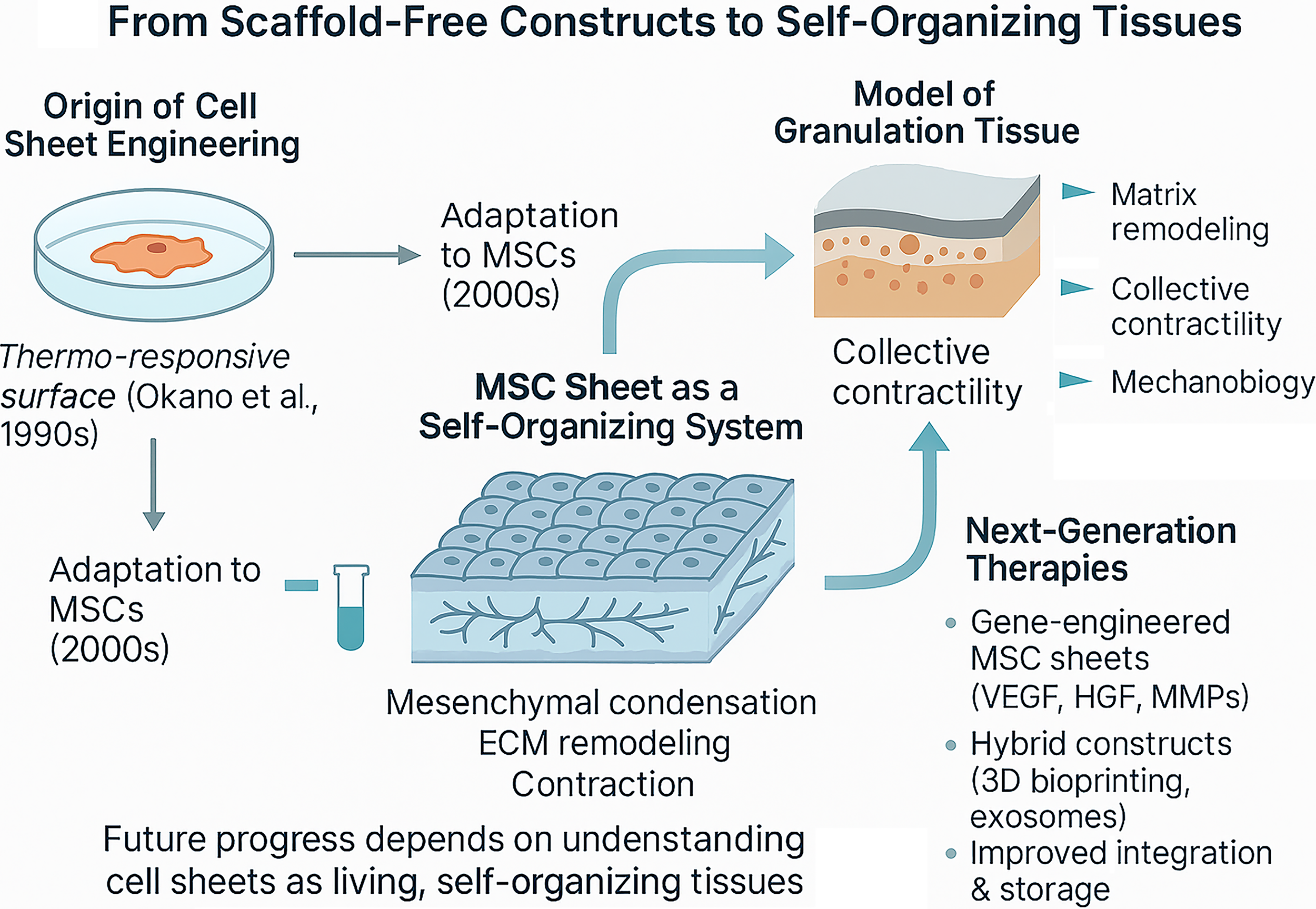
Historical development of MSC sheet engineering and its further progress prospects as of current status of published data. MSC, mesenchymal stromal cells.
Technological limitations and perspectives for practical advances
Analysis of available studies reveals that regardless of target tissue or model system, reported effects of MSC sheets are typically described in stereotypic terms: “enhanced angiogenesis,” “growth factor secretion,” or “improved cell survival.” However, in-depth mechanistic investigation remains scarce, and most reports lack assessment of structural remodeling or spatial organization of the constructs. This indicates that the field still relies largely on phenomenological observations rather than understanding of mechanism.
Although laboratory-scale protocols for generating CS are established, translating it to a standard, manufacturing-grade processes remains a major challenge. The main limitation is lack of knowledge on intrinsic processes of self-organization during sheet maturation with some of those mentioned in above sections. Without understanding the principles governing cellular density, cell-to-ECM ratio, and the stages of functional maturation, it is hard to define consistent technological parameters or quality criteria.24,86 In contrast to cell suspension, which can be evaluated using established potency and validation schemes, CS is a dynamic biological system with variable and difficult-to-quantify functional activity.
Significant variability arises from donor traits, MSC source, seeding density, and culture conditions.86,87 Such heterogeneity complicates data comparison across laboratories and hinders process standardization. Physical fragility of the constructs makes them sensitive to mechanical handling—CS can easily deform or tear during detachment or transfer, necessitating use of supporting membranes and specialized tools.88,89 Further obstacles rise at the stage of sheet fixation to recipient tissues—although intrinsic adhesiveness is preserved due to retained ECM components, attachment to mobile or mechanically active organs (e.g., heart) may result in partial displacement or loss of the CS. Several studies proposed solutions involving fibrin glue, biocompatible adhesives, or hybrid membranes to improve the interface stability. 90
Storage and transportation represent additional bottlenecks, as viability of CS outside an incubator rarely exceeds 24–48 h, and no established protocol for short-term preservation or cryostorage exists.91,92 Collectively, these issues hinder development of a standardized technological workflow essential for transition from experimental protocols to industry.
Taken together, we may define the current state of what might be described as “technological saturation.” Most practical directions have already been explored, and the therapeutic effects are reproducible across various models. Nevertheless, the fundamental understanding of CS formation dynamics and mechanism of action remains limited. Without deeper insights into these processes, further technological progress seems unlikely and existing achievements would remain difficult to scale.
Perspectives for MSC Sheets Application in Fundamental Research
Present state of MSC sheet technology is marked by an evident paradox: despite the abundance of experimental data describing their impact on regeneration, the mechanisms of their formation and therapeutic effects remain insufficiently understood. For further progress, the focus might shift toward the fundamental study of the CS as a self-organizing biological system.
Study of mechanobiology and morphogenesis
One of the most promising directions for future research is investigation of mechanical aspects of connective tissue morphogenesis. Contraction, compaction, and remodeling of the ECM observed during sheet formation indicate the crucial role of mechanical tension, cell adhesion, and cytoskeletal dynamics. Use of direct tension mapping, deformation analysis, and mechanotransduction assays could help to elucidate the forces that maintain the structural stability and self-organization of CS93,94 as a model of connective tissue formation.
Recent experimental studies confirm that self-organization of MSC is accompanied by emergence of ordered topological patterns reflecting the mechanical and geometric constraints of the system. It was demonstrated that the spatial distribution of topological defects within a CS predicts subsequent condensation and chondrogenic differentiation processes. 95 These findings highlight MSC sheets as model systems of self-organization in mesenchyme and its derivatives.
To move beyond describing topological patterns toward understanding their biological significance, researchers may focus on deciphering the specific signal cascades that translate mechanical tension into cellular decisions. Key targets might include:
mechano-induced transcriptional coactivators YAP/TAZ, whose nuclear/cytoplasmic shuttling serves as a direct readout of cellular tension
93
and may predict zones of active remodeling and commitment within the sheet; effectors of the Rho-ROCK pathway, which directly governs actomyosin contractility—the engine of condensation. Their spatial mapping would allow the correlation of force fields within the sheet with observed topological defects; strain-sensitive adhesion molecules, such as phosphorylated forms of focal adhesion kinase (FAK) and paxillin. Their analysis will reveal how cell–matrix attachment points evolve during sheet maturation and stress redistribution.
Beyond these canonical pathways, several less obvious but potentially pivotal mechanisms warrant investigation in the context of sheet self-organization: integrin-mediated mechanosensing via caveolae; 96 regulation of the nuclear scaffold and its role in mechanotransmission; 97 phase separation in cytoskeleton and adhesion complex organization. 98
Understanding heterogeneity and spatial tissue organization
Equally promising is exploration of spatial and functional heterogeneity within CS using modern omics approaches—such as single-cell RNA sequencing and spatial transcriptomics. This may reveal differences between CS regions to trace the transition of cells from initial proliferative state in suspension to a mature or committed state within CS. These techniques lay grounds for understanding how cell and environmental heterogeneity impacts self-organization of connective tissues. 99
The principal goal of spatial omics in this context may be to test specific hypotheses arising from comparison of CS to living tissues (e.g., GT), rather than merely cataloging differences. For instance:
do avascular sheets establish gradients of hypoxia-mimicking factors characteristic of a wound core (e.g., HIF-1α-dependent genes like VEGFA, CA9)? can a transient state analogous to activated GT myofibroblast be identified by coexpression of markers such as ACTA2 (α-SMA), TAGLN (SM22α), and profibrotic genes POSTN or CTHRC1? Is the spatial segregation of chondrogenic (SOX9+) and osteogenic (RUNX2+) cell pools linked to local differences in matrix stiffness or cell packing density?
Provided clues are not confining the researcher’s interest yet; these aspects can be definitely investigated to certain extent using CS as a model, and hardly one can pass the question of tissue-specificity of these traits once they are established. Briefly, tissue of different origins may have contrasting molecular patterns of connective tissue organizations and MSC sheets are a great model to investigate them.
Cellular interactions with the microenvironment and GT
Another direction concerns the interaction between the transplanted CS and host tissue microenvironment. Preclinical studies have shown that MSC sheets stimulate GT formation, promote angiogenesis, and prevent fibrosis, acting as active regulators of the microenvironment rather than mechanical substitutes for damaged tissues. 52 These findings highlight that CS efficacy is determined primarily by their capacity to create conditions for successful healing and regeneration rather than by direct structural integration.
GT is a transitory system formed by coordinated interplay of stromal cells, matrix components, and vasculature. In our studies, MSC sheet transplantation has accelerated GT maturation but also induced formation of a stable connective tissue layer featuring organized mature vessels with vivid mural component, indicating shift from fragile, leaky sprouts to stabilized microvessels. 47
Direct validation of the “sheet as GT model” hypothesis could come from comparative spatiotemporal proteomic/transcriptomic analyses that may help to establish the following:
matrix fingerprint: head-to-head comparison of composition and organization of ECM proteins (primarily fibronectin and type III collagen as hallmarks of early/organizing GT) in a mature sheet in vitro and in forming GT in vivo at a comparable stage (e.g., day 5–7); myofibroblast transformation signature: identification of shared expression patterns for markers of the contractile phenotype (α-SMA, transglutaminase-2) and crucial regulators of stroma-to-myofibroblast transition (TGF-β, CTGF/CCN2) in mature CS and GT specimen; secretory portrait: analysis of overlapping spectra of secreted factors in conditioned medium from CS and wound exudate/fluid focusing on angiocrine (Angiopoietin-1, HGF) and immunomodulatory (PGE2, TSG-6) molecules that determine repair outcomes.
Genetic and molecular engineering of cell sheets
Extra opportunities arise from genetic modification to tune the biological properties of MSC sheets. Expression of angiogenic, anti-inflammatory, or antifibrotic factors may enhance the therapeutic effects and reduce donor-dependent variability. The introduction of regulatory gene cassettes or exosome-mediated signaling systems could enable the creation of sheets with predefined secretory profiles.62,67
To transcend simple factor overexpression, genetic engineering may be a tool for hypothesis-driven deconstruction of self-organization, for example:
testing morphogen gradients: creating MSCs with optogenetically controllable expression of a desired morphogen (e.g., TGF-β1 or BMP2) could allow researchers to artificially establish spatial gradients within the sheet and observe whether they guide condensation patterns and lineage commitment in a predictable manner, directly testing the role of endogenous patterning; dissecting matrix feedback loops: CRISPRi/a-mediated knock-down or constitutive activation of genes encoding specific ECM receptors (e.g., integrin α5 or β1 subunits) or cross-linking enzymes (e.g., lysyl oxidase, LOX) would clarify the causal relationship between matrix deposition, stiffness, and the emergent sheet architecture. engineering synthetic cellular communication: introducing synthetic notch (synNotch) receptors that activate user-defined transcriptional programs upon cell–cell contact could be used to program self-organization rules from the ground up, transforming the sheet into a truly programmable morphogenetic system.
100
Integration with bioengineering and technological approaches
Understanding adhesion and matrix remodeling after transplantation would allow optimizing sheet properties (thickness, porosity, maturation degree, and mechanical stability) prior to implantation. At the same time, development of efficient preservation and transportation remains a vital task. Promising directions include hypothermic and partially cryogenic storage, as well as mechanical detachment techniques that minimize structural damage. 89
Integrating CS technology with other areas of regenerative medicine such as 3D bioprinting, tissue microengineering, and exosome-based therapy opens new perspectives. CS may serve as modular building blocks for multilayered hybrid constructs or as sources of extracellular vesicles with reproducible contents.19,94
The convergence with bioengineering should focus on creating modular, tunable platforms to dissect structure–function relationships.
controlling the initial “mechanical niche”: using micropatterned substrates with defined stiffness gradients to seed MSC sheets would allow researchers to preprogram the initial mechanical context and study its long-term impact on self-organization, testing whether sheet morphology and fate are imprinted by early mechanical cues; designing heterotypic sheet interfaces: 3D bioprinting of MSC sheet fragments alongside endothelial cell spheroids or neural crest cell aggregates could create simplified, spatially controlled models of tissue interfaces (e.g., osteochondral unit, neurovascular niche) to study how MSC sheets orchestrate complex regenerative crosstalk in a more physiological, yet reductionist, setting. harnessing exosomes as programmable messengers: rather than using naive exosomes, engineering MSC sheets to produce exosomes loaded with specific miRNAs or inhibitors (e.g., targeting YAP or ROCK) could transform the sheet from a bulk factor source into a spatiotemporally controlled delivery system for mechanistic probes or therapeutic modulators.
101
Conclusion
Three decades of research in CS technology have demonstrated remarkable technological progress and a broad range of potential applications, yet not in a clinically reproducible product. This situation stems not from limitations of the object itself, but rather from an incomplete understanding of its biological essence.
Extensive expansion of designations for CS use without advance of methodological foundations is unlikely to yield substantial progress. Sticking to an existing paradigm: “place a sheet over a lesion,” no longer expands scientific understanding, and the field must transition toward studies aimed at discovery of intrinsic self-organization principles rather than simply demonstration of therapeutic outcomes. Thus, advance of the field requires an integrative approach that combines cell biology, biophysics, and engineering. Viewing the MSC sheet as a self-organizing biological entity will not only improve its reproducibility but also bring us to understanding the fundamental principles of connective tissue regeneration.
Shifting the focus from empirical application toward studying the CS as a model of connective tissue regeneration opens new perspectives. The sheet recapitulates the key stages of development (e.g., mesenchyme condensation) and early tissue repair (provisional matrix synthesis, cell migration, angiogenesis), remodeling, and contraction. Investigating these processes in a controlled in vitro setting allows MSC sheets to be regarded as an experimental platform to unveil the mechanisms of tissue healing and fibrosis.
Summarizing the three domains of MSC sheets technology is provided in Figure 2 with a focus on novel applications for physiology research and studies of in vivo–related process in development and tissue repair.
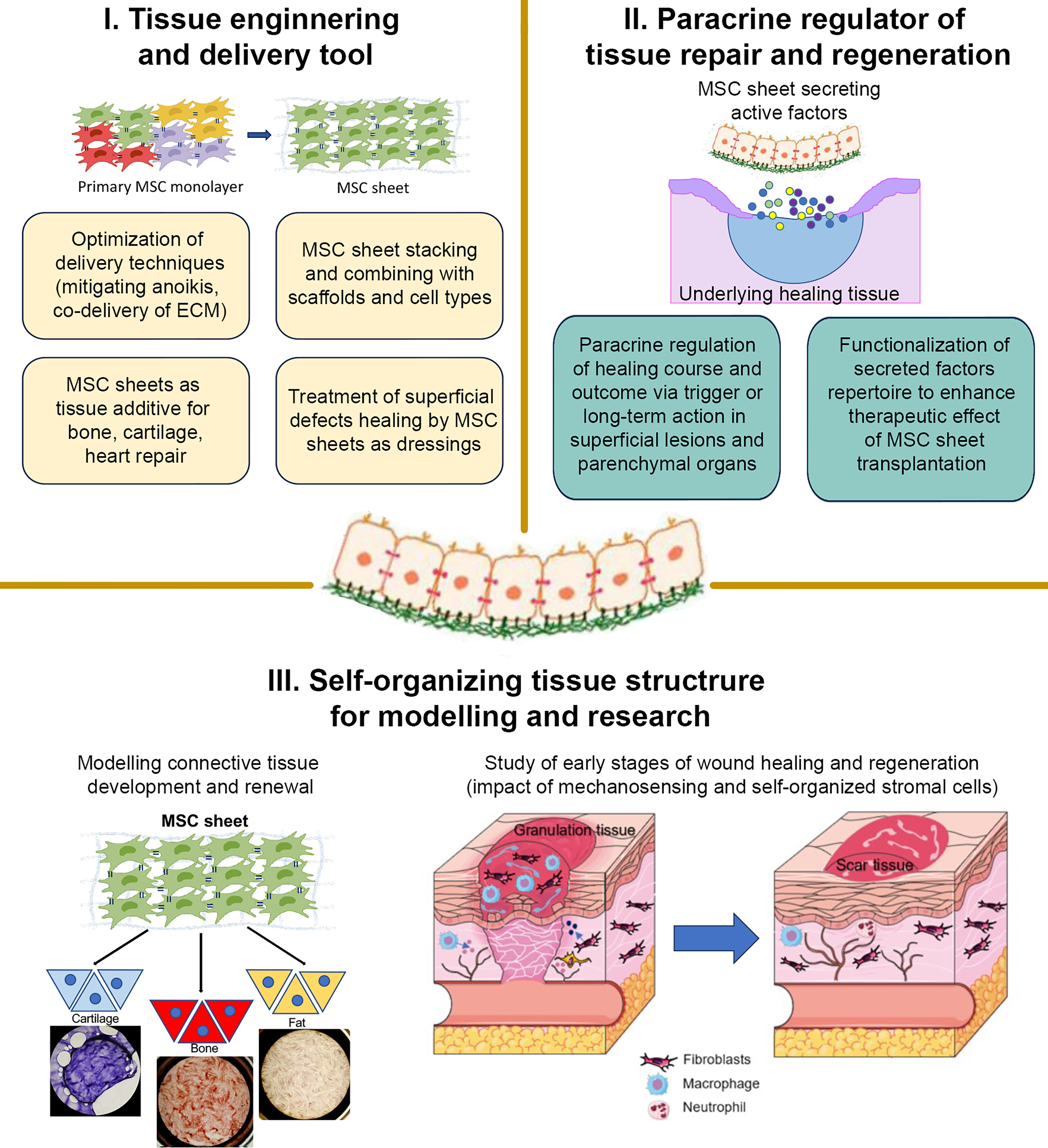
Three domains of MSC sheet engineering application in regenerative biology and medicine. MSC, mesenchymal stromal cells.
Further advance of this technology depends on the integration of fundamental and engineering approaches. Described comprehensive framework may bridge the existing gap between experimental and clinical stages, transforming CS technology from a delivery or research method into a reproducible therapeutic platform.
Authors’ Contributions
N.A.A.: First draft preparation and final text approval. V.S.G.: Published data collection and analysis and draft editing. M.A.V.: Published data collection and analysis and draft editing. R.Y.E.: Image preparation and published data analysis. Y.-C.H.: Final text editing and approval. P.I.M.: Supervision, final proofs, image proofs, and funding acquisition.
Footnotes
Acknowledgments
The authors express their gratitude to the staff of the Laboratory of Medical Bioengineering for valuable discussions and experimental contributions that inspired the conceptual framework of this review. The authors also acknowledge ChatGPT use for language polishing purposes only under author supervision. All intellectual content, analysis, and interpretations are solely those of the authors.
Author Disclosure Statement
No competing financial interests exist.