
Abstract
Noncoding RNAs (ncRNAs) comprise a significant portion of the transcriptome and contribute to various cellular and molecular processes and disease development. Although many functional roles for ncRNAs have been discovered, the regulatory functions of many require further elucidation. Intervertebral disk degeneration (IDD) is a leading cause of lower back pain and represents a major social and financial challenge worldwide. This review summarizes the literature on long ncRNAs and circular RNAs as novel regulators in IDD development. IDD diminishes quality of life, underscoring the importance of research on its pathogenesis and the identification of novel therapeutic targets. The definitive cellular and molecular mechanisms underlying IDD pathogenesis remain unidentified, and no effective treatment is available. Both genetic and environmental or lifestyle factors are associated with IDD development. Therefore, understanding the exact molecular mechanisms of IDD could yield novel therapeutic targets.
Introduction
Noncoding RNAs (ncRNAs) are important regulatory elements encoded by the genome with crucial functions in various cellular processes. ncRNAs are divided into two categories: housekeeping ncRNAs and regulatory ncRNAs. Housekeeping ncRNAs comprise transfer RNAs, ribosomal RNAs, small nucleolar RNAs, small nuclear RNAs, guide RNAs, and telomerase RNAs. Regulatory ncRNAs are classified into two principal groups: (1) small ncRNAs with a length less than 200 nucleotides, consisting of microRNAs (miRNAs), small interfering RNAs, and PIWI-interacting RNAs; and (2) long noncoding RNAs (lncRNAs) with a length exceeding 200 nucleotides (Maxmen, 2013), including circular RNAs (circRNAs) (Bagheri Moghaddam et al., 2022).
The intervertebral disk confers mobility and flexibility to the spine. It consists of fibrocartilaginous tissue and connects the vertebrae. Morphologically, the intervertebral disk comprises the nucleus pulposus (NP), the annulus fibrosus, and the cartilage endplate (Wise et al., 2020). Maintaining homeostasis is crucial for the NP, as it secretes a complex extracellular matrix (ECM) that includes type II collagen and proteoglycans (Wang et al., 2013). NP cells undergo phenotypic transition due to cellular and biochemical changes during intervertebral disk degeneration (IDD), leading to reduced proteoglycan synthesis and events that weaken the structural integrity of the disks.
The precise cellular and molecular mechanisms underlying IDD pathogenesis remain largely unknown, which explains why no effective treatment has been introduced. Both genetics (e.g., polymorphisms in genes encoding collagen types I, IX, and XI and aggrecan) and environmental factors or lifestyle (e.g., lack of exercise and exposure to mechanical pressure) are involved in IDD development (Battié et al., 2004; Nakamichi et al., 2016). Understanding the molecular mechanisms of IDD could lead to the identification of new therapeutic targets. This review summarizes the roles of lncRNAs and circRNAs as novel regulators in the development of IDD.
lncRNAs
lncRNAs lack open reading frames and contain RNA-processing signals such as poly(A) tails and 5′ caps. They are often transcribed by RNA polymerase II (Guttman et al., 2009; Mercer and Mattick, 2013). lncRNAs can be categorized into five subgroups based on their location and properties: sense, antisense, bidirectional, intergenic, and intronic. Sense lncRNAs overlap with exons of another transcript on the same strand, whereas antisense lncRNAs overlap with exons on the opposite strand. Bidirectional lncRNAs are transcribed in close genomic proximity to neighboring coding transcripts on the opposite strand. Intergenic lncRNAs are located between two genes, and intronic lncRNAs are transcribed within an intron of a second transcript (Ponting et al., 2009).
lncRNAs are involved in a wide range of biological processes, including the regulation of gene expression due to their structural diversity (Mirzadeh Azad et al., 2019; Taheri Bajgan et al., 2022). They participate in the regulation of protein-coding gene translation, gene transcription, epigenomic regulation, RNA turnover, chromatin organization, tumor development, stem cell differentiation, immune responses, inflammation-related processes, and genome defense (Briggs et al., 2015; Ferraiuolo et al., 2010).
Mounting evidence suggests that lncRNAs can be used as biomarkers and therapeutic targets in both normal and abnormal processes, such as cartilage development, degeneration, and regeneration (Fu et al., 2015; Gholipour et al., 2022; Li et al., 2017; Liu et al., 2014). Despite the volume of research conducted on the contribution of lncRNAs to the occurrence and development of IDD, their role still warrants clarification. Furthermore, substantial evidence indicates that lncRNAs regulate gene transcription by sponging miRNAs in IDD (Chen et al., 2015; Gholipour et al., 2022; Wang et al., 2015). Several studies have demonstrated the expression of different lncRNAs in degenerative and normal human NP samples, indicating their involvement in disk degeneration (Sampara et al., 2018).
In this review, lncRNAs are categorized into two groups based on their expression levels: upregulated and downregulated. This categorization helps to better understand the potential roles and regulatory functions of these lncRNAs in the biological processes and pathological mechanisms of IDD.
Upregulated lncRNAs
Several lncRNAs are upregulated in IDD progression, including lncRNA-CIR, RP11-296A18.3, PART1, HCG18, HOTTIP, GAS5, TUG1, ZFAS1, JPX, OIP5
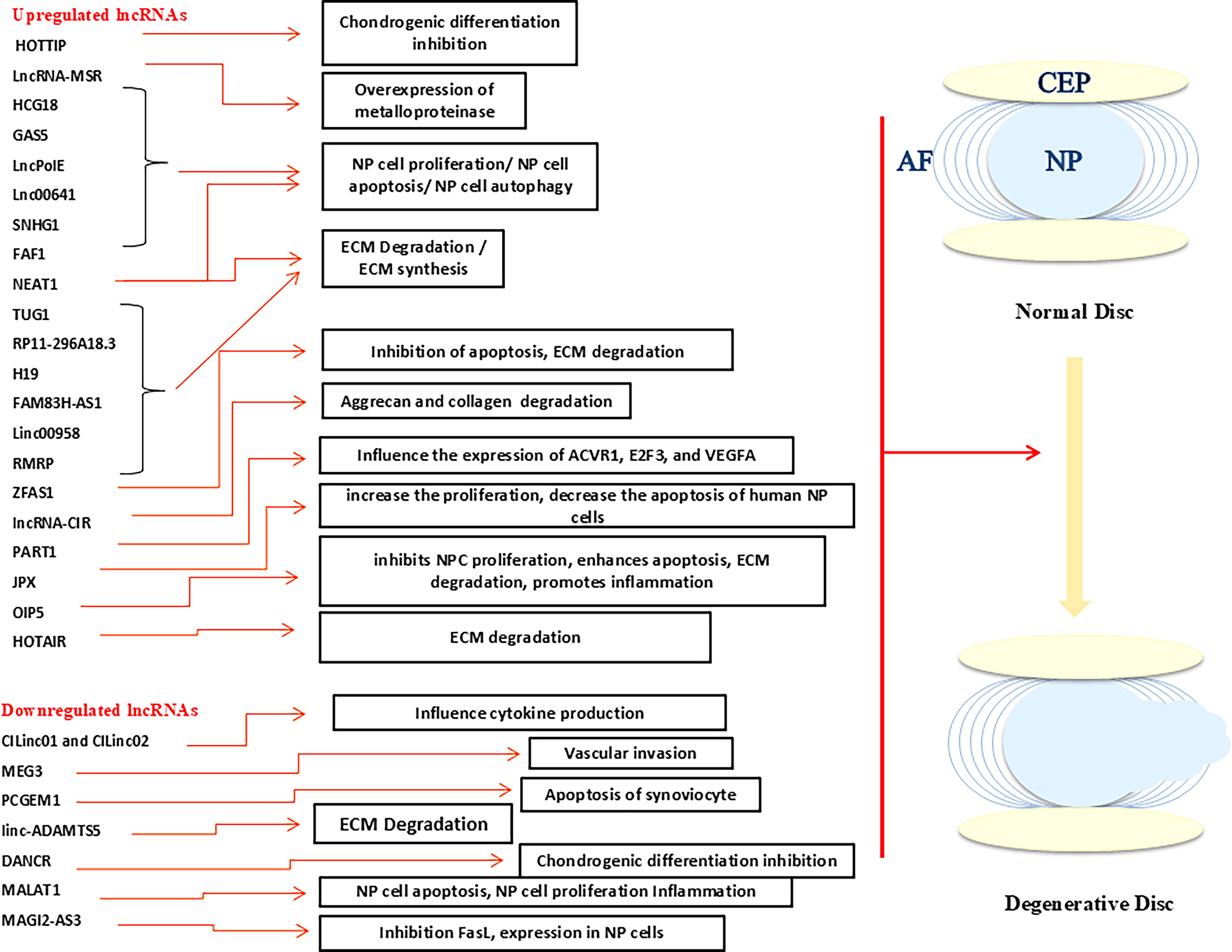
Schematic view of upregulated and downregulated long noncoding RNAs (lncRNAs) involved in the development of intervertebral disk degeneration (IDD). CEP, cartilage end plate; NP, nucleus pulposus; AF, annulus fibrosus.
Wang et al. (2017) found that RP11-296A18.3 increased HIF1A expression by acting as a sponge for miR-138, resulting in the promotion of neural progenitor cell proliferation and ECM synthesis. Hence, targeting RP11-296A18.3 in IDD tissues appears to be a promising approach for improving IDD.
PART1 regulates the expression of ACVR1, E2F3, and VEGFA through interaction with hsa-miR-34a and hsa-miR-148a (Zhao et al., 2016). It also negatively regulates the expression of miR-190a-3p (Zhang et al., 2021). lncRNA PART1 has been shown to have either an oncogenic or suppressive role in tumors (Jin et al., 2020; Pei et al., 2019; Sun et al., 2018; Zhang et al., 2021; Zhao et al., 2016), making it a valuable prognostic biomarker and therapeutic target in hepatocellular carcinoma, tongue squamous cell carcinoma, and nonsmall cell lung cancer. It is involved in the apoptosis and proliferation of prostate cancer cells. The expression of PART1 increases in patients with IDD, showing that this positive regulation occurs in the central NP tissue (Kang et al., 2018; Li et al., 2017; Lv et al., 2018; Sun et al., 2018; Zhang et al., 2019).
The lncRNA HCG18 functions as a molecular sponge that suppresses proliferation and promotes apoptosis of NP cells by interacting directly with miR-146a-5p, leading to increased IDD development. Additionally, HCG18 promotes ECM degradation by regulating the MIR4306/EPAS1 axis in degenerated NP cells (Cao et al., 2023; Xi et al., 2017).
HOTTIP is a regulator of HoxA genes; it concurrently upregulates and downregulates HoxA13, indicating the suppression of integrin-α1 in osteoarthritis (OA) chondrocytes. Cartilage development and destruction are regulated by HOTTIP through the modulation of integrin-α1, either through epigenetic regulation via DNMT-3B or transcriptional regulation via HoxA13. Similarly, suppression of miR-101 led to coordinated DNMT-3B activation and integrin-α1 repression, contributing to cartilage degeneration (Kim et al., 2013).
GAS5 negatively regulates cell survival and is expressed in several cancers. Song et al. showed that human GAS5 was upregulated in IDD and involved in apoptosis and autophagy by indirectly regulating miR-21. Wang et al. concluded that lncRNA GAS5 silenced NP cells and that GAS5 knockdown might be an effective therapeutic strategy for IDD treatment (Song et al., 2014; Wang et al., 2019).
TUG1 is a lncRNA whose regulatory gene is located on chromosome 22q12.2. An oncogene, TUG1 participates in cell proliferation, apoptosis, differentiation, and metastasis (Wang et al., 2017). TUG1 expression is significantly upregulated in degenerated intervertebral disk NP tissues and cells, indicating that TUG1 may play a role in the onset and progression of IDD (Tang et al., 2020). TUG1 is implicated in the apoptosis and ECM degradation of degenerated intervertebral disk NP cells by targeting the miR-26a/HMGB1 axis, which might be related to the NF-κB pathway activation. Recent studies have pointed to a potential specific regulatory connection between TUG1 and miR-26a (Tang et al., 2020). The Wnt/β-catenin pathway acts as a degeneration-promoting pathway in the process of IDD. The increase in TUG1 leads to the activation of this pathway, resulting in increased apoptosis, increased ECM degradation, and ultimately the progression of IDD. Therefore, blocking the Wnt/β-catenin pathway is considered a theoretical strategy for protecting NP cells against injuries caused by IDD (Chen et al., 2017).
JPX, a significant lncRNA that affects cell proliferation and apoptosis, is a critical element in IDD. It inhibits human NP cell proliferation and promotes apoptosis by regulating Hippo-YAP signaling via the miR-18a axis. Moreover, the regulatory role of miR-18a-5p/HIF-1A is such that JPX overexpression can increase the proliferation and decrease the apoptosis of human NP cells. In addition, the interaction between JPX and miR-18a-5p initiates a downstream signaling cascade and regulates proliferation and apoptosis in human NP cells (Yang et al., 2021).
One type of lncRNA is OIP5 antisense RNA 1 (OIP5-AS1). A prior study demonstrated that OIP5-AS1 was highly expressed in IDD and exerted its effects by reducing proliferation, accelerating apoptosis, promoting ECM degradation, and inducing inflammatory responses through the targeting of miR-25-3p (Che et al., 2021).
The activation of the Wnt/β-catenin pathway leads to IDD; accordingly, the inhibition of HOTAIR expression could be a potential target for IDD treatment. The results decisively demonstrate that HOTAIR, by activating the Wnt/β-catenin pathway, promotes disk degeneration through the induction of senescence, apoptosis, and ECM degradation. Consequently, silencing HOTAIR attenuates this degenerative process, positioning HOTAIR as a potential therapeutic target against IDD (Zhan et al., 2019).
lncRNA-MSR, a TMSB4 pseudogene, is upregulated in injured cartilage and stimulated in chondrocytes in response to mechanical stress. Liu et al. (2016) showed that in chondrocytes, lncRNA-MSR competitively regulated the expression of TMSB4 by interacting with miR-152.
The mammalian Polε complex is composed of four subunits: PolE1 (catalytic subunit), PolE2, PolE3, and PolE4. Li et al. (2019) proposed that the abnormal expression of lncPolE could play a role in the development of IDD by inhibiting PolE1 in iron-deficient environments.
Wang et al. (2019) revealed that long intergenic non-protein-coding RNA 641 (LINC00641) could affect collagen II and matrix metalloproteinase 3 expressions. NP cells show an increase in LINC00641 levels and a decrease in miR-153-3p levels. LINC00641 regulates autophagic cell death by targeting miR-153-3p and ATG5. Linc00641 may inhibit the proliferation and differentiation of C28/I2 cells by targeting miR-320a, consequently exacerbating cartilage tissue damage during OA. These findings could identify a novel therapeutic target for the management of arthritis (Han et al., 2024).
Tan et al. (2018) demonstrated that increased SNHG1 levels promoted the proliferation of NP cells by suppressing miR-326 expression. Additionally, they found a negative correlation between miR-326 expression and SNHG1 levels in tissues affected by disk degeneration.
During IDD development, reactive oxygen species can be produced in the form of H2O2 in NP cells. H19 could promote H2O2-induced degenerative alterations via cell senescence, increasing ADAMTS-5 and MMP levels and collagen I, while suppressing NP cell proliferation by activating the Wnt/β-catenin signaling pathway (Wang et al., 2018).
A prior study showed that the expression of FAF1 was increased in IDD and had a positive correlation with the severity of disk degeneration. This suggests that FAF1 could serve as a novel indicator for the early detection of IDD and as a promising therapeutic target (Mi et al., 2018).
Wei et al. (2019) discovered that FAM83H-AS1 expression was higher in degenerated NP samples than in normal NP samples. The study also observed a positive correlation between FAM83H-AS1 expression and disk degeneration grade. Furthermore, FAM83H-AS1 was found to promote NP cell growth and modulate ECM expression by targeting Notch 1.
Zhao et al. (2019) suggested that dysregulated LINC00958 expression might significantly contribute to IDD development by targeting miR-203 in NP cells.
Previous research proposed that lncRNA RMRP could accelerate the progression of IDD by interacting with miR-206, offering a promising new strategy for treating IDD (Wang et al., 2018).
Nuclear-enriched abundant transcript 1 (NEAT1) is essential for the integrity of the nuclear paraspeckle substructure. In lncRNA–mRNA microarray analyses, NEAT1 expression is higher in IDD. Ruan et al. showed that NEAT1-induced ECM degradation might involve the ERK1/2/MAPK signaling pathway. By preventing NP ECM degradation, NEAT1 could offer a new potential molecular target for IDD treatment (Lan et al., 2016; Naganuma and Hirose, 2013; Ruan et al., 2018; Shang et al., 2023).
The lncRNAs LINC00917 and CTD-2246P41.1 are crucial to IDD development by influencing SPHK1 to regulate angiogenesis (Chen et al., 2015). Chen et al. (2015) revealed that microarray analysis found the lncRNAs LINC00917, CTD-2246P4.1, CTC-523E23.5, RP4-639J15.1, RP11-363G2.4, AC005082.12, MIR132, and RP11-38F22.1 to be differentially expressed, suggesting they may have important roles in the development of IDD.
Downregulated lncRNAs
Some lncRNAs downregulated in IDD progression include MAGI2-AS3, CILinc01, CILinc02, MEG3, PCGEM1, linc-ADAMTS5, DANCR, and MALAT1 (Fig. 1) (Wang et al., 2017).
MAGI2-AS3 is a tumor-suppressor lncRNA in diseases such as breast and bladder cancer (Du et al., 2019; Wang et al., 2018). It is decreased in IDD and negatively regulates the expression of FasL in NP cells. As a result, the overexpression of MAGI2-AS3 might be effective for IDD treatment (Cui et al., 2020).
In knee OA cartilage, CILinc01 and CILinc02 show differing expression levels and regulate cytokine production during chondrocyte inflammation. This indicates that they may play a crucial role in mediating inflammation-induced cartilage degeneration in OA (Pearson et al., 2016).
Kang et al. (2016) demonstrated that PCGEM1 functioned as a sponge lncRNA for miR-770, controlling proliferation, apoptosis, and autophagy. They proposed PCGEM1 as a potential therapeutic target for IDD and OA.
Linc-ADAMTS5 and RREB1 are negatively correlated with ADAMTS5 expression in clinically relevant NP tissues. These findings suggest that RREB1 collaborates with linc-ADAMTS5, a ncRNAs, to suppress ADAMTS5 expression, ultimately impacting the ECM degeneration of the intervertebral disk (Wang et al., 2017).
SOX4 can directly bind to the promoter of lncRNA DANCR and augment its expression. Consequently, knocking down DANCR could reverse the stimulatory effect of SOX4 on the proliferation and chondrogenesis of synovium-derived mesenchymal stem cells. These data underscore the importance of SOX4 in the proliferation and differentiation of synovium-derived mesenchymal stem cells (Fan et al., 2018; Malakootian et al., 2018; Zhang et al., 2015).
MALAT1 levels are decreased in IDD NP cells. The upregulation of MALAT1 expression restrains IDD by suppressing inflammation, inhibiting NP cell apoptosis, and promoting cell proliferation (Zhang et al., 2017). All upregulated and downregulated lncRNAs significant in IDD development are presented in Table 1. Overall, lncRNAs play crucial roles in the pathogenesis and progression of spinal and skeletal diseases through the regulation of gene expression, modulation of the cell cycle and proliferation, influence on the ECM, regulation of inflammatory responses, and control of apoptosis and cellular senescence. These mechanisms position lncRNAs as promising therapeutic targets. Nevertheless, further research is essential to enhance our understanding of their biological functions and therapeutic potential (Ma et al., 2024; Yao et al., 2024).
Susceptible Long Noncoding RNAs in Intervertebral Disk Degeneration
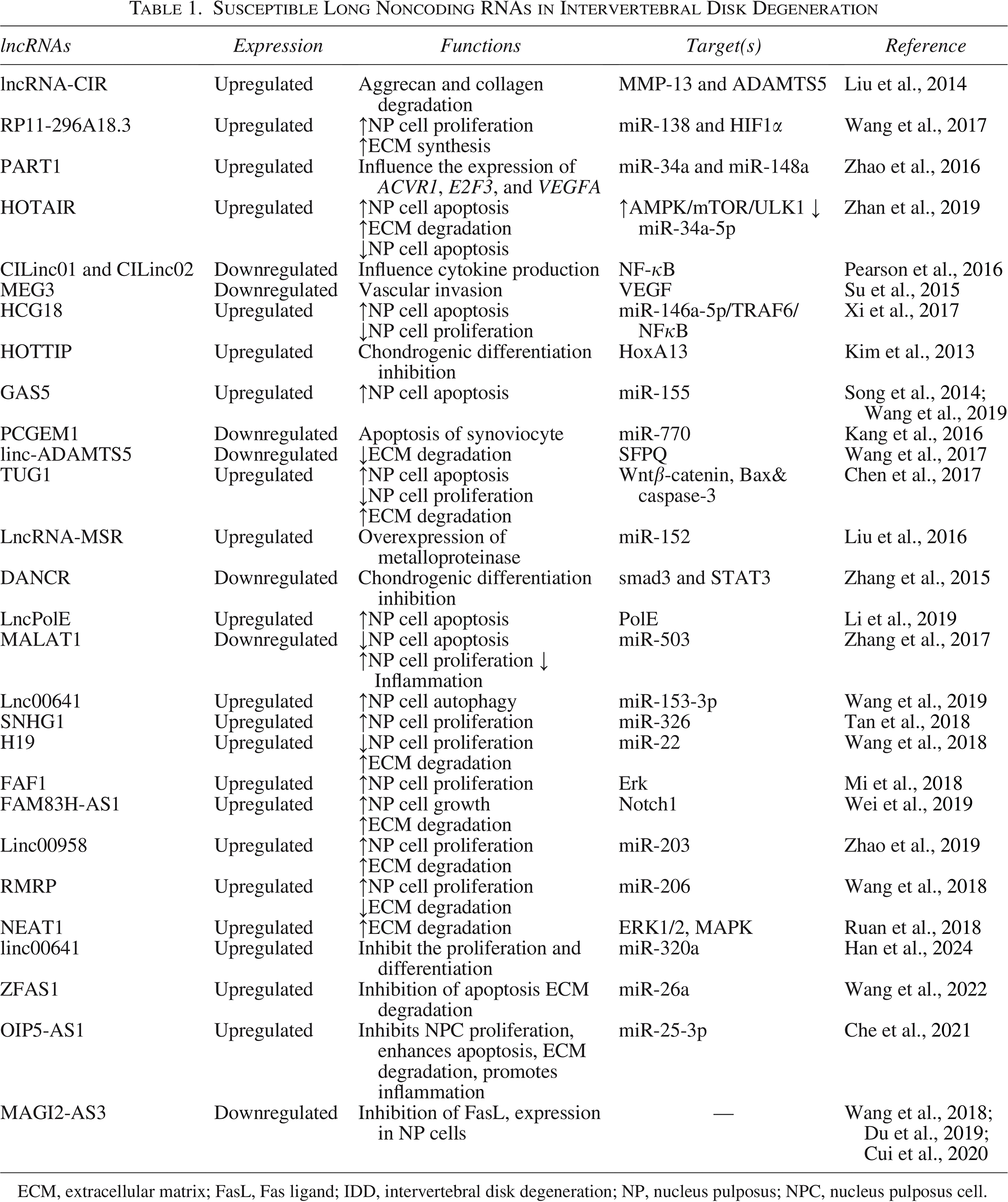
ECM, extracellular matrix; FasL, Fas ligand; IDD, intervertebral disk degeneration; NP, nucleus pulposus; NPC, nucleus pulposus cell.
circRNAs
circRNAs, another type of lncRNA, were first identified in virions infecting plants in 1976 by Sanger et al. (1976) Chiefly located in the cytoplasm, circRNAs have a closed circular structure connected through covalent bonds and lack open-access termini. They are resistant to hydrolysis by numerous RNA exonucleases and exhibit higher thermal stability than linear RNAs (Bagheri Moghaddam et al., 2022; Chen, 2020; Li et al., 2022).
circRNAs are divided into seven groups: (1) exonic circRNAs (the most common type, containing at least one exon from a single gene); (2) intronic circRNAs (generated through circularization of at least one intron from a single gene); (3) exonic–intronic circRNAs (comprising at least one exon and one intron); (4) intronic transfer RNAs (a special type of circRNA synthesized via the pretransfer RNA intron splicing mechanism); (5) read-through circRNAs (formed by the circularization of two exons from different genes); (6) fusion circRNAs (aberrant circRNAs produced after back-splicing of chimeric messenger RNA [mRNA] transcripts); and (7) mitochondrial circRNAs (produced from mitochondrial genes) (Jeck and Sharpless, 2014; Kristensen et al., 2019; Liu et al., 2020). Although the functions of circRNAs have not yet been fully defined, studies have demonstrated their involvement in numerous diseases, including OA (Shen et al., 2019), cancer (Shang et al., 2019), and cardiovascular (Aufiero et al., 2019; Bagheri Moghaddam et al., 2022), neurodegenerative, and immunological disorders (Akhter, 2018; Floris et al., 2017; Lodde et al., 2020). One well-established role of circRNAs is their function as competing endogenous RNAs that sponge miRNAs and regulate the expression of target mRNAs (Taheri Bajgan et al., 2022; Tang et al., 2020). In addition, circRNAs are significant regulators of IDD (Song et al., 2018), and their abundance in NP cells contributes substantially to the regulation of inflammation, NP cell apoptosis, and ECM metabolism—all processes involved in the development and progression of IDD (Li et al., 2022; Yang et al., 2021). Nonetheless, the precise role of circRNAs in IDD remains unclear.
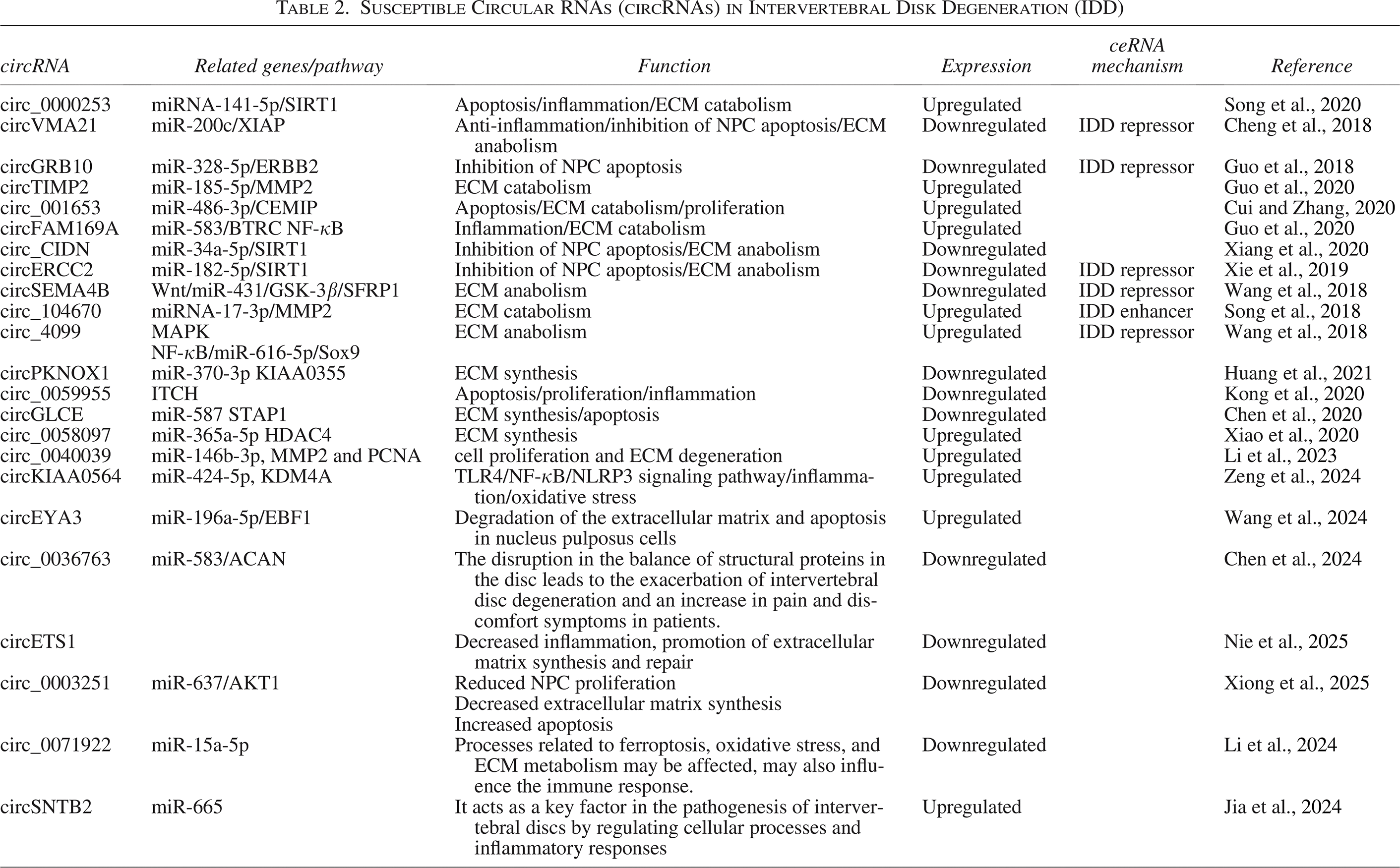
Song et al. (2020) demonstrated that exosomes derived from degenerative NP cells exhibited upregulation of exosomal circ_0000253, suggesting its potential role in promoting IDD through the absorption of miRNA-141-5p and subsequent downregulation of SIRT1. The expression of circTIMP2 is upregulated in IDD. Overexpression of circTIMP2 enhances ECM degradation and inhibits ECM synthesis. Correspondingly, miR-185-5p is sequestered by circTIMP2, potentially leading to upregulation of target genes associated with ECM degradation (Guo et al., 2020).
Cui and Zhang (2020) reported that the downregulation of circ_001653 could alleviate NP cell apoptosis. The miR-486-3p/CEMIP axis contributes to metabolic imbalance in the ECM. circFAM169A promotes IDD progression through the miR-583/BTRC signaling pathway (Guo et al., 2020). circ_104670 is highly expressed in NP tissues of IDD and acts as a competing endogenous RNA during NP degradation (Song et al., 2018). circ_4099 may promote ECM synthesis and secretion in NP cells, protect against IDD degeneration, and restore NP cell function (Wang et al., 2018).
Furthermore, circ_0058097 sponges miR-365a-5p and upregulates the expression of histone deacetylase 4 (HDAC4), a downstream target gene. Accordingly, it induces morphological changes in endplate chondrocytes and promotes ECM degradation and endplate cartilage degeneration (Xiao et al., 2020). Cheng et al. (2018) demonstrated that through the miR-200c–XIAP pathway, circVMA21 had the potential to reduce inflammatory cytokine–induced apoptosis of NP cells and restore the balance between anabolism and catabolism within the ECM.
In addition, the involvement of the circGRB10/miR-328-5p/ERBB2 signaling pathway in IDD progression suggests that circGRB10 could serve as a potential therapeutic target for IDD (Guo et al., 2018). In an ex vivo IDD model, circCIDN was shown to contribute to IDD development (Xiang et al., 2020). circCIDN acts as a sponge for miR-34a-5p, leading to compression-induced impairment of NP cells by inhibiting SIRT1 (Xiang et al., 2020). Xie et al. (2019) were the first to provide evidence indicating that activation of the miR-182-5p/SIRT1 axis through circERCC2 could ameliorate IDD by enhancing mitophagy and preventing apoptosis. The researchers suggested that circERCC2 might represent a promising therapeutic target for IDD. circSEMA4B acts as a miR-431 sponge, competing with SFRP1 or GSK-3β for miR-431 binding. Through the Wnt signaling pathway, it suppresses the degenerative process induced by IL-1β in NP cells (Wang et al., 2018).
Elevated levels of circPKNOX1 increase collagen II, aggrecan, and SOX9 expression while decreasing ADAMTS4, ADAMTS5, MMP3, and MMP13 expression. circPKNOX1 functions as a sponge for miR-370-3p, relieving inhibition of KIAA0355 expression and influencing IDD progression (Huang et al., 2021). Kong et al. (2020) reported that in vitro downregulation of circ_0059955 induced apoptosis and cell cycle arrest in NP cells, whereas in vivo overexpression of circ_0059955 reduced IDD in a puncture-induced rat model. Therefore, circ_0059955 may serve as a potential therapeutic target for IDD.
Furthermore, circGLCE targets the miR-587/STAP1 axis, leading to NP cell inhibition and ECM degradation, thereby mitigating IDD. circGLCE may serve as a potential therapeutic target for IDD treatment (Chen et al., 2020). Additionally, expression of circ_0040039 is elevated in NP tissues compared with normal tissues. This upregulation may contribute to IDD development by interfering with NP repair and regeneration through the upregulation of MMP2 and proliferating cell nuclear antigen mediated by miR-146b-3p (Li et al., 2023). circKIAA0564 is abnormally overexpressed in IDD and is associated with inflammation and oxidative stress in NP cells. It regulates KDM4A by competing for miR-424-5p and activates the TLR4/NF-κB/NLRP3 signaling pathway. Its reduction decreases inflammation and oxidative stress induced by IL-1β, and these findings may contribute to the development of targeted therapies for IDD (Zeng et al., 2024). circEYA3 plays a significant role in the progression of IDD by regulating the miR-196a-5p/EBF1 axis. EBF1 acts as a transcription factor involved in the degradation of the ECM and apoptosis in NP cells. The circEYA3/miR-196a-5p/EBF1 axis could be considered a novel therapeutic target for nonsurgical treatments of IDD (Wang et al., 2024).
circ_0036763, influenced by U2AF2 derived from exosomes of bone marrow mesenchymal stem cells, reduces IDD by regulating the miR-583/ACAN axis. These findings may serve as a novel therapeutic target for IDD (Chen et al., 2024). In a prior study, in both in vitro and in vivo experiments, PLGA microspheres loaded with si-circETS1 effectively delivered small interfering RNAs to NP tissue, which resulted in diminished circETS1 expression, decreased inflammation, promoted ECM synthesis and repair, and ultimately delayed IDD progression (Nie et al., 2025). According to the conducted research, the expression of circ_0003251 was significantly reduced in NP cells and degenerated NP tissues. Increased expression of circ_0003251 led to enhanced proliferation of NP cells, increased ECM synthesis, and reduced apoptosis. Mechanistically, circ_0003251 acted as a sponge for miR-637, inhibiting the suppression of AKT1 and reducing NP cell degeneration. PLGA microspheres successfully delivered circ_0003251, enhanced its therapeutic effect, and delayed the progression of IDD in vivo. circ_0003251 has been identified as a promising biomarker and therapeutic target for IDD. Delivery via PLGA microspheres ensures its effective application and presents a new strategy for the treatment of IDD (Xiong et al., 2025). The circ_0071922/miR-15a-5p–mRNA signaling network may influence IDD by regulating ECM metabolism, oxidative stress, and apoptosis in NP cells. These findings could serve as potential biomarkers and therapeutic targets for the management of IDD (Li et al., 2024).
One study found that circSNTB2 was significantly increased in NP tissues of the lumbar disc herniation (LDH) group and in NP cells treated with tumor necrosis factor α. miR-665 binds to circSNTB2, with cullin 4A (CUL4A) identified as a downstream target gene of miR-665. Reduction of circSNTB2 led to increased proliferation and decreased apoptosis of NP cells, an effect that was reversed by the inhibition of miR-665. Increased CUL4A counteracted the effects of miR-665 on the proliferation and apoptosis of NP cells. In vivo experiments showed that decreased circSNTB2 reduced thermal and mechanical pain caused by LDH and NP injury. By regulating miR-665 and CUL4A, circSNTB2 modulates the proliferation and apoptosis of NP cells, providing a rationale for targeted therapy of LDH (Jia et al., 2024). In general, studies have shown that several different circRNAs play a role in the progression of IDD (Table 2 and Figs. 2 and 3).
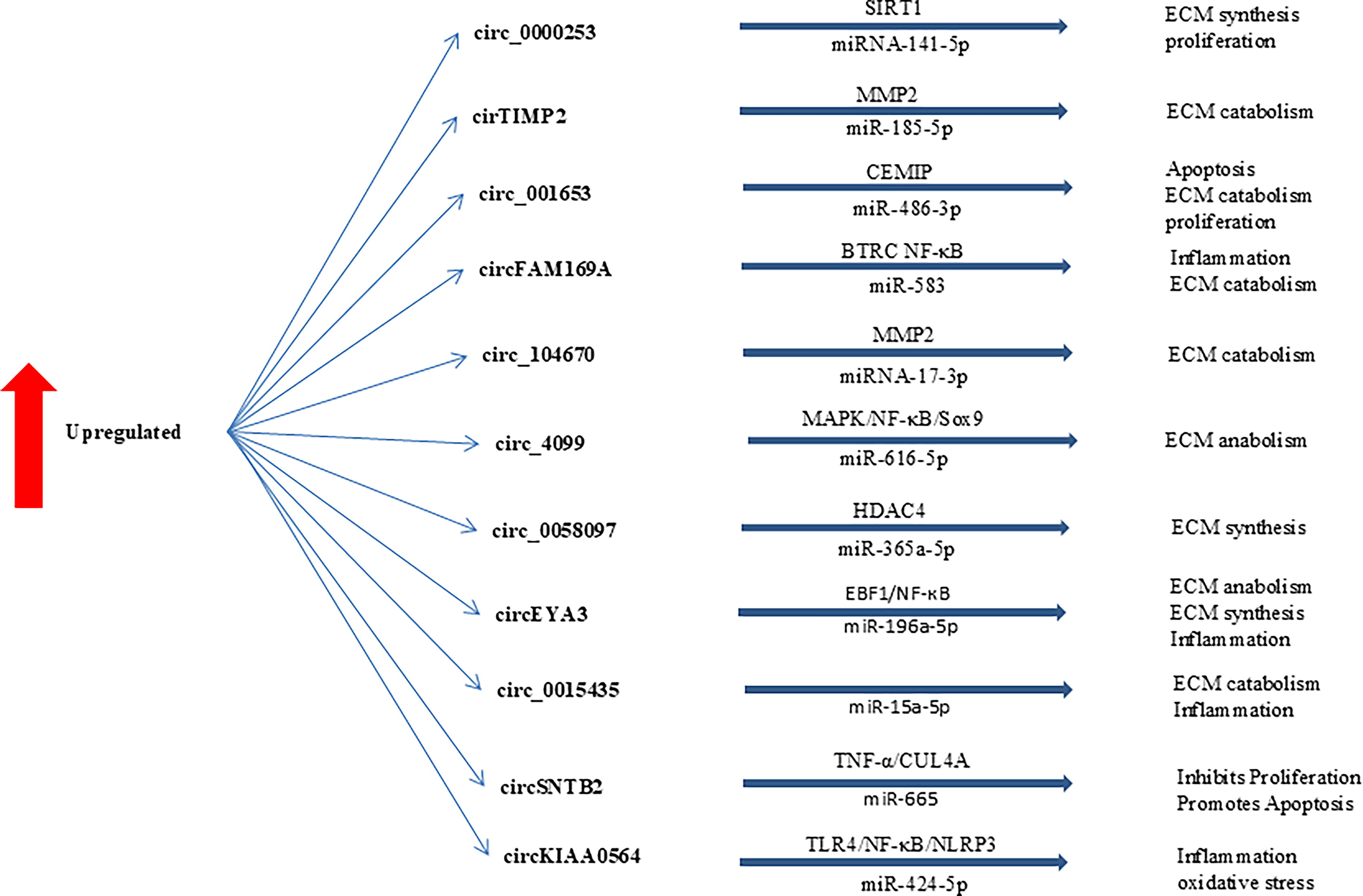
Upregulated circRNAs involved in the regulation of IDD development. ECM, extracellular matrix; IDD, intervertebral disk degeneration.
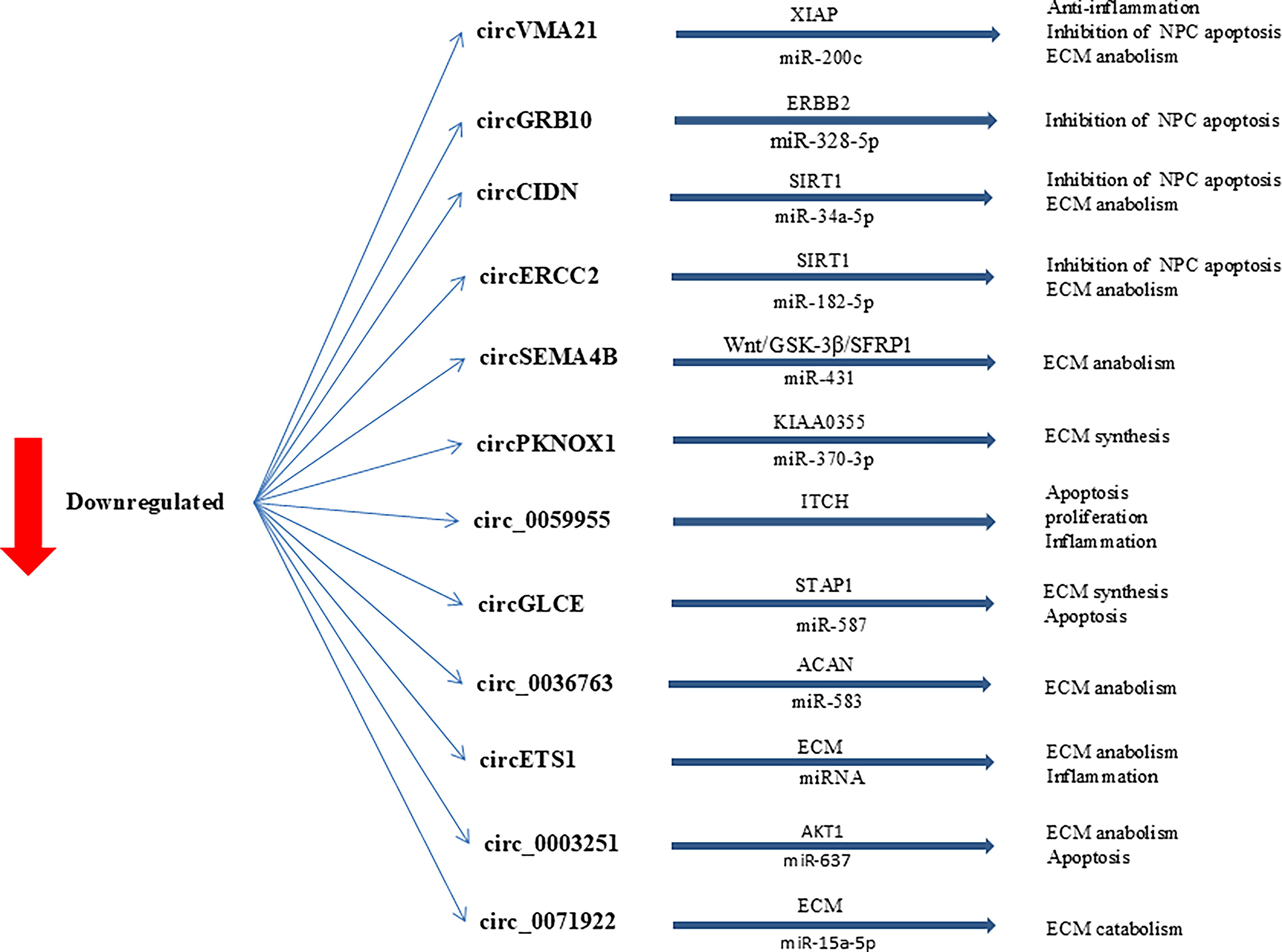
Downregulated circRNAs involved in the regulation of IDD development. ECM, extracellular matrix; IDD, intervertebral disk degeneration.
Susceptible Circular RNAs (circRNAs) in Intervertebral Disk Degeneration (IDD)
Due to their stability and critical regulatory roles in disc cellular homeostasis, NP cell proliferation, apoptosis, and ECM metabolism, circRNAs are an important research focus for IDD repair (Chang et al., 2025; Jiang et al., 2021). Targeting dysregulated lncRNAs and circRNAs offers novel therapeutic strategies, and modulating their expression could restore normal cellular functions (Jiang et al., 2022). The diagnostic and therapeutic potential of lncRNAs and circRNAs offers opportunities for early diagnosis, personalized therapy, and improved patient outcomes.
Limitations
lncRNAs and circRNAs discussed in this study were identified based on expression changes; nevertheless, their precise mechanisms of action and functional roles in IDD remain unclear. In addition, some studies lack experimental validation or clinical evidence to support the proposed therapeutic applications of lncRNAs and circRNAs.
Conclusions
IDD is a common degenerative condition characterized by deterioration of the intervertebral discs that separate the spinal vertebrae, leading to pain in the back or neck and often radiating to the legs or arms. Once a disc is damaged, it cannot regenerate. To date, no investigations have fully elucidated the precise mechanisms underlying IDD. Therefore, exploring the molecular and genetic markers involved in the disease’s development and early diagnosis is of considerable interest.
Further insights into the pathogenesis of degenerative disease may be achieved by clarifying the biological roles of ncRNAs in regulating the pathological processes of IDD and elucidating their underlying molecular mechanisms.
This knowledge could help devise prevention strategies and contribute to the development of molecular therapies. Numerous studies have attempted to understand the development and progression of IDD; still, additional research is needed to examine changes in the expression of both coding and noncoding genes in IDD to uncover its precise molecular signaling pathways and regulators.
Foreseeable future research could focus on developing machine learning models to predict novel lncRNAs and circRNAs that regulate IDD. Artificial intelligence–based tools can analyze large-scale transcriptomic datasets to identify key regulatory ncRNAs with high accuracy, providing comprehensive insights into their roles in IDD progression.
Footnotes
Author Disclosure Statement
All authors have read and approved the data presented in the article and declare that there are no conflicts of interest.
Funding Information
This work was supported by a research grant from Bam University of Medical Sciences (grant 401000007; IR.MUBAM.REC.1401.003) and the Research Deputyship of Rajaie Cardiovascular Institute.
Authors' Contributions
M.M., M.J., and A.G.: Contributed to databases search and the writing of the first and final draft and to write the tables and draw the figures. M.O.: Contributed to concept, the writing of the review, and supervising and revising the whole draft, figures, and tables. All the authors have reviewed the final manuscript.