
Abstract
Radiation therapy is a key treatment for many cancers, but patient responses vary due to differences in genomic stability and DNA repair capacity. Increasing evidence highlights the role of Alu elements, the most abundant short interspersed nuclear elements in the human genome, as regulators of cellular stress responses. This review summarizes current knowledge on Alu biology, including their structure, retrotransposition, methylation, and expression, with a focus on their activation by ionizing radiation and their influence on DNA damage, repair pathways, gene regulation, and immune signaling. Radiation exposure has been shown to activate Alu elements, resulting in genomic destabilization, impaired repair, and pro-inflammatory responses. Consistent with this, Alu-associated mutations and microsatellite instability were observed at loci linked to key cancer genes (e.g., TP53, BRCA2), aligning with increased loss of heterozygosity and exon mutational burden. In addition, Alu hypomethylation further promotes microsatellite instability, recombination, defective repair, and is associated with therapeutic resistance. In parallel, circulating cell-free DNA fragments containing Alu sequences have also been correlated with delivered radiation dose and clinical toxicity, underscoring their potential as minimally invasive biomarkers. Taken together, these findings position Alu activation as both a mechanistic driver of radio-response heterogeneity and a minimally invasive biomarker for biodosimetry and toxicity risk stratification.
Introduction
Radiotherapy (RT) remains a cornerstone of cancer treatment, administered to nearly half of all patients, curing about 40% compared with 49% by surgery and 11% by chemotherapy. Advances in delivery methods have transformed RT from basic two-dimensional external beam RT to highly precise techniques such as intensity-modulated RT, image-guided RT, stereotactic approaches, and proton therapy. These innovations allow dose escalation to tumors while sparing healthy tissues, particularly in complex anatomical sites and pediatric patients (West and Barnett, 2011). Despite these improvements, long-term complications including fibrosis, bowel obstruction, and incontinence continue to impair quality of life (Dalsania et al., 2021). Fibrosis may involve multiple organs, while acute side effects such as erythema, mild mucositis, dermatitis, and cystitis are usually reversible but may still interrupt treatment and worsen outcomes (Davy and Heathcote, 2021). Different procedures can be used to evaluate treatment-related toxicity, but each has its own limitations as discussed in Table 1. Thus, there is a need to unravel the molecular mechanisms of radiation-induced toxicity and to stratify patients according to the risks and complications. Current radiation dosing plans are mostly based on population-based models, especially the Quantitative Analysis of Normal Tissue Effects in the Clinic (QUANTEC) model, which correlates delivered doses with observed toxicities. Even though QUANTEC offers population-based dose-toxicity guidelines, it overlooks individual radiosensitivity. Reliable biodosimetric biomarkers are urgently required to personalize RT, enabling risk-adapted treatment and improving patient outcomes through biologically informed dose adjustments (Bentzen et al., 2010; Lockney et al., 2022). The development of reliable biodosimetric biomarkers promises to shift RT from population-based dosing toward truly personalized, risk-adapted treatment regimens that adjust doses according to a patient’s unique biological response.
Alu Insertions in Cancer-Related Genes Associated with Radiosensitivity (West and Barnett, 2011)

In recent years, circulating cell-free DNA (cfDNA) has been identified as a noninvasive biomarker to assess radiation-induced toxicity in cancer patients. Released into the bloodstream from dying cells through apoptosis and necrosis, cfDNA including tumor-derived fragments (ctDNA) can be dynamically monitored, offering insights into disease status and treatment response. Among cfDNA components, repetitive elements such as Alu sequences are one of the most prominent representatives of the cfDNA components because of their abundance and ubiquity throughout the entire genome. Alu elements, ∼300 bp SINEs comprising ∼10% of the genome, are abundant and detectable in cfDNA (Shaban et al., 2022; Fig. 1). Nonetheless, there are challenges of sequence similarity across subfamilies in addition to epigenomic complexity. Hypomethylation is a contributing factor to resistance to cancer (Park et al., 2021). Most recently, the Next-generation Sequencing of UnMethylated Alu method has been invented that is capable of specific recognition of unmethylated Alu repeats across over 130,000 individual Alu elements (Jorda et al., 2017). However, nonspecific binding to similar sequences often interferes with accurate identification and quantification (Gussakovsky and McKenna, 2021). Overcoming these technical limitations is essential for advancing the use of Alu RNA as reliable biomarkers in radiation biodosimetry and other clinical applications.
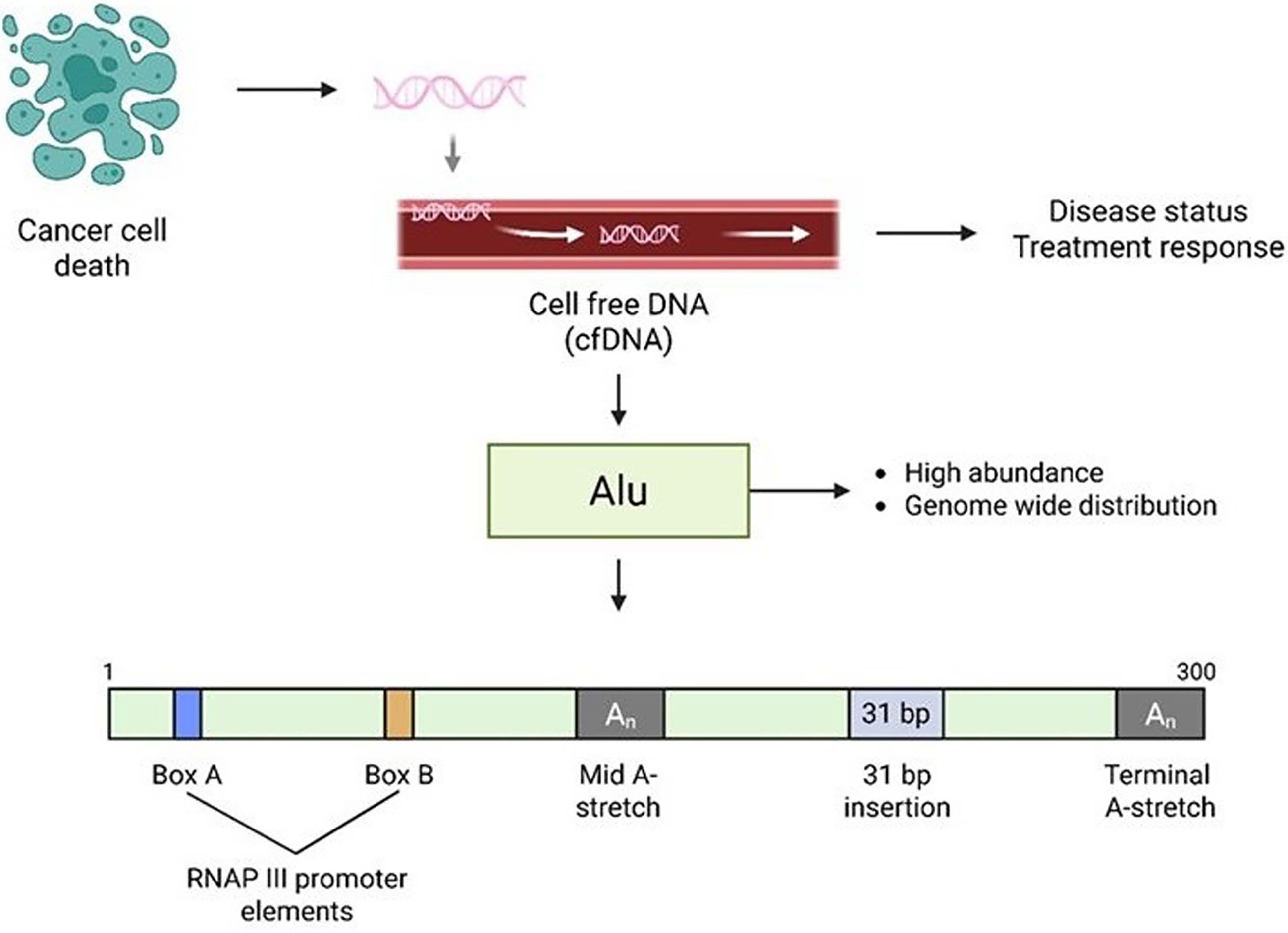
Schematic representation of Alu elements in cell-free DNA (cfDNA) as a biomarker for cancer. Following cancer cell death, fragmented genomic DNA is released into the circulation as cell-free DNA (cfDNA). Among these fragments, Alu elements are highly abundant and distributed throughout the genome, making them useful candidates for monitoring disease progression and treatment response. The Alu sequence includes RNA Polymerase III promoter elements (Box A and Box B), a mid A-stretch, a 31 bp insertion, and a terminal A-stretch. These features contribute to the transcriptional regulation and potential mutational impact of Alu insertions in the genome.
The review aims at discussing the biological rationale, clinical utility, and limitations of using Alu element quantification as a biomarker of radiation toxicity. We explore the potential of incorporating Alu-based cfDNA measurements into RT protocols to allow adaptive dose adjustments based on individual biological responses. In addition, we discuss future directions such as combining Alu quantification with advanced liquid biopsy techniques, genomic profiling, and AI-driven predictive models to improve the personalization of radiation therapy.
Structural Evolution and Multifaceted Roles of Alu Elements in Gene Regulation and Genome Dynamics
Alu elements are short interspersed nuclear elements (SINEs) named after the AluI restriction site, present in over half of these repeats. They are enriched in gene-dense regions of the human genome, often overlapping CpG islands and transcription factor motifs, and can function as enhancer-like elements whose activity evolves with age, supporting their role as proto-enhancers (Su et al., 2014). Alu sequences are originated from the 7SL RNA of the signal recognition particle (SRP), which regulates translation of nearly one-third of human proteins. Homology between 7SL RNA and Alu monomers suggests that deletions within ancestral 7SL RNA fragments gave rise to independent left and right Alu monomers, which later fused in a head-to-tail manner to form the modern dimeric structure (Hasler and Strub, 2006). Once integrated, Alu elements can retrotranspose through transcription by RNA polymerase II, binding to the SRP9/14 heterodimer, and recruitment to the 40S ribosome, where ORF2 protein mediates reverse transcription and reinsertion of Alu cDNA.
Among subfamilies, compared with Alu J, AluS, and AluY retain retrotranspositional capacity with AluY showing the highest activity due to its recent evolutionary origin (Lin et al., 2015). These findings suggest that AluS and AluY subfamilies demonstrate greater functional relevance in current genomes. In addition to mobility, Alu elements restructure the genome by promoting insertional mutagenesis, recombination, and epigenetic modifications. The present mechanisms of gene expression modifications by these subfamilies include cryptic exon splicing, mRNA decay, and formation of inverted repeats (Lee et al., 2024). Interestingly, over 90% of adenosine-to-inosine (A-to-I) edits that are catalyzed by Adenosine Deaminase Acting on RNA (ADAR) enzymes are also regulated by Alu-derived RNAs, which influence pluripotency. With the help of a number of splice sites, Alus create transcriptomic diversity, predominantly in 3′UTRs, which are regarded as important areas in post-transcriptional regulation (Gussakovsky and McKenna, 2021). Translation is also controlled by the Alu sequences using trans-acting ribonucleoproteins. Studying these processes is challenging because inverted Alu repeats in 3′UTRs form extensive dsRNA structures.
Techniques for Detecting and Characterizing Alu Elements
Understanding the molecular mechanisms underlying the diverse regulatory functions of Alu elements is essential for deciphering their clinical significance. To implement these mechanistic results in clinical practice, it is necessary to be aware of the experimental tools used to study Alu elements and their presence in biological fluids. Photoactivatable-Ribonucleoside-Enhanced Crosslinking (PAR-CLIP) (Fig. 2) has been used to map Alu RNA–protein interactions (Alu-RBP). In addition to PAR-CLIP, visible light cross-linking using methylene blue and formaldehyde treatment can also be used to map Alu-RBP interactions precisely (Guillemin et al., 2021; Hafner et al., 2010; Liu et al., 2000; Patton et al., 2020). These methods offer a mechanistic understanding of the impact of Alu transcripts on RNA stability, splicing, and translation. Thus, such techniques bridge basic Alu biology with their emerging clinical relevance.
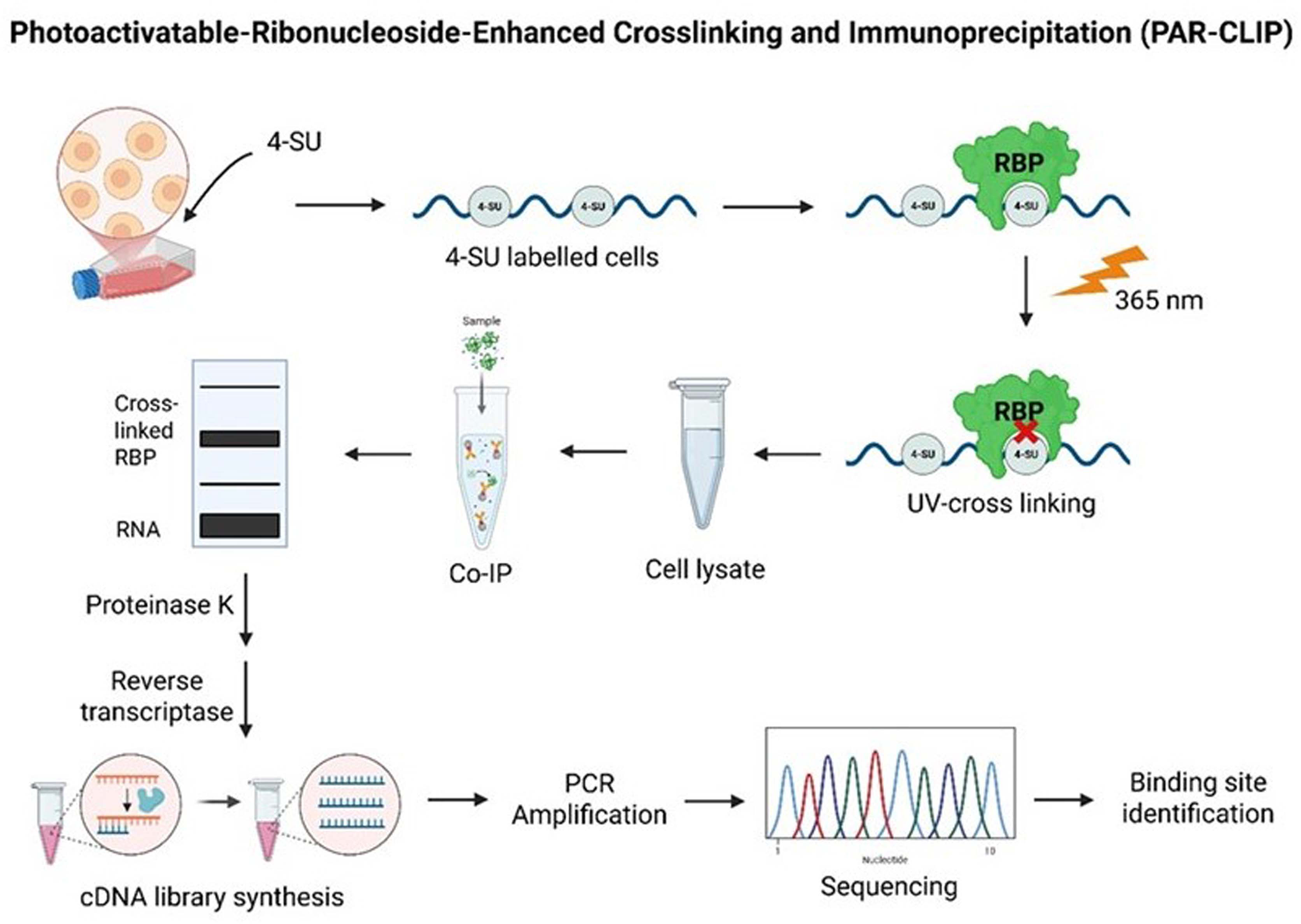
Schematic overview of the photoactivatable-ribonucleoside-enhanced crosslinking and immunoprecipitation (PAR-CLIP) method for identifying RNA-binding protein (RBP) interaction sites. Photoactivatable ribonucleoside analogue of 4-thiouridine (4-SU) is used to label cells and is inserted into nascent RNAs. When exposed to UV light at 365 nm, 4-SU-labeled RNA is covalently crosslinked to interacting RBPs. Thereafter, cells are lysed and the crosslinked RBP–RNA complexes are immunoprecipitated (Co-IP). The isolated RNAs are converted into cDNA libraries via reverse transcription after the proteinase K digestion. These libraries are PCR-amplified and sequenced to identify the exact binding sites of RBPs toward target RNAs.
With this background, more accurate detection of Alu insertions and other genetic changes can now be achieved using new sequencing methods. Advanced approaches, particularly optimized next-generation sequencing (NGS), now enable the reliable detection of rare mobile element insertions (MEIs), which are critical for patients with ancestral backgrounds predisposed to these variants and are often missed in conventional analyses. For instance, re-evaluation of NGS data of putative hereditary breast and ovarian cancer cases utilizing the MEI detection tool Mobster discovered a new Alu insertion in Ataxia Telangiectasia Mutated (gene/protein) (ATM) intron 54 (ATM:c.8010 + 30–8010 + 31 insAluYa5). Functional studies demonstrated that this variant drives skipping of exon 54, producing frameshift and probably nonfunctional transcripts. This variant was present in several unrelated individuals, suggesting its pathogenic potential (Klein et al., 2023). Recently, a strategy that integrates inter-Alu-PCR with specific NGS has been used to sensitively measure the presence of microsatellite instability (another characteristic of genomic instability in cancer). This approach allows for the efficient capture of poly A-containing genomic regions in short amplicons (80–150 bp), which is also suitable for low-input DNA from liquid biopsies. These approaches target polyadenine repeats located within the Alu elements. Alu-associated microsatellite instability (MSI) can be detected at extremely low levels of somatic indel frequencies (0.05–1.5%) using customized MSI-tracer software, which has both practical and diagnostic benefits (Yu et al., 2021). This highlights the importance of including MEI detection in clinical sequencing pipelines to enhance diagnostic outcomes and reveal hidden drivers of cancer susceptibility. In support of this, an investigation of gastric cancer established the utility of a branched DNA (bDNA)-based Alu assay for measuring the concentration of cell-free DNA (cfDNA) in the serum of patients. Cell-free DNA levels among gastric cancer patients were significantly higher than those in patients with benign gastric disease and healthy individuals (p < 0.05). Unlike other traditional tumor markers, such as Carcinoembryonic Antigen (CEA), CA19-9, CA72-4, and CA50, which have weak or no significant correlations, CFD is more sensitive, especially for the early detection of tumors (Qian et al., 2017a). Based on this concept, a new method, RealSeqS, has been developed to identify aneuploidy in plasma cfDNA, whereby approximately 350,000 repetitive elements are amplified using a single primer. To extend the usefulness of repetitive element analysis, researchers have proposed a machine learning framework, Alu Profile Learning Using Sequencing (A-PLUS), to learn more comprehensive differences in the profiles of repetitive elements in cancer patients and noncancer controls. A-PLUS was applied to more than 7600 samples of 11 tumor types, with a sensitivity of 40.5% and specificity of 98.5%. When combined with aneuploidy data and protein markers, the sensitivity improved to 51% (Rudin and Thompson, 2001).
Furthermore, imaging technologies now allow the visualization of radiation-induced changes in the chromatin architecture surrounding Alu elements, offering insights into DNA damage response mechanisms. For instance, to examine the radiation-induced dynamics of chromatin organization involving Alu elements, a combination of immunostaining and COMBO-FISH procedures has been developed. The procedure involves computer-designed probes to Alu consensus sequences. This method is also applied in combination with super-resolution localization microscopy, which allows visualization and quantitative analysis of Alu repeats and heterochromatin structures at the nanoscale level in irradiated cells (Hausmann et al., 2020; Krufczik et al., 2017). Similarly, radiation-induced chromatin changes in the SkBr3 breast cancer cell line have been analyzed using single-molecule localization microscopy. The interaction between GNPs and ionizing radiation resulted in a dose- and nanoparticle-dependent rearrangement of the chromatin network (Schafer et al., 2024). These techniques enable the tracking of dose-dependent chromatin rearrangements during DNA repair after exposure to low-linear energy transfer (LET) radiation. This can be of great value in comprehending the spatial dynamics of Alu elements in the radiation response.
It was also shown that real-time qPCR of circulating human Alu and LINE-1 DNA can be used as a valid method to determine the burden of subcutaneous and intratibial tumors in xenografts, the results of which are comparable to the standard methods of measuring human tumors (bioluminescence imaging and calipers) (Mishra et al., 2021). This underscores the emerging multifunctional role of Alu elements, not only in contributing to genomic instability and radiosensitivity but also as versatile tools for early detection and molecular monitoring of cancer through liquid biopsy.
Alu Elements and Genes Associated with Radiosensitivity
Alu elements and DNA damage response modulation
Inherited and acquired variations in radiosensitivity are largely governed by genetic differences in DNA repair, apoptosis, and oxidative stress pathways. Syndromes caused by mutations in key DNA repair genes, particularly those in the nonhomologous end joining pathway, such as XRCC2, XRCC3, XRCC4, XRCC5, XRCC8, and PRKDC are strongly linked to cellular radiosensitivity and immunodeficiency. Notably, mutations in ATM (as seen in ataxia-telangiectasia) result in extreme radiosensitivity (West and Barnett, 2011). Double-strand break (DSB) repair outcomes can be assessed by measuring both unrepaired and misrepaired DNA. In this context, Alu repetitive elements, which are abundant throughout the genome, serve as important markers in Southern blot-based assays to quantify unrepaired DSBs, offering a genome-wide view of persistent damage. Additionally, repair fidelity can be assessed by hybridization to a specific 3.2-Mbp NotI restriction fragment on chromosome 21. In a comparative study, fibroblasts from a radiosensitive breast cancer patient and ATM-deficient cells exhibited significantly higher levels of misrepair than normal controls, despite only modest increases in unrepaired DSBs detected via the Alu-based assay (Alsbeih et al., 2021).
Alu elements and genomic instability
Interestingly, Alu elements themselves may contribute to radiosensitivity by mediating genomic instability or modulating expression of genes involved in DNA repair and stress responses. In line with this, genomic profiling research has revealed that an even distribution of Alu elements is essential for ensuring genomic stability, and Alu deletions can be oncogenic initiators. A deletion of a single Alu at the 2000 bp site was observed in 83% of the cancer cases and 3% of the healthy controls in a study of 64 breast cancer patients and 64 healthy controls (p < 0.0001; OR 149), showing that it is strongly associated with cancer. Another less-intense band was observed at 1700 bp, which was more common in cancer patients (74%). In addition, the DNA banding patterns were found to be distinctly different in the blood samples of cancer patients and healthy individuals, but not in the tissue samples. These results indicate that Alu deletions in peripheral blood DNA can be used as early and noninvasive biomarkers of breast cancer and possibly as an indication of predisposition to radiation-induced genomic instability (Gazi et al., 2022). Beyond deletions, the repetitive nature and high copy number of Alu elements have enabled innovative molecular applications.
Transcription, splicing, or mRNA stability can be disturbed by Alu insertions near the regulatory regions or introns, resulting in MSI and loss of heterozygosity (LOH). An analysis of a microsatellite site within the TP53 intron (TP53-Alu) indicated that LOH was correlated with more frequent TP53 exon mutations in colorectal cancer (p = 0.0047), whereas MSI in TP53-introns was related to the occurrence of early stage disease (TNM II) (Liu et al., 2019). In breast cancer, longitudinal sampling of 123 patients revealed MSI in 70.7%, with 39.1% at TP53-Alu. LOH was often associated with alleles 399 and 404. Reduced expression of hMLH1, hMSH2, and TP53 was observed following chemotherapy, with a significant reduction in hMSH2, which was strongly correlated with MSI/LOH and secondary tumors. These results indicate that TP53-Alu can be used as a prognostic marker of genomic stress (Kamat et al., 2012).
Activation of Alu elements following radiation could interfere with DNA repair pathways, either by promoting aberrant A-to-I RNA editing and/or antisense transcription or chromatin remodeling. This dysregulation can disrupt homologous recombination, particularly in the case of Alu-mediated splicing or recombination of key genes, such as BRCA1 or ATM, which decreases the cellular radiosensitivity. Notably, over 100 retroelement (RE) insertions, including pathogenic Alu insertions, have been detected in cancer-related genes such as BRCA1, BRCA2, MSH2, and APC. One such striking instance is the Portuguese founder mutation c.156157 insAlu in BRCA2, which significantly increases the risk of hereditary breast and ovarian cancer. Emerging evidence indicates that several Alu insertions are population-specific. An example of this is BRCA2 c.34073408 insAlu, which is most frequently observed in people of African ancestry, and the analysis of the haplotype confirmed a shared lineage across 40 carriers. Similarly, BRCA2 c.94519452 insAlu and ATM c.73747375 insAlu are probable European foundational mutations, whereas BRCA2 c.50075008 insAlu demonstrates early indications of a Latin American background impact (Qian et al., 2017b). However, reports of such RE insertions are limited because of the limitations of conventional detection systems, including Array Comparative Genomic Hybridization (aCGH) and Multiplex Ligation-dependent Probe Amplification (MLPA). Thus, fundamental genetic testing should also involve the efficient detection and interpretation of RE insertions, which can accurately predict cancer risk.
Plasma cfDNA and alu elements as biomarkers of treatment toxicity in cancer
Although most decisions regarding the radiation dose in cancer therapy are based on physics-based dosimetry and population averages, they do not generally represent individual biological responses to therapy. This may lead to over- or under-treatment of some patients; hence, there is a need for personalized indicators to measure treatment effect and toxicity. One promising biomarker for this purpose is circulating cell-free DNA (cfDNA), which is released in response to tissue damage by radiation. In view of this a radiation toxicity assay, known as RadTox, was developed. It utilizes Alu retrotransposon sequences to measure the quantity of cfDNA. The results of this assay in preclinical models demonstrated that cfDNA levels correspond to the entire radiation dose in the body and increase in response to stressors, including trauma or sepsis.
Based on this, a clinical trial was conducted on patients with prostate cancer who underwent photon or proton RT. Prostate cancer was chosen based on the fact that radiation therapy is mainly used to treat Prostate cancer, allowing clearer attribution of toxicity to radiation rather than other treatments. The results demonstrated that the increase in cfDNA levels before and during the first week of therapy was rapid and patient-specific, with higher levels observed in patients receiving photon therapy than in those receiving proton therapy. These levels were positively related to the integral body dose, and a few patients were found to have abnormally high cfDNA at the onset of treatment, indicating a biological tendency toward radiotoxicity (Lockney et al., 2020; Paul et al., 2020). Further clinical evaluation in 54 prostate cancer patients revealed that baseline and early cfDNA levels were significantly correlated with grade 2 gastrointestinal (GI) toxicity, both acute and late, but not with genitourinary (GU) toxicity. Specifically, acute GI toxicity was associated with cfDNA levels measured on days 0–5 of treatment (p < 0.005), and late GI toxicity was linked to pretreatment cfDNA levels (p = 0.021). These results support the notion that cfDNA is a predictive biomarker of GI toxicity and indicate that it could be a real-time toximeter to supplement dose–volume histogram data and allow biologically informed treatment changes. A multi-institutional study is currently being conducted to explore this method further (Lockney et al., 2022). Another study in Rhesus macaques indicated that the level of cfDNA increased drastically following radiation exposure. A study using a branched DNA (bDNA) test revealed that the increase in cfDNA was dose-dependent and occurred within days following total body irradiation (1–6.5 Gy). Small doses (0–2 Gy) were distinctly differentiated from larger doses (5.5–6.5 Gy), which were potentially fatal. The alterations in the cfDNA of monkeys were similar to those observed in human patients following high radiation exposure, indicating that cfDNA may be used as a fast and dependable biomarker of early triage and dose evaluation across species (Okunieff et al., 2024). The test has already been used in the United States and the European Union in a Clinical Laboratory Improvement Amendments-approved format. However, certain challenges remain. These challenges can be addressed by simplifying the sample processing procedures, enhancing the assessment of the test, and setting more precise values of the cfDNA cutoff to correspond to various levels of radiation exposure to improve triage and risk assessment (Okunieff et al., 2024). In prostate RT, rises in Alu cfDNA had the strongest relationship with GI toxicity compared with genitourinary toxicity, which is in line with the idea of rectal lining mucosal damage rather than cancer cell damage. These observations directly favor the application of Alu cfDNA as a surrogate endpoint for normal tissue radiosensitivity and radiation toxicity. Notably, these observations are the reason why it is believed that changes in circulating Alu cfDNA are strongly indicative of radiation-induced damage to normal tissues and not a widespread cell death.
Alternative cfDNA approaches beyond alu elements
Recent studies have explored alternative methods for monitoring treatment response and radiation-induced injury using alternative cfDNA-based methods, focusing on genome-wide or methylation-specific signatures rather than Alu-derived cfDNA. In such studies, repetitive element quantification was not used, but rather the indices of chromosomal instability and cell-type-specific patterns of methylation to learn more about the radiation-induced molecular changes. In patients receiving RT for lung, esophageal, and head and neck cancers, genome-wide instability quantified as an I-score demonstrated a dramatic decrease one month after RT, which was correlated with a decrease in tumor burden and indicated the potential of using cfDNA as a means of detecting minimal residual disease (Cho et al., 2023). In another study, methylation-defined brain-derived cfDNA (BncfDNA) was also evaluated as a predictor of radiation-induced brain injury (RBI) in patients having brain metastases. High levels of BncfDNA were measured during periods of RBI and reduced after the patient recovered, regardless of tumor regression, showing that cfDNA can indicate normal tissue damage caused by radiation (Makranz et al., 2023). Similarly, cell-type-specific methylation mapping of the circulating DNA fragments of endothelial, epithelial, and liver cell types showed dose-dependent and tissue-specific increases in circulating DNA fragments of these cell types in both animal models and in breast cancer patients after radiation (McNamara et al., 2023). These findings highlight that cfDNA composition dynamically reflects tissue damage beyond the tumor site and can serve as a sensitive biomarker for radiation-induced systemic effects. Although these studies did not entail the Alu-based measurement of cfDNA, they all contribute to the general idea that cfDNA is indicative of tumor response and radiation-induced tissue damage. Collectively, these studies offer useful complementary evidence to support the greater clinical potential of cfDNA as a noninvasive biomarker in radiation oncology, which is consistent and complementary to the current attempts to investigate various cfDNA signatures, such as Alu elements, to accurately track the effects of RT.
Diagnostic, prognostic, and therapeutic monitoring potential of alu cfDNA
Beyond radiation therapy, cfDNA is gaining traction as a non-invasive liquid biopsy tool in oncology for monitoring disease progression and treatment responses. In a meta-analysis of 36 studies, it was revealed that ALU sequence levels, particularly ALU 115 and ALU 247, were significantly elevated in cancer patients compared with healthy individuals, those with benign conditions, or patients at different cancer stages (Shaban et al., 2022). In patients with locally confined breast cancer undergoing neoadjuvant chemotherapy, dynamic changes in cfDNA, particularly ALU 115 and ALU 247 fragment levels, were found to correlate with therapeutic outcomes. In a study of 65 patients, kinetics of ALU 115 from cycle 1 to 6 of chemotherapy decreased in patients who achieved complete remission, but increased in nonresponders (p = 0.033). Response was, however, not predicted by DNA integrity indices and traditional tumor markers such as CEA and CA15-3 (Lehner et al., 2013). On the same note, comparing patients with lung or breast cancer to healthy people, both Alu index and LINE-1 methylation levels were also found to be very different. Hypomethylation at LINE-1 demonstrated better diagnostic capacity of lung and breast cancer with AUCs of 0.848 and 0.890, respectively, whereas the value of Alu index was 0.742 and 0.724 in the same order. Although longer Alu fragment concentrations were found to be associated with the Alu index, regression analysis indicated that LINE-1 methylation was a better marker of these cancers (Park et al., 2021).
The results are reinforced by a bigger sample of 167 breast cancer patients, in which ALU 247 and DNA integrity index were significantly reduced in the postoperative plasma samples in comparison with their presurgical counterparts. Higher ALU 247 levels were associated with metastasis, poor disease-free survival (p = 0.005), and lower tumor-infiltrating lymphocytes in ER-negative subtypes (p = 0.01). ALU 247 emerged as an independent prognostic factor in both univariate and multivariate analyses [HR 1.3, p = 0.017], and its inclusion in nomogram models significantly improved prediction accuracy (C-index = 0.823) (Nair et al., 2023). Recently, we have also shown that DNA integrity index was higher for chemoresistant oral cancer samples when compared with chemosensitive samples (Patel et al., 2024).
Recent evidence supports the diagnostic and prognostic value of Alu-based cfDNA quantification in cancer. The systematic meta-analysis by Taghizadeh-Teymorloei et al., demonstrated that the circulatory Alu cfDNA (in particular, ALU115 and ALU247) had a high pooled sensitivity (0.88) and specificity (0.86) for the detection of colorectal cancer (Taghizadeh-Teymorloei et al., 2024). Moreover, a high cfDNA integrity index was strongly associated with low survival rates and advanced disease stage, suggesting the prognostic utility of Alu-derived cfDNA. These results point to the possibility of Alu cfDNA as a translational, strong, and minimally invasive biomarker in disease diagnostics, prognostics, and monitoring of treatment-related toxicities. Together, these findings highlight the broader clinical utility of cfDNA and Alu-based measurements as minimally invasive tools for early prediction of toxicity, treatment response, and long-term outcomes across different cancer types. By offering real-time, biologically relevant insights, cfDNA profiling has the potential to transform cancer care through personalized monitoring and therapeutic adaptation.
Mechanistic Basis for Alu-Mediated Radioresistance
The role of Alu elements in therapy resistance, such as radioresistance, is increasingly recognized, but their mechanisms are complex. Alu-mediated genomic instability, whether caused by insertional mutagenesis, nonallelic homologous recombination, or hypomethylation-mediated activation, is a possible hypothesis that could explain the loss or silencing of tumor suppressor genes (including ATM, and BRCA1/2) (Oubaddou et al., 2023; Pastor et al., 2009). These perturbations affect the fidelity of DNA repair processes and promote cell survival after genotoxic stress. Radiation-induced transcription of Alu RNAs produces double-stranded RNA (dsRNA) intermediates, which in turn can activate innate immune sensors, such as MDA5 and TLR3, and induce a paradoxical prosurvival inflammatory response to type I interferon signaling and chronic Nuclear Factor kappa-light-chain-enhancer of activated B cells (NF-kB) activation (Aune et al., 2022). Such inflammation can transform the tumor immune microenvironment by augmenting immunosuppressive cytokines (e.g., IL-6 and IL-10) and bringing inflammatory control immune cells together, allowing tumor immune evasion (Landskron et al., 2014). Moreover, RNAs derived from Alu may complex with RBPs and disrupt the stability of RNAs and their translation, such as those of stress response regulators, such as HSP70 or BCL2, to promote postradiation survival (Goswami et al., 2023; Pandey et al., 2011). The combination of these mechanisms indicates that Alu activation during radiation stress can both destabilize the genome and reprogram the immune microenvironment, contributing to radioresistance and treatment failure.
Alu DNA methylation and therapeutic perspectives
Environmental exposures and therapeutic interventions such as radiation have significant impacts on DNA methylation patterns, particularly within repetitive elements, with important consequences for genomic stability and cancer progression. RT, one of the pillars of cancer treatment, is known to cause both genetic and epigenetic alterations that can contribute to heterogeneity, radioresistance, and relapse in tumors (Aypar et al., 2011). DNA hypomethylation causes genomic instability, which accelerates aging and cancer. In this regard, a new biotechnology approach using Alu-targeting small interfering RNA (siRNA) has been established to reprogram the methylation of Alu repetitive elements within human cells. This method showed a negative relationship between the level of Alu methylation and endogenous DNA damage in normal white blood cells. Alu-targeting siRNA treatment increased Alu methylation levels, enhanced cell growth, alleviated DNA damage, and improved resistance to genotoxic stress. This indicates that Alu methylation may be used to stabilize the genome by restoring it, which would have potential applications in normalizing radiation-induced toxicity of normal tissues and in reversing the genomic instability observed in aging in regards to hypomethylation. However, this is not the case in cancer cells. Alu methylation is already reduced in many tumors, leading to the activation of these elements. This action can facilitate genomic instability and allow cancer cells to resist therapy (Jorda et al., 2017). In these instances, further enhancement of Alu activity may render cancer cells sensitive to radiation or chemotherapy. Therefore, Alu-targeted therapies may have two roles: (1) to protect normal cell activities by enhancing methylation and (2) targeting cancer cells by taking advantage of their unstable Alu activity (Patchsung et al., 2018).
Recent investigations into pediatric glioma stem cells exposed to repeated radiation doses revealed that while a few fractions had minimal effects, extended fractionated radiation led to widespread DNA methylation changes. These mutations were localized in regulatory areas, such as enhancers, alternative splicing sites, and repetitive sequences and accompanied by dose-dependent morphological and proliferative changes. This type of epigenetic reprogramming is probably a cellular adaptation to radiation-induced stress that could affect treatment outcomes (Danielsson et al., 2020). Together with findings from studies on radiofrequency electromagnetic fields (RF-EMF), which showed that RF-EMF exposure selectively alters methylation of LINE-1 and ribosomal repeats but not Alu elements, these data underscore the dynamic nature of repetitive element methylation in response to environmental and therapeutic radiation (Ravaioli et al., 2023). Understanding and manipulating these epigenetic responses, for example, with the aid of Alu siRNA-mediated methylation recovery, could offer new approaches to enhance genomic stability, overcome therapy resistance, and improve therapeutic efficacy in cancer.
Therapeutic Implications and Future Directions
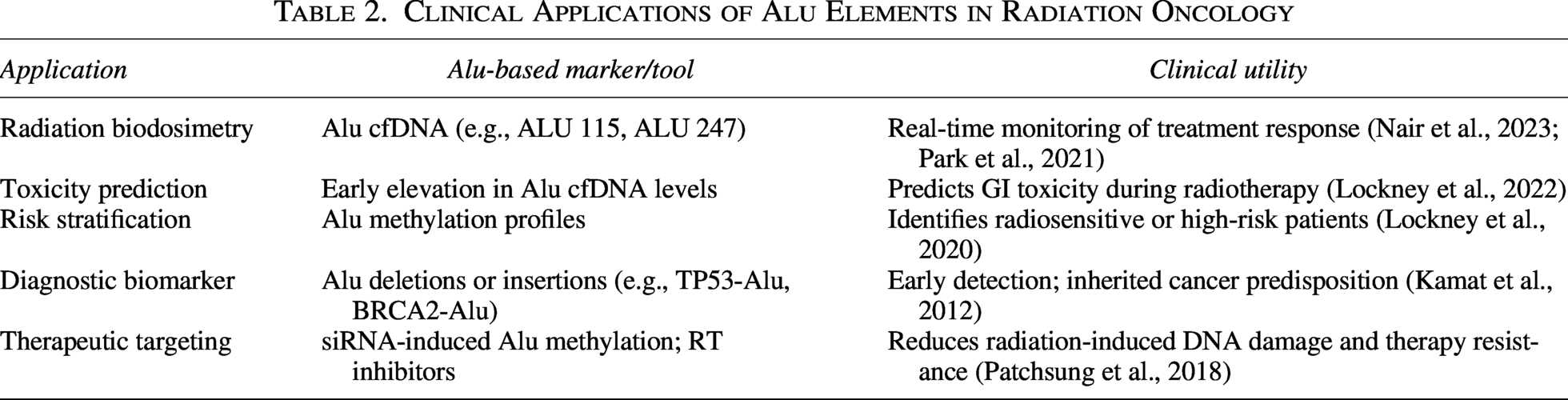
Understanding the role of Alu elements in the radiation response opens new possibilities for personalized cancer therapy. Targeting Alu activity may help reduce radiation-induced damage and improve treatment effectiveness. A potential solution is restoring Alu methylation status with siRNAs, which has been observed to reduce DNA damage and improve genomic stability in experimental animal models. In addition, reverse transcriptase-inhibitory drugs, which are widely used in antiviral therapy, can prevent Alu retrotransposition and mutagenesis. There is potential of silencing Alu transcripts that cause therapy resistance through the use of RNA-targeting methods, including antisense oligonucleotides. Monitoring Alu-derived cfDNA in blood can result in real-time adjustments to the radiation dose. The integration of these molecular approaches with conventional RT can serve to overcome the resistance and minimize the toxicity. Future studies should aim to establish the validity of these strategies in clinical practice, as well as the dynamics of Alu element regulation with other cell signaling pathways. Incorporating Alu-based diagnostics and therapeutics into cancer treatment will bring us one step closer to real-life personalized and adaptive RT. Table 2 provides an overview of the clinical applications of Alu elements in cancer (West and Barnett, 2011).
Clinical Applications of Alu Elements in Radiation Oncology
Conclusion
Alu elements, once considered non-functional repetitive sequences, are now recognized as active regulators of genome stability, gene expression, and cellular responses to stress, including radiation. Their activation by ionizing radiation contributes to DNA damage, defective repair processes, and disturbed immune signaling, which can underlie variations in RT effects. The presence of Alu sequences in circulating cell-free DNA offers a promising, minimally invasive tool for assessing radiation-induced toxicity and predicting treatment response. Furthermore, experimental strategies aimed at restoring Alu methylation or suppressing Alu activity have shown potential for reducing genomic instability and enhancing radioprotection. As our understanding of Alu biology deepens, these elements may serve not only as biomarkers but also as therapeutic targets. The future integration of Alu-based assays into clinical workflows could enable more precise and personalized RT and improve the management of radiation-related side effects. Continued research on the functional consequences of Alu activation will be critical for advancing both diagnostics and therapeutics in oncology.
Authors’ Contributions
M.P. collected the related article. C.J., M.P., and S.P. drafted and wrote the article. R.G. contributed to the design of the review and assisted in drafting and revising the article. All authors reviewed and approved the final version.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
No funding was received.