
Abstract
Introduction:
The collection and preservation of postmortem genetic material from recently deceased animals of rare and endangered species represent a critical yet underexplored avenue in conservation biology. While extensive research has been conducted on the human postmortem interval (PMI), there is a notable gap in understanding the postmortem preservation of germplasm in endangered species.
Objectives:
This study aimed to investigate the dynamics of apoptosis in various tissues of the Yangtze sturgeon at different postmortem time points, and to provide a reference for identifying the optimal time window for germplasm preservation in rare and endangered fish in the wild.
Methods:
Terminal deoxynucleotidyl transferase-mediated deoxyuridine triphosphated nick-end labeling assay and tissue morphology analyses were used to investigate apoptosis in the brain, heart, fin, liver, gonad, muscle, spleen, and skin of the Yangtze sturgeon at five different time points 0, 4, 8, 12, and 16 hours postmortem.
Results:
The results revealed a dynamic pattern of apoptosis. All tissues exhibited a time-dependent increase in apoptotic rate, indicating a clear correlation between PMI and apoptosis progression. This temporal pattern underscores the importance of timely genetic resource preservation, as the integrity of genetic material deteriorates progressively after death. Histomorphological analysis further demonstrated progressive degradation of tissue structure, especially in metabolically active tissues such as the gonad and fin.
Conclusion:
Based on the findings, we recommend that the genetic resources of the Yangtze sturgeon be preserved as soon as possible after death, particularly within the first 12 hours when tissue integrity remains sufficient for viable cell isolation or cryopreservation. This window is critical for metabolically active tissues, which show marked changes over time and may be important for postmortem identification. Further research should explore cryopreservation and antioxidant treatments to extend the preservation window for germplasm resources, ensuring the long-term viability of these valuable genetic materials.
Introduction
Yangtze sturgeon (Acipenser dabryanus), also known as Dabry’s sturgeon, is a critically endangered flagship species endemic to the Yangtze River, historically distributed across its main stem and several tributaries. 1 However, by the late 20th century, natural reproduction of the Yangtze sturgeon had ceased due to the construction of water conservancy projects, overfishing, and significant habitat alterations, indicating that the wild population was unable to be self-sustaining.2,3 In July 2022, Yangtze sturgeon was affirmed to be extinct in the wild by the International Union for Conservation of Nature, 4 meaning that no viable, naturally reproducing wild populations remain.
Despite the implementation of numerous policies and protective measures, such as an artificial propagation and release program and the action plan for conservation of the Yangtze sturgeon (2015–2030), the wild population remains unrecovered in the short term. Furthermore, the species’ genetic diversity continues to decline due to potential inbreeding in captive populations. As a result, the continuation of the Yangtze sturgeon species faces more severe challenges. It is urgent to maintain the diversity of the Yangtze sturgeon and protect its germplasm resources. Current conservation efforts primarily focus on habitat restoration,5,6 preservation of living organisms, 7 and physiological and biochemical research.8,9 However, research on the rescue and conservation of Yangtze sturgeon germplasm resources remains insufficient relative to the species’ critical conservation needs. 10
In vitro preservation methods for germplasm resources typically involve the storage of specimens, tissue samples, 11 cells,12–14 or nucleic acids. 15 However, the application of these methods often lacks precision and may be applied randomly or without strategic planning, particularly in response to sudden mortality events caused by pathogenic infections, accidents at breeding facilities, or other unforeseen circumstances. Such uncoordinated approaches can result in missed opportunities to preserve tissues and organs with higher viability, thereby hindering the effective rescue and conservation of endangered species.
Given the limited research on postmortem germplasm preservation in fish, we sought to adapt methodologies16–19 commonly used in human postmortem interval (PMI) studies in forensic science and criminal investigation. In this study, the TUNEL (terminal deoxynucleotidyl transferase-mediated deoxyuridine triphosphate [dUTP] nick-end labeling) assay and morphological changes were used to investigate apoptosis in the brain, heart, fin ray, liver, gonad, muscle, spleen, and epidermis of 1-year-old juvenile Yangtze sturgeon at five PMIs: 0, 4, 8, 12, and 16 hours. Our findings offer a novel theoretical framework for determining the optimal time window for the rescue and preservation of endangered fish germplasm under extreme or postmortem conditions. The results indicate that, under ambient conditions of 25°C, the most effective period for preserving cellular resources in the Yangtze sturgeon is within the first 12 hours following death. Beyond this window, tissue degradation may limit their suitability for cell-based applications. This study not only provides a scientific basis for germplasm preservation in the Yangtze sturgeon but also offers valuable insights for developing preservation strategies for genetic materials in other endangered aquatic species.
Materials and Methods
Experimental fish and treatment
The fish used in the experiment were 1-year-old juvenile Yangtze sturgeon, obtained from the breeding base of the Chinese Sturgeon Research Institute, China Three Gorges Group Co., Ltd. (Yichang, Hubei). All individuals were confirmed to be healthy based on external examination, showing no visible signs of disease, injury, or abnormal behavior. The 25 juveniles were reared in circular concrete tanks (area: 8.3 m2; water depth: 0.42 m; volume ≈ 3486 L) for 2 weeks during which they were fed a commercial juvenile pellet feed (Shandong Shengsuo Feed Technology Co., Ltd., China), with a crude protein content ≥42% and crude fat ≥8%, twice daily to apparent satiation. The temperature of the dechlorinated tap water was maintained at 25°C (±0.5°C), and dissolved oxygen was maintained above 5.0 mg/L. After acclimatization, 25 fish (28.8 cm ± 2.26 cm average in length, 251.2 g ± 32.8 g average in weight) were randomly selected, and 5 fish were selected at each time interval point for testing.
Tissue preparation and histology
Juvenile Yangtze sturgeon were euthanized using tricaine methanesulfonate (MS-222) at a concentration of 200 mg/L at a room temperature of 25°C. To ensure animal welfare, the MS-222 solution was buffered to a neutral pH (approximately pH 7.0) using sodium bicarbonate before use. Fish were exposed to the buffered solution for 10–25 minutes until complete cessation of movement and response to external stimuli was observed, confirming euthanasia. Following euthanasia, the sturgeons were placed in a breeding tank under controlled environmental conditions. The room temperature was consistently maintained at approximately 25°C throughout the 16-hour observation period to ensure stable storage conditions and minimize external factors that could affect sample quality. Samples of eight tissues: brain, heart, fin, liver, gonad, muscle, spleen, and epidermis, were collected at 0, 4, 8, 12, and 16 hours postmortem. The tissues were trimmed into 5–8 mm blocks and fixed in 4% (w/v) paraformaldehyde for 20–30 hours. Following fixation, the tissues were washed three times with Phosphate-Buffered Saline (PBS) buffer to remove residual fixative. Dehydration was then performed using a graded ethanol series, after which the tissues were transferred into molten paraffin wax maintained at 56°C. Continuous paraffin sections with a thickness of 5 μm were subsequently prepared. Some sections were used for TUNEL staining, while others were stained with hematoxylin and eosin (H&E). The H&E-stained sections were examined under a light microscope using a double-blind method. Key observations included nuclear morphology (condensation/dissolution), structural integrity of tissues, and signs of autolysis.
TUNEL assay
Paraffin-embedded sections (5 µm) were used for in situ detection of DNA strand breaks in apoptotic cells. Sections were sequentially immersed in xylene I for 15 minutes, xylene II for 15 minutes, absolute ethanol I for 5 minutes, absolute ethanol II for 5 minutes, 85% ethanol for 5 minutes, and 75% ethanol for 5 minutes, followed by a rinse in distilled water to complete the deparaffinization and rehydration process. Subsequent staining was performed according to the manufacturer’s protocol using the in situ Cell Death Detection Kit, Peroxidase (POD) (Roche, Basel, Switzerland). Tissue sections were treated with proteinase K working solution and incubated at 37°C for 30 minutes. Sections were then washed three times with PBS for 5 minutes. Then tissue sections were incubated with permeabilization solution at room temperature for 20 minutes, followed by washing with PBS (pH 7.4) three times for 5 minutes each. A freshly prepared mixture of reagent 1 [Terminal deoxynucleotidyl transferase (TdT)] and reagent 2 (dUTP) in a 2:29 ratio was applied to cover the tissue sections. Slides were incubated in a humidified chamber at 37°C for 2 hours. After three additional PBS washes (5 minutes each), endogenous peroxidase activity was blocked by incubation in 3% hydrogen peroxide (prepared in methanol) for 15 minutes at room temperature in the dark, followed by another set of PBS washes (3 × 5 minutes). Reagent 3 (converter-POD) was then applied, and the slides were incubated at 37°C for 30 minutes, followed by PBS washes as above. 3,3,-Diaminobenzidine (DAB) substrate was added, and color development was monitored under the Olympus IX 71 microscope (Olympus Corporation: Shinjuku-ku, Tokyo, Japan). The reaction was stopped by rinsing with tap water once the nuclei showed brown staining in apoptotic cells. Sections were counterstained with Harris hematoxylin for approximately 3 minutes, differentiated in 1% hydrochloric acid alcohol, blued with ammonia water, and rinsed thoroughly. Finally, the slides were dehydrated through graded alcohols (75%, 85%, absolute ethanol I and II, 6 minutes each), cleared in xylene for 5 minutes, and mounted with neutral resin.
Statistical analysis
All sections derived from five biological replicates per experimental group (specific tissue at a specific PMI) were scanned using a high-resolution digital slide scanner (Leica Aperio AT2, Leica Biosystems, Inc., Germany), generating the raw image data. The images were then analyzed using the Counting function of Image-Pro Plus 6.0 software to quantify TUNEL-positive (apoptotic) cells, following established protocols.20,21 For each tissue sample at each time point, five sections were randomly selected. From each section, five nonoverlapping high-power fields (HPFs; 400× magnification) were randomly chosen for analysis. For each selected HPF, both the number of TUNEL-positive (apoptotic) cells and the total number of recognizable cells were counted.
The percentage of apoptotic cells (PAC) for each tissue sample was calculated using the following formula: PAC % = (Total counted apoptotic cells/total number of recognizable cells) × 100%.
All quantitative data on the PAC were derived from five biological replicates per experimental group (specific tissue at a specific PMI). The normality of PAC data distribution within each group (defined by tissue type and PMI, n = 5 per group) was initially assessed using the Shapiro–Wilk test. Homogeneity of variances among groups being compared (i.e., different time points within a single tissue type, or different tissue types within a single time point) was evaluated using Levene’s test. Due to the relatively small sample size per group and observations of non-normal distribution and/or variance heteroscedasticity for some datasets, nonparametric statistical methods were employed for primary hypothesis testing to ensure robustness.
Two main types of comparisons were performed: (1) Comparison of PAC across different PMI within each tissue type. To investigate the temporal dynamics of apoptosis within each of the eight tissues (brain, fin, heart, gonad, liver, muscle, skin, and spleen), the Kruskal–Wallis H test was performed independently for each tissue. This test compared the median PAC values among the five PMI groups (0, 4, 8, 12, and 16 hours). If the Kruskal–Wallis H test indicated a statistically significant overall difference (p < 0.05), Dunn’s post hoc test with Bonferroni correction for multiple comparisons was subsequently conducted to identify which specific PMI groups differed significantly from each other. (2) Comparison of PAC across different tissue types at each specific PMI. To determine if PAC levels differed among the eight tissue types at each individual PMI (0, 4, 8, 12, and 16 hours), the Kruskal–Wallis H test was performed independently for each of the five time points. This test compared the median PAC values among the different tissue types at that specific PMI. If a significant overall difference was found (p < 0.05), Dunn’s post hoc test with Bonferroni correction was used for pairwise comparisons between tissue types.
Descriptive statistics, including mean, standard deviation (SD), and median for PAC in each group, were calculated and are presented in Table 1. All statistical analyses were performed using R software (R Version 4.5.0) utilizing packages including dplyr for data manipulation, rstatix for Dunn’s test, and Kruskal–Wallis test execution, rcompanion for generating compact letter displays for significance notation, and ggplot2 for data visualization. 22 A p-value < 0.05 was considered statistically significant for all tests.
Descriptive Statistics for the Percentage of Apoptotic Cells in Various Tissues of Yangtze Sturgeon (Acipenser dabryanus) at Different Postmortem Intervals
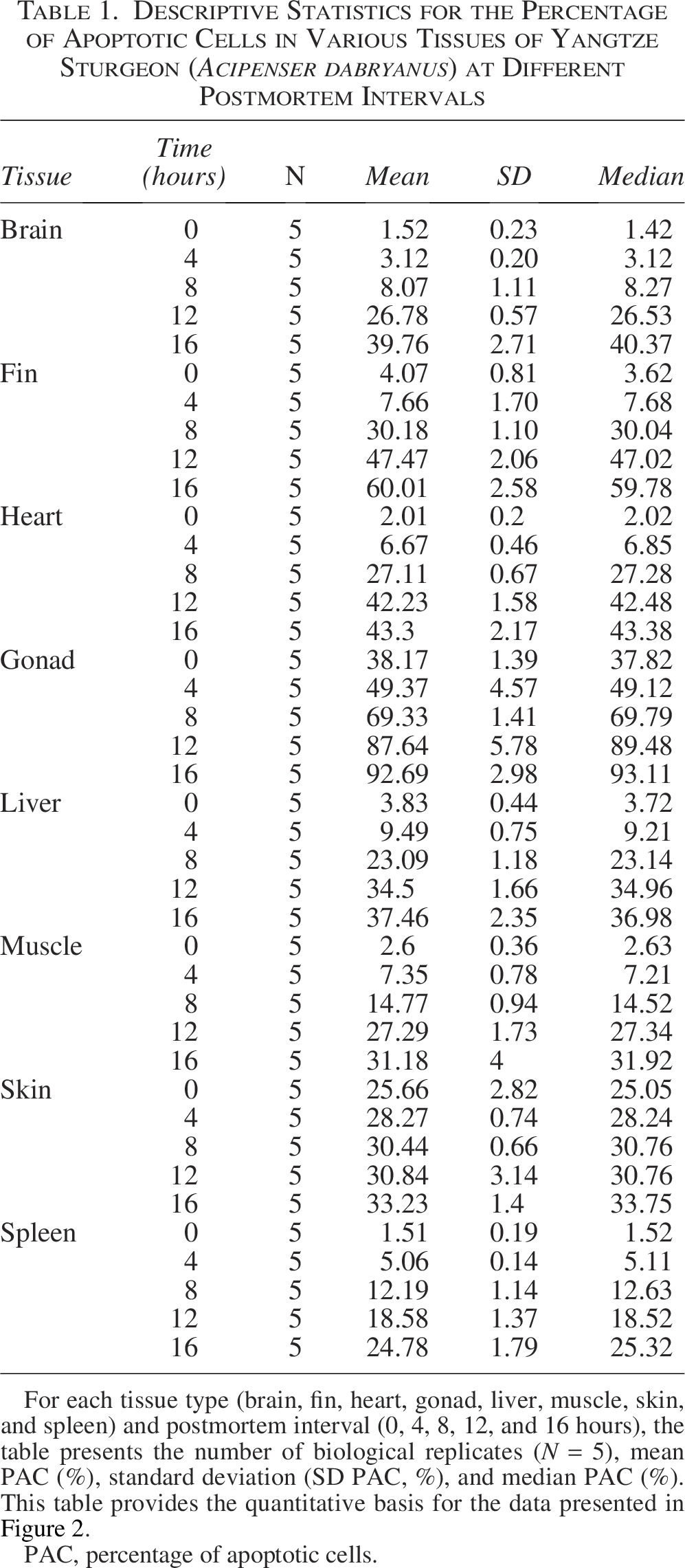
For each tissue type (brain, fin, heart, gonad, liver, muscle, skin, and spleen) and postmortem interval (0, 4, 8, 12, and 16 hours), the table presents the number of biological replicates (N = 5), mean PAC (%), standard deviation (SD PAC, %), and median PAC (%). This table provides the quantitative basis for the data presented in Figure 2.
PAC, percentage of apoptotic cells.
Results
Apoptosis in various tissue cells of Yangtze sturgeon during the early postmortem period
During the early PMI, no significant structural changes were observed in the eight tissues compared to the 0-hour time point, except for a slight reduction in mucus secretion in the fin and skin (Table 2). Histological examination and in situ analysis of apoptotic cells in Yangtze sturgeon revealed that apoptotic nuclei were stained brownish-yellow or brown, with irregular nuclear morphology and variable sizes, and that the apoptotic cells were more abundant at the edges of tissue than in the central regions (Fig. 1A1–H1, A2–H2). Compared with other tissues, the highest density of apoptotic cells at 4 hours postmortem was observed in the gonadal tissues (49.37 ± 4.57%), followed by the skin tissues (28.27 ± 0.74%), while the apoptotic cell levels in other tissues showed a relatively low, generally not exceeding 10% (Table 1). As the postmortem interval extended, the rate gradually decreased. In the skin tissue, the PAC remained relatively stable throughout the entire PMI range, fluctuating between 10% and 20%. In the early postmortem period (0–8 hours), there were no statistically significant differences in apoptotic rates at different time points within the same tissue (Fig. 2A).
Summary of Morphological Changes Observed in Different Tissues of the Yangtze Sturgeon (Acipenser dabryanus) Over a 16-Hour Postmortem Period
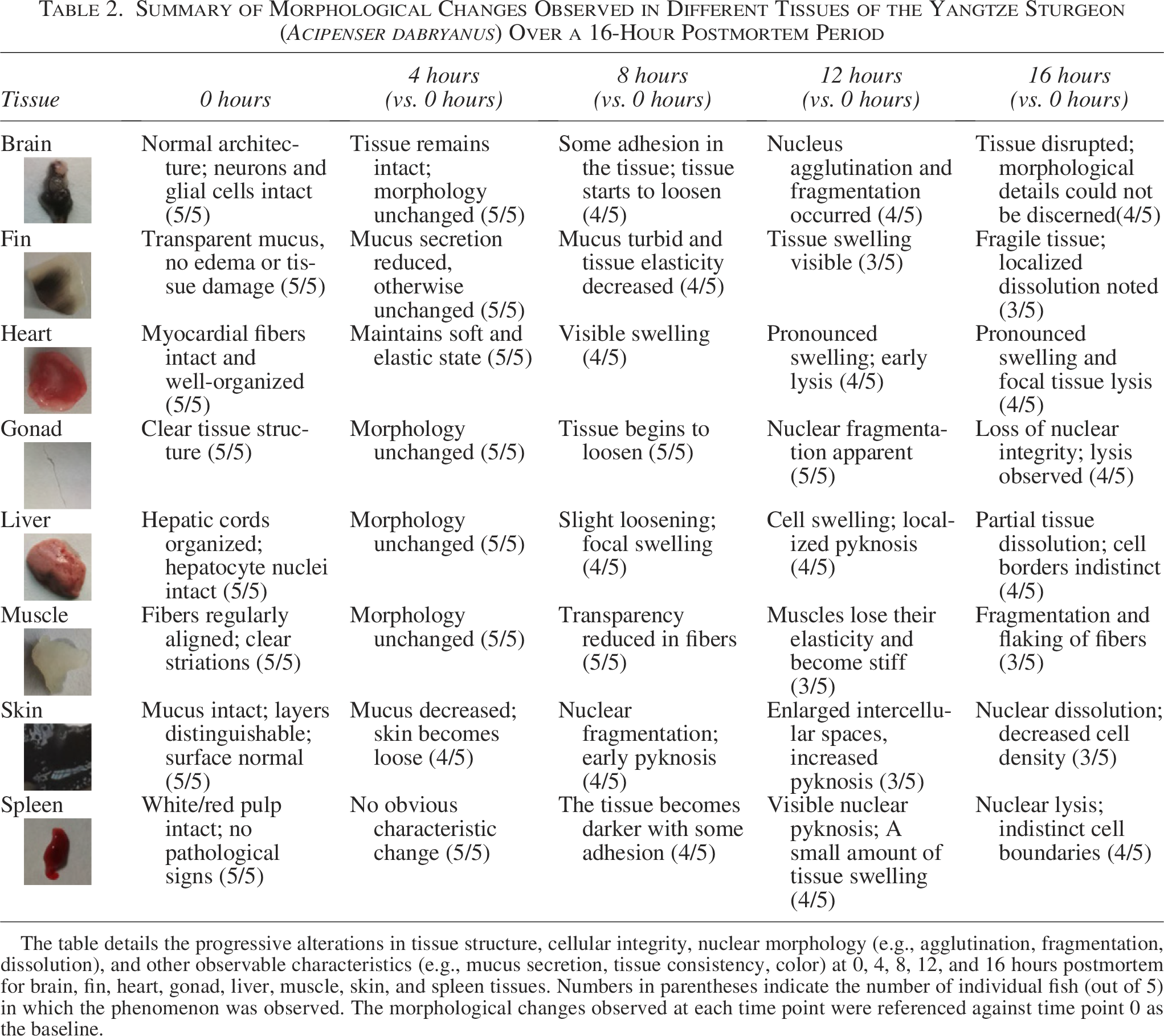
The table details the progressive alterations in tissue structure, cellular integrity, nuclear morphology (e.g., agglutination, fragmentation, dissolution), and other observable characteristics (e.g., mucus secretion, tissue consistency, color) at 0, 4, 8, 12, and 16 hours postmortem for brain, fin, heart, gonad, liver, muscle, skin, and spleen tissues. Numbers in parentheses indicate the number of individual fish (out of 5) in which the phenomenon was observed. The morphological changes observed at each time point were referenced against time point 0 as the baseline.
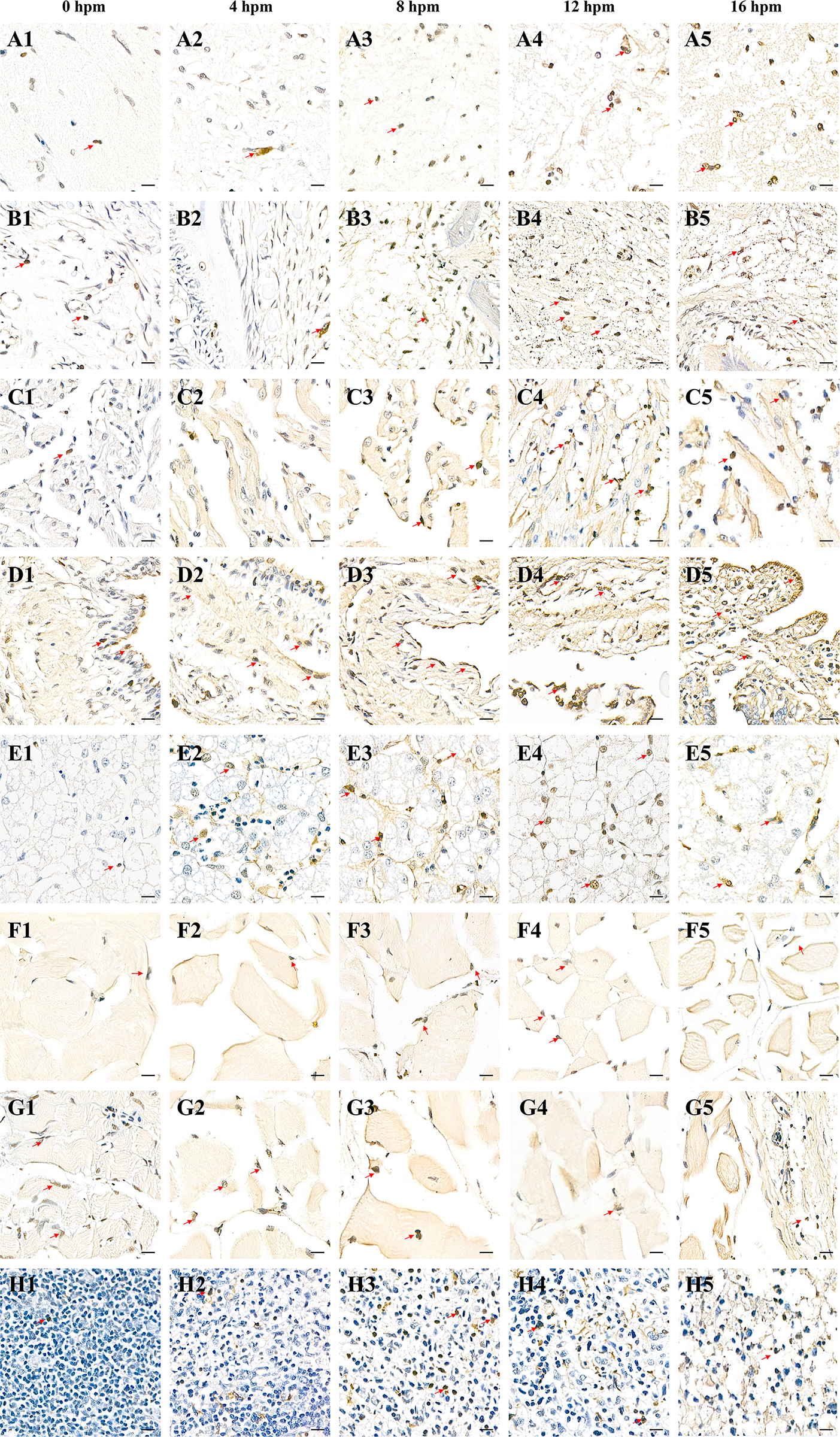
Representative photomicrographs of histomorphological changes and TUNEL-positive cells in various tissues of Yangtze sturgeon (Acipenser dabryanus) at different postmortem intervals (PMI). Rows
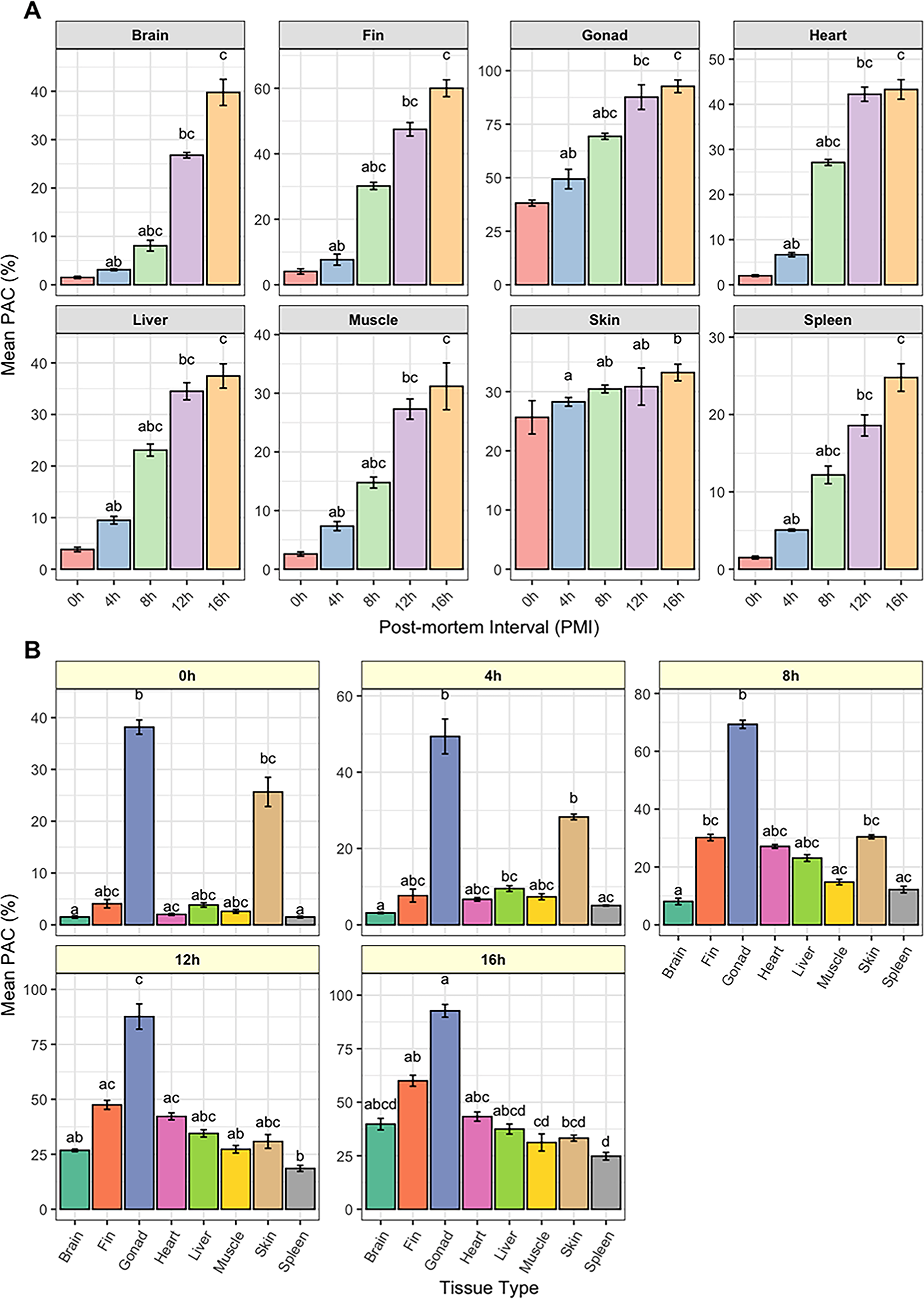
Dynamics of apoptotic cell percentage (PAC) in various tissues of the Yangtze sturgeon (Acipenser dabryanus) across postmortem intervals (PMIs).
Apoptosis in various tissue cells of Yangtze sturgeon during the medium postmortem period
As the PMI extended from 8 to 12 hours, all tissues exhibited significant morphological changes (Table 2). The mucus on the skin and fin tissues lost their previous elasticity and became rigid. Internal tissues, including the heart, spleen, gland, liver, brain, and muscle, began to exhibit mild nuclear pyknosis, visible nuclear fragmentation, and occasional nuclear dissolution (Fig. 1A3–H3, A4-H4).
A significant increase in PAC was observed across most tissues, except for the skin (Fig. 2A). At 8 hours postmortem, the average PAC showed a marked rise compared to the 4-hour time point (Table 1). Specifically, gonad tissue exhibited an increase from 69.33 ± 1.41% at 8 hours to 87.64 ± 5.78% at 12 hours, heart tissue rose from 27.11 ± 0.67% to 42.23 ± 1.58%, brain tissue rose from 8.07 ± 1.11% to 26.78 ± 0.57%, and fin tissue increased from 30.18 ± 1.10% to 47.47 ± 2.06%. These results underscore a marked elevation in apoptotic activity during this interval. Notably, the PAC demonstrated a continuous upward trajectory, approaching or even surpassing a 50% apoptosis rate. Compared with other tissues, the apoptotic rates in liver, muscle, skin, and spleen exhibited only a relatively modest upward trend. For instance, liver tissue showed an increase from 23.09 ± 1.18% to 34.5 ± 1.66%, muscle tissue from 14.77 ± 0.94% to 27.29 ± 1.73%, and spleen tissue from 12.19 ± 1.14% to 18.58 ± 1.37%, with the overall increase in these tissues not exceeding approximately 13%. Overall, the increase in PAC across all tissues was statistically significant (p < 0.05) when compared to the early PMI (0 hours), emphasizing the temporal dynamics of cellular apoptosis following death (Fig. 2B).
Apoptosis in various tissue cells of Yangtze sturgeon during the later postmortem period
During the later stages of the postmortem period, significant postmortem changes were observed in all tissues, including nuclear fragmentation, lysis, and cell dissolution. These changes resulted in the fragmentation of tissue structures, with widening intercellular spaces and a noticeable reduction in cell count. The boundaries of cells became less defined, and some cells completely dissolved or disappeared (Fig. 1A5–H5).
Concurrently, a progressive elevation in PAC was detected in all tissues between 12 and 16 hours postmortem, suggesting a continued apoptotic response beyond the early postmortem phase. This increase was particularly pronounced in the brain and fin, where PAC rose by 12.98% and 12.54%, respectively, by 16 hours (Table 1). In contrast, only modest increases were observed in the gonads, liver, and skin. Among these, the spleen exhibited the greatest rise, with PAC increasing from 18.58 ± 1.37% to 24.78 ± 1.79%, while the heart showed the smallest change, with PAC increasing slightly from 42.23 ± 1.58% to 43.30 ± 2.17%. Statistically significant differences in apoptotic rates were noted when compared to the 0-hour postmortem baseline (p < 0.05); however, no significant differences were found between the 12- and 16-hour time points (p > 0.05), as shown in Figure 2.
Discussion
The conservation of germplasm resources is a cornerstone of biological research, especially for the preservation of genetic diversity in rare and endangered species.23,24 Recently, global efforts have intensified to collect, preserve, and utilize germplasm resources, recognizing their pivotal role in breeding and biodiversity conservation.25,26 The postmortem collection of germplasm resources from rare and endangered species has attracted increasing attention due to its potential applications in species restoration and even de-extinction initiatives.27–29
Our study applied the TUNEL assay, a widely recognized technique for detecting DNA fragmentation in situ.30,31 While TUNEL is commonly applied as an indicator of apoptosis, 32 it is important to note that the assay labels DNA strand breaks that may also arise from necrosis, autolysis, or other forms of cell damage.33,34 Therefore, TUNEL-positive signals reflect the presence of DNA fragmentation rather than apoptosis exclusively. To assess postmortem cellular integrity, we used this assay to evaluate DNA fragmentation patterns in eight different tissues of Yangtze sturgeon at five PMIs (0, 4, 8, 12, and 16 hours). Given the assay’s sensitivity but limited specificity, we performed strict optimization and control of experimental conditions, such as fixation time, enzyme concentration, and incubation parameters, to minimize technical variability and reduce potential artifacts in TUNEL staining.
The results of this study demonstrate that the PAC in various tissues of the Yangtze sturgeon increases progressively with the extension of the PMI, indicating a time-dependent apoptotic pattern. Notably, in most tissues, such as the gonads, fins, and heart, the PAC exceeded 40% at 16 hours postmortem. This phenomenon is consistent with previous research, which has demonstrated that as the PMI increases, nuclear DNA undergoes regular degradation within a specific timeframe. 35 Additionally, due to differences in autolysis rates and DNA degradation kinetics among different cell types within the same organism, the proportion of apoptotic cells varies across tissues. In this study, germ cells in gonadal tissues exhibited a higher rate of apoptosis compared to other tissues, which may be related to the fact that the interstitial cells in gonadal tissue are more sensitive to enzymatic action, androgenase stops, or contain specific degrading enzymes to accelerate degradation, etc.36–38 Although the skin displayed a relatively high PAC during the early postmortem stages, its apoptosis level showed limited variation with increasing PMI. This suggests that the physiological response of the skin to postmortem conditions may be relatively stable. By 16 hours postmortem, apoptosis had further increased in all tissues; however, at this stage, the sturgeon’s body showed evident signs of autolysis, and the structural integrity of the tissues was severely compromised.
It is important to note that apoptosis is an active, ATP-dependent process, which rapidly ceases after death due to the shutdown of energy metabolism. Therefore, the dynamic changes in TUNEL signals observed in this study are more likely attributable to passive postmortem DNA degradation and the inherent limitations of the detection method, rather than ongoing, biologically regulated apoptosis. Future work will use fresh tissues to quantify DNA degradation dynamics through multimodal approaches. These will include quantitative analyses of DNA degradation (e.g., gel electrophoresis), systematic assessment of DNA integrity via Quantitative Polymerase Chain Reaction (qPCR) amplification efficiency, and mechanistic studies on the interplay between tissue autolysis (e.g., cathepsin-mediated proteolysis) and nuclease-driven DNA fragmentation.
Our observation of a significant increase in the rate of apoptosis in tissues such as the brain, muscle, liver, and fin over time suggests a time-dependent pattern of cellular degradation. These findings are consistent with previous studies,39–41 suggesting that apoptosis in these tissues may serve as a potential biomarker for estimating early PMIs. In contrast, tissues such as the spleen and heart exhibited a comparatively slower increase in apoptotic rates, possibly reflecting a stronger resistance to postmortem conditions or slower cellular degradation processes. Apoptotic cells in our study were mainly concentrated at the tissue periphery, with fewer detected in central areas. This may result from technical factors related to fixation. Since fixatives penetrate from the outside in, incomplete preservation of nuclear structures in the core may occur, even after 24 hours in freshly prepared 4% paraformaldehyde. Additionally, autolysis or chromatin loss in deeper regions could reduce TUNEL signal detection. These observations underscore the need to optimize tissue size, fixative volume, and fixation time to ensure uniform preservation.
Additionally, our study observed that the onset of rigor mortis occurred between 8 and 12 hours postmortem, with full rigidity observed at 12 hours. Data analysis of apoptotic rates across tissues revealed that most cells were in an advanced state of apoptosis at this critical time point. Although apoptosis continued to increase beyond 16 hours, the tissue structure had already undergone severe degradation by this stage. This suggests that the 12-hour timepoint represents a critical threshold in the intrinsic degradation of tissues. The process of rigor mortis is known to be influenced by environmental factors, particularly ambient temperature, which directly affects the rate of ATP depletion and onset of muscle stiffening. Previous studies have demonstrated that lower postmortem temperatures can significantly delay the onset of rigor mortis and apoptosis, thereby preserving tissue integrity for longer periods. Conversely, higher temperatures accelerate enzymatic and oxidative degradation processes, leading to earlier structural and cellular breakdown.42,43 Given that our study was conducted under room temperature conditions, it is plausible that the observed timing of rigor mortis and increased apoptosis reflects a more rapid postmortem deterioration compared to samples maintained at refrigerated or hypothermic conditions. This raises the important consideration that postmortem preservation strategies, including temperature control, may critically influence the time window available for the viable recovery of germline or stem-like cells.
Autolysis likely commenced sometime between 12 and 16 hours postmortem, potentially contributing to structural alterations in the tissues. This change may also contribute to the limitations of the TUNEL assay in detecting late-stage DNA fragmentation, or it could indicate that the degradation of nuclear material has surpassed the sensitivity threshold of the method. During later stages of death, certain tissues seem to exhibit delayed apoptotic responses. For instance, in cardiac and skin tissues, the delayed apoptosis may be linked to the interaction with other forms of cell death, such as necrosis and autophagy, or associated with changes in the expression of some genes. Further investigation is warranted to explore the tissue-specific regulatory mechanisms involved in postmortem cellular degradation.
Conclusions
In summary, this study provides an important reference for determining the postmortem window suitable for germplasm preservation in the Yangtze sturgeon under ambient conditions of approximately 25°C. Furthermore, our findings offer preliminary evidence that the brain, muscle, liver, and fin may represent optimal tissue types for estimating the PMI in this species. Based on TUNEL assays and histomorphological analysis, we observed that cellular apoptosis and tissue degradation progressively increased over time, with substantial structural compromise occurring beyond 12 hours after death. These findings suggest that within the first 12 hours, tissues may still retain sufficient cellular integrity for potential applications involving viable cell isolation or cryopreservation.
Beyond this period, however, extensive autolysis and loss of tissue architecture may limit their suitability for preservation in the form of living cell resources. At that point, the remaining options may include DNA or RNA extraction for molecular studies, or even preservation as fixed or archival specimens. Therefore, the 12-hour mark should not be interpreted as a rigid threshold, but rather as a preliminary time reference under specific environmental conditions. The optimal preservation method and time window may vary depending on tissue type, species-specific characteristics, and external factors such as temperature and humidity. Further research is needed to establish standardized protocols and to determine the most appropriate preservation forms for different purposes under varying postmortem conditions.
Authors’ Contributions
Conceptualization: J.L. and H.D.; methodology: J.L., H.D., and B.W.; validation: S.J. and Y.C.; formal analysis: J.L.; investigation: H.D. and Y.C.; data curation: Y.C. and S.J.; writing—original draft: J.L. and B.W.; writing—review and editing: J.L. and H.D.; and visualization: B.W. All authors have read and agreed to the published version of the article.
Footnotes
Acknowledgments
The authors wish to thank Xin Zhu and Wei Wang for providing help in the fish breeding, and Xun Zhao for technical assistance during tissue collection and laboratory work.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work was financially supported by the Fundamental Research Funds for the Central Government-Guided Local Science and Technology Development Special Program of Hubei Province (2024CSA072), the Hubei Province Natural Science Foundation Innovative Development Joint Fund Project (2025AFD314), and the Hubei Key Laboratory of Three Gorges Project for Conservation of Fishes (2020).
Compliance with Ethical Standards
The study was carried out by the guidelines and regulations of the National Institute of Health Guide for the Care and Use of Laboratory Animals and was approved by the Institutional Review Board on Bioethics and Biosafety of the Chinese Sturgeon Research Institute.