
Abstract
Aim:
The poultry sector is currently witnessing heavy demand for its products especially meat, which is expected to intensify in the coming years. However, while Japanese quail displays a lot of promise to help meet the soaring demands, its sustainable production requires assisted reproduction via sperm cryopreservation. Hence, the current study was designed to elucidate the impact of cryopreservation on Japanese quail semen quality, antioxidant potential, and mitochondrial activity and intra-species variation in terms of freeze-tolerance.
Materials and Methods:
Semen was collected individually from seven mature males, diluted with NaCl extender and cryopreserved. Samples were analyzed for sperm motility, plasma membrane and acrosomal integrity, viability, DNA fragmentation, and biochemical parameters at the fresh collection, post-dilution, post-cooling, post-equilibration, and post-thaw stages of freezing.
Results:
Sperm motility, plasma membrane and acrosomal integrity, viability, antioxidant potential, scavenging capacity, and mitochondrial activity were reduced (p < 0.05) and DNA fragmentation was increased (p < 0.05) at all the stages of cryopreservation. Further, all the parameters were negatively correlated with DNA fragmentation during cryopreservation. The percent incline rates for DNA fragmentation and decline rates for the rest of the parameters in individual birds showed intra-species variation (p < 0.05) with respect to freeze-tolerance.
Conclusion:
Japanese quail semen quality, antioxidant potential, and mitochondrial activity are severely affected by the freezing process and the level of freeze-resilience varies among individuals.
Introduction
Animal propagation and conservation for the future are topics of global interest as semen freezing has made sperm revival and its utilization possible after prolonged storage. 1 However, cells under chilling temperatures go through multiple shocks that disrupt their normal biological framework. In vivo, spermatozoa are protected against reactive oxygen metabolites with a network of natural antioxidants spread across the sperm and seminal fluid. It operates by three different mechanisms: inhibiting oxidants from reacting with cell components, breaking free radical reactions, and eliminating their products via cellular macromolecules. 2 This protection mechanism is diminished when semen is subjected to in vitro manipulation and reactive oxygen species (ROS) attack the double bonds in the polyunsaturated fatty acids (PUFA) of membranes to produce oxidative tension. Such inconsistencies in the redox pathway ultimately cause metabolic and functional disruption and compromise the sperm quality. 3
Avian sperm in particular contains very little cytoplasm having minute antioxidant quantities and relatively large plasma membrane surface area with lots of PUFAs. 4 When limited quantities of antioxidants available are overwhelmed by the uncontrolled ROS proliferation, PUFAs make the cells highly prone to freeze-thaw impairment. The ROS extracts the hydrogen atoms from membrane PUFAs and propagates the production of lipid alkyl radicals. Subsequently, it reacts with oxygen, giving rise to the lipid peroxyl radical that could remove hydrogen from another fatty acid in a chain reaction. These events not only decrease plasma membrane fluidity, but also cause inactivation of membrane-bound enzymes/antioxidants/receptors and increase its permeability to unwanted ions. 5 Freezing could also stimulate a pre-acrosomal reaction and thus affects its function and sperm viability.
Mitochondrial status is correlated with the functional competencies of the sperm, 1 but the mitochondrion is also reported as the most sensitive organelle against cryopreservation. When the scavengers are depleted in the cell to control ROS, undue oxidation establishes a wide spectrum of pathologies like declined mitochondrial membrane potential (MMP), uncoupling of electron transport chain, causing the adducts formation with mitochondrial proteins, decreased ATP synthesis, increased free radical production, and ultimately low sperm motility, viability and DNA integrity. 6 When the condensed state of nuclear DNA is attacked by ROS, it produces single and double stranded breaks in the DNA, induction of abasic sites, base modifications or DNA crosslinking.4,7 Consequently, abnormally packaged chromatin affects post-fertilization embryo development via reduced conception or embryo cleavage rates.
The world population is on the rise with an estimated figure of 9.8 billion by 2050, and poultry is the most desired meat type among people. 8 Japanese quail has long served as an animal model for research but no attention is given to its propagation as a food source. Besides large scale farming, the small body size of Japanese quail is also suitable for small scale farming. Moreover, experimental research has shown that this bird quickly responds to selection for body weight.9,10 However, Japanese quail need assisted reproduction via sperm cryopreservation for its propagation and establishment of semen banks because it is extremely sensitive to the introduction of new mates, inbreeding, and transportation stress. 11 Therefore, it is essential to not only define the changes in frozen sperm but also any intra-species variation to develop effective semen preservation protocols and improve sperm quality. To our knowledge, no studies have been performed on avian species for individual variation with regard to freeze-tolerance, so the current study investigated the extent of decline in semen quality, antioxidant activity, and mitochondrial status during cryopreservation and intra-species variation in terms of freeze-tolerance.
Materials and Methods
For this study, all chemicals were purchased from Sigma-Aldrich, Co., 3050 Spruce Street, St Louis, USA.
Ethical statement
The Ethics Committee of Pir Mehr Ali Shah Arid Agriculture University Rawalpindi, Pakistan, approved the study for the use of birds.
Management of experimental birds
Seven mature male Japanese quail (age: 1 year; weight: 250 g) were purchased from local breeders and managed in the Avian Research Station, Pir Mehr Ali Shah-Arid Agriculture University, Rawalpindi, Pakistan. The experimental birds were kept in sand floor pens (length: 182.88 cm; width: 91.44 cm; height: 45.72 cm) and offered commercially available quail breeder feed (approximately 30 g/bird/day). Fresh water was provided ad libitum and the photoperiod was maintained at 16 hours light.
Experimental design
Semen was collected from seven mature males twice a week (April 2022 to May 2022) and processed individually. Samples were evaluated at fresh, post-dilution (37°C), post-cooling (20°C), post-equilibration (4°C), and post-thaw stages of freezing. All sperm quality parameters (motility, plasma membrane and acrosome integrity, viability, DNA integrity) were assessed simultaneously with each individually collected ejaculate from each bird (7 birds × 4 replicates = 28 ejaculates). Each of the three biochemical parameters (mitochondrial status, total antioxidant potential [TAP], free radical scavenging capacity) were assessed with separate semen collection from each bird (7 birds × 3 biochemical parameters × 4 replicates = 84 ejaculates) and processed for the freezing stages. Hence, 112 ejaculates were collected in total for all the parameters evaluated in the current study. This study design was followed due to the limitation of small semen volume (40–70 µL) collected as a single ejaculate from each Japanese quail.
Semen collection and initial evaluation
Semen was collected via the manual massage method 12 in a graduated plastic vial in the morning hours (06:00 AM) and semen volume was measured via micropipette. Initial sperm motility of each ejaculate was obtained by mixing 1 µL fresh semen in 50 µL of phosphate-buffered saline (PBS) (pH 7.2, 300 mOsm/kg) and placing a drop of prepared mixture on a pre-warmed glass slide (37°C) under a phase contrast microscope (×400, Olympus BX20, Japan). Ejaculates with initial motility >80% were processed further and sperm concentration was measured by Neubauer haemocytometer (Marienfeld, Germany) under a phase contrast microscope (×400, Olympus BX20, Japan).
Extender preparation and freezing
The NaCl extender (pH 8.0; Osmolarity 410 mOsm/kg) was prepared by mixing 35 mM glucose, 20 mM N-tris(hydroxymethyl)methyl-2-aminoethanesulphonic acid, and 150 mM sodium chloride in double distilled water (100 mL). 13 From each ejaculate, semen required for the fresh assessment was separated and rest of the sample was diluted with NaCl extender (1:15) for the freezing process. The extended semen (37°C) was cooled to 20°C in 10 minutes (1.70°C/min) and then cooled to 4°C in 15 minutes (1.06°C/min). Cryoprotectant dimethylacetamide (1%) was added in the sample and equilibrated for 4 minutes. Samples were filled in 0.25 mL French straws (IMV, L’Aigle, France), placed over liquid nitrogen vapors (5 cm above LN2) for 4 minutes, and immersed into LN2 (−196°C) container. The straws were thawed at 25°C for 30 seconds in water bath for semen analysis.
Semen quality assays
Sperm motility
Sperm motility was evaluated by placing a drop of the semen sample on a glass slide (37°C) and observing it under a phase contrast microscope (400X). The percentage of motile sperm was evaluated on a scale ranging from 0% to 100%. 14
Sperm plasma membrane integrity
Sperm plasma membrane integrity was assessed through the hypo-osmotic swelling (HOS) test. 15 The HOS solution was prepared by adding 1 g of sodium citrate to distilled water and the volume was raised to 100 mL. Semen samples (2.5 µL) were mixed with 50 μL of an HOS solution (100 mosm/kg) and incubated at 25°C for 30 minutes. A drop of incubated sample was placed on a pre-warmed slide (37°C) and fixed in 2% glutaraldehyde. Spermatozoa having swollen heads and coiled/swollen tails were categorized as normal with intact plasma membranes. A total of 200 spermatozoa were studied at four separate fields under a phase contrast microscope (1000X with oil immersion).
Sperm viability
Sperm viability was examined via Lake glutamate solution that was prepared by fortifying sodium glutamate (0.01735 g), potassium citrate (0.00128 g), sodium acetate (0.0085 g), and magnesium chloride (0.000676 g) in 100 mL of distilled water. Water-soluble nigrosin (5 g) and water-soluble eosin-bluish (1 g) were mixed with Lake glutamate solution. One drop of semen was added to 12 drops of stain and mixed. A smear was prepared on a glass slide, fixed and air dried. The smear provided a clear background and contrast between the unstained live sperm and the darkly pink-stained dead sperm. A total of 200 spermatozoa were counted on a slide under a phase contrast microscope (Olympus BX20, Japan; 1000×, oil immersion).
Sperm acrosomal integrity
Sperm acrosomal integrity was evaluated through Giemsa stain 16 that was prepared by adding 3 g of giemsa and 2 mL of PBS (pH 7.0) into 35 mL of water. Smears of the semen were made on a pre-warmed glass slides, air dried, fixed in neutral formal-saline (5% formaldehyde) for 30 minutes, and kept in giemsa stain for 1.5 hours. Sperm with normal acrosomes were evenly and darkly stained while sperm with abnormal or ruptured acrosomes were unevenly stained or unstained. A total of 200 sperm were studied in at least four different areas under a phase-contrast microscope (Olympus BX20, Japan; 1000X, oil immersion).
Sperm DNA fragmentation
Sperm DNA fragmentation was determined via aniline blue stain. 14 The staining solution was prepared by adding aniline blue (5 g) in 100 mL of PBS, filtered, and pH adjusted to 3.5 with the addition of 2% glacial acetic acid. Semen (3 µL) was spread on a glass slide and air dried. Slides were dipped into aniline blue stain for 5 minutes, flushed with distilled water, and air dried. A count of 200 sperm was taken using a phase contrast microscope (1000X, oil immersion). The spermatozoa stained dark blue had fragmented DNA and the unstained sperm had intact DNA.
Biochemical assays
Free radical scavenging capacity
Free radical scavenging activity was evaluated via DPPH assay (2,2-diphenyl-1-picrylhydrazyl).
3
Sperm pellet obtained after centrifugation (10 minutes; 1500 ×g) was washed with PBS (pH 7.4 at 25°C; Sigma-Aldrich, USA) and centrifuged again (1500 ×g) for 10 minutes. Then sperm pellets were diluted with PBS to obtain a final concentration of 50 million sperm/mL. A 40 µL of sperm suspension was added to 560 µL of DPPH working solution (3.2 mg DPPH [Sigma-Aldrich, USA] in 100 mL of 80% methanol [BDH, UK]) and given vigorous shaking followed by 1 hour of incubation. DPPH solution (600 µL) was taken as a control and a blank was prepared by mixing 560 µL of methanol in 40 µL of diluted sample. Then, absorbance was taken in a spectrophotometer (537 nm) and free radical scavenging capacity calculated:
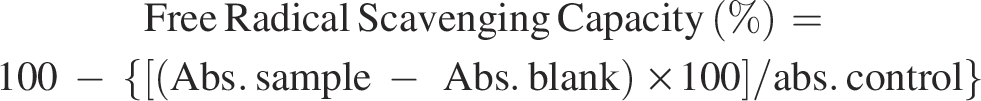
Total antioxidant potential
The ferric reducing antioxidant power (FRAP) assay was used to measure the total antioxidant potential of the semen. Working FRAP solution was prepared by mixing acetate buffer (3.1 g sodium acetate trihydrate [Merck, Germany], 16 mL glacial acetic acid [Merck, Germany] and increased to 1 L with distilled water), 2,3,5-triphenyltetrazolium chloride (TPTZ) (0.031 g TPTZ [Sigma-Aldrich, USA] per mL of 40 mM HCl [Merck, Germany]), and ferric chloride (Merck, Germany) (0.054 g in 10 mL distilled water) in the ratio 10:1:1. For the standard, 50 µL of standard ferrous sulphate was added and dissolved in 1 mL of FRAP reagent. Semen samples (20 μL) were mixed with FRAP reagent (400 μL) and readings of standard and samples were obtained in a spectrophotometer (593 nm). The FRAP value for the samples were calculated as follows:
FRAP value of semen (μmol/mL) = (Absorbance of sample/Absorbance of standard) × FRAP value of standard
Activity of mitochondrial dehydrogenases
The MTT [3-(4, 5-dimethyl-thiazol-2-yl)−2, 5-diphenyltetrazolium bromide] reduction assay 3 was employed to evaluate the activity of mitochondrial dehydrogenases in sperm cells. A water-soluble yellow dye (MTT) was absorbed by the viable cells and reduced to water-insoluble blue formazan via mitochondrial dehydrogenases. The MTT stock solution was prepared by mixing 5 mg MTT (Sigma-Aldrich) per mL of PBS. Then, MTT stock solution (2.5 µL) was added to the semen sample (25 µL) and the absorbance value was recorded immediately and after 1 hour of the incubation at 37°C in a spectrophotometer (550 nm). Later, the rate of MTT reduction was calculated by the difference between the initial and final readings.
Statistical analysis
Data for all the parameters are presented as mean ± SEM. The data for the stages of cryopreservation and individuals were analyzed via two-factor factorial ANOVA in MSTAT-C® software (version 1.42, Michigan State University, East Lansing, MI, USA) with completely randomized design. In addition, overall percent decline and incline rates of the parameters were calculated in individual birds from the fresh stage to post-thaw and one-way ANOVA was performed in MSTAT-C®. The collective mean and SEM of all birds were also calculated for each parameter at each stage and one way ANOVA was applied. When the F-ratio was obtained significant (p < 0.05), Fisher’s protected least square difference test was applied to compare means. Pearson’s correlation was performed with the SPSS statistical package (SPSS 16.0 for Windows; SPSS, Inc, Chicago, IL) on all the semen parameters studied at the fresh stage and during cryopreservation.
Results
Effect of cryo-storage on sperm motility
The data on the effect of stages of cryopreservation on sperm motility in individual Japanese quail (n = 7) are presented in Figure 1. In the fresh semen, the highest (p > 0.05) sperm motility was recorded in bird 4 (91.25 ± 2.39) followed by bird 7, 6, 1, 5, 3, and 2. At post-dilution, bird 4 (87.50 ± 1.44) and bird 6 (87.50 ± 1.44) maintained higher (p > 0.05) sperm motility compared with other individuals. At post-cooling, higher (p > 0.05) sperm motility was recorded in bird 1 (68.75 ± 1.25), bird 6 (68.75 ± 1.25) and bird 4 (67.50 ± 1.44) compared with other individuals. At post-equilibration, birds 4, 6, 1, and 3 maintained higher (p > 0.05) sperm motility (58.75 ± 1.25, 57.50 ± 2.50, 57.50 ± 1.44, and 57.50 ± 1.44) compared with birds 5, 2, and 7. At post-thaw, bird 4 maintained the highest (p < 0.05) sperm motility (25.00 ± 0.00) compared to bird 1 (21.25 ± 1.25), bird 3 (18.75 ± 1.25), bird 6 (17.50 ± 2.50), bird 5 (16.25 ± 2.39), bird 2 (12.50 ± 1.44) and bird 7 (7.50 ± 1.44). Overall, bird 4 presented the lowest (p < 0.05) and bird 7 presented the highest (p < 0.05) percent decline rate for motility compared with other individuals during cryopreservation (Table 1). Moreover, a significant decrease (p < 0.05) in sperm motility was observed with the combined mean of all birds at each successive stage (Fresh: 89.82 ± 0.57; Post-dilution: 86.25 ± 0.38; Post-cool: 66.60 ± 0.70; Post-equilibration: 56.42 ± 0.79; Post-thaw: 16.96 ± 2.16).
Effect of stages of cryopreservation (fresh, post-dilution, post-cooling, post-equilibration, and post-thaw) on sperm motility (%) of individual Japanese quail (n = 7). Different superscripts indicate statistically significant differences (p < 0.05).
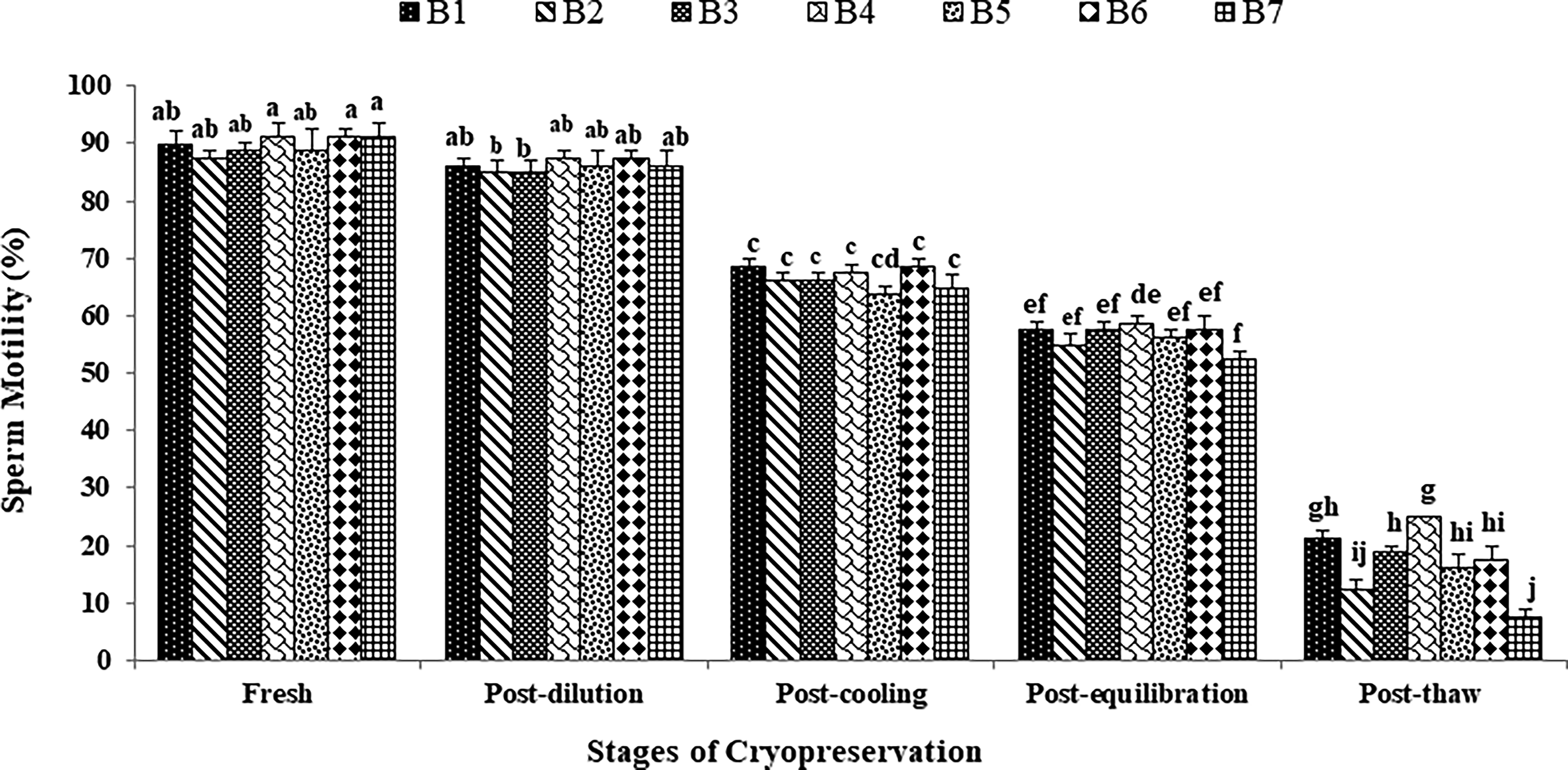
Overall Decline and Incline Rate (%) of Japanese Quail (n = 7) Sperm Quality, Antioxidant Activity, and Mitochondrial Status from Fresh Collection to Post-Thaw Stage of Cryopreservation
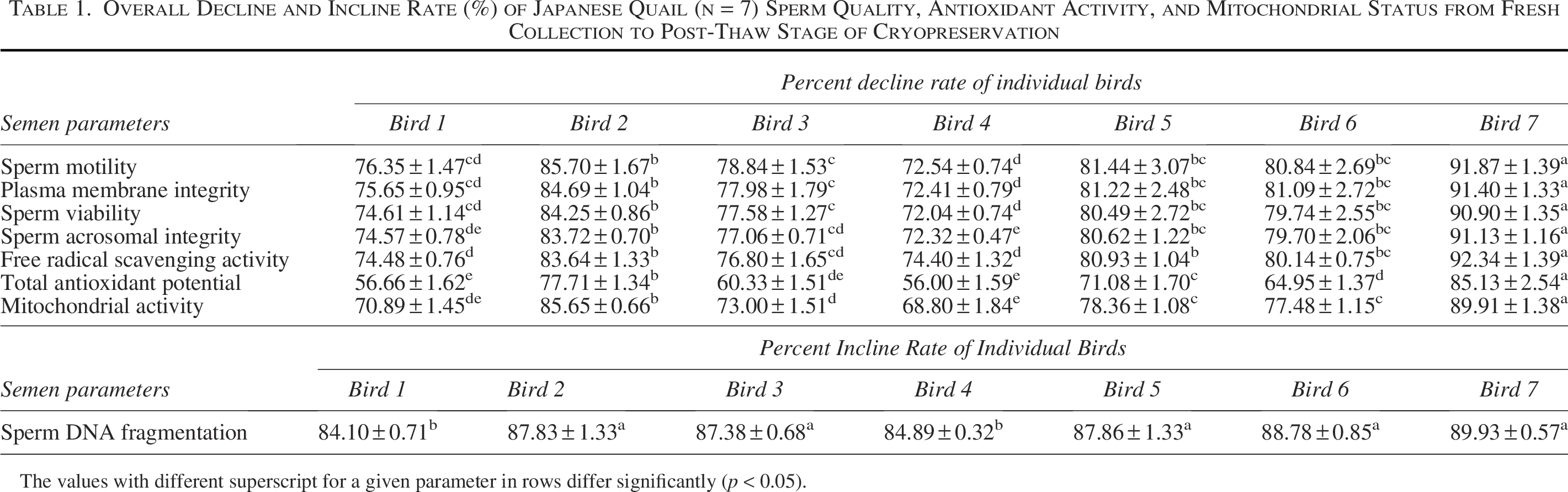
The values with different superscript for a given parameter in rows differ significantly (p < 0.05).
Effect of cryo-storage on sperm plasma membrane integrity
The data on the effect of stages of cryopreservation on sperm plasma membrane integrity in individual Japanese quail (n = 7) are presented in Figure 2. In the fresh semen, the highest (p > 0.05) sperm membrane integrity was recorded in bird 4 (95.25 ± 1.44) followed by bird 7, 6, 1, 5, 3, and 2. At post-dilution, bird 4 (88.75 ± 1.93) and bird 6 (88.75 ± 1.49) maintained higher (p > 0.05) sperm membrane integrity compared with other individuals. At post-cooling, bird 6 (70.00 ± 1.41), bird 1 (69.50 ± 1.26), and bird 4 (68.25 ± 1.60) presented higher (p > 0.05) plasma membrane integrity whereas at post-equilibration, bird 4, 6, 3, and 1 maintained higher (p < 0.05) sperm membrane integrity (59.75 ± 0.95, 58.75 ± 2.29, 58.50 ± 1.55, and 58.50 ± 1.76 respectively) compared with other individuals. At post-thaw, bird 4 maintained the highest (p < 0.05) sperm membrane integrity (26.25 ± 0.48) compared with bird 1 (22.75 ± 0.85), bird 3 (20.25 ± 1.44), bird 6 (17.75 ± 2.59), bird 5 (17.25 ± 2.06), bird 2 (14.00 ± 0.91), and bird 7 (8.25 ± 1.38). Overall, bird 4 presented the lowest (p < 0.05) and bird 7 presented the highest (p < 0.05) percent decline rate for membrane integrity compared with other individuals during cryopreservation (Table 1). Moreover, a significant decrease (p < 0.05) in sperm membrane integrity was observed with the combined mean of all birds at each successive stage (Fresh: 93.42 ± 0.55; Post-dilution: 87.25 ± 0.45; Post-cool: 67.35 ± 0.76; Post-equilibration: 57.21 ± 0.90; Post-thaw: 18.07 ± 2.22).
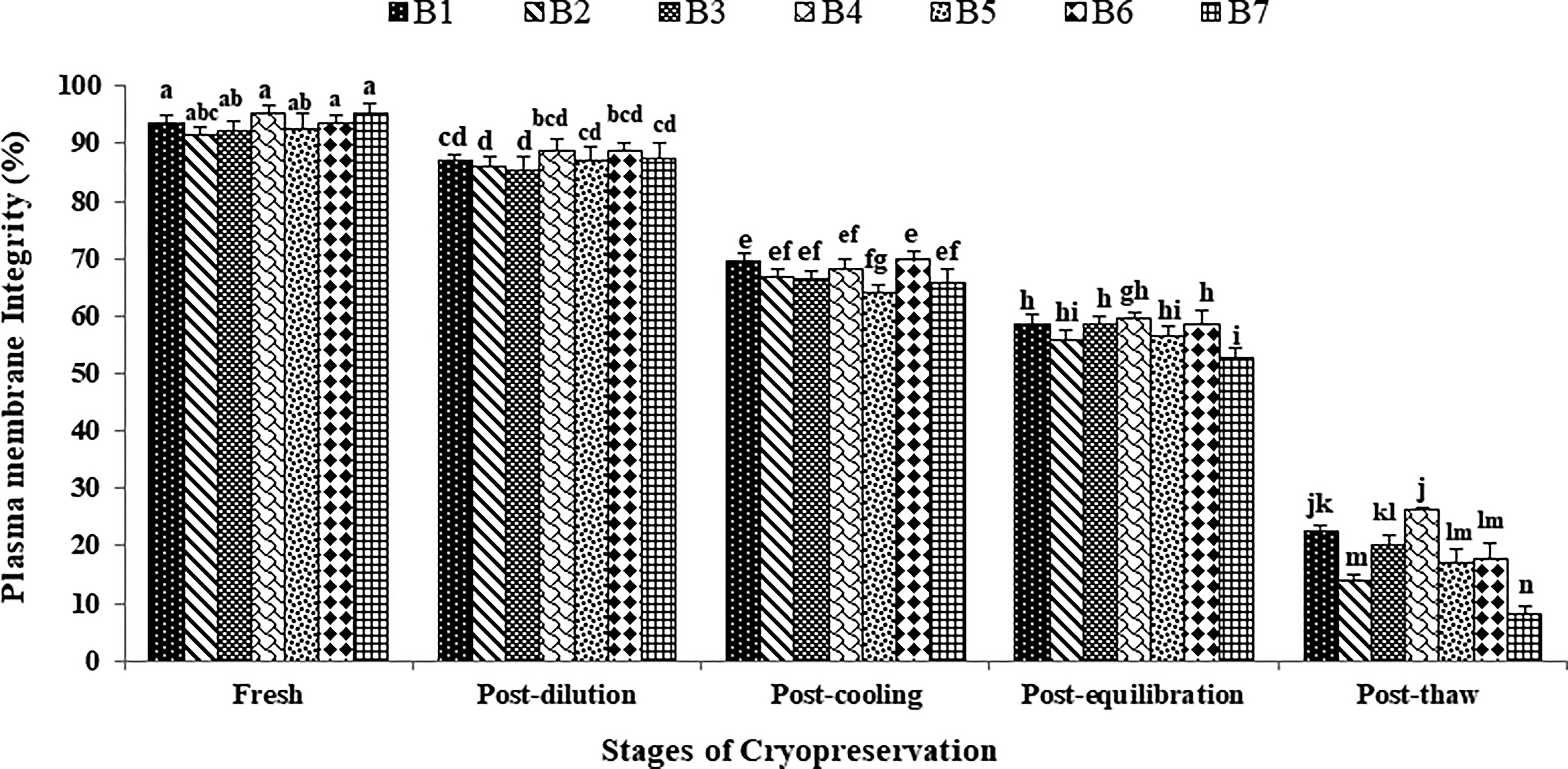
Effect of stages of cryopreservation (fresh, post-dilution, post-cooling, post-equilibration, and post-thaw) on sperm plasma membrane integrity (%) of individual Japanese quail (n = 7). Different superscripts indicate statistically significant differences (p < 0.05).
Effect of cryo-storage on sperm viability
The data on the effect of stages of cryopreservation on sperm viability in individual Japanese quail (n = 7) are presented in Figure 3. In the fresh semen, the highest (p > 0.05) sperm viability was recorded in bird 4 (95.75 ± 1.31), followed by bird 7, 6, 1, 5, 3, and 2. At post-dilution, bird 4 (89.25 ± 1.65) and bird 6 (89.00 ± 1.29) maintained higher (p > 0.05) sperm viability followed by bird 7, 5, 1, 2, and 3. At post-cooling, bird 6 (71.00 ± 1.35), bird 1 (70.25 ± 1.18), and bird 4 (69.00 ± 1.47) presented higher (p < 0.05) sperm viability whereas at post-equilibration, bird 4, 6, 1, and 3 maintained higher (p < 0.05) sperm viability (60.25 ± 1.11, 59.50 ± 2.53, 59.25 ± 1.60 and 58.75 ± 3.30) compared with other individuals. At post-thaw, bird 4 maintained the highest (p < 0.05) sperm viability (26.75 ± 0.63) compared with bird 1 (23.75 ± 0.75), bird 3 (20.75 ± 0.95), bird 6 (18.75 ± 2.29), bird 5 (18.00 ± 2.20), bird 2 (14.50 ± 0.65), and bird 7 (8.75 ± 1.38). Overall, bird 4 presented the lowest (p < 0.05) and bird 7 presented the highest (p < 0.05) percent decline rate for sperm viability compared with other individuals during cryopreservation (Table 1). Moreover, a significant decrease (p < 0.05) in sperm viability was observed with the combined mean of all birds at each successive stage (Fresh: 93.85 ± 0.52; Post-dilution: 87.75 ± 0.43; Post-cool: 67.89 ± 0.85; Post-equilibration: 57.75 ± 0.90; Post-thaw: 18.75 ± 2.24).
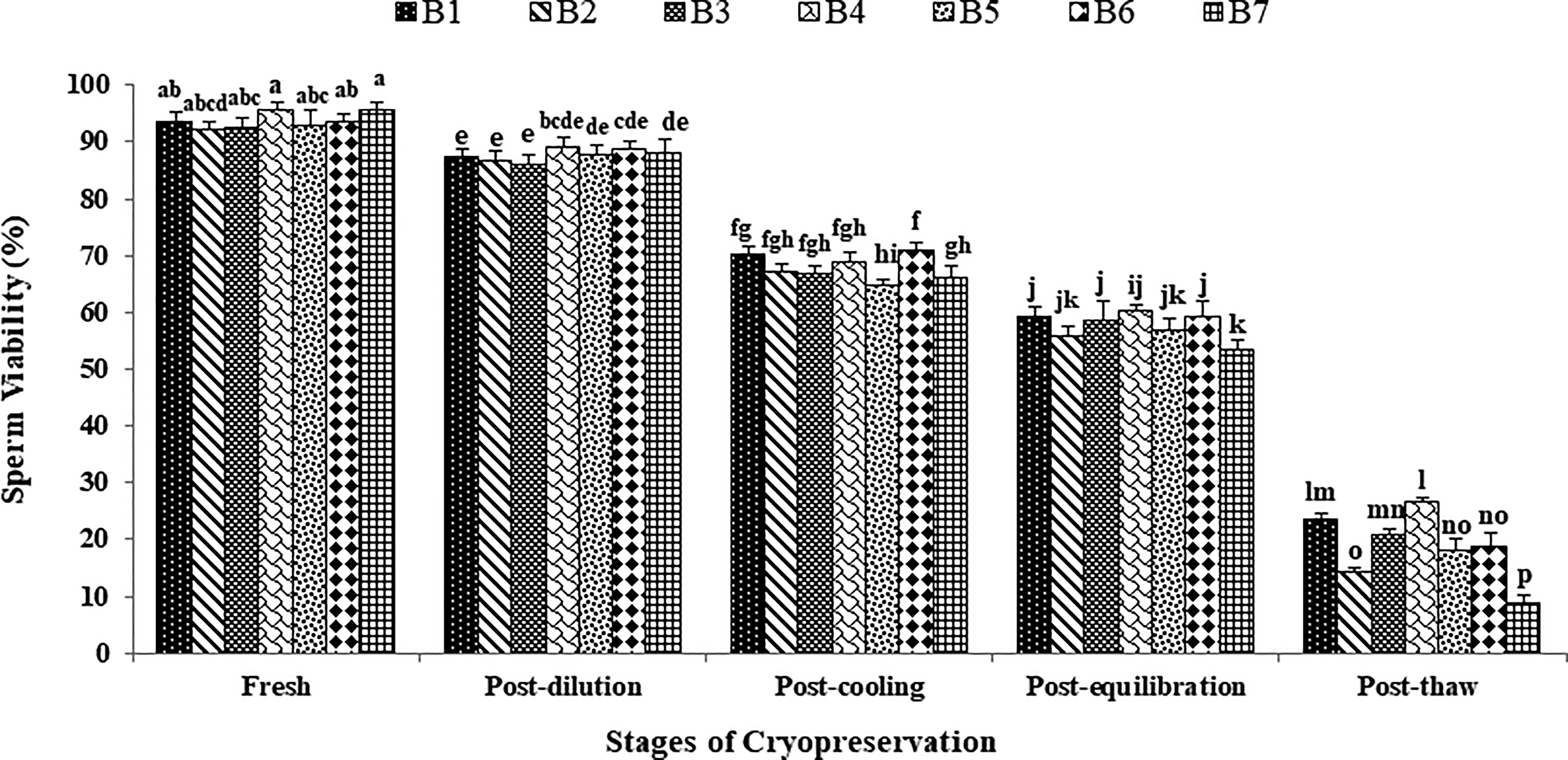
Effect of stages of cryopreservation (fresh, post-dilution, post-cooling, post-equilibration, and post-thaw) on sperm viability (%) of individual Japanese quail (n = 7). Different superscripts indicate statistically significant differences (p < 0.05).
Effect of cryo-storage on sperm acrosomal integrity
The data on the effect of stages of cryopreservation on sperm acrosomal integrity in the individual Japanese quail (n = 7) are presented in Figure 4. In the fresh semen, bird 4 presented the highest (p > 0.05) sperm acrosomal integrity (95.75 ± 0.95) followed by bird 7, 6, 1, 5, 3, and 2. At post-dilution, bird 4 (89.50 ± 1.76) and bird 6 (89.50 ± 1.19) maintained higher (p > 0.05) sperm acrosomal integrity, whereas at post-cooling, bird 6 (71.50 ± 1.19), bird 1 (70.75 ± 1.31), and bird 4 (69.50 ± 1.19) presented higher (p < 0.05) sperm acrosomal integrity compared with other individuals. At post-equilibration, birds 4, 6, 3, and 1 maintained higher (p < 0.05) values (60.75 ± 0.95, 59.75 ± 2.59, 59.00 ± 1.47, and 59.75 ± 1.60) compared with birds 5, 2, and 7. At post-thaw, birds 4 and 1 maintained the highest (p < 0.05) sperm acrosomal integrity (26.50 ± 0.50 and 23.75 ± 0.48) compared with bird 3 (21.25 ± 0.48), bird 6 (19.00 ± 2.00), bird 5 (18.00 ± 0.82), bird 2 (15.00 ± 0.58), and bird 7 (8.50 ± 1.19). Overall, birds 4 and 1 presented the lowest (p < 0.05) and bird 7 presented the highest (p < 0.05) percent decline rate for acrosomal integrity compared with other individuals during cryopreservation (Table 1). Moreover, a significant decrease (p < 0.05) in sperm acrosomal integrity was observed with the combined mean of all birds at each successive stage (Fresh: 93.50 ± 0.49; Post-dilution: 88.08 ± 0.55; Post-cool: 68.58 ± 1.01; Post-equilibration: 58.87 ± 0.64; Post-thaw: 20.58 ± 1.69).
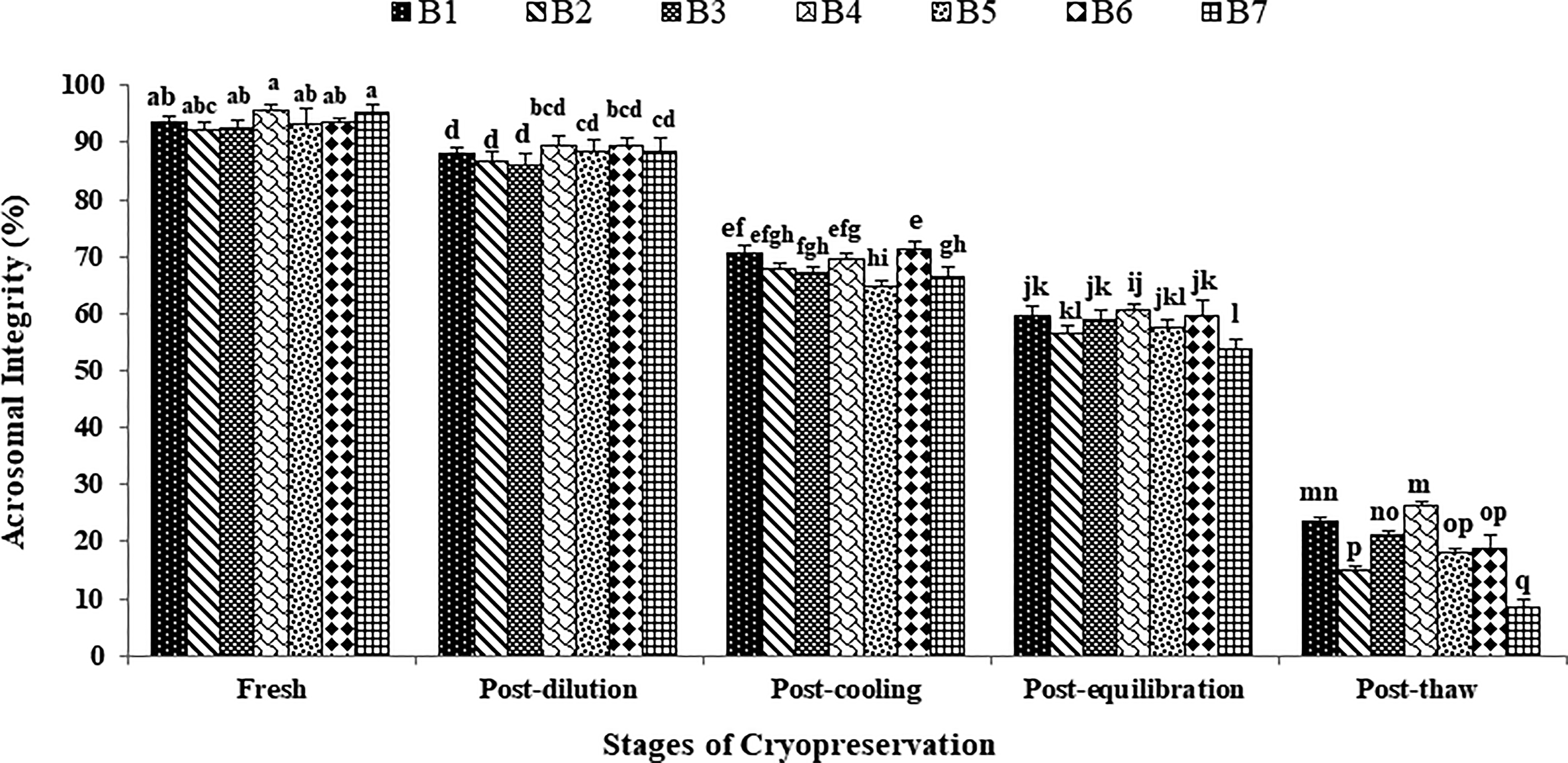
Effect of stages of cryopreservation (fresh, post-dilution, post-cooling, post-equilibration, and post-thaw) on sperm acrosomal integrity (%) of individual Japanese quail (n = 7). Different superscripts indicate statistically significant differences (p < 0.05).
Effect of cryo-storage on sperm DNA fragmentation
The data on the effect of stages of cryopreservation on sperm DNA fragmentation in individual Japanese quail (n = 7) are presented in Figure 5. In the fresh semen, bird 4 presented the lowest (p > 0.05) sperm DNA fragmentation (8.50 ± 0.29) followed by birds 7, 6, 1, 5, 3, and 2. At post-dilution and post-cooling, birds 4, 6, and 1 maintained lower (p > 0.05) sperm DNA fragmentation compared with other individuals. At post-equilibration, birds 4, 6, 3, and 1 maintained lower (p < 0.05) sperm DNA fragmentation (39.00 ± 0.82, 39.25 ± 0.95, 39.75 ± 1.03, and 40.00 ± 0.91 respectively) compared with birds 5, 2, and 7. At post-thaw, birds 4 and 1 maintained the lowest (p < 0.05) sperm DNA fragmentation (76.50 ± 1.19 and 78.00 ± 0.91) compared with birds 3 (79.25 ± 0.25), bird 6 (80.25 ± 0.95), bird 5 (82.75 ± 1.11), bird 2 (84.25 ± 0.25), and bird 7 (87.00 ± 0.71). Overall, birds 4 and 1 presented the lowest (p < 0.05) and bird 7 presented the highest (p < 0.05) percent incline rate for DNA fragmentation compared with other individuals during cryopreservation (Table 1). Moreover, a significant increase (p < 0.05) in sperm DNA fragmentation was observed with the combined mean of all birds at each successive stage (Fresh: 9.39 ± 0.26; Post-dilution: 13.60 ± 0.38; Post-cool: 31.00 ± 0.72; Post-equilibration: 40.42 ± 0.47; Post-thaw: 81.14 ± 1.40).
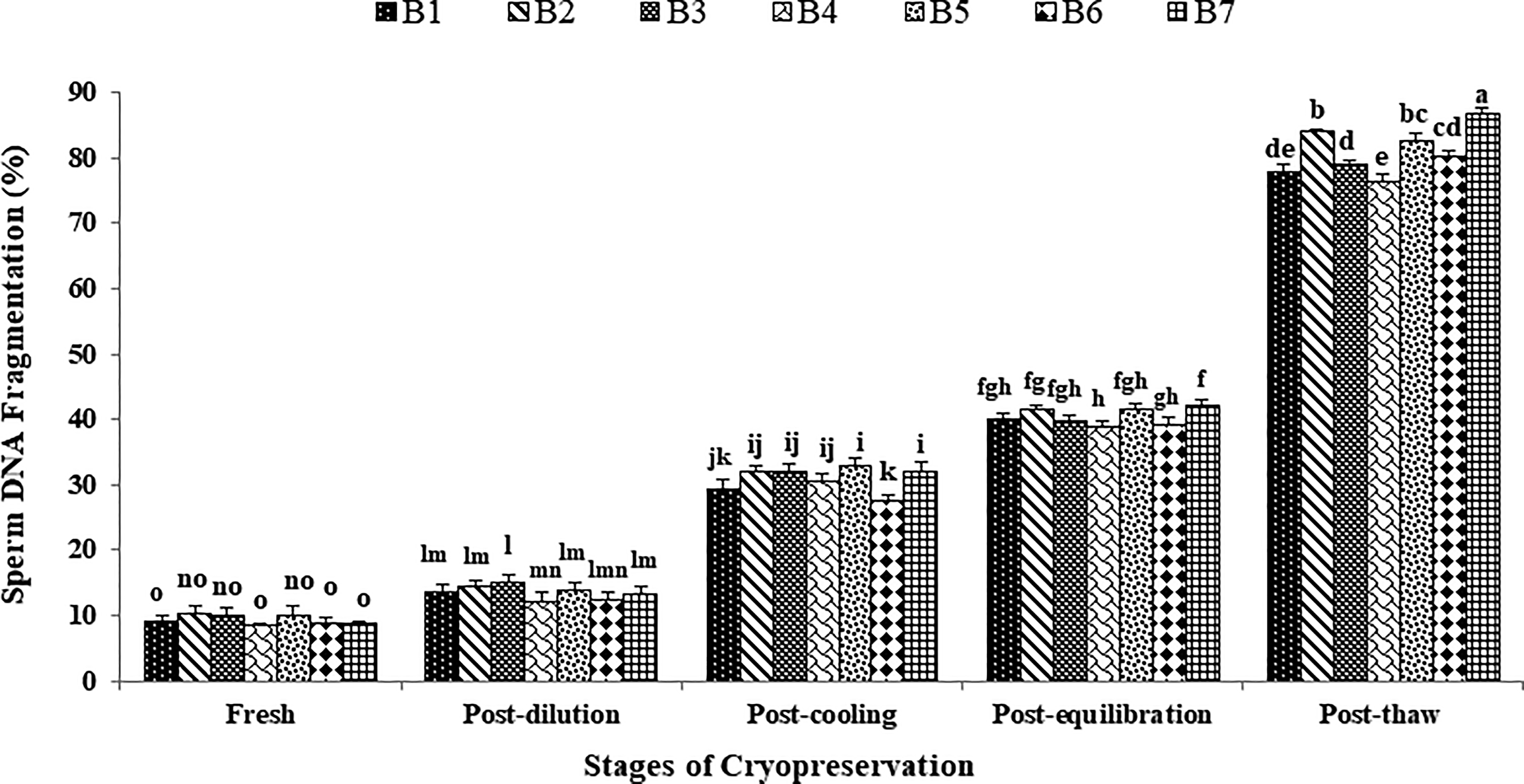
Effect of stages of cryopreservation (fresh, post-dilution, post-cooling, post-equilibration, and post-thaw) on sperm DNA fragmentation (%) of individual Japanese quail (n = 7). Different superscripts indicate statistically significant differences (p < 0.05).
Effect of cryo-storage on free radical scavenging activity
The data on the effect of stages of cryopreservation on free radical scavenging activity (%) in individual Japanese quail (n = 7) semen are presented in Table 2. In the fresh semen and at the post-dilution stage, the highest (p < 0.05) scavenging activity was recorded in bird 4 followed by bird 7, 6, 1, 5, 3, and 2. At post-cooling, higher (p < 0.05) scavenging activity was recorded in birds 4, 6, 1, and 7, whereas at post-equilibration, birds 4 and 6 maintained higher (p < 0.05) scavenging activity compared with other individuals. At post-thaw, bird 4 maintained the highest (p < 0.05) scavenging activity compared with birds 1, 3, 6, 5, 2, and 7. Overall, birds 4 and 1 presented lowest (p < 0.05) and bird 7 presented the highest (p < 0.05) percent decline rate compared with other individuals during cryopreservation (Table 1). Moreover, a significant decrease (p < 0.05) in free radical scavenging activity was observed with the combined mean of all birds at each successive stage (Fresh: 75.15 ± 1.35; Post-dilution: 72.58 ± 1.25; Post-cool: 57.72 ± 1.24; Post-equilibration: 39.62 ± 1.39; Post-thaw: 14.68 ± 1.82).
Effect of Stages of Cryo-Storage on Scavenging Activity, Antioxidant Potential and Mitochondrial Status in the Individual Japanese Quail Semen (n = 7)
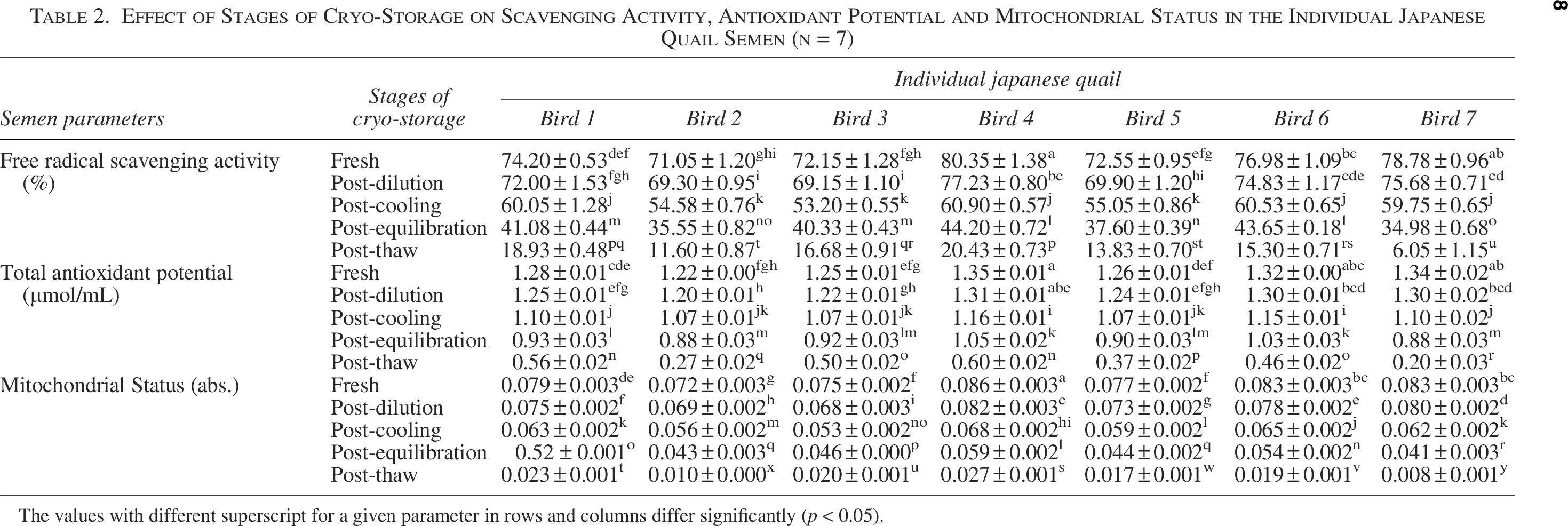
The values with different superscript for a given parameter in rows and columns differ significantly (p < 0.05).
Effect of cryo-storage on FRAP
The data on the effect of stages of cryopreservation on FRAP (µmol/mL) in individual Japanese quail (n = 7) semen are presented in Table 2. In the fresh semen and at the post-dilution, birds 4 and 7 presented higher (p < 0.05) TAP followed by birds 6, 1, 5, 3, and 2. At the post-cooling and post-equilibration, birds 4 and 6 maintained higher (p < 0.05) TAP compared to other individuals. At post-thaw, birds 4 and 1 maintained higher (p < 0.05) TAP compared with birds 3, 6, 5, 2, and 7. Overall, birds 4 and 1 presented the lowest (p < 0.05) and bird 7 presented the highest (p < 0.05) percent decline rate for TAP compared with other individuals during cryopreservation (Table 1). Moreover, a significant decrease (p < 0.05) in FRAP was observed with the combined mean of all birds at each successive stage (Fresh: 1.28 ± 0.01; Post-dilution: 1.25 ± 0.01; Post-cool: 1.10 ± 0.01; Post-equilibration: 0.95 ± 0.02; Post-thaw: 0.46 ± 0.05).
Effect of cryo-storage on mitochondrial dehydrogenases activity
The data on the effect of stages of cryopreservation on activity of mitochondrial dehydrogenases (abs.) in individual Japanese quail (n = 7) sperm are presented in Table 2. In the fresh sperm and at the post-dilution, the highest (p < 0.05) mitochondrial activity was recorded in bird 4 followed by birds 7, 6, 1, 5, 3, and 2. Similarly, at the post-cooling and post-equilibration stages, the highest (p < 0.05) mitochondrial activity was recorded in bird 4 compared with other individuals. At post-thaw, bird 4 maintained the highest (p < 0.05) mitochondrial activity compared with birds 1, 3, 6, 5, 2, and 7. Overall, bird 4 presented the lowest (p < 0.05) and bird 7 presented the highest (p < 0.05) percent decline rate for mitochondrial activity compared with other individuals during cryopreservation (Table 1). Moreover, a significant decrease (p < 0.05) in mitochondrial activity was observed with the combined mean of all birds at each successive stage (Fresh: 0.079 ± 0.001; Post-dilution: 0.075 ± 0.002; Post-cool: 0.060 ± 0.001; Post-equilibration: 0.115 ± 0.067; Post-thaw: 0.017 ± 0.002).
Correlation of Japanese quail semen quality and biochemical parameters at fresh collection and during cryopreservation
Overall correlation of semen parameters in Japanese quail at fresh collection and during cryopreservation (post-dilution, post-cooling, post-equilibration, and post-thaw) is presented in Table 3. At fresh collection, sperm motility, plasma membrane and acrosomal integrity, viability, and scavenging activity positively correlated (p < 0.01) with each other and negatively correlated (p < 0.01) with DNA fragmentation. During cryopreservation, all semen quality and biochemical parameters were strongly and negatively correlated (p < 0.01) with DNA fragmentation while positively correlated (p < 0.01) with each other.
Correlation Among Semen Quality and Biochemical Parameters of Japanese Quail at Fresh Collection and During Stages (Post-Dilution, Post-Cooling, Post-Equilibration, and Post-Thaw) of Cryopreservation
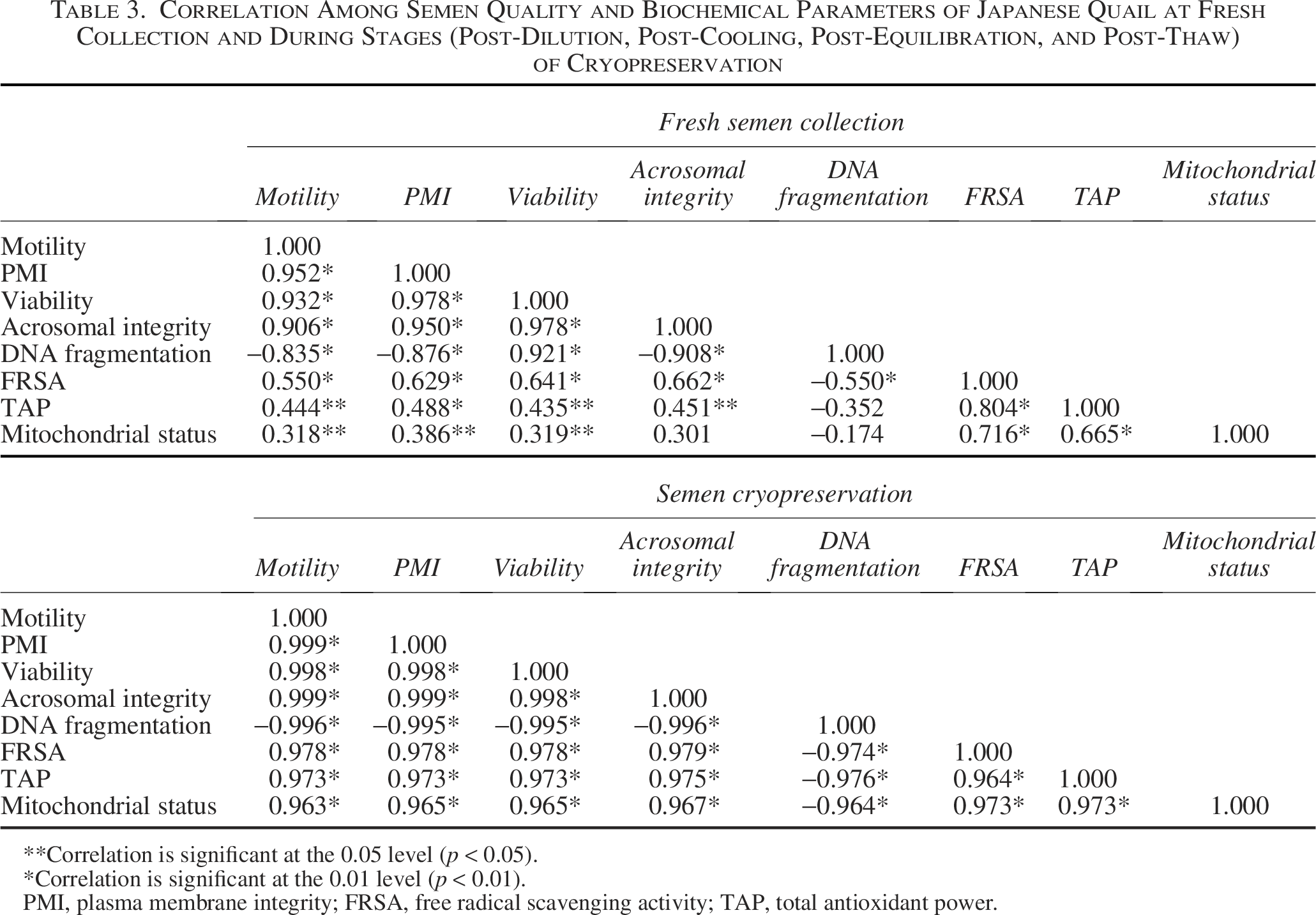
Correlation is significant at the 0.05 level (p < 0.05).
Correlation is significant at the 0.01 level (p < 0.01).
PMI, plasma membrane integrity; FRSA, free radical scavenging activity; TAP, total antioxidant power.
Discussion
The application of avian semen cryopreservation is not yet commercialized for poultry production due to its extreme sensitivity to freezing methods and variable response among species. The cryopreservation process is associated with various physical and chemical stressors imposed on the cells like disturbed energy production or anti-oxidative imbalance. 1 Such fluctuations result in low sperm viability and motility, increased membrane oxidation, DNA fragmentation, apoptosis, and loss of fertility. 17 There are reports of a 30% to 60% reduction in poultry sperm motility at post-thaw and electron microscopy revealed 60% of the sperm organelles bearing irreversible damage.1,18 In addition, individuals even within a species may vary in terms of freeze-tolerance. Hence, the current study was conducted to assess the decline in sperm quality, antioxidant potential, and mitochondrial performance as well as intra-species variation in Japanese quail at various stages of cryopreservation.
During freezing, highly fragmented DNA and declined MMP is known to significantly contribute to the loss of sperm motility. Furthermore, halted oxidative phosphorylation, disrupted tricarboxylic acid cycle, and inactivated antioxidant enzymes are the primary causes of mitochondrial dysfunction amid freezing. 19 The present study showed that sperm motility, viability, mitochondrial activity and TAP declined (p < 0.05) and DNA fragmentation increased (p < 0.05) from fresh collection to the post-thaw stage in all Japanese quail individually and combined. Normally, mitochondria contain a transition pore that briefly opens and elevates the permeability of inner mitochondrial membrane up to the solutes of 1500 Da. This allows calcium and limited ROS to move across the organelle for various physiological needs. Sperm chilling forces a prolonged opening of this pore, causing a rapid collapse of MMP, osmotic shock (that ruptures the mitochondrial membrane), subsequent loss of ATP and unrestrained passing of ROS. This ROS flourishes throughout the sperm and overwhelms the antioxidants, impeding the sperm’s fertilization ability or results in necrosis.19,20 Moreover, mitochondria transfer ATP to the sperm tail microtubules 21 for motility but the freeze-thaw process can reduce motility by 25–75% as a consequence of mitochondrial damage. 22 This probably explains the parallel decline of both parameters during cryopreservation. Furthermore, in the current study, TAP and mitochondrial activity did not illustrate a strong negative correlation with DNA fragmentation at fresh stage. It may be because the DNA is packed inside the nucleus and perhaps it is affected by freezing only when ROS has established itself in a self-perpetuating cycle. 19 So, during cryopreservation, all quality and biochemical parameters showed a strong and negative correlation (p < 0.01) with DNA fragmentation. It has been shown that cryopreservation leads to a decrease in DNA methylation and release of apoptosis-inducing factors from mitochondria that trigger DNA fragmentation. 22 Furthermore, the freeze led decline in antioxidants and viability renders key genes (related to fertility potential) prone to various modifications like sudden arrest or induction of gene transcription, initiation of final transduction pathways, replication errors, or transversions of GC to AT. 23
Avian sperm plasma membrane contains a higher percentage of PUFAs that maintain fluidity and help in fusion, but this characteristic also makes the avian sperm highly susceptible to freeze-thaw damage. Moreover, acrosomal collapse and loss of its contents are also attributed to the freezing process. 2 The present study recorded the decline (p < 0.05) in sperm membrane and acrosomal integrity with reduction (p < 0.05) in the free radical scavenging capacity throughout the cryopreservation stages in all Japanese quail. It has been stated that the freezing process reduces the scavenging capacity of antioxidants like superoxide dismutase as well as diminishes glutathione by approximately 78% compared with fresh semen. 24 Small quantities of ROS are useful for the sperm binding to zona pellucida and acrosome reaction, but after depletion of the antioxidants, production of ROS multiplies and targets the plasma membrane. 14 The presence of more than two carbon-carbon double bonds in the PUFAs mainly adjacent to the methylene group makes the methylene carbon-hydrogen bonds weaker. Hence, hydrogen in PUFAs becomes more vulnerable to abstraction and once stripped off, the attacking free radical is stabilized as a conjugated diene radical, set to be oxidized again in a continuous loop. 25 Therefore, the deficiency of antioxidants is the prime cause of severe plasma membrane abrasion, ions/intracellular enzymes’ leakage, and inhibition of respiration during freezing. Moreover, osmotic stress expands the sub-acrosomal region and oxidative stress ruptures the outer acrosomal membrane which impairs the fertility potential. 24 A strong positive correlation (p < 0.01) among semen scavenging capacity, sperm plasma membrane, and acrosomal integrity in the current study indicates a clear association between the availability of antioxidants and the intact sperm membranes as a determinant of proper sperm function and subsequent fertilization.
Inter-individual variation could serve as the reliable predictor of suitability for semen freezing as all the individuals having high-quality fresh semen may exhibit variable freeze-tolerance.10,26 In the present study, birds 4 and 7 showed the highest (p < 0.05) sperm quality and biochemical activity compared with other individuals at the fresh stage. However, as the freezing stages progressed, bird 4 maintained the higher values while bird 7 showed a sharp decline (p < 0.05) at post-thaw compared to other individuals. Moreover, bird 1 ranked fourth at the fresh stage, showed the second highest (p < 0.05) sperm quality at post-thaw. This intra-species variation with respect to freeze-tolerance is further reinforced by the lowest percent decline rates (p < 0.05) in birds 4 and 1 for the sperm parameters compared with other individuals. These findings suggest that each animal adjusts at a different pace to the same cooling/freezing protocol. One possible explanation is the variable degrees of chromatin compaction. Individuals with strictly compact chromatin are least affected by the freeze damage in contrast to the individuals with slightly relaxed chromatin. 27 Japanese quail displays strong sperm competition, which may have led to such individual differences. 10 However, in the current study, at the fresh stage, all the individual samples had ≥80% motility, and the variable freeze tolerance among individuals is still unanswered. Perhaps the individuality of this feature is under the genetic control 27 as shown in a previous study; greater variability was identified in the sequences of GPx-4 genes in both fertile and infertile men. 28 Although such speculations need future investigation for their confirmation, 29 some proteins have been identified to be linked with the post-thaw changes and their expression patterns were involved with freeze-tolerance.30,31
The severe decline in the overall sperm quality and biochemical performance in the Japanese quail during cryopreservation point toward directing efforts to improve its freezability with the help of exploring the underlying molecular mechanisms and the use of additives. This would improve the effectiveness of semen banking, enhance the breeding efficiency, and allow the planning and management of breeding/propagation programs at optimal times, for not just Japanese quail but other avian species too. Semen banking would also increase sample accessibility to the breeders and researchers enabling the wider use of this freezing technology.
Conclusion
Japanese quail semen quality, antioxidant potential, and mitochondrial status severely decline during the freezing process. Moreover, sperm motility, plasma membrane and acrosomal integrity, viability, mitochondrial status, antioxidant potential, and free radical scavenging capacity strongly and negatively correlate with DNA fragmentation at all the steps of freezing. Intra-species variation is also identified with respect to freeze-tolerance in all the sperm quality and biochemical parameters evaluated. The extent to which the studied parameters and inter-individual variation are related to the fertility of cryopreserved sperm in Japanese quail needs further exploration.
Authors’ Contributions
S.Z.: Management of Study Animals (lead), Investigation (lead), Methodology (lead), Data acquisition (Lead), Formal Analysis (lead), Data Curation (lead), Validation (supporting), Writing—Original draft (lead), Writing—Review and editing (lead). B.A.R.: Conceptualization (lead), Project administration (lead), Supervision (lead), Resources (lead), Writing—Review and Editing (equal), Validation (supporting). S.A.: Conceptualization (lead), Resources (lead), Visualization (lead), Validation (lead), Writing—Review and editing (equal).