
Abstract
Plant germplasm is critical for maintaining biodiversity and mitigating genetic erosion. While cryopreservation has emerged as an increasingly vital approach for conserving plant genetic resources, its routine application remains limited due to species- and explant-specific preservation requirements. In many instances, additional experimentation is necessary to develop genotype-specific protocols to achieve satisfactory survival rates. In addition, the technical operation is time-consuming and labor-intensive. Moreover, the survival rate of explants post-cryopreservation from many species remains suboptimal. PVS-based vitrification cryopreservation of dormant buds represents an ideal strategy to address these challenges, leveraging the buds’ inherent cold hardiness and robust regrowth capacity. This review focuses on four key determinants of successful PVS-based cryopreservation: the collection time and physiological status of dormant buds, shoot tip dissection, PVS exposure duration, and recovery culture. The insights provided herein may facilitate the rapid development of new protocols or user-friendly procedures for a broader range of plant species, enabling wider adoption by institutions and laboratories, and thereby enhancing global efforts in germplasm conservation.
The Conservation of Plant Species
Preserving the diversity of plant genetic resources is crucial for environmental protection and breeding, as these resources serve as strategic assets for safeguarding national food security, promoting ecological sustainability, and ensuring energy security.1B2 -4 Currently, in situ preservation and seed banks are the most employed methods for conserving plant germplasm.1,5,6 However, in situ preservation is vulnerable to adverse weather, natural disasters, diseases, and pest invasions.1,6 For clonal plants, germplasm resources require annual field cultivation, demanding significant labor and financial investment for maintenance.3,7-9 Seed bank preservation is limited to species with dryness or cold sensitive-seeds, low seed viability, or inaccessible seeds.4,10,11 Thus, alternative approaches such as cryopreservation have been explored to maximize the genetic potential of germplasm while mitigating risks associated with traditional methods.
Cryopreservation involves cooling cells or tissues to ultra-low temperatures (typically −196°C in liquid nitrogen [LN]), halting all cellular division and metabolic processes, and enabling theoretically indefinite storage.4,12,13 This method also minimizes somaclonal variation, a critical advantage for germplasm preservation.1,7,14 Cryopreservation is widely recognized as the safest and most cost-effective long-term strategy for conserving plant genetic resources,7,11,13,15,16 attracting substantial attention from geneticists and breeders, and becoming indispensable for plant germplasm conservation.14,17,18 To date, nearly all major agricultural and horticultural crops have research reports with respect to cryopreservation, and various plant tissues and organs, including shoot tips, dormant buds, callus, embryonic axes, seeds, pollen, and root tips, could survive post-cryopreservation.19,20 However, large-scale routine application remains restricted to few species due to genotype-specific and explant-specific requirements. For most species or different genotypes of the same species, post-cryopreservation survival rates are still low and cannot meet the preservation requirements (>40% recovery rate). 21 Thus, adapting cryopreservation protocols to multiple species and a wide array of genotypes remains a significant challenge.
Dormant bud cryopreservation
Dormant bud cryopreservation offers an ideal strategy to address these challenges, leveraging the natural cold hardiness and strong post-cryopreservation regrowth capacity of dormant buds. Sakai (1960) conducted the first cryopreservation study, demonstrating that winter bud sticks of poplar and willow could survive in LN. 15 Since then, dormant bud cryopreservation has gained increasing attention for conserving germplasm of woody plants, particularly fruit trees.22-24 Dormant buds have proven to be efficient and cost-effective materials for cryopreservation, with simple protocols applicable to twigs of some temperate and tropical woody species. 25 A study on cryopreservation of apple germplasm using dormant buds and in vitro shoot tips found that the time and labor for cryopreservation of dormant buds were 40% and 50% of those required for cryopreservation of in vitro shoot tips, respectively. 26 Currently, the USDA-ARS facilities in Fort Collins and Corvallis preserve 2155 apple germplasm accessions using twigs with dormant buds. 4 Slow-freezing cryopreservation of apple twigs with dormant buds has achieved 90% survival after 10 years of storage. 23 Traditional slow-freezing protocols for dormant buds involve dehydrating bud segments to low moisture content followed by gradual low-temperature acclimation.4,26,27 However, this technique relies on expensive controlled-rate freezers to achieve freeze-induced dehydration and is primarily suited for cold-tolerant species and undifferentiated cultures and apices, which limits its widespread use.8,20,28,29 In recent years, an increasing number of studies have reported successful dormant bud cryopreservation using PVS-based protocols. This method involves exposing explants to concentrated cryoprotectant solutions for dehydration and could induce a glassy state (vitrification) in cellular water to prevent ice crystallization and tissue damage during immersion in LN; thereby, ice crystallization is prevented and plant tissues survive.8,10,12,30
Compared with slow freezing, PVS-based cryopreservation is simpler to perform, suitable for larger and complex organs, achieves higher post-cryopreservation survival rates, and requires no specialized equipment.10,31 This protocol has been applied to over 100 plant species and is gaining prominence as a promising alternative to slow freezing for long-term plant genetic resource conservation.1,10,30
PVS-based vitrification cryopreservation of dormant buds has achieved high regeneration rates in numerous species, including blackcurrant cultivars, 32 Malus, 33 Salix, 34 Vaccinium, 35 cherry, 7 persimmon,12,13,36 Betula pendula Roth, 37 Astragalus membranaceus (Fisch.) Bge. var. mongholicus (Bge.) Hsiao, 38 and Gentiana straminea Maxim. 39 Moreover, dormant bud cryopreservation has also exhibited distinguished superiority over the shoot tips of in vitro plants, such as Rosa canina, R. rubiginosa, 40 R. pomifera, 16 persimmon,41,42 and Actinidia arguta. 43 Additionally, the possibility of genetic instability is minimized, and plant regeneration post-cryopreservation is easier than other propagules since the shoot tip of dormant bud is totipotent and could sprout directly or produce multiple shoots or bulblets in vitro without forming callus as an intermediate stage (Fig. 1). As a result, PVS-based vitrification cryopreservation holds great promise as an effective preservation strategy for perennial plants producing dormant buds whatever is woody and herbaceous plants. This review provides updated and comprehensive information on the development and recent progress of dormant bud cryopreservation with high regrowth levels across diverse species (Table 1).
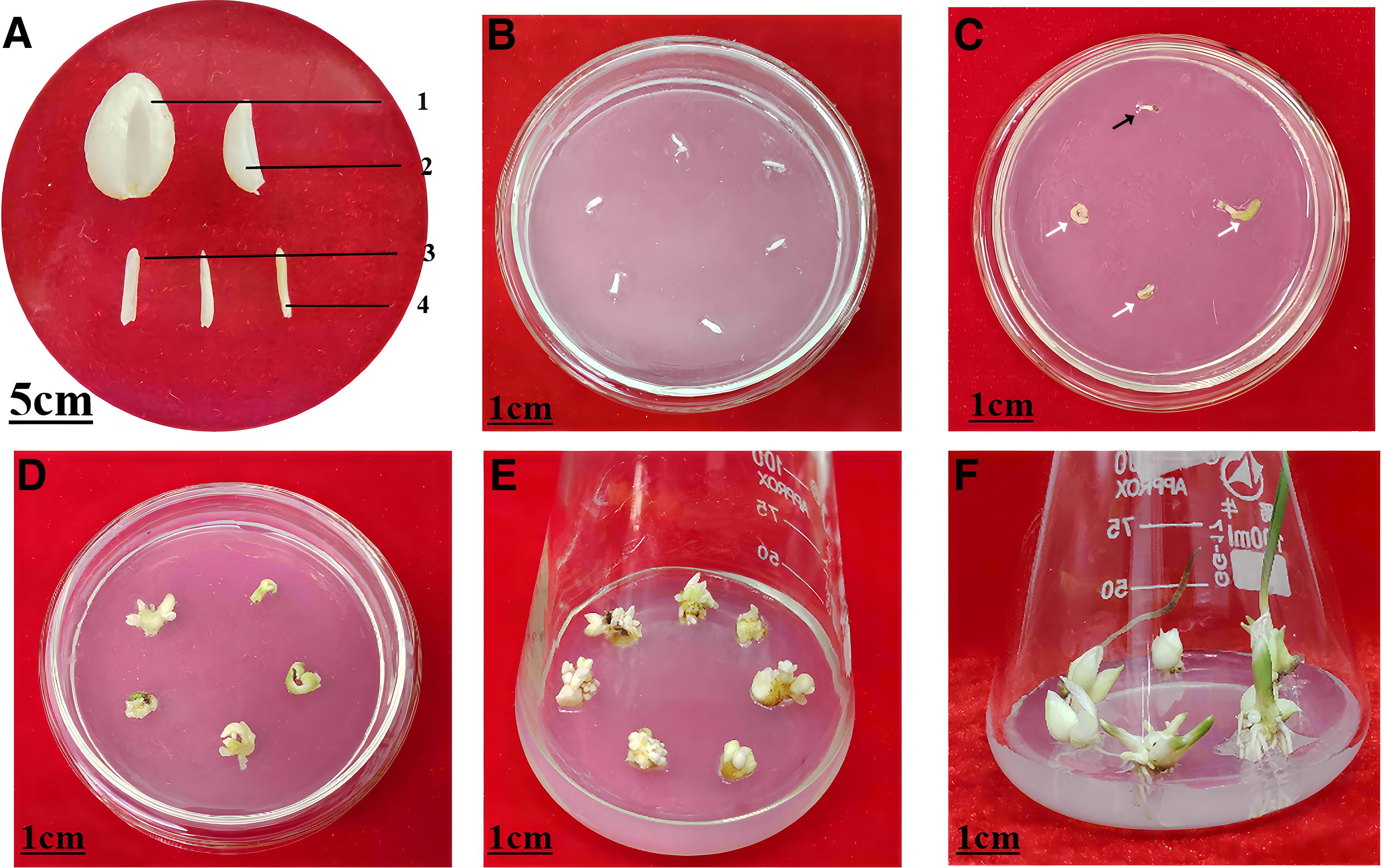
The anatomical structure of bulbs of Fritillaria przewalskii and the recovery process of dormant buds after cryopreservation.
Outcome of Cryopreservation of Dormant Buds by PVS-Based Vitrification Protocol
LS, loading solution; LN, liquid nitrogen; NAA, 1-naphthaleneacetic acid; TDZ, Thidiazuron; BAP, Benzylaminopurine; PVP, Polyvinylpyrrolidone; DKW, Driver Kuniyuki & Walker.
PVS-based vitrification methods
Several vitrification-based cryopreservation technologies have been successfully developed for dormant buds, including vitrification, 52 droplet-vitrification (DV), 53 and encapsulation-vitrification (EV). 54 The standard vitrification protocol involves treating explants with a loading solution (LS; typically 2 mol/L glycerol + 0.4 mol/L sucrose, pH 5.7), 55 followed by high-concentration vitrification solutions with the purpose of explant dehydration. Common types are PVS1 (22% glycerol + 13% ethylene glycol + 13% polyethylene glycol + 15% DMSO), 56 PVS2 (30% glycerol + 15% ethylene glycol + 15% DMSO + 0.4 mol/L sucrose), 52 and PVS3 (50% glycerol + 50% sucrose). 55 After PVS treatment, explants are plunged into LN for storage. For plant regeneration, explants are rapidly thawed in a water bath, exposed to an unloading solution (MS medium with 1.2 mol/L sucrose, pH 5.7) 5 to remove cryoprotectants, and finally recovered on hormone-supplemented medium. In DV, cryoprotectant solutions are applied as single droplets on aluminum foil to enable ultra-rapid cooling and thawing.57,58 EV combines vitrification and encapsulation-dehydration, involving explant encapsulation in alginate beads, followed by loading and PVS dehydration.5,59 Critical factors must be considered for applying dormant bud cryopreservation to germplasm conservation. This review summarizes current research on PVS-based cryopreservation of dormant buds and highlights factors enhancing survival and regeneration. Our goal is to stimulate further research and development in dormant bud cryopreservation, supporting the preservation of genetic diversity in perennial plants.
Plant species and genotypes
Genotype significantly affects the recovery rate in cryopreservation of most plant species, though its impact is less pronounced in PVS-based vitrification cryopreservation of dormant buds. Forsline applied slow-freezing cryopreservation to 64 apple accessions, with 60 of them achieving an average recovery growth rate exceeding 65%. 22 For Malus, 180 samples representing 32 species were cryopreserved using dormant buds over a 10-year period, yielding an average post-cryopreservation survival rate of 39%. 60 In Pyrus, cryopreservation of dormant buds from 35 accessions across 21 species showed that the PVS2 method enabled more genotypes to meet the 40% survival baseline. 60 Similarly, vitrification of dormant buds from 20 apple accessions resulted in shoot tip viability ranging from 58.3% to 74.2%. 61 Guyader et al. cryopreserved dormant buds from 15 Malus and Pyrus cultivars, recording average recovery rates of 31.8% and 26.9%, respectively. 46 In DV of six persimmon accessions, survival rates exceeded 79%. 62
The strong universality of PVS-based vitrification within genera is further demonstrated in blackcurrant, 32 Pyrus, 50 almond, 63 and grapevine. 64 Even conspecific plants from diverse ecological environments achieved favorable results using dormant buds. For example, dormant buds from 24 apple varieties exhibited >70% survival, and protocols optimized for cold-hardy North American apple cultivars were adaptable to less cold-hardy varieties in Denmark’s temperate oceanic climate. 65 Jenderek et al. confirmed no significant differences in post-cryopreservation viability among dormant buds from three apple cultivars grown across distinct U.S. locations. 66 Additionally, a Shimane-developed persimmon dormant bud protocol was successfully applied to 16 temperate cultivars from Japan, China, and Korea, achieving an 89% average shoot formation rate. 13 Growing interest in dormant bud cryopreservation has driven protocol development for deciduous trees and shrubs, with varying success reported in peony, 67 blueberry, 35 aspen, 68 currant, 69 little walnut, English walnut, and black walnut. 49 Subsequent studies extended vitrification to herbaceous species, including G. straminea Maxim.,39,70 Codonopsis pilosula (Franch.) Nannf.,70,71 and A. membranaceus. 38 The above research results indicate that dormant buds enable cryopreservation of numerous accessions within a genus, with most achieving acceptable regrowth.
Physiological state of dormant bud
Developing highly efficient cryopreservation protocols for germplasm banks requires optimizing not only the operation technology but also the physiological status of the shoot tips,5,44,72 which is mainly determined by the extent of cold acclimation.25,73-75 The reason is that adequate natural cold acclimation induces cell dehydration22,24,76,77 and promotes synthesis and accumulation of freezing tolerance solutes, such as soluble sugars, sugar alcohols, organic acids, amino acids, and polyamines in plants. 78 Most dormant bud studies collect branches or scions in December–February, as early fall samples often lack cryotolerance and are susceptible to suffer freeze damage.75,77 However, optimal timing varies by species and region. For peony, buds collected in late November—late February yielded 66%–74% regrowth, but dropped to <1% in late March. 67 In Finland, the regeneration of hybrid aspen buds post-cryopreservation increased from 9.8% (September) to 75% (October–February). 68 Dracocephalum austriacum buds collected in late December achieved 100% regeneration, declining to 76% in February and 30% in March. 79 Moreover, late December collection is critical for high survival in Pyrus, 50 blueberry,35,79 American elm, 72 and A. arguta. 43
It is often desirable to collect materials in their cold-hardiest state from the field. However, different accessions or species exhibit distinct acclimation kinetics, and the degree of cold hardiness may vary annually due to temperature fluctuations, so this state is not always easy to discern or test. In temperate species such as apple and pear, it has been documented that buds should only be harvested after the plants have been exposed to subzero temperatures for a minimum of 72 hours. 77 Ravish (2023) 26 reported on dormant buds harvested from subtropical zones, where temperatures do not drop below 12°C in any season. Moisture levels in buds during three winter months—when leaf abscission occurred—were used as a criterion. During natural acclimation, buds are reported to lose water content to varying extents in December, January, and February. The results showed that buds harvested in February with the lowest moisture level were optimal for preservation. In addition, cold acclimation can be artificially induced and significantly improves the post-cryopreservation survival rate of shoot tips. Jenderek et al. found that blueberry buds harvested under conditions where the average maximum air temperature was below 11.2°C showed higher post-cryopreservation viability, ranging from 52% to 80%, in Fort Collins, CO. 35
For in vitro plants, cold acclimation is a highly effective method, typically achieved by exposing them to low temperatures (0°C–4°C) or alternating between daytime exposure at 20°C and nighttime exposure at 3°C for more than 2–3 weeks. 80 For field-grown plants, woody branches or organs bearing dormant buds can be stored under low-temperature conditions for further acclimation, which enables the dormant buds to tolerate the stresses incurred during cryopreservation. The most common approach involves enclosing the material in polyethylene bags and storing it in a refrigerator at −5°C to 3°C for 1 week to 3 months prior to use, which generally enhances cold hardiness while maintaining plant viability.75,77 However, the optimal storage duration varies across species. Towill and Ellis observed that apple branches stored for 6–8 months could be successfully cryopreserved, whereas the regenerative capacity of sweet cherry branches stored for more than 4–6 months declined significantly. 25 For herbaceous perennials, subterranean organs with dormant buds must be collected pre-soil freezing, though cold acclimation is often insufficient. Our studies showed that excavating C. pilosula 71 and F. przewalskii subterranean organs in October—early November and storing them in moist soil within polyethylene bags at 2°C–4°C for 1–3 months could gradually improve dormant bud survival rate post-cryopreservation with prolonged the preservation time (unpublished, Fig. 1). Notably, A. membranaceus dormant buds collected in March achieved 100% survival after 2 weeks of artificial cold acclimation. 38
Shoot tip size
Shoot tip size significantly affects post-cryopreservation recovery. 27 Optimal sizes range from 1 to 3 mm 27 with most protocols using about 1 mm.27,59,81-83 Cryopreservation of 0.5–1 mm shoot tips dissected from dormant buds of American elm, 72 Diospyros kaki Thunb.,13,84 and almond 63 all achieved 80%–100% regeneration. Smaller apices are susceptible to vitrification solution toxicity, reducing vitality and survival, and larger apices, conversely, exhibit poor cryoprotectant permeability, leading to incomplete dehydration, and high water content reduces cold tolerance of shoot tips and ultimately results in lower survival. 71
However, dissecting 1 mm-long shoot tips from growing plantlets is time-consuming and often reduces explant vigor. In contrast, a simple method can enable efficient isolation of small shoot tips from dormant buds. First, dormant buds can be slittered at an appropriate angle, exposing inner shoot tips, and then excised with a scalpel to yield abundant, vigorous tips rapidly.38,39,70,71 Moreover, our research found 2–4 mm shoot tips from F. przewalskii dormant buds achieved optimal survival by vitrification (unpublished), and even whole dormant buds of C. pilosula in big volume also exhibited strong survival and regeneration ability, 71 which suggested that vitrification was a viable long-term preservation method for larger explants and could greatly simplify the steps of shoot tip dissection.
Shoot tip preculture
Preculture is a treatment in which shoot tips are cultured in a medium supplemented with high concentrations of sugar, DMSO, sorbitol, or mannitol. 80 These substances could increase medium osmotic pressure leading to cell dehydration or permeate cells and reduce free water, which could reduce the cell freezing point and then further decrease the formation of ice crystals. 80 Sucrose is the most widely used sugar in preculture, with proven efficacy for dormant bud cryopreservation in B. pendula Roth., 37 G. straminea Maxim., 39 Tamarix taklamakanensis, 45 Prunus dulcis, 3 and almond. 63 Typically, shoot tips are cultured in medium adding 0.3–1.0 mol/L sucrose for 1–7 days,5,27,51 but optimal preculture duration must be determined experimentally as species-specific variation exists. Preculture is unnecessary for dormant buds collected at optimal times, such as persimmon, 84 C. pilosula (Franch.) Nannf., 71 and F. przewalskii (unpublished).
Loading
Loading involves immersing materials in cryoprotectant solutions at room temperature for a specified period to reduce cellular water content and prevent osmotic shock caused by PVS solution. 6 Commonly used loading solutions include 0.2 mol/L glycerol + 0.4 mol/L sucrose or 60% PVS2, with a 20-minute exposure—a routine parameter across most species and explant types.5,9 Loading solutions have been repeatedly validated for in vitro-grown and dormant bud shoot tip cryopreservation.2,13,85 Rasl et al. found that while loading did not affect survival, it significantly inhibited regeneration of Lamiaceae dormant buds cryopreserved via DV. 79 Notably, satisfactory survival of D. kaki Thunb. dormant buds was achieved via DV without loading. 84
Plant vitrification solutions
Application of concentrated vitrification solutions is critical for cryopreservation: they accelerate dehydration, lower freezing points, and induce vitrification (amorphous solidification) of cellular fluids, which avoid the formation of large amount of ice crystals and thereby reduce ice-induced injury. 60 PVS2 is the most widely used vitrification solution.6,37,52,86 DMSO as a key component exhibits lower toxicity at 0°C–5°C than at 20°C–25°C. 27 Explants can tolerate longer PVS2 exposure at lower temperatures; thus, PVS2 treatment is typically performed at 0°C. 27 For example, axillary buds of greenhouse-grown Rosa exposed to PVS2 at 0°C via DV retained 100% survival even after 90-minute exposure, whereas room-temperature PVS2 treatment significantly reduced survival. 87 Similar findings were reported for shoot tips of Nephelium ramboutan-ake, 88 mint, 89 and Populus alba L. 2 Optimal PVS2 exposure varies by species, with 20–120 minutes most common. For instance, DV of shoot tips from test-tube plantlets of Rosa, 40 potato, 90 Colocasia esculenta var. esculenta, 91 shallot, 92 kiwifruit, 93 and yacon 94 used 20–60 minutes PVS2 exposure at 0°C. In contrast to dormant buds, they exhibited broad PVS2 tolerance due to high dehydration and cold stress resistance. A. arguta dormant buds treated with PVS2 for 120–300 minutes at 0°C achieved >60% survival 43 ; G. straminea Maxim. buds exposed for 20–60 minutes yielded >65% survival 39 ; R. pomifera buds treated for 10–30 minutes achieved >58% survival 16 ; and R. canina and R. rubiginosa dormant buds treated for 0–30 minutes via DV achieved >90% regeneration. 40 When dormant buds of C. pilosula were incubated for 20, 40, 60, and 80 minutes at room temperature, all resulted in a high survival rate of more than 65% using vitrification. 71 For A. membranaceus dormant buds, 20, 50, and 80 minutes incubation resulted in a survival rate of more than 80%. 38 In cryopreservation of persimmon dormant buds, the survival rate increased with the longer exposure duration, and the best exposure time was 90 minutes by vitrification. 12 In another report, persimmon dormant buds exposed to PVS2 for 10, 20, and 30 minutes obtained a survival rate higher than 60%. 13 10, 20, and 30 minutes incubation in PVS2 attained a survival rate of higher than 58% in cryopreservation of dormant buds of R. pomifera.
PVS3 is also a kind of widely used plant vitrification solution and is recommended for samples sensitive to chemical toxicity and tolerant of osmotic stress, while its penetrative property generally necessitates a longer exposure time compared with PVS2.27,95 Kami et al. demonstrated that a balanced composition of PVS A3 solution can be more beneficial for the recovery of cryopreserved samples compared with the original PVS2 solution, while PVS3 solution is recommended to use for samples sensitive to chemical toxicity and tolerant to osmotic stress. 96 In mint shoot tips, PVS3 has shown lower toxicity to sensitive shoot tips than PVS2.27,89 Bae et al. reported an average regrowth rate of 61.5% in 26 strawberry accessions cryopreserved using the PVS3-based DV protocol. 97 In the cryopreservation of dormant pear buds, results indicated that PVS3 yielded the highest viability after cryopreservation compared with other cryoprotectants, including honey, PVS2, and PVS4. 98
Thawing and unloading
When the germplasm is needed, thawing is first carried out, which is essential for all cryopreservation protocols.27,90,99 Severe cellular damage often occurred during thawing, such as shrinkage and cytoplasmic melting, which were observed in peripheral cells of G. straminea Maxim. 70 and C. pilosula (Franch.) Nannf. (unpublished) after thawing. Slow-cooled dormant twigs are typically thawed slowly at 0°C–4°C for 24 hours36,42,100 or at 37°C for 3–4 minutes. 68 In contrast, PVS-based vitrification of dormant buds uses rapid thawing in 37°C–40°C water baths to prevent recrystallization,5,27,54,101 while DV often uses 20–30 minutes of room-temperature thawing.5,27,102
In order to avoid the PVS toxicity to shoot tips post-thawing and to prevent uncontrolled rehydration of shoot tips, unloading is necessary.5,27,103 Unloading is typically washing the explants in MS medium with 1.2 mol/L sucrose for 20 minutes. 54 However, A. membranaceus 38 and F. przewalskii (unpublished) dormant buds achieved high survival without unloading, which suggested dormant bud was a wonderful preservation material.
Post-Cryopreservation Regeneration of Dormant Buds
The development of efficient regeneration methods for explants retrieved from LN is of utmost importance in the cryopreservation process. Conventional propagation methods such as cuttings and grafting have certain limitations, as grafting is highly susceptible to internal factors (e.g., compatibility, nutrient and moisture content of the scion, cambial activity) and external factors (e.g., atmospheric temperature, soil composition). Therefore, it is complex and unstable and needs superior skill to complete. 104
It is well established that shoot tip in vitro culture is the most critical technique in micropropagation, wherein shoot tips are often easily cultured to regenerate multiple shoots without undergoing a callus phase. Cryopreserved shoot tips derived from dormant buds, which possess meristems and have a relatively large volume, are more likely to sprout or produce multiple shoots, thereby offering high proliferation efficiency and genetic stability. 71
The establishment of a recovery system, first and foremost, depends on the acquisition of aseptic shoot tips. Therefore, preventing contamination is technically pivotal when preparing shoot tips from buds under sterile conditions. Because the dormant buds are tightly wrapped, the shoot tip, being the innermost part of the dormant bud, is often in a sterile state. Shoot tips can be obtained by peeling off the outer one or two layers of the bud, followed by surface sterilization. The bud is then longitudinally bisected at an angle with a scalpel to expose the shoot tip, which is finally dissected using the scalpel tip. This method yields a survival rate of nearly 100%, 38 which is time-efficient and simple, and ensures the viability and sterility of the shoot tips.
Recovery media are generally based on MS, 1/2 MS, modified MS, or WPM basal media, supplemented with high cytokinin and low auxin concentrations to induce shoot morphogenesis.12,13,72,84 The most commonly used cytokinin is 6-benzylaminopurine (BA), with concentrations ranging from 0.05 to 4 mg/L. 18 In some species, BA is used in combination with indole-3-butyric acid, indole-3-acetic acid, gibberellic acid, and 1-naphthaleneacetic acid. Reactive oxygen species-induced oxidative stress is regarded as a major factor causing explant death, which severely limits the further development of plant genetic resource cryopreservation.105-107 Exogenous application of antioxidants and anti-stress agents, such as vitamin C, ascorbic acid, and melatonin, has been shown to improve the recovery of cryopreserved plantlets.72,88,108
Given that cryostorage is associated with freezing and desiccation stresses, shoot tips undergoing recovery growth are typically cultured in the dark for 3–7 days due to freeze-induced damage; subsequent enhanced lighting may be required to support morphogenesis.18,70 Exposure of tissues to light during the early recovery phase after LN treatment has been found to increase damage in shoot apex cultures. 109 Choudhary et al. reported that buds of six Morus species showed no recovery when directly cultured under light conditions, whereas viability ranging from 12% to 100% (with an average of 52%) was achieved when culture conditions were modified to include dark incubation. 26 In the cryopreservation of dormant buds of G. straminea Maxim., 70 persimmon, 36 A. arguta, 43 Amygdalus communis L., 63 R. canina and R. rubiginosa, 40 R. pomifera, 16 and pears, 50 dark incubation was adopted too.
Genetic Stability of Cryo-Derived Plants
Maintaining the genetic stability of cryostored materials is of paramount importance for plant germplasm conservation. Shoot tips possess unique advantages: they contain meristems capable of direct germination or producing multiple shoots. Multiple shoot morphogenesis not only provides high proliferation efficiency but also exhibits strong genetic stability, making it an attractive alternative for graft propagation. 71 For testing the genetic fidelity of post-cryopreservation regenerants, morphometric analysis has served as a simple and cost-effective method. Pawłowska and Szewczyk-Taranek observed phenotypic stability in regenerants of pentaploid roses by DV. 110 Kwaśniewska (2017) performed biometric analyses of cryo-derived regenerants of R. pomifera “Karpatia” during the stages of shoot multiplication, rooting, and acclimatization, which revealed no changes compared with non-cryopreserved samples. 16
In cryopreservation studies on Dioscorea rotundata Poir., 111 apple, 112 sugarcane, 113 Dendranthema grandiflora Tzvelev, 114 and grape, 115 progenies regenerated from shoot tips have shown genetic stability compared with nonfrozen materials. However, occasional reports have documented genetic variations in plantlets regenerated through cryopreservation.114,116B117 -119 According to Martín et al., 120 such genetic variations may be associated with tissue culture conditions. It is widely acknowledged that callus formation during tissue culture can also induce genetic variability and thus should be avoided.27,74,121,122 Shoot tips directly germinating or producing multiple shoots minimize variation risk. 71 Kulus found EV better preserved genetic stability than DV or vitrification, likely due to physical protection of explants. 123 Therefore, ensuring genetic fidelity requires attention to variation-inducing factors and further research to address these issues.
Cytology and Histology Studies During Cryopreservation
Changes and damage to shoot tips during the process of cryopreservation can be observed via histological sectioning. Freezing injury is linked to increased vacuolization: the more vacuolized plant cells are, the more water needs to be removed to prevent lethal ice crystallization. 124 Hence, it is crucial to dehydrate tissues to avoid lethal intracellular freezing prior to rapid freezing, thereby improving post-cryopreservation survival rates. 26 High concentrations of sucrose and glycerol—key components of loading solution (LS) and PVS2—induce hyperosmotic stress in cells, resulting in cellular dehydration. After dehydration with the loading solution, cells in shoot tips lose water and exhibit slight plasmolysis. Subsequent exposure of the shoot tips to higher osmotic pressure via PVS2 treatment causes further cell dehydration, leading to further plasmolysis, which is conducive to cell survival.125-127
Freezing and rewarming treatments could cause severe damage to buds, primarily characterized by cell wall breakage, blurred nuclear boundaries, cytoplasmic disintegration, and nucleolar disappearance.128,129 Following vitrification-based cryopreservation, surviving cells are mainly observed in the apical meristem and youngest leaf primordia, which remain intact. The phenomenon has been documented in the cryopreservation of potato, 130 banana, 124 and apple. 130 The reason lay in small cells in the apical meristem, possessing a high nucleocytoplasmic ratio, dense cytoplasm, and no vacuoles, which did not freeze during cryopreservation and exhibited a higher survival rate compared with mature cells characterized by large vacuoles and small nuclei. 130 Zeng et al. reported that during the cryopreservation of papaya shoot tips, large vacuoles fragmented into small vacuoles and the endoplasmic reticulum underwent swelling. 131 Similarly, Takashi et al. observed numerous small vacuoles, piroplasms, and swollen structures. These findings suggest that cells may employ such adaptive strategies to enhance their survival post-cryopreservation. Due to the requirement for large quantities of shoot tips in physiological and omics research, studies in this field remain limited. Fortunately, intact dormant buds of some plant species could be successfully preserved, 70 which could provide conditions for unraveling the complex molecular mechanisms of vitrification-based cryopreservation.
Conclusion and Future Prospects
Success in PVS-based cryopreservation depends on five key factors: dormant bud collection time and physiological status, shoot tip dissection, PVS exposure duration, and recovery culture. Dormant bud collection during optimal physiological status (December–February) is critical. Cryopreserved shoot tips from dormant buds—with meristems and larger volumes—are easily dissected, sprout readily, produce multiple shoots, and offer high post-cryopreservation recovery efficiency and genetic stability. Dormant buds tolerate a broad range of cryoprotectant exposure times, though excessive or insufficient duration is fatal to in vitro shoot tips. PVS-based cryopreservation of dormant buds offers advantages, including simplified handling, high survival rates, and broad applicability to herbaceous and woody plants, making it a highly promising strategy for large-scale plant genetic resource cryobanking. This review aims to facilitate the development of new, user-friendly protocols for more species, enabling wider adoption by institutions and enhancing germplasm preservation efforts.
Footnotes
Author Disclosure Statement
The authors named in this article declare that they are responsible for the work conducted in this study and accept full liability for any claims related to the content of this article.
Funding Information
This research was funded by the National Natural Science Foundation of China (No. 81960683), the Science and Technology Project of Gansu Province (No. 21JR1RA263, 24YANA007, 23YFFA0027, 24CXKA009, 22CX8NA082, and 25JRRA1178), and the Industrial Support Project of Colleges and Universities in Gansu Province (No. 2020C-09 and 2024CYZC-40).