
Abstract
Introduction:
Neisseria meningitidis, the primary pathogen of epidemic meningococcal meningitis, is a typical fastidious bacterium with stringent nutritional requirements and high environmental sensitivity, making routine preservation challenging. Current cryopreservation methods lack quantitative efficacy evaluation and suffer from low viability.
Objective:
This study aimed to screen optimal permeable cryoprotectants (CPAs) for fastidious bacteria typified by N. meningitidis via cytotoxicity, cryopreservation viability, and ice crystal inhibition assays and elucidate the underlying protective mechanism.
Materials and Methods:
Viable cell counting with ANOVA and Tukey’s test assessed the cytotoxicity and post-cryopreservation survival of N. meningitidis treated with 10/15/20% glycerol and 5%, 10%, 15% dimethyl sulfoxide (DMSO), formamide (FMD), and propylene glycol (PG) in PBS/brain heart infusion (BHI) at 37°C for 15 min and frozen at −80°C for 48 hours. Differential scanning calorimetry (DSC) analyzed thermodynamic parameters of CPA solutions, and cryomicroscopy determined ice-inhibiting properties by observing ice crystal morphology and growth rate.
Results:
Cytotoxicity assays showed N. meningitidis survival rates of 88.14%, 83.85%, and 81.62% in BHI broth with 5% DMSO, 5% FMD, and 5% PG, respectively. After 48 hours of −80°C cryopreservation, 5% DMSO exhibited the best protective effect in both PBS and BHI (p < 0.05), with survival rates of 64.93% and 63.11%. DSC analysis revealed decreased crystallization enthalpy (ΔHc) and melting temperature (Tm) of DMSO with increasing concentration in BHI. Cryomicroscopy confirmed 5% DMSO most effectively inhibited ice crystal growth (921.57 μm2 in PBS, 814.20 μm2 in BHI within 50 s).
Conclusion:
5% DMSO in BHI broth is the optimal permeable CPA for N. meningitidis and other fastidious bacteria. Its potential mechanism is that 5% DMSO balances bacterial osmotic pressure, alleviates solute and ice crystal damage, and balances ice inhibition with biocompatibility to enhance bacterial survival.
Introduction
Neisseria meningitidis is the primary pathogen responsible for epidemic meningococcal meningitis and septicemia. Globally, this bacterium causes approximately 300,000 infections annually, with a mortality rate as high as 10%−15%, and about 20% of survivors suffer from permanent neurological sequelae. 1 N. meningitidis is a fastidious, encapsulated, aerobic Gram-negative diplococcus with a kidney-bean morphology, and its optimal growth conditions are 35°C–37°C with 5%−10% CO2. It forms gray, non-hemolytic, convex, smooth colonies with distinct edges on blood agar, yet it is prone to autolysis during the stationary phase. Most strains are able to ferment maltose and grows on various media, including blood agar and tryptic soy agar. 2 The cell envelope beneath the bacterial capsule consists of an outer membrane, a peptidoglycan layer, and a cytoplasmic inner membrane. The outer leaflet of the outer membrane is predominantly composed of lipopolysaccharides (LPSs), while the inner leaflet comprises proteins and phospholipids. The phospholipid fraction contains proteins that primarily regulate the transport of nutrients and metabolic products and is dominated by phosphatidylethanolamine, accompanied by varying amounts of phosphatidylglycerol, cardiolipin, and phosphatidic acid. 3 Currently, the development of serogroup B N. meningitidis vaccines is a priority for controlling invasive meningococcal disease, yet the immunogenic design is hindered by the structural similarity between its capsule and human antigens. 4 With the advancement of vaccine technology, vaccine design based on molecular diagnostic data has become increasingly precise, enabling the formulation of effective immunization strategies targeting epidemic serogroups and their variants. 5 Maintaining the phenotypic and genetic characteristics of original strains via low-temperature preservation technology is critical to ensuring the stability and authenticity of strains required for vaccine development.
In current research, although cryopreservation is widely applied for sample storage, its specific effects on bacterial structure and antigen expression remain to be fully elucidated to optimize preservation conditions and guarantee the accuracy and efficacy of diagnostic assays and vaccine development. Glycerol or skim milk is commonly used as a cryoprotectant (CPA) for N. meningitidis preservation, 6 whereas a study by Laura et al. demonstrated that cotton swab impregnation storage at −80°C without additional CPAs also achieves favorable long-term preservation efficacy. 7 Although these established methods provide feasible approaches for strain preservation, their practical application is accompanied by notable limitations. For instance, skim milk, as a macromolecular CPA, is susceptible to brand and batch variations, resulting in poor reproducibility and an increased risk of exogenous microbial contamination. 8 The cotton swab impregnation method, while simple to operate, has a limited physical adsorption capacity, which makes it difficult to precisely control bacterial viability and distribution uniformity. This shortcoming renders it unable to meet the quantitative preservation requirements of standardized, highly reproducible living biobanks for vaccine development. Cryopreservation is currently the gold standard for long-term storage of microbial strains; however, ice crystal formation, osmotic stress, and chemical toxicity of CPAs during freezing may induce cell membrane damage, protein denaturation, and a significant reduction in bacterial viability. 9 Existing studies on N. meningitidis cryopreservation have obvious limitations: most focus on a single CPA or fixed concentration, lacking comparative analyses of different permeable CPAs in various base solutions. The underlying mechanisms by which key thermodynamic properties of CPAs affect cell survival remain unclear. In addition, the cytotoxicity of CPAs against N. meningitidis has not been fully quantified, which may lead to impaired strain activity or functional loss post-preservation. Importantly, these research gaps are not unique to N. meningitidis; systematic data on the cryoprotective efficacy, application status, and limitations of these four CPAs remain scarce in most fastidious bacteria, including Haemophilus influenzae and N. gonorrhoeae, and this knowledge gap underscores the urgency and innovation of establishing a dedicated CPA screening system for fastidious microorganisms, as performed in the present study.
To address these aforementioned issues, this study conducted a multidimensional analysis to evaluate the effects of four permeable CPAs at low, medium, and high concentrations with PBS and brain heart infusion (BHI) broth as base solutions on the survival rate of N. meningitidis after cryopreservation at −80°C and determine the cytotoxicity of each CPA at 37°C for 15 minutes. Additionally, this study also measured the crystallization temperature (Tc), crystallization enthalpy (ΔHc), melting temperature (Tm), and melting enthalpy (ΔHm) of permeable CPAs in different base solutions via differential scanning calorimetry (DSC) and dynamically monitored ice crystal growth area using cryomicroscopy. Collectively, the findings of this study can provide an optimized basis for the cryopreservation system of N. meningitidis and similar fastidious bacteria and offer specific technical references for the standardized construction, quality control, and long-term stable preservation of living biobanks, thereby supporting the high-quality development of related pathogenic research, diagnostic reagent development, and vaccine development.
Materials and Methods
Bacterial culture and identification
N. meningitidis (ATCC 43828) was obtained from the Respiratory Tract Pathogen Laboratory, Institute of Infectious Disease Prevention and Control, Chinese Center for Disease Control and Prevention. The bacterial strains were inoculated onto Columbia blood agar plates and incubated at 37°C for 12 hours for bacterial harvesting. Species identification was performed using the Analytical Profile Index (API) biochemical identification strips (bioMérieux, France).
Preparation of cryoprotectants
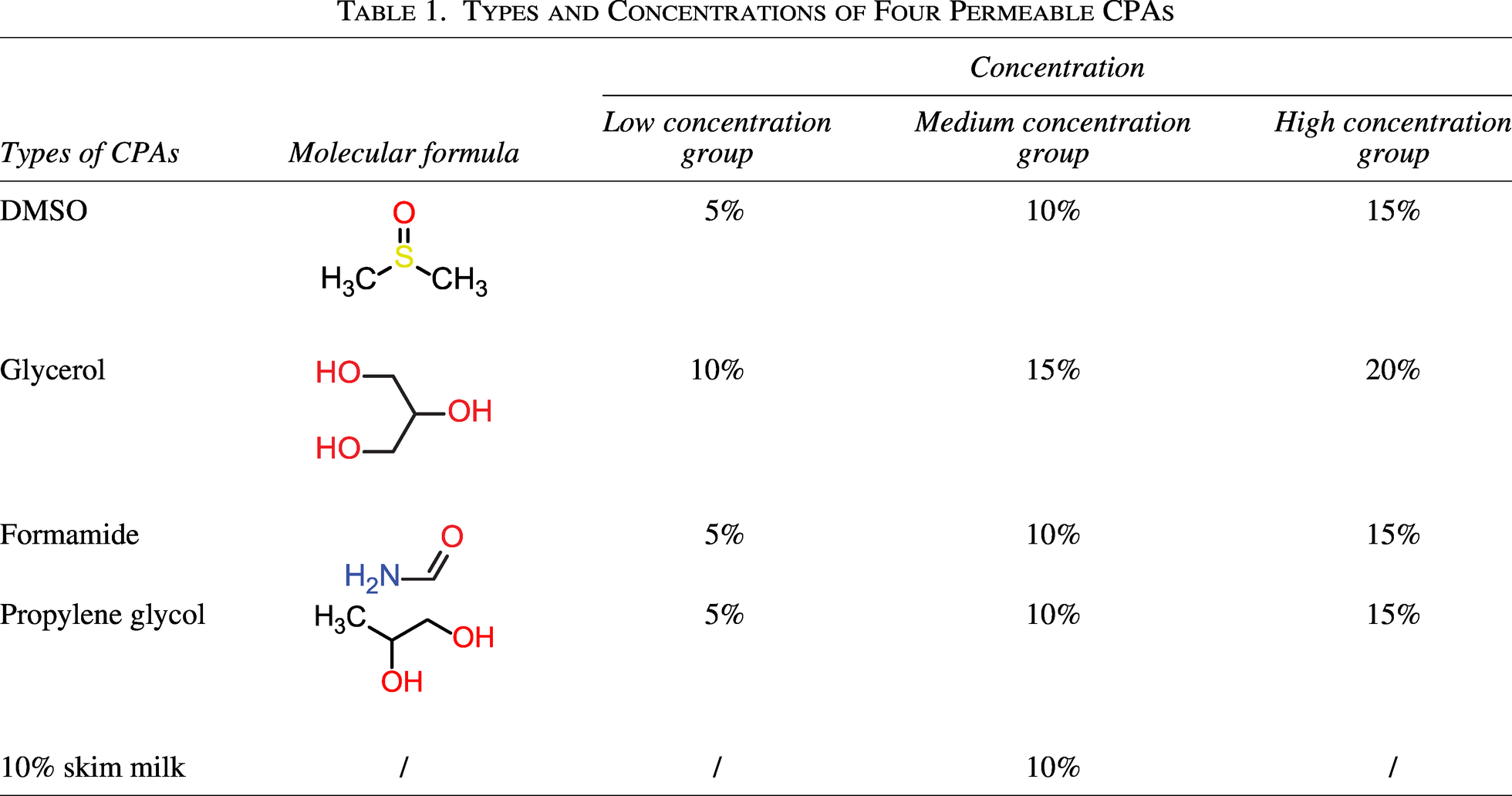
The types and concentrations of permeable CPAs used in this study are listed in Table 1. Glycerol, propylene glycol (PG), and BHI broth were sterilized by autoclaving at 121°C for 15 min, 10% skim milk was autoclaved at 121°C for 5 min, formamide (FMD) was sterilized by filtration through a 0.22 μm sterile filter membrane, dimethyl sulfoxide (DMSO) and PBS were purchased as sterile commercial products. All CPAs were prepared at twice the final working concentration as indicated in Table 1 for subsequent 1:1 (v/v) mixing with bacterial suspensions.10% skim milk was used as a positive control.
Types and Concentrations of Four Permeable CPAs
Cytotoxicity assay of permeable cryoprotectants
Sterilized CPA stock solutions were mixed with bacterial suspensions at a volume ratio of 1:1 (v/v), resulting in the final working concentrations of CPAs as specified in Table 1. The CPA-bacterial mixtures were transferred to cryovials (1 mL per vial) and incubated at 37°C for 15 min. All samples were subjected to gradient dilution with corresponding BHI broth or PBS before and after co-incubation, and the viable count was determined on Columbia blood agar plates to calculate the survival rate for cytotoxicity assessment. The survival rate was expressed as N1/N0 × 100%, where N0 and N1 represented the viable counts before and after the co-incubation process, respectively.
Assessment of N. meningitidis viability after cryopreservation at −80°C
The CPA-bacterial mixtures were placed into a metal block controlled-rate freezing container (cooling rate of 1°C/min) and cryopreserved at −80°C for 48 h. Cryovials were then rapidly removed from the freezer and thawed in a 37°C water bath with gentle shaking for 3–5 min until complete thawing. Thawed samples were gently mixed, subjected to serial gradient dilutions, and plated on Columbia blood agar for viable counting. The protective efficacy of each CPA was quantitatively evaluated based on the bacterial survival rate post-cryopreservation.
Differential scanning calorimetry experiment
DSC measurements were performed using a DSC 8500 differential scanning calorimeter (PerkinElmer, USA). To simulate the actual freezing and thawing processes of the experimental system, the temperature profiles of PBS and BHI broth were determined using a precision thermometer, and the results are presented in Supplementary Data. The optimized DSC program was set as follows: equilibrating at an initial temperature of 25°C for 1 min, cooling to −80°C at a rate of 1°C/min and holding for 2 min, then heating back to 25°C at a rate of 20°C/min and holding for 1 min to terminate the program. Crystallization temperature, crystallization enthalpy, melting temperature, and melting enthalpy were recorded and analyzed for each sample.
Cryomicroscopy observation
Cryomicroscopic analysis was conducted using a Linkam cryomicroscopy system (BSC196 Biological Cryo-stage, UK). A 2 μL aliquot of each CPA solution was placed on the cryo-stage and covered with a glass coverslip to ensure clear imaging during thermal cycling. The thermal program was executed as follows: cooling to −50°C at 20°C/min and holding for 2 min; heating to −12°C at 10°C/min and holding for 1 min; heating to −5°C at 0.5°C/min and holding for 1 min; further heating at 0.1°C/min until only a single ice nucleus remained in the field of view and holding for 1 min; finally cooling again at 0.1°C/min. Images were captured at 1 s intervals to monitor ice crystal growth, and the ice crystal area was quantified using ImageJ software.
Statistical analysis
All experimental data were analyzed using GraphPad Prism 11.0 software (GraphPad Software, USA). Each experiment was independently replicated three times, and data were expressed as mean ± standard deviation (SD). One-way ANOVA was performed for multi-group comparisons, with the Brown-Forsythe test confirming homogeneity of variance (p > 0.05). Tukey’s test was used for pairwise comparisons between groups, p < 0.05 was considered statistically significant.
Results
Strain identification results
N. meningitidis colonies grown on Columbia blood agar plates (Fig. 1a) exhibited a gray, non-hemolytic, round, convex, smooth, moist morphology with distinct edges. Biochemical identification via API strips yielded colorimetric results (Fig. 1b) that were consistent with the phenotypic characteristics of N. meningitidis, confirming the strain as N. meningitidis.

Identification of N. meningitidis.
Effects of permeable cryoprotectant types and concentrations on the cytotoxicity of N. meningitidis
Fig. 2 illustrates the cytotoxic effects of CPA types and concentrations (in PBS or BHI broth) on N. meningitidis. As shown in (Fig. 2a), the cytotoxic survival rate of all CPAs in PBS decreased gradually with increasing concentrations. Among these, 5% DMSO exhibited the lowest cytotoxicity with a survival rate of 73.35%, followed by 10% Gly (56.31%) and 5% FMD (60.52%) with relatively low cytotoxicity. The cytotoxic survival rate of 5% PG (43.28%) was significantly lower than that of 5% DMSO (p < 0.05). The cytotoxic survival rate of the PBS control group was only 4.51%.
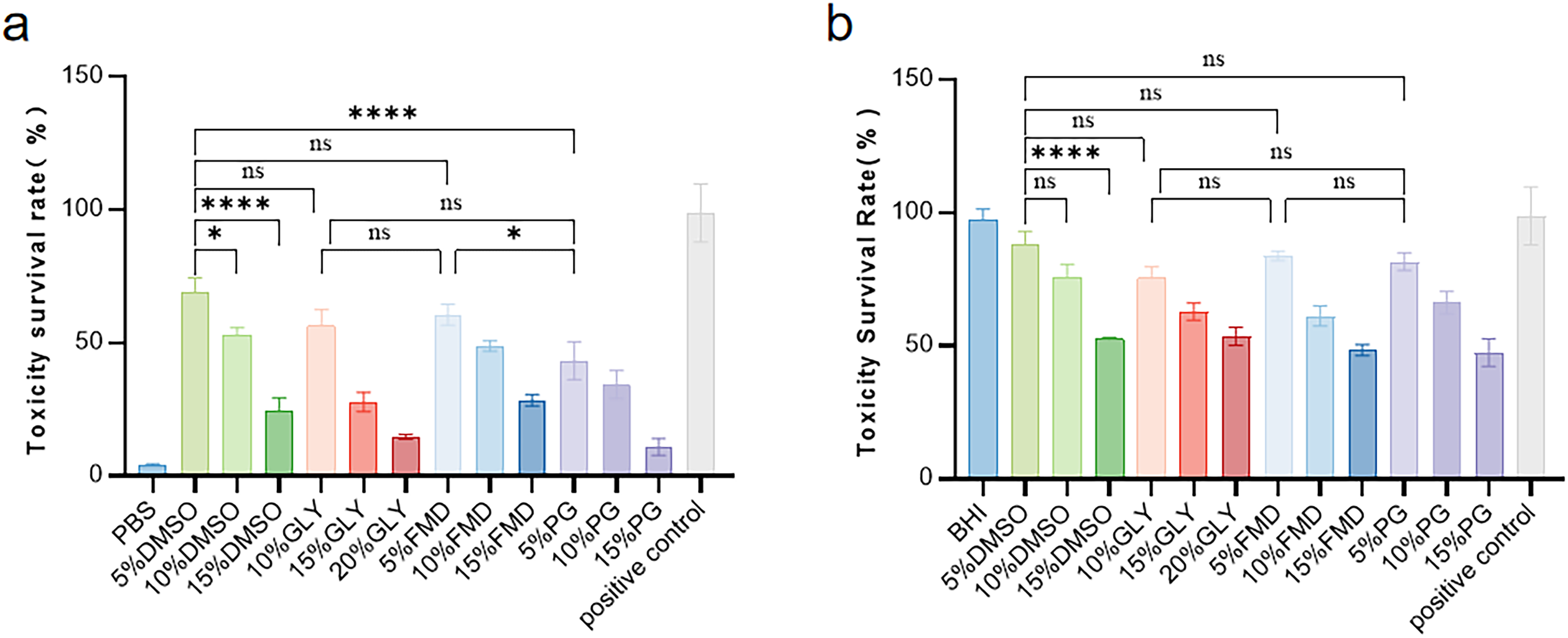
Toxicity of different protective agents on survival rate of N. meningitidis in different basic media.
A similar concentration-dependent cytotoxic trend was observed in BHI broth (Fig. 2b), with no significant differences among low-concentration permeable CPAs. 5% DMSO still showed the lowest cytotoxicity (88.14% survival rate), followed by 5% FMD (83.85%) and 5% PG (81.62%) with moderate cytotoxicity. 10% Gly presented the highest cytotoxicity with a survival rate of 75.25%, and the cytotoxic survival rate of the BHI control group was 94.0%.
Comparison of Fig. 2a and b revealed that the overall cytotoxic survival rate in BHI broth was higher than that in PBS, and positive control group exhibited a relatively high cytotoxic survival rate of 91.1%.
Effects of permeable cryoprotectant types and concentrations on N. meningitidis viability after cryopreservation
Fig. 3 analyzes the effects of CPA types and concentrations on N. meningitidis viability following 48 hours of cryopreservation at −80°C. In PBS (Fig. 3a), the survival rate of all permeable CPAs decreased with increasing concentrations, except for the FMD group. 5% DMSO achieved the highest survival rate of 64.93% (p < 0.05), followed by 10% Gly (45.69%) and 10% FMD (51.10%) with relatively high viability. The 5% PG group had the lowest survival rate (28.23%), and the PBS control group only reached a survival rate of 5.0%.
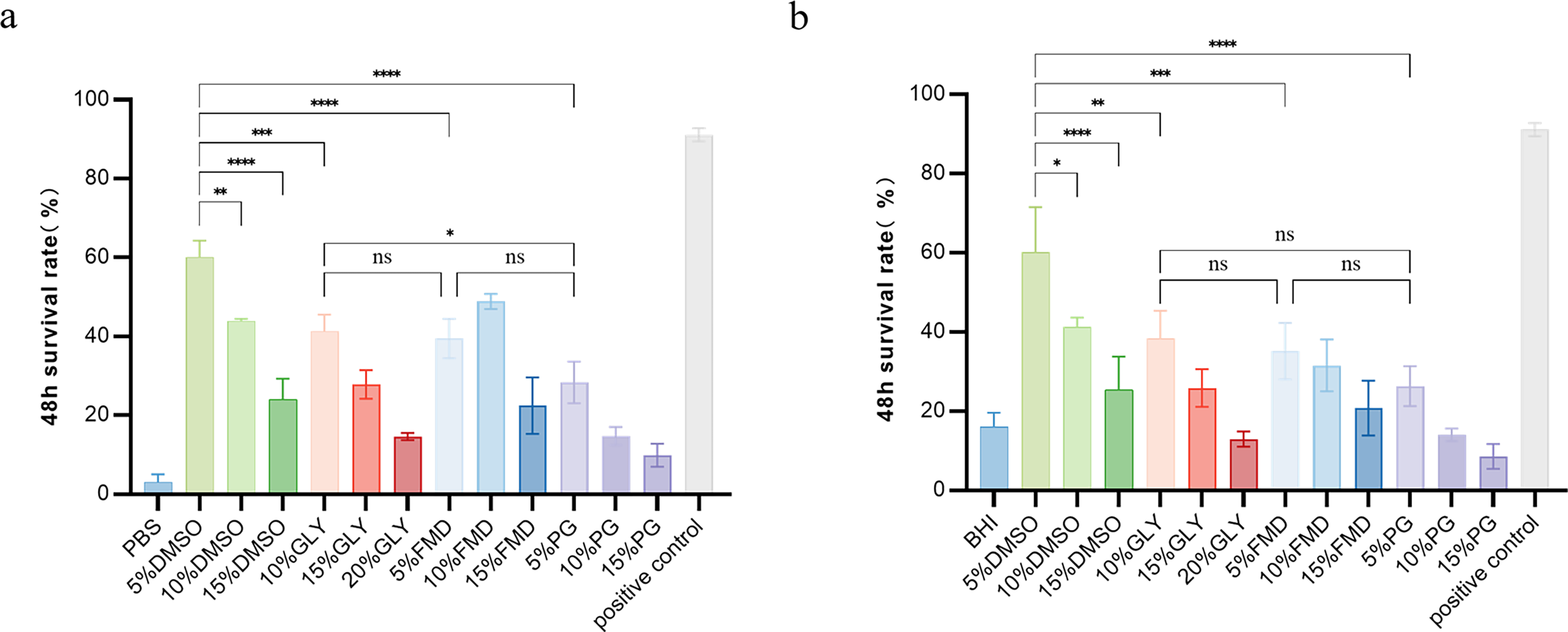
Survival rate of N. meningitidis after 48 hours frozen storage in different CPAs in different base solutions.
A consistent concentration-dependent trend was observed in BHI broth (Fig. 3b). 5% DMSO still maintained the highest survival rate (63.11%, p < 0.05) after 48 hours of cryopreservation, followed by 10% Gly (40.88%) and 5% FMD (42.09%). The 5% PG group showed the lowest survival rate (26.33%), and the survival rate of the BHI control group was 20.0%.
Comparison of Fig. 3a and b indicated no significant difference in survival rates between the two base solutions, and positive control group achieved a survival rate of 91.56%.
Analysis of thermophysical parameters of different permeable cryoprotectants and base solutions
In PBS, the Tc values of the four permeable CPAs were random with no obvious concentration-dependent trend (Fig. 4a). The ΔHc decreased gradually with increasing CPA concentrations (Fig. 4a). Among the four CPAs, 15% DMSO exhibited the lowest ΔHc (139.503 J/g), indicating the strongest ice crystal inhibition capacity during freezing. Additionally, 20% Gly (143.82 J/g) and 15% PG (145.42 J/g) showed relatively low ΔHc values. The Tm presented a regular increasing trend with rising CPA concentrations (Fig. 4c). For example, the Tm of 5% DMSO was −5.43°C, while that of 15% DMSO decreased to −13.2°C. The variation trend and magnitude of Tm for Gly and PG groups were consistent with DMSO. The ΔHm decreased with increasing CPA concentrations (Fig. 4c), and 20% Gly had the lowest ΔHm (145.755 J/g), followed by 15% DMSO (130.548 J/g) and 15% FMD (165.330 J/g).
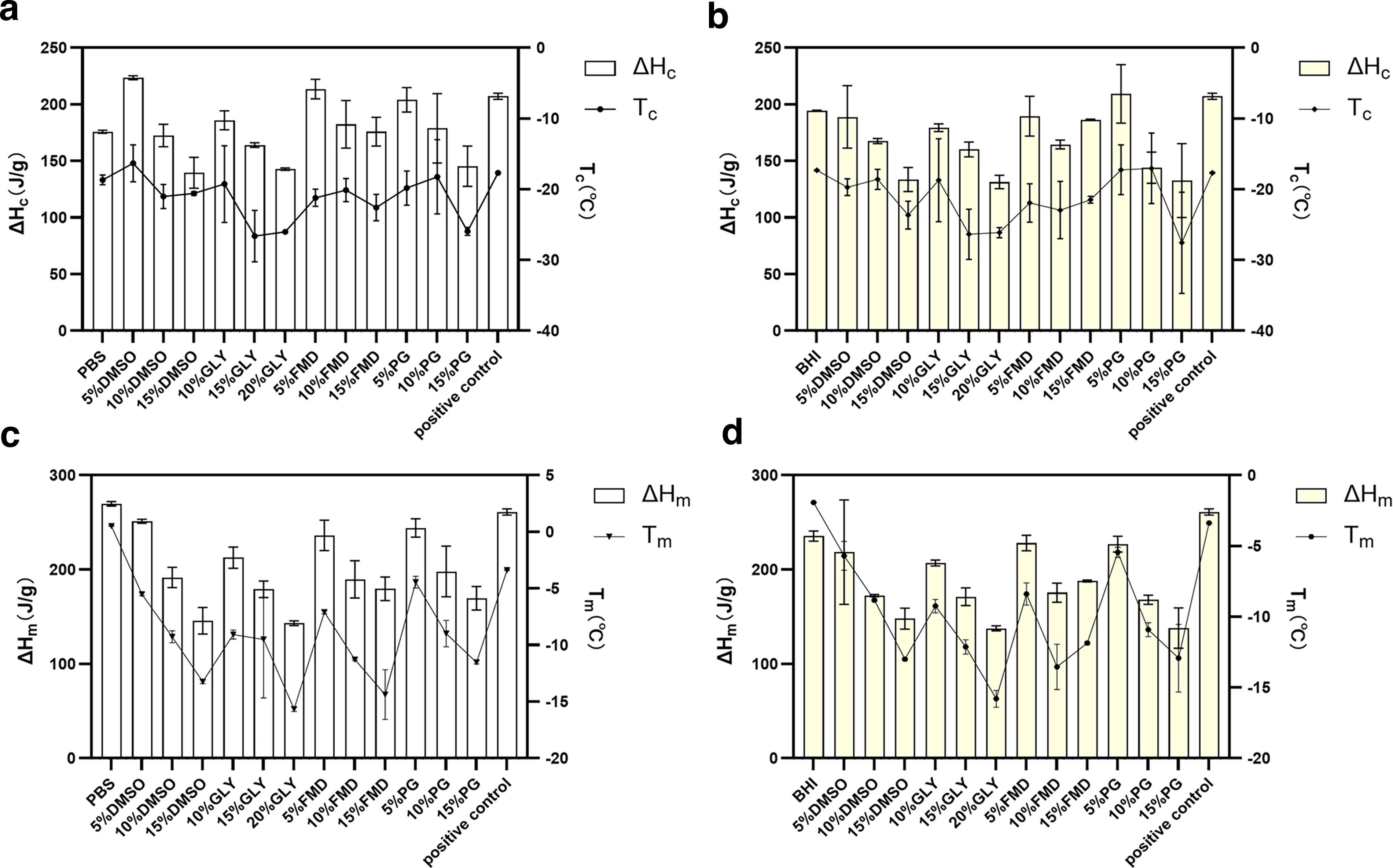
Key Parameters of Thermal Physical Properties of Protective Agents in Each Group.
In BHI broth, the Tc of CPAs was also random without a clear concentration trend (Fig. 4b), consistent with PBS. Except for FMD, the ΔHc of all CPAs decreased with increasing concentrations (Fig. 4b). 15% DMSO still had the lowest ΔHc (123.1335 J/g) with the strongest ice inhibition ability, followed by 20% Gly (138.328 J/g) and 15% PG (133.64 J/g). For Tm (Fig. 4d), all CPAs showed a regular increasing trend with rising concentrations except the FMD group. For instance, the Tm of 5% DMSO was −5.28°C, and that of 15% DMSO was −12.95°C; the variation trend of Tm for Gly and PG was similar to DMSO. The Tm values of 5%, 10%, and 15% FMD were −8.67°C, −12.62°C, and −11.76°C, respectively. The ΔHm of all CPAs decreased with increasing concentrations except the FMD group (Fig. 4d). 20% Gly exhibited the lowest ΔHm (134.77 J/g), followed by 15% DMSO (141.09 J/g) and 15% FMD (139.68 J/g).
In summary, permeable CPAs in both BHI broth and PBS exhibited consistent thermophysical trends. Based on key thermophysical parameters, the ice crystal inhibition capacity followed the order: high concentration > medium concentration > low concentration for DMSO, Gly, and PG groups, while for FMD it was medium concentration > high concentration > low concentration. The ice inhibition effect of BHI broth was slightly higher than that of PBS, with no significant difference between the two base solutions.
Analysis of single ice crystal growth in different permeable cryoprotectants and base solutions
The growth area of single ice crystals for each group of permeable CPAs in a PBS base solution from 0 s to 50 s is shown in Fig. 5a. In Fig. 5b, the growth area of 5% DMSO was approximately 921.57 µm2, 10% Gly was about 1140.03 µm2, 5% FMD was around 1317.33 µm2, and 5% PG was roughly 2432.94 µm2. Fig. 5c presents the measurement of single ice crystal growth area for each group of permeable CPAs in BHI base solution from 0 s to 50 s. In Fig. 5d, the growth area of 5% DMSO was about 814.20 µm2, 10% Gly was approximately 1027.86 µm2, 5% FMD was around 1043.96 µm2, and 5% PG was roughly 1631.56 µm2.
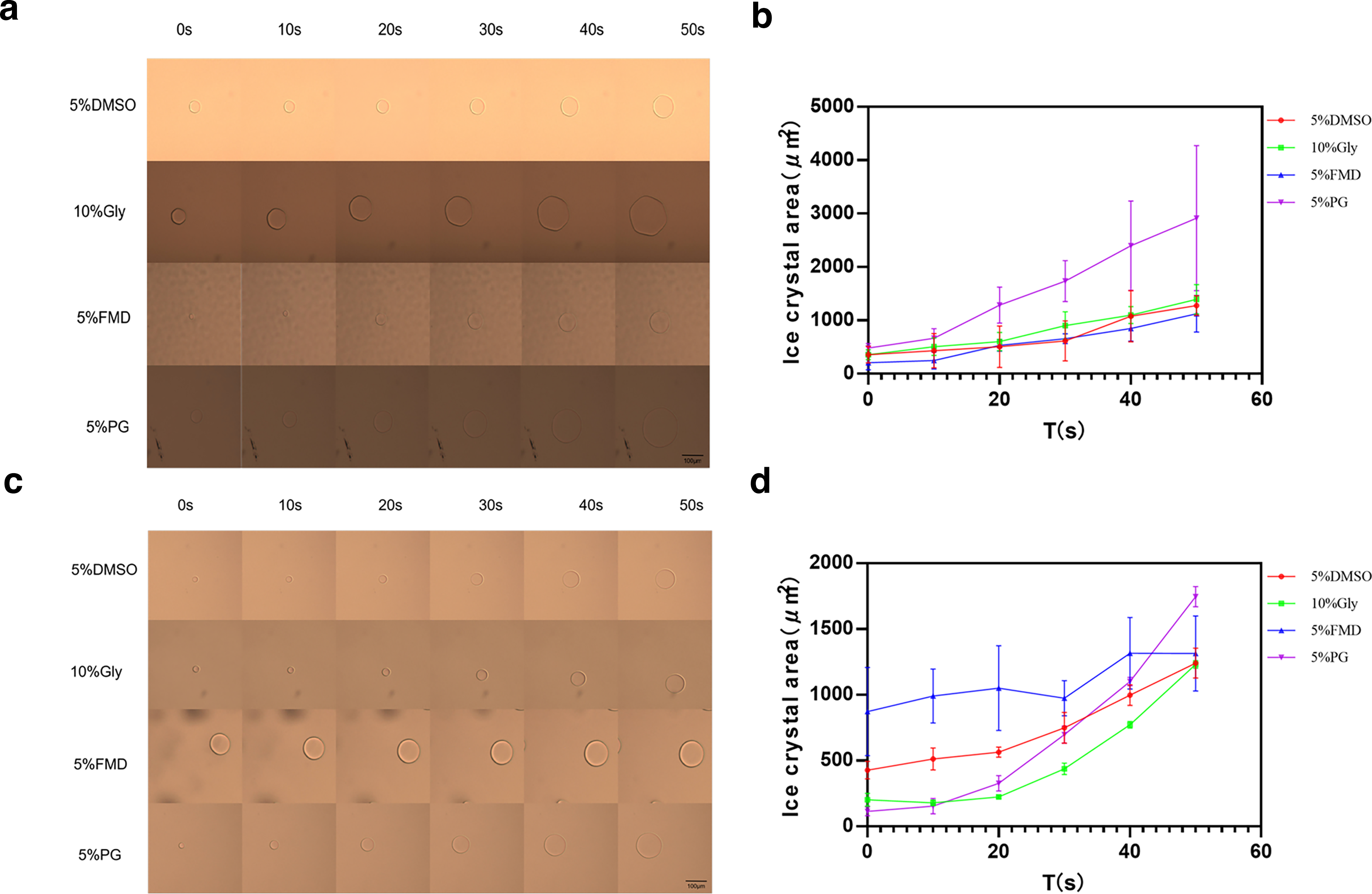
Single crystal growth area of each group of protective agent.
Therefore, 5% DMSO in BHI base solution exhibited the slowest ice crystal growth, followed by 5% DMSO in PBS base solution, and then in sequence by 10% Gly in BHI base solution, 5% FMD in BHI base solution, 10% Gly in PBS base solution, 5% FMD in PBS base solution, 5% PG in BHI base solution, and 5% PG in PBS base solution.
Discussion
Permeable CPAs feature low molecular weight and high water solubility, enabling rapid transmembrane penetration. They effectively mitigate cryoinjury by balancing intracellular and extracellular osmotic pressure, inhibiting intracellular ice crystal formation, and stabilizing protein tertiary structures. DMSO (C2H6OS), FMD (HCONH2), glycerol (C3H8O3), and PG (C3H8O2) were selected for systematic evaluation in this study, primarily because these four classic permeable CPAs have been extensively investigated in bacterial and mammalian cell cryopreservation, ensuring excellent comparability. While sharing similar characteristics of low molecular weight, high hydrophilicity, and rapid membrane permeability, they differ in chemical properties and protective mechanisms, which facilitates comparative optimization of cryoprotective efficacy. Structural differences in these CPAs determine their distinct ice-inhibiting potential. DMSO and FMD are small-molecule strong polar compounds: the sulfur-oxygen double bond (S = O) in DMSO forms robust hydrogen-bonded networks with water molecules, which disrupts the ordered arrangement of water and effectively inhibits ice crystal nucleation and growth. 10 Ding et al. 11 demonstrated that human bone marrow mesenchymal stem cells exhibited DNA damage and impaired cellular function after cryopreservation with 10% DMSO, whereas Renata et al. 12 reported that reducing DMSO concentration to 5% effectively preserved the multipotent differentiation potential and long-term engraftment capacity of hematopoietic stem cells while alleviating cytotoxicity. For FMD, its carbonyl (C = O) and amino (–NH2) groups enable the formation of stable hydration complexes, 13 and Diogo et al. 14 confirmed that 12.2% and 15% FMD improved zebrafish sperm quality and hatching rates post-thawing. In contrast, glycerol and PG belong to polyols with relatively larger molecular weights than DMSO and FMD and are rich in hydroxyl groups (–OH). Their protective mechanism mainly relies on enhancing solution viscosity via hydration, partially replacing water molecules to physically retard ice crystal growth kinetics. Ogata et al. 15 verified that 3% glycerol combined with optimized freezing/thawing rates improved canin DSC results in this study revealed that with increasing concentrations of permeable CPAs, the ΔHc and the ΔHm decreased progressively, accompanied by a regular reduction in Tm in both BHI broth and PBS. This indicated enhanced ice-inhibiting efficacy, which was consistent with the slowed ice crystal growth observed via cryomicroscopy and aligned with the findings of Katarína et al. 16 Notably, BHI broth exerted a superior cryoprotective effect compared with PBS, likely attributed to abundant proteins and carbohydrates in BHI that provided additional cryoprotection and further suppressed ice crystal formation during freezing.
The classic two-factor hypothesis of cryoprotection postulates that cells suffer two primary forms of damage during freezing: mechanical injury induced by ice crystal formation and solute osmotic damage caused by extracellular solution concentration.17,18 This study extended this framework to N. meningitidis and other fastidious bacteria, and our results demonstrated that CPA efficacy was not simply positively correlated with ice-inhibiting capacity. Although low-concentration DMSO did not exhibit the strongest ice-inhibiting activity in DSC assays, it yielded the highest bacterial survival rate post-cryopreservation. This finding suggested that alleviating solute osmotic damage is equally critical for N. meningitidis survival. DMSO and FMD, with rapid transmembrane penetration, rapidly equilibrated intracellular and extracellular osmotic pressure, thereby mitigating dehydration-induced damage in the early freezing phase. 19 In contrast, glycerol and PG penetrate cell membranes more slowly, which may enable milder water efflux during slow-rate freezing and confer better protection under specific conditions. 20 Thus, the optimal CPA concentration represents a delicate balance between ice inhibition requirements and the reduction of chemical damage from osmotic stress and cytotoxicity. In BHI broth, 5% DMSO achieved the optimal balance between ice inhibition and biocompatibility whereas PBS, lacking organic buffering components, exacerbated CPA cytotoxicity and consequently led to reduced bacterial survival rates compared with BHI.
N. meningitidis and other Gram-negative fastidious bacteria possess a unique outer membrane rich in LPS and porins, and the presence of a polysaccharide capsule further increases cell surface complexity. 21 Low temperatures disrupt membrane lipid fluidity, trigger lipid phase transition and membrane rupture. The differential protective efficacy of CPAs in this study may stem from their specific interactions with the bacterial outer membrane. Andrey et al. 22 reported that DMSO enhances membrane permeability to hydrophilic molecules and cryoprotective activity via pore formation in the outer membrane. Glycerol and PG, by virtue of their strong hydration capacity, 23 stabilize membrane protein conformations and maintain the pericellular hydration layer, thereby preventing excessive dehydration and membrane denaturation. In BHI broth, nutrient components may facilitate membrane stabilization and mitigate CPA-induced membrane perturbation, thereby endowing low-concentration DMSO with superior protective efficacy. In contrast, high concentrations of CPAs are prone to impairing membrane integrity and elevating the risk of autolysis.
Beyond ice inhibition and membrane protection, additional mechanisms including oxidative stress mitigation and metabolic regulation may contribute to CPA-mediated cryoprotection. Reactive oxygen species (ROS) accumulate during freezing/thawing cycles, inducing lipid peroxidation and protein oxidative damage, 24 DMSO and glycerol may scavenge free radicals and activate cellular antioxidant defense systems via their intrinsic antioxidant properties, thereby alleviating oxidative damage.25,26 Moreover, CPAs may regulate bacterial metabolic pathways and affect post-thaw recovery capacity. The differential protective effects of BHI and PBS are likely associated with metabolic substrate availability, BHI provides essential nutrients to enhance bacterial stress resistance, while PBS limits metabolic recovery post-thawing. Future studies should quantify intracellular ROS levels and antioxidant enzyme activities to further elucidate the oxidative stress-related protective mechanisms of CPAs in N. meningitidis cryopreservation.
The significance of this study goes beyond the screening of optimal CPAs for N. meningitidis, more importantly, it provides a theoretical basis and experimental reference for the cryopreservation of fastidious bacteria and other hard-to-culture microorganisms. It should be noted that this study was conducted using only a single standard strain (ATCC 43828). Future validation with clinical isolates will be crucial to further confirm the generalizability of the current findings. Establishing an efficient and stable living microbial biobank is a critical infrastructure for safeguarding microbial biodiversity, preserving clinical strain resources, advancing vaccine development, and supporting basic microbiological research. In addition, it will be necessary to explore the stability of the optimized CPA protocol during long-term preservation and compare its efficacy with that of liquid nitrogen storage, so as to further enhance the practical application value of this study. By elucidating and optimizing cryoprotective strategies from molecular mechanisms to cellular effects, we can improve the long-term viability and genetic stability of precious microbial strains, thereby laying a solid foundation for the sustainable preservation and utilization of microbial resources.
Conclusion
5% DMSO in BHI broth is the optimal permeable CPA for N. meningitidis and other fastidious bacteria. Its potential mechanism is that 5% DMSO balances bacterial osmotic pressure, alleviates solute and ice crystal damage, and balances ice inhibition with biocompatibility to enhance bacterial survival.
Footnotes
Acknowledgments
No competing financial interests exist.
Authors’ Contributions
H.J.: Methodology, investigation, visualization, and writing—original draft. N.G.: Methodology, investigation, visualization, and writing—original draft. L.X.: Investigation and formal analysis. Y.L.: Formal analysis and methodology. G.L.: Investigation and methodology. J.X.: Methodology and formal analysis. Y.X.: Conceptualization, methodology, and writing—review and editing. Y.X.: Re-sources and writing—review and editing. Z.S.: Conceptualization, project administration, and writing—review and editing. Q.W.: Conceptualization, project administration, and writing—review and editing.
Compliance with Ethical Standards
This article does not contain any studies with human participants or animals performed by any of the authors.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by
Supplemental Material
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.