
Abstract
Introduction:
Rhea americana are ecologically important, as they disperse seeds and participate in the food chain of various predators in South America. However, populations are declining in the wild.
Objectives:
The objective was to characterize the resilience of greater rhea spermatozoa to cryopreservation by assessing a commercial semen extender supplemented with different concentrations of dimethyl sulfoxide (DMSO).
Methods:
Spermatozoa from the vas deferens of six mature males were evaluated for kinetic parameters, viability, membrane functionality, and morphology. Samples were first diluted in OvodylTM plus 6%, 10%, 14%, or 18% (v/v) DMSO, loaded in 0.25-mL straws frozen, and plunged in liquid nitrogen. After one week, samples were thawed and reevaluated. Sperm resilience to freezing temperatures was analyzed through preservation efficiencies (percentage of postfreezing values relative to the fresh ones).
Results:
Marked interindividual variability was observed, particularly in motility-related parameters. Fresh sperm motility ranged from 15.5% to 61.0%, while postthaw values did not exceed 11%. Similarly, viability ranged from 59% to 71% in fresh samples and from 3% to 26% postthaw. Membrane functionality varied from 64% to 95% in fresh samples compared with 7%–43% postthaw. Sperm morphology ranged from 66% to 86% in fresh samples. Coefficient of variation analysis confirmed high variability for motility parameters, whereas other variables showed moderate to low dispersion, indicating a heterogeneous response among individuals to the cryopreservation procedure. There was no difference (p < 0.05) between the different DMSO concentrations evaluated. The average efficiency of preserving total motility, progressive motility, viability, and membrane functionality in 14% DMSO was 18.3 ± 11.6%, 47.0 ± 28.5%, 21.0 ± 5.9%, and 27.1 ± 11.6%, respectively.
Conclusion:
DMSO concentrations did not influence postthaw sperm quality. However, marked interindividual variation, particularly in motility-related parameters, played a key role in sperm resilience to cryopreservation.
Introduction
Preserving and maintaining repositories of biological samples of DNA, somatic cells, tissues, gametes, and embryos, as well as their associated metadata enhance strategies for the protection of endangered species. 1 For avian species, semen conservation is the most widely used method for ex situ in vitro conservation of avian genetic resources and preservation of rare breeds, as it is the only noninvasive and economically viable technique available. 2
Greater rheas (Rhea americana) are birds of ecological importance in South America, particularly as seed dispersers and as part of trophic interactions with large predators. 3 In some Latin American countries, they are also raised for the production of meat, leather, and feathers. 4 Despite their importance, rheas are currently classified as near threatened, and population declines may have ecological consequences in the ecosystems they inhabit. 5 Thus, it is necessary to apply strategies aimed at the conservation of this species, among which the cryopreservation of male gametes represents a promising tool for future genetic management via assisted breeding.
Several factors are important in sperm cryopreservation protocols: the choice of the appropriate extender; the selection of cryoprotectants and their ideal concentrations; and the optimization of freezing and thawing rates. This guarantees good postthawing results and may vary according to the species. 2 Extenders should help maintain pH, osmolarity, provide sperm with energy, and prevent clumping by lowering sperm concentration. 6 In birds, commercial semen extenders are commonly used. OvodylTM is a good example that contains elements providing a favorable environment for the cell. 7 It has been used for sperm preservation in domestic species, such as the turkey (Meleagris gallopavo),8,9 as well as on wild species such as the black grouse (Tetrao tetrix) or the capercaillie (Tetrao urogallus). 10
Dimethyl sulfoxide (DMSO) is an intracellular cryoprotectant that has been used for semen cryopreservation in various wild bird species such as the Ring penguin (Aptenodytes patagonicus), 11 the whooping crane (Grus americana), 12 the harpy eagle (Harpia harpyja), 13 or the ostrich (Struthio camelus), another species from the ratite group. 14 Its use has been reported in different concentrations (4%, 8%, 12%, and 16% v/v) with promising results. 14 Regarding cryopreservation methods, a wide variety of freezing rates has been reported for avian semen, including slow, fast, and ultra-fast protocols. In emus (Dromaius novaehollandiae), a freezing protocol included 30 minutes of equilibration at 5°C, followed by a cooling rate of −11°C/min from 5°C to –140°C, which yielded promising results. 15 Since emus and rheas are phylogenetically closely related, as both belong to the ratite group, adopting the same freezing rate could be a viable alternative for the latter.
Despite these advances, avian sperm preservation remains challenging, particularly in ratites, where reduced sperm motility and variable sperm quality have frequently been reported. 16 These limitations highlight the species-specific sensitivity of avian sperm to cryoinjury and reinforce the need for optimization of cryoprotectant concentrations and freezing protocols. Given the lack of information on sperm cryopreservation in greater rheas, we hypothesized that DMSO concentration would influence postthaw sperm quality parameters, and that individual variation among males could affect sperm resilience to cryopreservation. Therefore, the objective of this study was to evaluate the effects of different DMSO concentrations (6%, 10%, 14%, and 18%) in a commercial extender (OvodylTM) on postthaw sperm quality, as well as to assess individual variability in response to the cryopreservation process.
Methods
All animal experiments comply with the Animal Research: Reporting of In Vivo Experiments guidelines. All experimental procedures were performed in accordance with ethical standards of the National Institutes of Health guide for the care and use of laboratory animals (NIH Publications No. 8023, revised 1978), as they are in accordance with the Brazilian National Council for the Control of Animal Experimentation. Therefore, experiments were approved by the Ethics Committee on the Use of Animals of the Federal Rural University of the Semi-Arid Region (UFERSA; Process no. 23091.001423/2020-8) and the Chico Mendes Institute for Biodiversity Conservation (ICMBio; no. 73638-1).
The reagents used in this study were obtained from Sigma-Aldrich (St. Louis, MO), except when reported.
Animals
The animals used for the study were obtained from the Center for the Multiplication of Wild Animals of our institution (CEMAS/UFERSA; Scientific register N° 12.492-0004/IBAMA), located at 5° 11′S and 37° 20′W, at an average altitude of 16 m in a region characterized by a semiarid tropical climate, presenting Köppen climatic classification as dry and very hot, with an average annual temperature of 27.4°C.
The rhea paddocks had a dimension of 20.0 × 10.0 m. The rheas were fed with concentrated feed based on corn, wheat bran, and soybean, fruits were also provided, and water was provided ad libitum daily. Annually, a small group of individuals at CEMAS/UFERSA is culled for population control and used for research. Six adult rheas in reproductive age, 3.1 ± 0.6 years old, and with an average weight of 23.9 ± 1.2 kg, were part of the study.
Sperm collections took place during the 2021, 2022, and 2023 breeding seasons (two animals per year). Animals were captured and restrained manually. After being weighed, males were sedated before euthanasia following the protocol of Pollock et al. 17 After mechanical restraint, animals were weighed and subjected to anesthesia, with a combination of preanesthetic medication: Xylazine Hydrochloride (Xilazin® 2% at a dose of 1 mg/kg, manufacturer Syntec, Tamboré, São Paulo, Brazil) and Ketamine Hydrochloride (Ketamine® 10% at a dose of 15 mg/kg, Vetnil, Louveira, São Paulo, Brazil) administered intramuscularly. Once sedation was achieved, animals received an anesthetic dose of Thiopental (Thiopanthal sodium® 1 g at a dose of 150 mg/kg, Cristalia, São Paulo, Brazil) as an induction drug. Once the anesthetic plane was achieved, animals were euthanized by administering potassium chloride (Potassium chloride® 19.1% at a dose of 2.56 mEq/kg, Equiplex, Goiânia, GO, Brazil) intracardially. Animal’s death was confirmed by cardiorespiratory arrest. All organs and carcasses were utilized for morphological studies. Remaining biological material was subsequently disposed of by incineration through a licensed third-party company contracted by the university, in accordance with institutional and environmental regulations.
Sperm recovery and initial analysis
The vas deferens were collected, wrapped in gauze moistened with 0.9% saline solution at 38°C, and transported to the laboratory within 10 minutes for processing. Subsequently, the vas deferens were dissected from excessive adjacent tissue and washed externally with saline solution at 38°C. A flotation technique was used to recover the spermatozoa. Briefly, the ducts were separated and sliced with a scalpel in a Petri dish containing 2-mL saline solution at 38°C. After 5 minutes in a static position, tissues were removed and the sperm suspension evaluated. 18
The recovered volume was measured using pipettes and graduated tubes. A 5-μL aliquot of the sperm suspension was diluted in 995 μL of 10% buffered formalin to estimate the concentration using a Neubauer counting chamber visualized under light microscopy (400×) (Nikon Eclipse E200, Nikon Instrument, Tokyo, Japan). The total number of recovered sperm cells was then calculated by multiplying the sperm volume and concentration. 18
Each collected sample was divided into equal aliquots and assigned to different experimental treatments (6%, 10%, 14%, or 18% DMSO), allowing direct within-individual comparisons among treatments.
Computer-assisted semen analysis
Sperm motility parameters were analyzed by computer-assisted sperm evaluation (IVOS 7.4 G; Hamilton-Thorne Research, Beverly, MA, USA) according to the settings previously established for the species by our team. 19 Temperature was set at 37°C. Thirty frames were acquired at a frame rate of 60 Hz, minimum contrast of 15, and minimum cell size of 9 pixels. The straightness (STR) threshold was set at 80%, and the average path velocity (VAP) cutoff was established at 10 µm/s. Additional settings included cell size of 9 pixels and cell intensity of 19. Static cell detection parameters were defined as head size ranging from 0.57 to 8.80 µm2, head intensity from 0.10 to 3.08 (arbitrary units), and elongation from 4% to 98%. Analyses were conducted at a magnification factor of 1.89 (dimensionless). A 3-μL of sperm suspension was placed in the chamber of a Leja slide (IMV Technologies, I’Aigle, France). Once the sample drift was stabilized, five independent and nonconsecutive microscopic fields were randomly selected and scanned. The following motility characteristics were determined: total motility (MOT; in %), progressive motility (PMOT; in %), path velocity (VAP; in μm/s), progressive velocity (VSL; in μm/s), curvilinear velocity (VCL; in μm/s), lateral amplitude (ALH; in μm), beat frequency (BCF; in Hz), straightness (STR; in %), linearity (Lin; in %), and velocity distribution (rapid, medium, slow and static—in %).
Assessment of sperm viability
To assess the integrity of the sperm plasma membrane, the methodology previously validated for rheas by Bezerra et al. 19 was followed. A 10-μL aliquot from each sample was incubated at 37°C for 10 minutes with 2 μL Hoechst 33342 (H342, Molecular Probes, Eugene, OR, U.S.A) diluted at 25 mg/mL in DMSO. Subsequently, 3 μL of propidium iodide (PI) (Thermo Fisher Scientific, Waltham, MA, USA) was added to each sample. The PI working solution (2 mg/mL) was prepared from a 25-mg/mL stock solution (25 mg PI dissolved in 1 mL DMSO; Sigma-Aldrich, Burlington, MA, USA) by diluting 80 μL of stock solution in 920 μL of phosphate-buffered saline (Sigma-Aldrich, Burlington, MA, USA). Samples were then incubated at 37°C for 8 minutes. Samples were observed with an epifluorescence microscope (Olympus B × 51TF, Tokyo, Japan), with a 100 W mercury discharge burner (U-LH100HG) as the fluorescence light source, and analyzed under a yellow filter U-FYW (540–585 nm) by counting 100 cells per sample. Spermatozoa with blue fluorescence (H342) were classified as viable, and those with red fluorescence (PI) were classified as dead.
Hypo-osmotic swelling test
Aliquots (5 μL) of the samples containing the sperm were added to 45 μL of distilled water (0 mOsm/l) and incubated at 37°C for 40 minutes to validate the HOST. After incubation, the sperm were observed under phase-contrast microscopy (×1000 magnification). A total of 100 sperm were counted in at least five fields and classified as reactive or nonreactive, based on the presence or absence of curled (swollen) tails, respectively. 19
Sperm morphology evaluation
To analyze sperm morphology, 5 μL of sperm samples were fixed and stained with 45 μL formaldehyde-Bengal rose solution (20 mL of distilled water; 0.58 g sodium citrate; 0.8 mL formaldehyde; 0.3 g pf Bengal Rose; CAQ Casa da Química, São Paulo-SP, Brazil) on a slide. Two hundred sperm were evaluated using light microscopy (×1000 magnification), and per slide were observed. Sperm morphological defects were classified according to the method previously reported by Bezerra et al. 18
Semen cryopreservation
Samples were frozen and thawed following the freezing and thawing rate proposed by Sood et al. 15 for emus (D. novaehollandiae). Briefly, samples were diluted in Ovodyl™ (IMV technologies, I’Aigle, France) at 5°C. Then, they were equilibrated for 30 minutes at 5°C and divided into four aliquots. Subsequently, a second dilution was performed with the diluent containing DMSO (Dinâmica Química Contemporânea LTDA, Indaiatuba, SP, Brazil) at four different final concentrations as 6%, 10%, 14%, and 18% (v/v), reaching a final concentration of 75 × 106 sperm/mL. Such cryoprotectant concentrations are within the range previously described for other ratite bird, the emu. 15 After adding the cryoprotectants, the samples were loaded into 0.25 mL straws (IMV Technologies, L’Aigle, France) and remained for another 5 minutes at 5°C. Finally, the straws were placed 7.5 cm above the nitrogen vapor at −140°C in a Styrofoam box (39 × 32 × 29 cm) for 15 minutes to then be immersed in liquid nitrogen and placed in cryogenic cylinders for storage. The cooling rate from 5°C to −140°C was estimated to be approximately −11°C/min, based on the freezing conditions described by Sood et al. 15 for emus and the positioning of straws in nitrogen vapor. After one week of storage, samples were thawed at 5°C for 40 seconds and evaluated as mentioned for fresh samples.
Sperm preservation efficiency
Sperm resilience to freezing temperature was further evaluated. The efficiency of cryopreservation was evaluated based on total motility, progressive motility, viability, and membrane functionality before and after freezing. For this purpose, the following formulas were used: (percentage of total motility after thawing/percentage of total motility before freezing) × 100; (percentage of progressive motility after thawing/percentage of progressive motility before freezing) × 100; (percentage of viable spermatozoa after thawing/percentage of viable spermatozoa before freezing) × 100; and (percentage of functional spermatozoa after thawing/percentage of functional spermatozoa before freezing) × 100. 20
Statistical analysis
Data were expressed as mean ± standard error of means (SEM). Normality was assessed using the Shapiro–Wilk test, and homoscedasticity was evaluated using Levene’s test. When necessary, data were transformed using the arcsine method. Comparisons between groups with normally distributed and homogeneous data were performed using analysis of variance, followed by Fisher’s test. For variables that remained heterogeneous even after arcsine transformation (ALH, BCF, STR, LIN, medium, broken tail, tailless sperm), statistical differences were determined using the nonparametric Kruskal–Wallis test. Results were considered statistically significant when p < 0.05. All analyses were performed using Jamovi software, version 2.3.19 (Sydney, Australia). Interindividual variability was assessed using the coefficient of variation (CV), calculated as (standard deviation/mean) × 100. 21
Results
Fresh semen
The number of sperm recovered ranged from 176.0 × 106 (Animal 3) to 539.0 × 106 (Animal 5). A mean of 348.0 ± 64.5 × 106 spermatozoa was recovered. Fresh semen computer-assisted semen analysis parameters are presented in Figure 1A–I, showing clear interindividual variation among males. Total and progressive motility varied between individuals, from 15.5% (Animal 1) to 61.0% (Animal 5) (Fig. 1A), with an average of 30.2 ± 8.0% (Table 1). Animal 2 exhibited a progressive motility of just 1% while the highest progressive motility was 21% in Animal 6 in the fresh sperm samples (Fig. 1B). Similar patterns were observed for velocity parameters (VAP, VSL, and VCL), indicating differences in sperm kinetic performance among males. The VAP of the fresh samples ranged from 57.4 μm/s in Animal 3 to 98.1 μm/s in Animal 6 (Fig. 1C). The VSL values ranged from 45.6 μm/s in Animal 3 to 76.8 μm/s in Animal 6 (Fig. 1D). Parameters such as straightness (STR) and linearity (LIN) showed lower dispersion, suggesting more consistent sperm trajectory patterns across individuals The highest Straightness and Linearity were observed in Animal 4 (82.0% and 52.0%, respectively) (Fig. 1H and I).
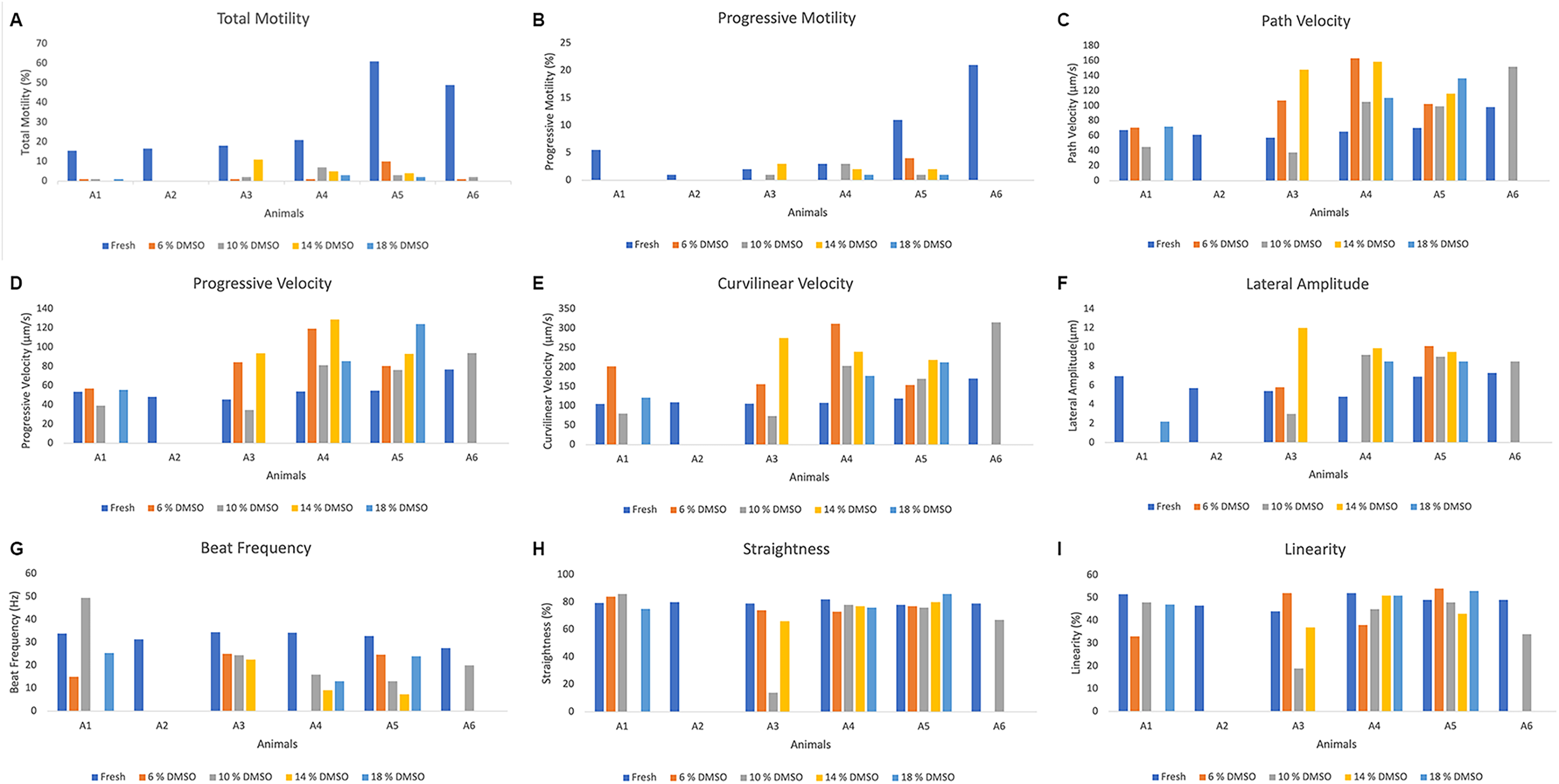
Individual values of sperm kinetic characteristics in greater rheas (Rhea americana; n = 6 animals) before and after cryopreservation in OvodylTM extender supplemented with different concentrations of dimethyl sulfoxide (DMSO).
Average Values (Mean ± SEM) of Fresh Sperm Characteristics in Greater Rheas (Rhea americana, n = 6 Animals)
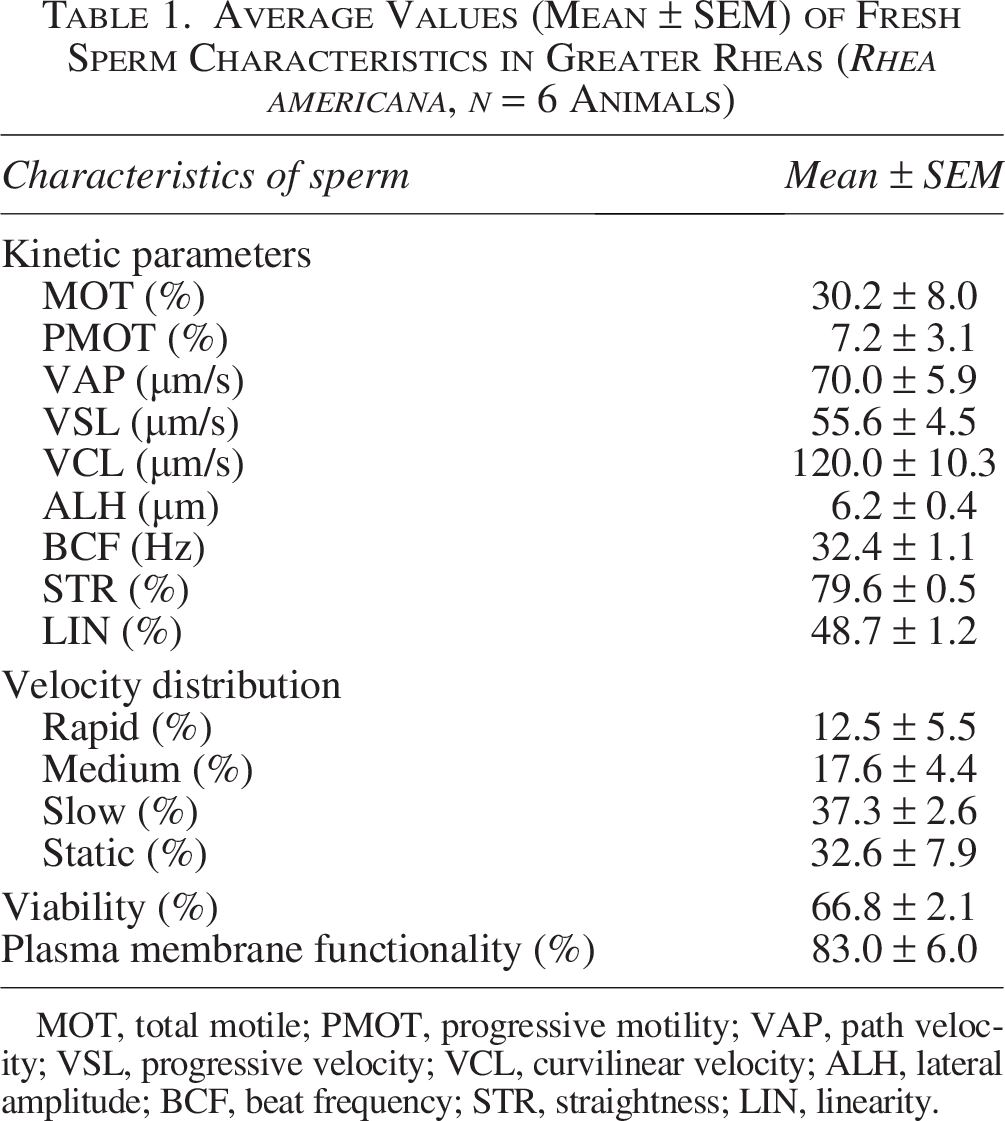
MOT, total motile; PMOT, progressive motility; VAP, path velocity; VSL, progressive velocity; VCL, curvilinear velocity; ALH, lateral amplitude; BCF, beat frequency; STR, straightness; LIN, linearity.
Sperm subpopulations in fresh samples are presented in Figure 2A–D, revealing a marked interindividual variation. The proportion of fast spermatozoa varied among individuals; the highest percentage of rapid sperm was present in Animal 6 (36%), while Animal 3 had only 3% (Fig. 2A). The highest percentage of medium spermatozoa (Fig. 2B) was in Animal 5 (39%); however, in the other animals, this percentage did not exceed 16%. Slow and static spermatozoa subpopulations were consistently predominant in all individuals, representing the largest fraction of spermatozoa. The percentage of slow spermatozoa (Fig. 2C) was greater than 30% in all animals, with Animal 2 having the highest percentage (49%). The lowest percentages of static spermatozoa (Fig. 2D) were 4% in Animal 5 and 15% in Animal 6; in the other animals, this value ranged between 34% and 54%.
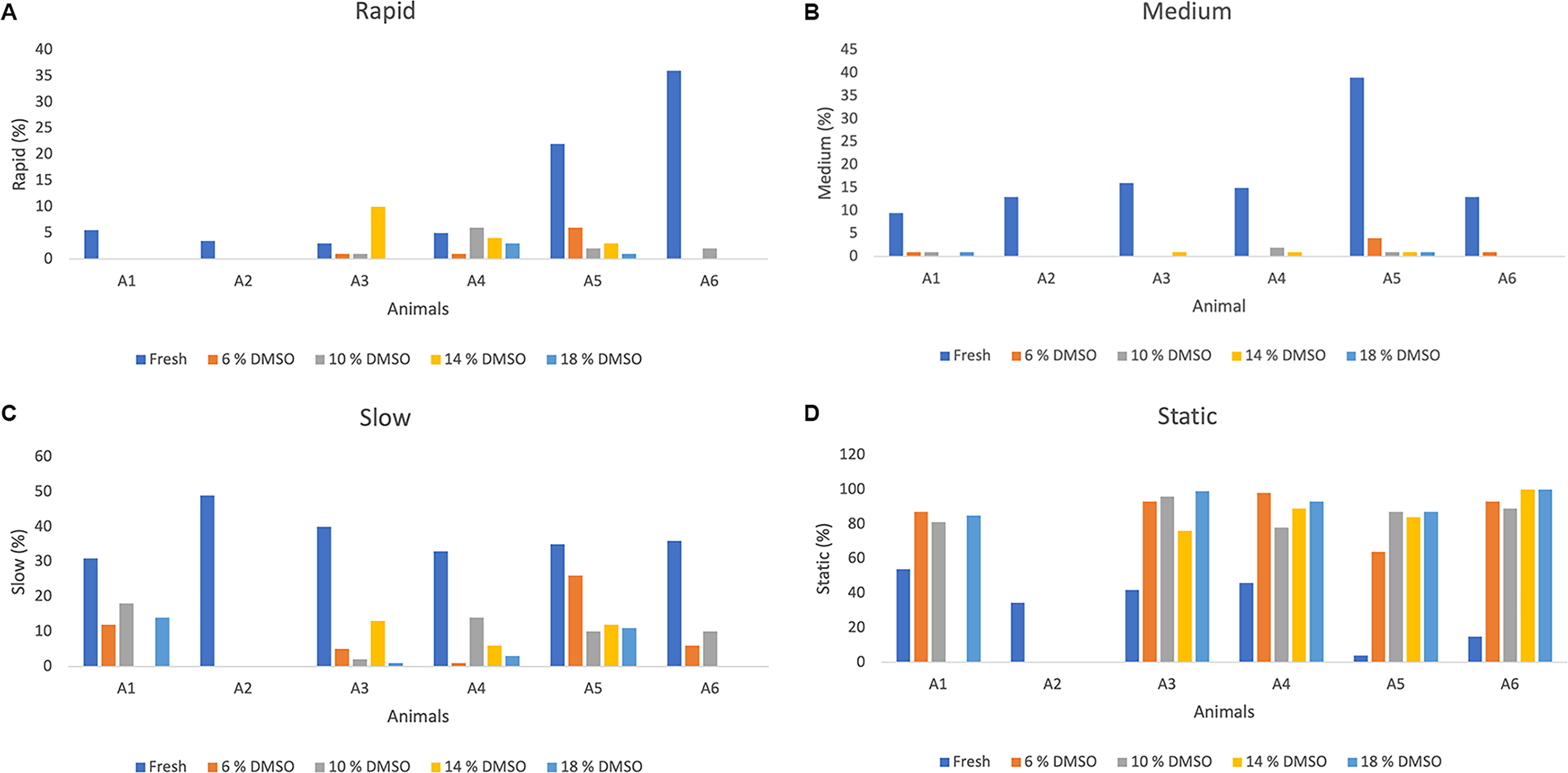
Distribution of sperm velocity (%) in greater rheas (Rhea americana; n = 6 animals) before and after cryopreservation in OvodyTM extender supplemented with different concentrations of dimethyl sulfoxide (DMSO) and evaluated through computerized semen analysis (CASA).
Viable sperm were labeled with blue fluorescence (Fig. 3A), and dead sperm were labeled in red fluorescence (Fig. 3B). Overall, viability values were high, with several individuals exceeding 70% (Animals 2, 4, and 5) (Fig. 3C), and the total average was 66.8 ± 2.1% (Table 1), indicating good sperm quality in fresh samples. Membrane functionality also showed consistently high values among individuals, although some variation was observed. The highest membrane functionality was observed in Animal 5 (95%) (Fig. 4), the lowest was 64.0% in Animal 1 (Fig. 4), and the total average was 83.0 ± 6.1% (Table 1). Both viability and membrane functionality exhibited less interindividual variability, suggesting that these parameters were more stable among males.
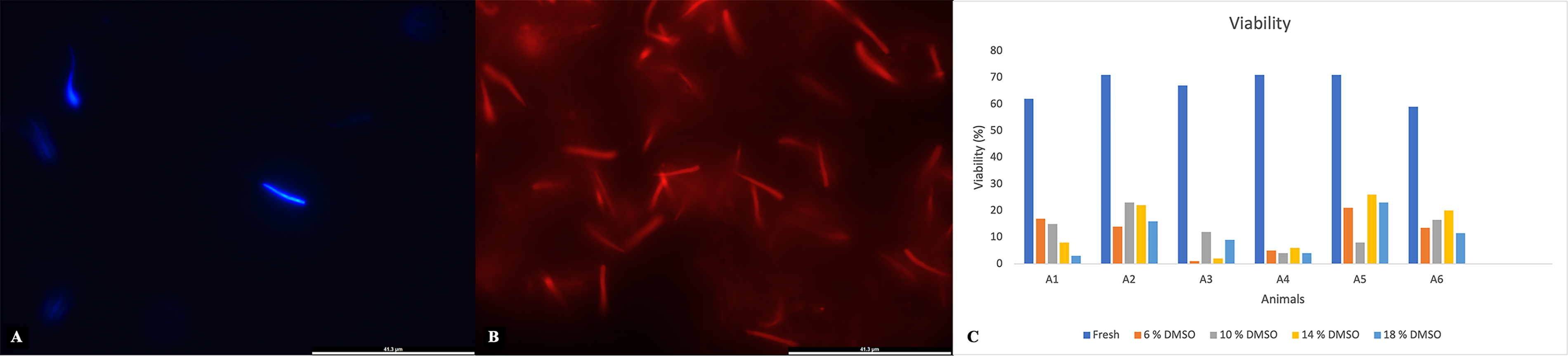
Percentages of sperm viability in greater rheas (Rhea americana; n = 6 animals) before and after cryopreservation in OvodylTM extender supplemented with different concentrations of dimethyl sulfoxide (DMSO).
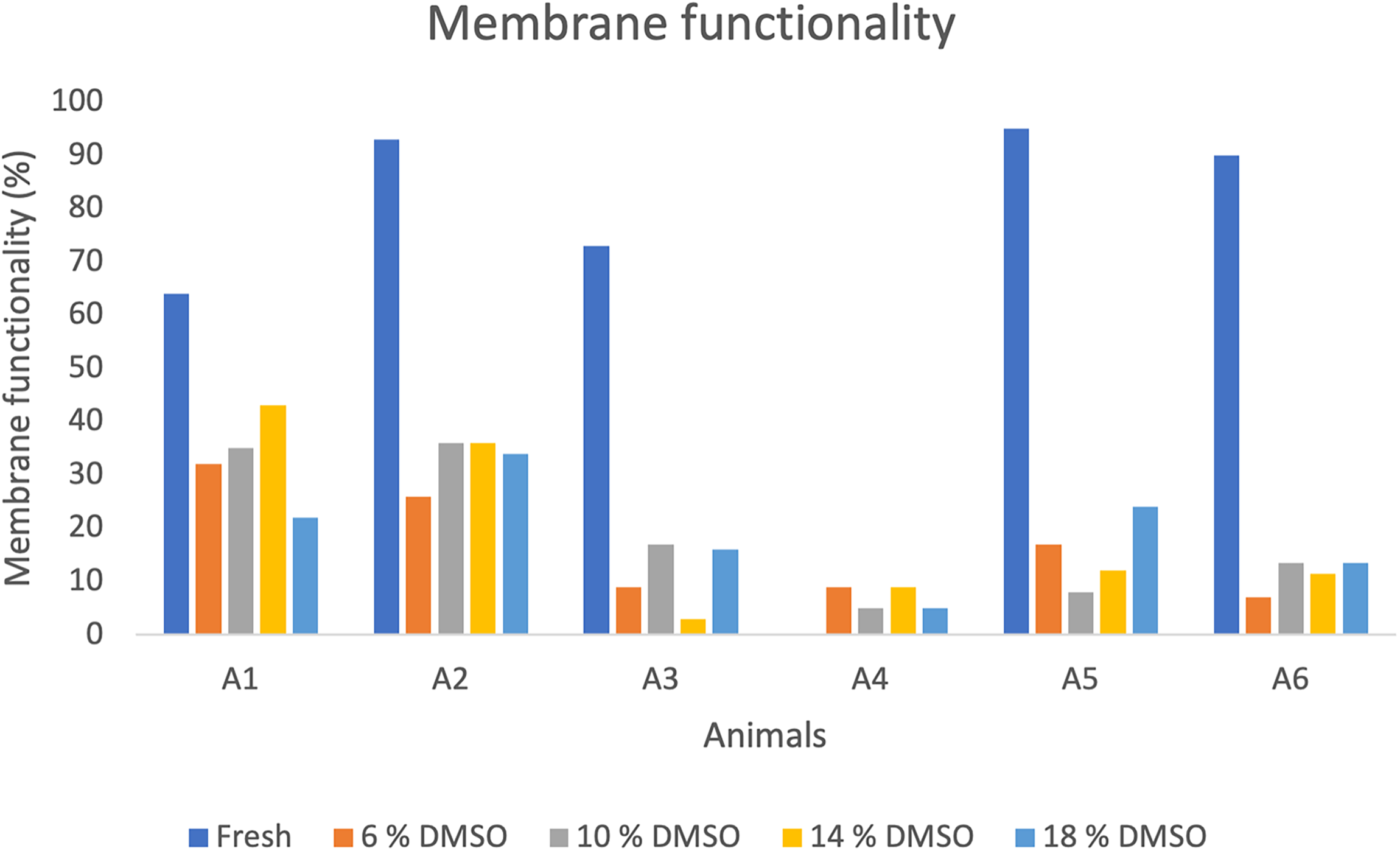
Percentages of sperm membrane functionality (%) in greater rheas (Rhea americana; n = 6 animals) before and after cryopreservation in OvodylTM extender supplemented with different concentrations of dimethyl sulfoxide (DMSO).
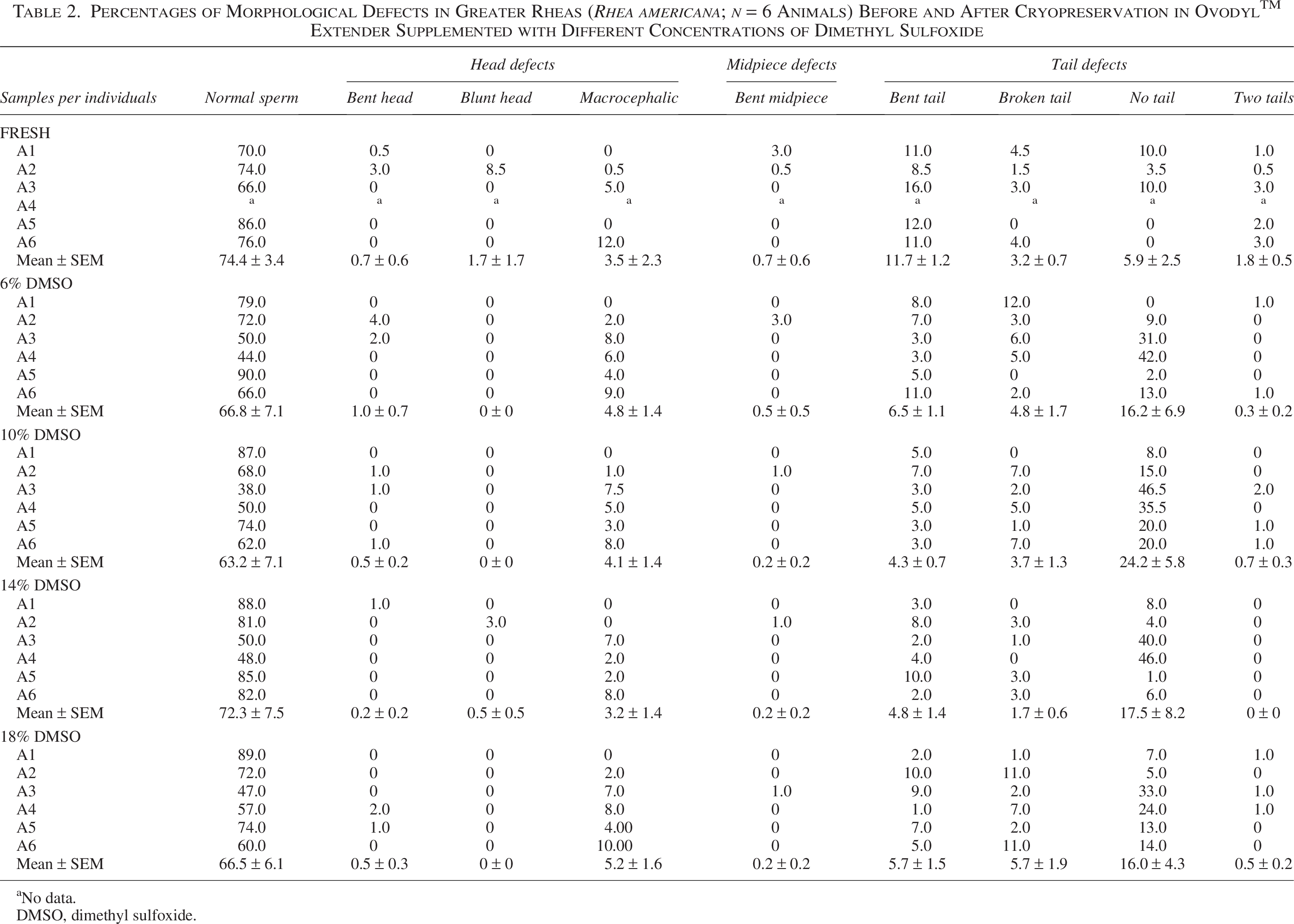
The average of normal sperm and detailed morphology data by animal are reported in Table 2. The highest percentage of normal sperm was found in Animal 5 (86%), and the most frequent defect was bent tail in Animal 3 (16%) (Table 2). Due to technical problems, it was not possible to evaluate the functionality and morphology of fresh sperm from Animal 4.
Percentages of Morphological Defects in Greater Rheas (Rhea americana; n = 6 Animals) Before and After Cryopreservation in OvodylTM Extender Supplemented with Different Concentrations of Dimethyl Sulfoxide
No data.
DMSO, dimethyl sulfoxide.
Postthawing kinetic parameters
Due to technical problems, it was not possible to evaluate the postthawing kinetic parameters of Animal A2, so only five animals were considered for this evaluation.
Postthawing sperm quality was markedly reduced in all treatments. Total and progressive motility remained close to zero in most cases, regardless of DMSO concentrations. Despite this overall reduction, a high degree of interindividual variability was observed (Fig. 1A). The highest postthawing sperm motility was achieved by Animal 3, with 11% motile sperm, when using 14% DMSO; however, for the same treatment, motility was zero in Animal 6 (Fig. 1A). After cryopreservation, progressive motility values were less than 5%, with Animals 1 and 6 showing zero in all treatments, and the highest value of 4% in Animal 5 in the treatment with 6% DMSO. Velocity parameters (VAP, VSL, and VCL) also varied among individuals. The highest values of VAP, VSL, and VCL after cryopreservation were achieved by Animal 4 with 6% DMSO (163.1, 119.3, and 312.3 µm/s, respectively).
Regarding the velocity distribution of frozen-thawed sperm, the percentage of rapid sperm was low in all treatments, with the highest value of rapid sperm (10%) being observed in Animal 3 with 14% DMSO (Fig. 2A). Overall, no significant differences were detected among DMSO concentrations for any of the evaluated parameters (p > 0.05) (Table 3), indicating that the cryoprotectant concentration did not influence postthaw sperm quality.
Average Values (Mean ± SEM) of Spermatozoa Characteristics in Greater Rheas (Rhea americana; n = 6 Animals) After Cryopreservation in Ovodyl™ Extender Supplemented with Different Concentrations of Dimethyl Sulfoxide
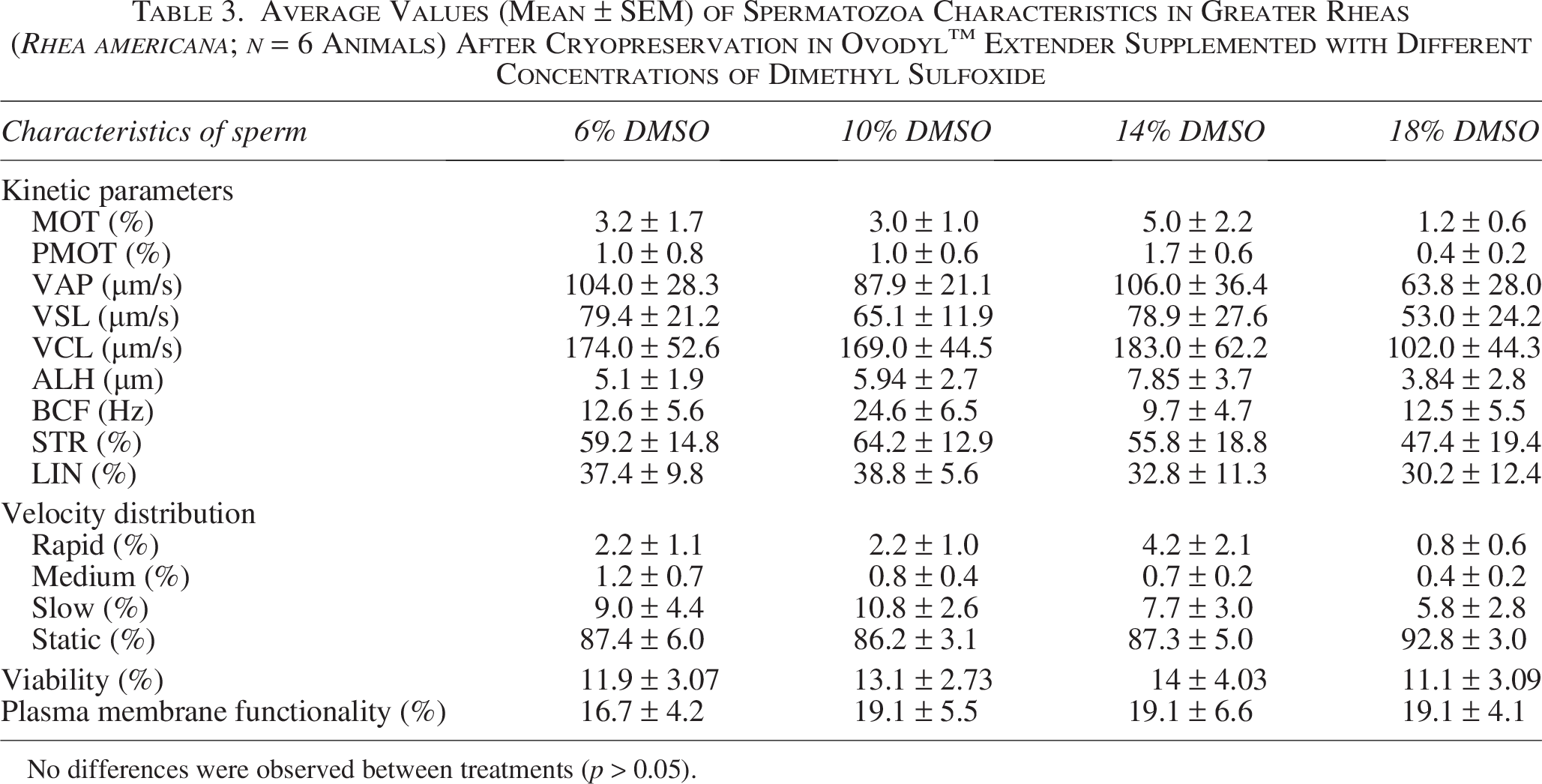
No differences were observed between treatments (p > 0.05).
A marked interindividual variability was observed. Remarkable efficiency (61%) with 14% DMSO was obtained for individual Animal 3 (Fig. 5A). However, in most treatments, efficiency was less than 10% (Fig. 5A). In Animals 1 and 6, there was no efficiency in preserving progressive motility in any treatment; however, in Animal 3, the 14% DMSO treatment showed 150% preservation efficiency (Fig. 5B). Reflecting a low initial motility before freezing and a preservation of this parameter. Regarding the average efficiency of preserving total motility, the 14% DMSO treatment showed 18.3 ± 11.6%, while 18% DMSO treatment showed 4.8 ± 2.7% (Table 4). The efficiency of preserving progressive motility was 47.0 ± 28.5% in 14% DMSO, while it was 7.3 ± 7.3% in 6% DMSO (Table 4).
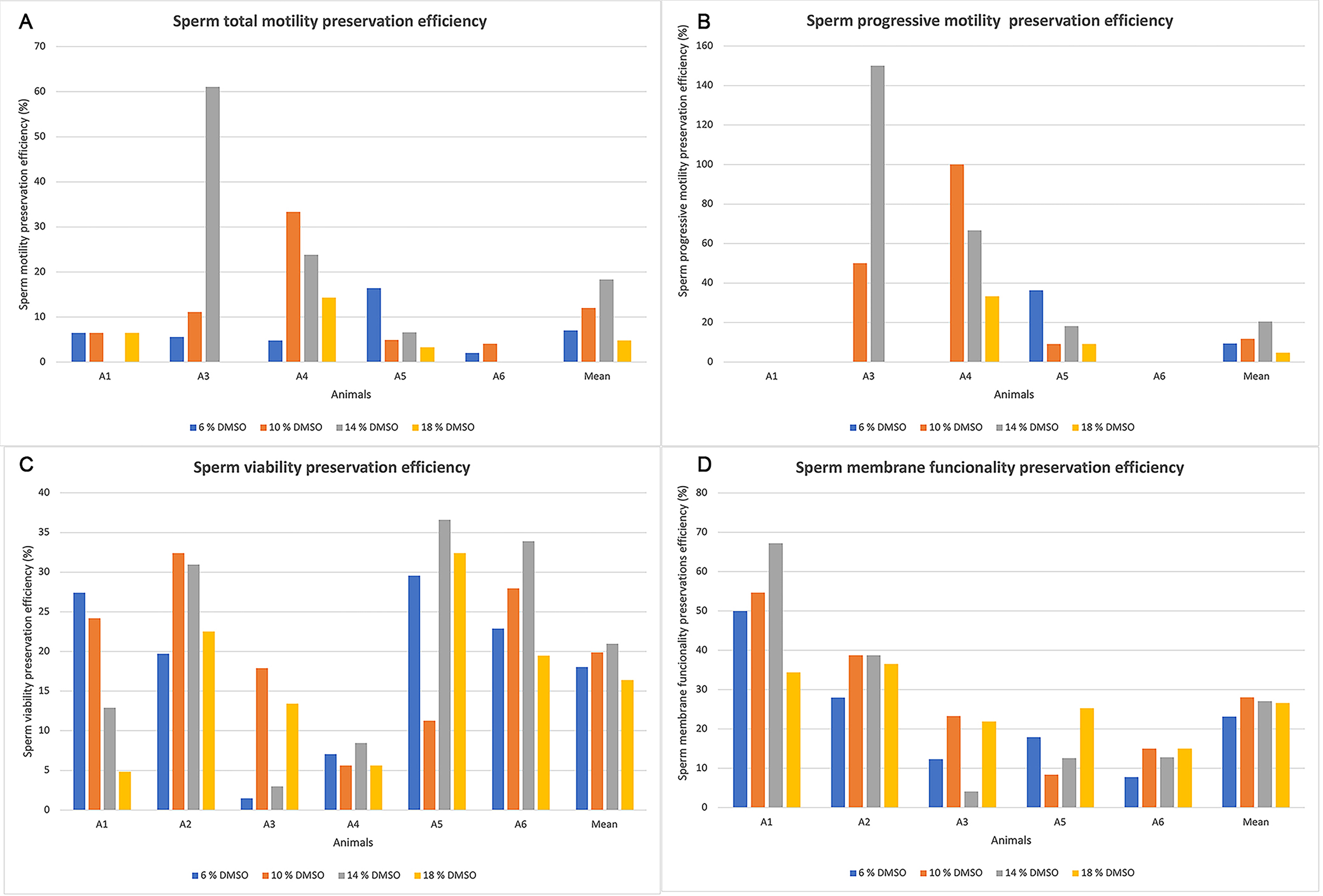
Individual values of sperm preservation efficiency in greater rhea (Rhea americana; n = 6 animals) after cryopreservation in OvodylTM extender supplemented with different concentrations of dimethyl sulfoxide (DMSO).
Average Values (Mean ± SEM) of Sperm Preservation Efficiency in Greater Rhea (Rhea americana; n = 6 Animals) After Cryopreservation in OvodylTM Extender Supplemented with Different Concentrations of Dimethyl Sulfoxide)
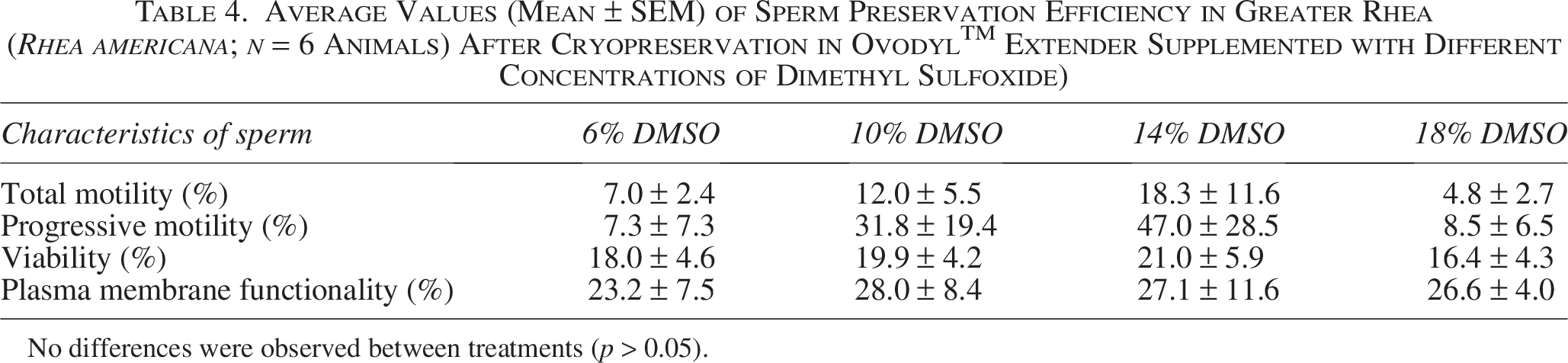
No differences were observed between treatments (p > 0.05).
Postthawing viability and membrane functionality
The percentage of live sperm after cryopreservation ranged from 3% (Animal 1; 18% DMSO) to 26% (Animal 5; 14% DMSO) (Fig. 3C). Concerning the efficiency in preserving viability after cryopreservation, slightly higher values were observed in the 14% DMSO condition, with some individuals exceeding 30% efficiency (Animals 2, 5, and 6) (Fig. 5C). Regarding the average efficiency of viability preservation, the 14% DMSO treatment was 21.0 ± 5.9%, while the 18% DMSO treatment was 16.4 ± 4.3 (Table 4).
Regarding plasma membrane functionality, the highest value (43%) observed was in Animal A1 using 14% DMSO (Fig. 4). No differences were observed between treatments (p > 0.05) (Table 3). Furthermore, the highest efficiency in preserving functionality (67.2%) was also in Animal 1 using 14% DMSO (Fig. 5D). The average efficiency of preserving membrane functionality in 10% DMSO was 28.0 ± 8.4%, while in 6% DMSO it was 23.2 ± 7.5% (Table 4).
Postthawing sperm morphology
The highest percentage of sperm with normal morphology, 90%, was observed in Animal 5 with 6% DMSO, while in Animal 3 with 10% DMSO, only 38.0% were morphologically normal (Table 2). The most evident defect was the detachment between head and tail (46.5%) in Animal 3 with 10% DMSO (Table 2, Fig. 6).
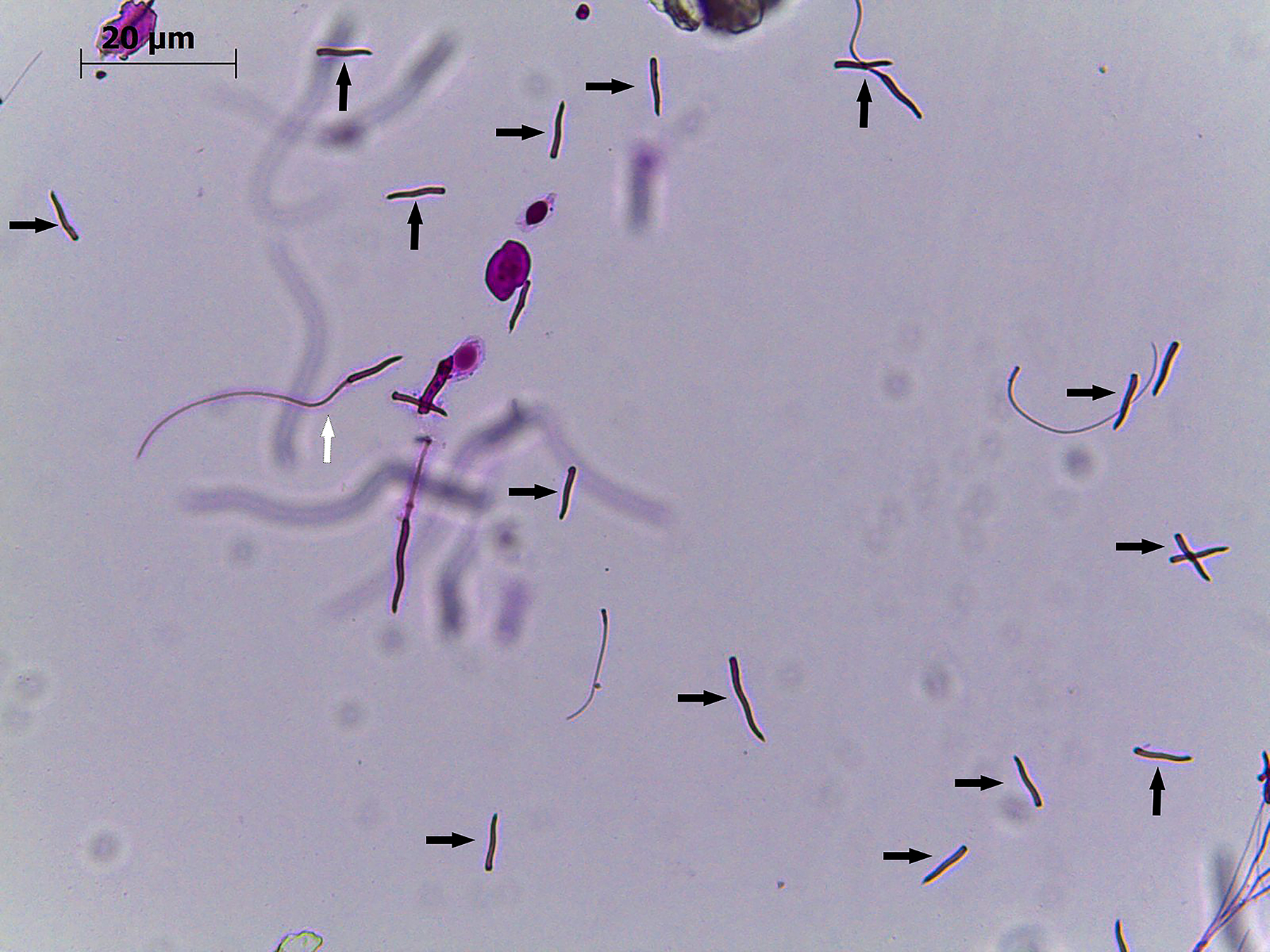
Head-tail detachments in rhea (Rhea americana) spermatozoa after cryopreservation with 14% dimethyl sulfoxide (DMSO) (Animal 4). Black arrows point to sperm heads where tail detachment occurred. White arrow points to a normal spermatozoon. Magnification 40×.
Interindividual variation
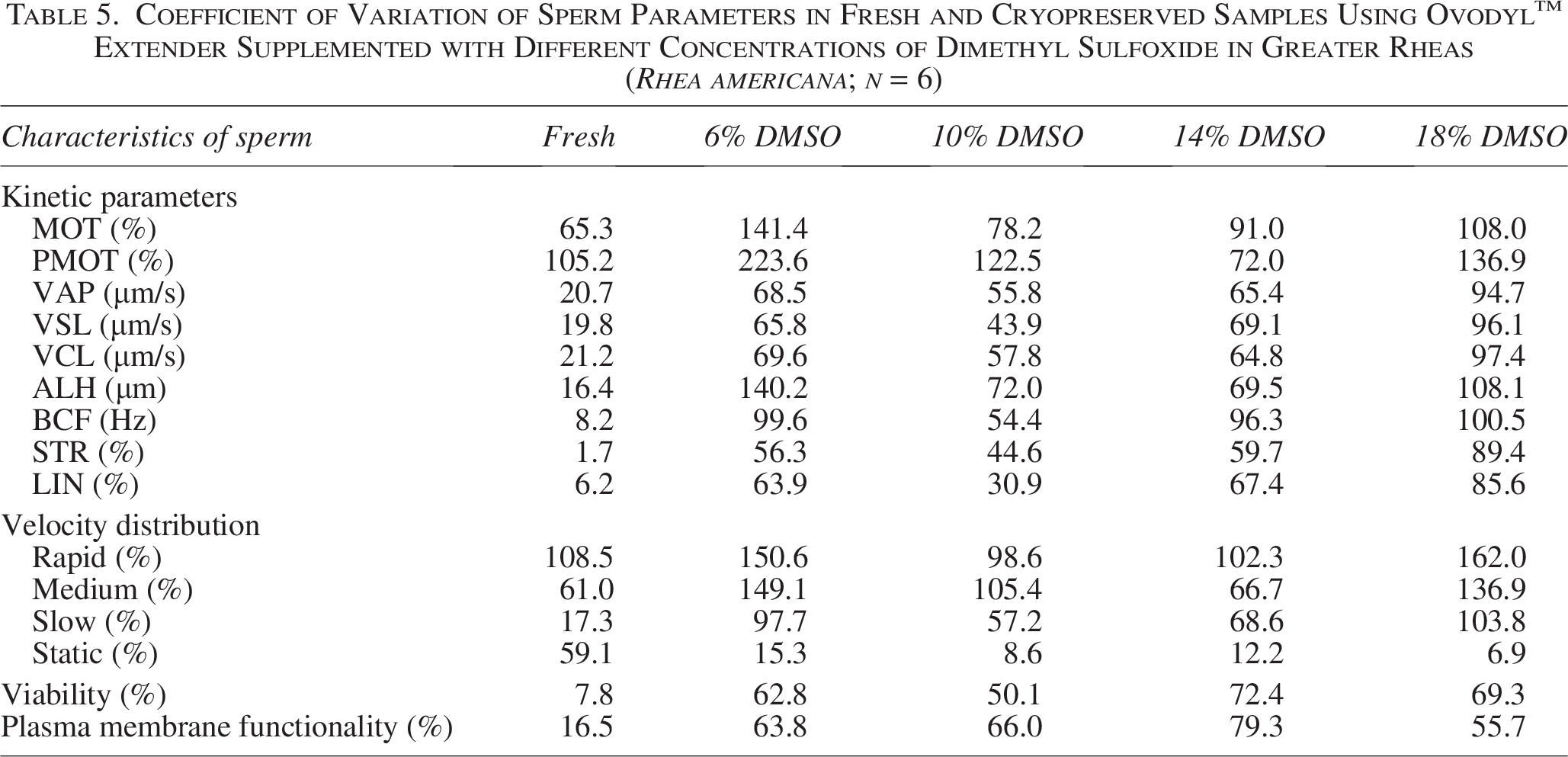
Analysis of the CV revealed high interindividual variability, mainly in parameters related to motility (Table 5). Total and progressive motility showed the highest CV values, both in fresh samples and after cryopreservation, indicating heterogeneous responses among individuals. In general, an increase in variability was observed after thawing, especially in sperm subpopulations (rapid, medium, and slow) (Table 5). In contrast, parameters such as BCF, STR, LIN, and viability showed less dispersion in fresh data, although they also showed increased variability after cryopreservation (Table 5).
Coefficient of Variation of Sperm Parameters in Fresh and Cryopreserved Samples Using Ovodyl™ Extender Supplemented with Different Concentrations of Dimethyl Sulfoxide in Greater Rheas (Rhea americana; n = 6)
Discussion
This is the first study to report the preservation of reproductive cells from rheas, representing a major step in the conservation of this species. The main challenge is the variation in the quality of fresh semen and the response to sperm freezing between individuals.
The mean total motility observed in this study was 30.2 ± 8.0. Recently, our team observed a mean value for kinematic parameters of rhea sperm, 14.6 ± 2.5%. 19 In the present study, the kinematic parameters were higher than those previously reported for this species. Since this is the first study reporting sperm cryopreservation in the greater rhea, comparisons with other ratite species provide important context. In emus, postthaw sperm quality has also been shown to decline markedly, with motility values remaining below 30% and accompanied by reductions in viability. 15 Furthermore, functional assessments, such as the ability to penetrate the perivitelline layer, were severely impaired after cryopreservation, even when some motility was preserved. These findings are consistent with the present results and indicate that low postthaw sperm performance is a common feature in ratites. Therefore, the outcomes observed in R. americana reinforce the inherent challenges of sperm cryopreservation in this group and highlight the need for further protocol optimization.
In the present study, total motility of fresh sperm ranged from 15.5% (Animal 1) to 61.0% (Animal 5). In addition, the high CV revealed a wide individual variability in the seminal parameters of rheas. High interindividual variability has also been reported in other avian species such as the rooster (Gallus domesticus), guinea fowl (Numida meleagris), 22 and long-tailed finch (Poephila acuticauda), 23 where marked differences between males and the presence of distinct sperm subpopulations were reported. In our study, the characteristics that varied the most were those related to motility parameters. It is known that sperm motility is strongly influenced by physiological conditions, maturation processes, and the reproductive environment. 24 In birds, spermatozoa acquire motor competence along the male reproductive tract and exhibit variable performance depending on intrinsic and extrinsic factors, which directly affect their fertilization capacity. 24 Furthermore, spermatozoa with greater motility are more successful in reaching and occupying sperm storage sites in the female reproductive tract, suggesting that motility differences between males are biologically relevant and may reflect underlying individual variability in sperm function. 24 Additionally, this variability appears to be increased by the cryopreservation process, as evidenced by the increased dispersion of postthaw parameters observed here. It is well established that avian spermatozoa are sensitive to freezing damage, and individual differences in membrane composition and osmotic tolerance can lead to uneven resistance to freezing and thawing procedures. 22 The underlying causes of the observed interindividual variability were not directly investigated in this study. However, differences in sperm quality and cryoresistance between males might be associated with intrinsic factors, such as individual physiological state, genetic makeup, and differences in sperm maturation processes. Since all animals were kept under similar environmental and management conditions, external factors such as diet or housing are less likely to explain the observed variability. Therefore, the heterogeneous response observed among males in this study reflects inherent biological differences that influence sperm resilience, reinforcing the importance of considering individual variability when developing and optimizing cryopreservation protocols for this species.
In addition to movement-related parameters, the viability, membrane functionality, and normal sperm morphology of Animal 1 were lower than those of most animals, 62.0%, 64.0%, and 70.0%, respectively. It is worth noting that Animal 1 was the oldest in the group (5 years), while the other animals were of similar age (∼3.0 years). It is known that age can influence seminal parameters in birds such as the North African houbara bustard and roosters, with older birds tending to have poorer-quality semen.25,26 However, in addition to age, it is possible that other intrinsic factors alter the seminal quality of rheas. It is also known that a dominance relationship between males has been observed in rheas. 27 It is possible that this dominance relationship influences sperm quality, as observed in wild sparrows (Passer domesticus). 28
Cryoprotectant type and concentration are key factors influencing the success of sperm cryopreservation. In the present study, DMSO was selected based on its widespread use and documented effectiveness in avian species.12,13,29–32 Another intracellular cryoprotectant, dimethylacetamide (DMA), has also been used in ratites 15 and presents a similar mechanism of action to DMSO,33,34 although structural differences may influence membrane interactions, osmotic tolerance, and toxicity. 35 In this way, we also encourage studies that compare the action of other cryoprotectants to DMSO.
The range of DMSO concentrations (6%, 10%, 14%, and 18%) was based on previous studies in birds, where both lower 36 and higher concentrations12,14,15,29 have shown variable success depending on the species.12,14,29 In emus, concentrations of 6% up to 24% of DMA were evaluated. 15 Values of 52 to 55 ± 2.3% membrane integrity were achieved at the use of 18% and 24% DMA, while a low membrane preservation rate was obtained with 6%–12% DMA after thawing. 15 Based on this variability, a range of concentrations was evaluated in the present study to explore their effects on rhea sperm cryosurvival.
Freezing at −140°C was selected for rheas based on previous results obtained in emus, where it proved to be more effective than freezing at −35°C. 15 This approach aims to reduce the exposure time of spermatozoa to the critical temperature range of −15°C to −60°C, where maximum damage is known to occur. 37 Under the conditions described by Sood et al., 15 exposure of the straws to −140°C rapidly reduces their temperature to approximately −78°C, thereby minimizing solute damage caused by ice crystal formation.
We reported a decrease in the values of kinetic parameters after thawing. The average total motility ranged from 1.2 ± 0.6% (18% DMSO) to 5.0 ± 2.2% (14% DMSO). Due to the cryoinjuries caused by the low cryopreservation temperature, the averages of cryopreserved samples are lower than fresh samples. The loss of sperm quality occurs mainly due to ice crystallization damage, oxidative stress, heat shock, and osmotic shock. 38 Overall, sperm motility results after cryopreservation of wild bird sperm remain low compared with those of several mammalian species.12,32,39 No statistical differences were observed between the postthawing treatments. In avian species, several studies have explored the use of antioxidant supplementation to mitigate cryodamage and improve postthaw sperm quality. 40 For example, the addition of glutathione has been shown to enhance sperm parameters and reduce oxidative stress in turkey semen after cryopreservation. 40 Although such approaches were not evaluated in the present study, they highlight potential strategies to improve sperm cryosurvival in rheas.
The mean efficiency of preserving total and progressive motility was not different. Although preservation efficiency is not commonly reported in studies on sperm cryopreservation from wild avian species, approximate comparisons can be inferred from published values of pre- and postthaw sperm parameters. Based on these data, the efficiency values for motility parameters observed in the present study are below the range reported for emus. 15 However, such comparisons should be interpreted with caution due to differences in methodologies, species, and experimental conditions.
The different responses observed among animals also reflected the preservation efficiency. It is known that avian and mammalian semen characteristics can differ between species, breeds, and lineages. 38 Semen from some individuals may be more resistant than others, like in wild boars. 41 Furthermore, even birds that initially had higher motility, such as Animals 5 and 6 (61% and 49%, respectively), had reduced or even zero motility after thawing, demonstrating low resistance to cryopreservation. It is also worth noting that the low initial motility observed in most fresh samples limits the potential for recovery after cryopreservation. In Animal 3, an efficiency in preserving progressive motility of 150% was observed with 14% DMSO. This occurred because the observed progressive motility was 3% after cryopreservation, while it was only 2% for the fresh sample. This shows that although the progressive motility values were very low, they were unchanged by the cryopreservation.
No differences were observed between treatments regarding the viability and functionality of the membrane. These metrics varied less, as shown by the low CV (Table 5). Some intrinsic characteristics of avian sperm make them more vulnerable to damage during the freezing process. For example, avian sperm have less cytoplasm and mitochondria and a higher amount of polyunsaturated fatty acids in the plasma membrane than some mammalian species, such as humans and cattle. 42 This higher concentration of fatty acids increases membrane fluidity, making it more susceptible to oxidation, especially during cryopreservation. Furthermore, it has been observed in Galliformes that sperm cholesterol content can influence sperm cryosurvival. 43 Cholesterol is also a key component of the plasma membrane, supporting its structure. 44 In rheas, there are no studies characterizing the sperm plasma membrane. It is possible that the composition of the sperm plasma membrane leads to variable sensitivity to freezing and thawing. This includes damage caused by the movement of lipids within the membrane during the transition phase and loss of lipids. Therefore, we recommend future studies to investigate the composition of the sperm plasma membrane to determine the best way to adapt cryopreservation protocols to the membrane composition.
It is known that the imbalance between cellular antioxidant defense systems and the production of reactive oxygen species (ROS) during freezing leads to oxidative stress. 45 In roosters, it has been reported that increased ROS can induce detrimental changes during the semen cryopreservation process.46,47 High levels of ROS can cause the destruction of nucleic acids, proteins, lipids, and carbohydrates, which lead to cell death. 48 A decline in mitochondrial activity and, consequently, motility may be associated with increased ROS formation. 49 Further investigations about the addition of antioxidants to the freezing medium for rhea sperm are warranted.
Morphology was one of the best-maintained parameters, with the largest number of defects observed in the tail. It is known that the tail is the most sensitive part of the avian sperm cell. 50 Other ultrastructural damage might have occurred in the membrane, mitochondria, chromatin, and other organelles; however, they were not explored in this study. Thus, we suggest further investigating fresh and thawed rhea spermatozoa to understand the damage that occurs at the ultrastructure level.
Some limitations have to be considered in the interpretation of the results. The small sample size (n = 6) is due to the limited availability of animals and the opportunistic nature of the sampling, which reduces the statistical power of the analyses. As rheas are a near-threatened wild species, we had an ethical commitment to minimize the number of animals used in the study. We therefore used six males already scheduled for euthanasia, avoiding the need to sacrifice additional individuals. Despite the limited sample size, the data remain highly valuable given the scarcity of information available in this species. Comparable or even smaller numbers of individuals have been reported in studies describing sperm parameters in other wild animals. This includes critically endangered species such as Southern muriqui (Brachyteles arachnoides) 51 or Bornean orangutans (Pongo pygmaeus), 52 or species of least concern, such as the six-banded armadillos (Euphractus sexcinctus), 53 crab-eating foxes (Cerdocyon thous), 54 or Griffon vultures (Gyps fulvus). 55 Furthermore, no control cryo-conditions could be included, as there are no previously established cryopreservation protocols for rheas. Therefore, all conditions were evaluated comparatively within the same experimental framework. Despite these limitations, the study provides the first insights into sperm cryopreservation in R. americana and establishes a basis for future investigations
Conclusion
This study provides the first evidence that sperm cryopreservation in R. americana is feasible but currently limited by low postthaw sperm quality and strong individual-dependent responses. No difference was observed between the different concentrations of DMSO. These findings highlight the need for protocol improvement, particularly regarding cryoprotectant selection and optimization of freezing conditions. Future studies should focus on improving the functional parameters of spermatozoa and validating fertilization capacity through artificial insemination assays. From a conservation standpoint, this information is essential to advance the understanding of sperm physiology in this species. Results emphasize that biobank strategies for this species should consider individual variability and prioritize the identification of males with greater cryotolerance to maximize the preservation of genetic resources.
Ethical Statement
All procedures were approved by the UFERSA Animal Ethics Committee, Process no. 23091.001423/2020-8.
Data Availability Statement
The data that support the findings of this study are available from the corresponding authors upon reasonable request.
Authors’ Contributions
L.G.P.B.: Conceptualization, methodology, writing—original draft preparation, and investigation. A.M.S.: Methodology and investigation. M.R.T.D.: Methodology and investigation. R.P.d.S.: Methodology and investigation. S.S.J.M.: Methodology and investigation. A.G.P.: Methodology and investigation. M.F.d.O.: Methodology and validation. P.N.J.N.: Writing—review and editing. P.C.: Writing—review and editing. A.R.S.: Conceptualization, writing—review and editing, investigation, supervision, and project administration.
Footnotes
Acknowledgments
The authors thank the team of the Center for Multiplication of Wild Animals/UFERSA for providing the biological material used in the study; IMV Technologies for providing the extender; and Prof. José Rodolfo Lopes de Paiva Cavalcanti for providing the microscope for the production of fluorescence photos.
Author Disclosure Statement
No conflicting financial interests exist.
Funding Information
This study was financed in part by the Coordination for the Improvement of Higher Education Personnel–Brazil (CAPES; Financial Code-001) and the National Council for the Scientific Development (CNPq; grant no. 303929/2018-9).