
Abstract
Background
Articular chondrocytes (ACs) secrete a variety of extracellular matrix components to maintain the functions of articular cartilage. Degeneration of ACs leads to the degeneration of articular cartilage and consequently to osteoarthritis. The secretion of bone marrow mesenchymal stem cells (BMSCs) is capable of protecting ACs from degeneration, and thus BMSCs are widely applied to treat osteoarthritis.
Objective
This study aims to explore whether BMSCs and ACs will affect the functions of each other through their secretions in the context of osteoarthritis.
Design
BMSCs and ACs isolated from rabbits were identified using flow cytometry and immunocytochemistry. Conditioned medium of BMSCs and ACs treated with 0, 5, 10, 20, and 40 ng/ml of tumor necrosis factor-alpha (TNF-α) were collected and used to treat ACs and BMSCs, respectively. The viabilities of ACs and BMSCs treated with condition medium were assessed using a Cell Count Kit-8 (CCK-8) kit. Reverse transcription-quantitative polymerase chain reaction (RT-qPCR), immunoblotting, and enzyme-linked immunosorbent assay (ELISA) methods were employed to evaluate the relative expression levels of genes and proteins, as well as the cytokine concentrations in the supernatant.
Results
Immunofluorescence and flow cytometry results indicated that the purity of isolated cells exceeded 95%. CCK-8 analysis showed that 6 hours of treatment with a conditioned medium did not affect the viability of BMSCs and ACs. However, treatment for 12 hours or longer significantly increased the viability of BMSCs (p < 0.05) and significantly decreased the viability of ACs (p < 0.01). RT-qPCR results demonstrated that the relative expression levels of Runx2 (1.15-3.91), Alp (1.06-2.84), TNF (BMSCs: 0.94-2.54; ACs: 1.03-2.64), IL6 (BMSCs: 0.98-2.78; ACs: 0.96-3.71), IL17A (BMSCs: 1.08-5.91; ACs: 0.90-4.20), and IL10 (BMSCs: 0.93-2.82; ACs: 0.89-2.25) genes in conditioned medium-treated BMSCs and ACs were dose-dependently elevated (p < 0.001) by TNF-α treatment. Immunoblotting analysis revealed that the expression levels of RUNX2 (0.53-0.86) and ALP (0.49-0.85) proteins were also dose-dependently elevated (p < 0.001) by TNF-α treatment. ELISA results showed similar TNF-α dose-dependent increases (p < 0.001) in the supernatant concentrations of pro-inflammatory cytokines TNF-α (BMSCs: 36.90 ± 0.75 to 199.38 pg/ml; ACs: 29.76 to 293.99 pg/ml), interleukin (IL)-6 (BMSCs: 4.96-48.24 pg/ml; ACs: 6.12-38.15 pg/ml), IL-17 (BMSCs: 3.06-28.99 pg/ml; ACs: 3.08-28.51 pg/ml), as well as the anti-inflammatory cytokine IL-10 (BMSCs: 6.34-65.02 pg/ml; ACs: 5.30-34.85 pg/ml).
Conclusion
Together, these results indicate a TNF-α-regulated bidirectional interaction between BMSCs and ACs, deepening our understanding of the pathogenesis of osteoarthritis and aiding in its prevention and treatment.
Keywords
Introduction
Articular chondrocytes (ACs) are metabolically active cells residing in articular cartilage, playing crucial roles in maintaining cartilage homeostasis by synthesizing and updating various extracellular matrix components, such as collagen, glycoproteins, proteoglycan, and hyaluronan.1,2 It is well recognized that the dysfunction of ACs, resulting from alterations in the inflammatory microenvironment, plays a key role in cartilage degeneration and the progression of osteoarthritis, a leading cause of disability in older adults.3,4 The function of ACs is influenced by many factors in their chemical and mechanical environments, including pro-inflammatory cytokines and growth factors.5-7 Therefore, modulating the microenvironment of ACs is an effective approach for alleviating osteoarthritis. For example, Xu and colleagues reported that harpagide could inhibit tumor necrosis factor-alpha (TNF-α)-induced inflammatory response in rat ACs, thereby alleviating osteoarthritis. 8
Recently, bone marrow mesenchymal stem cells (BMSCs) have been widely applied in the treatment of osteoarthritis due to their tissue repair and immunomodulatory abilities. For example, BMSCs have been found to protect against cartilage damage in animal models.9,10 Although the underlying mechanisms by which BMSCs protect cartilage are not fully elucidated, one recognized mechanism is that BMSC secretions mitigate the senescence and death of ACs. For example, Li and colleagues discovered that curcumin-primed human BMSC-derived extracellular vesicles can reverse interleukin (IL)-1β-induced catabolic responses of ACs by upregulating miR-126-3p. 11 Intriguing, the interaction between BMSCs and ACs is bidirectional, and the secretions of ACs can also in turn influence the function of BMSCs. For instance, Tang and colleagues found that paracrine secretions of ACs can facilitate the proliferation, chondrogenic gene expression, and cartilaginous matrix production of BMSCs both in vitro and in vivo. 12
Tumor necrosis factor-alpha is a mononuclear factor derived from activated mono-macrophages that exerts dose-dependent dual effects on osteogenic differentiation of BMSCs by activating necrosis factor receptors. 13 For example, Qin’s study found that upregulated expression of TNF-α suppresses the expression of mineralization and bone matrix-related genes, 14 while Yang’s study found that it increases the mineralization of extracellular matrix components. 15 This disparity is thought to be attributed to differences in TNF-α dose, species, and tissues. 13 In addition, TNF-α has been found to affect the viability and proliferation of human ACs. For instance, Lsyar et al. investigated the effects of TNF-α inhibition on human chondrocytes in monolayers and found that their viability and proliferation were significantly inhibited, 16 indicating that TNF-α might have positive effects on ACs. Notably, these investigations primarily focused on the effects of TNF-α on BMSCs or ACs individually and did not explore the effects of TNF-α on their interaction. Considering the joint as an integrated system, t interactions likely exist between BMSCs, ACs, and cytokines. Uncovering the intrinsic relationships among these components will enhance our understanding of the mechanisms underlying osteoarthritis. This knowledge can provide a more comprehensive theoretical foundation for the prevention and treatment of osteoarthritis.
In this study, we isolated primary BMSCs and ACs from rabbits and investigated the effects of conditioned medium derived from BMSCs or ACs, which were both treated with varying concentrations of TNF-α, on the relative expression levels of IL6, IL17A, TNF, and IL10 genes in ACs or BMSCs, as well as on the concentrations of cytokines IL-6, IL-17, TNF-α, and IL-10 in their corresponding supernatant. Our study explored the interaction between BMSCs and ACs in the presence of different concentrations of TNF-α in the microenvironment, which will deepen our understanding of the pathogenesis of osteoarthritis and the underlying mechanisms by which BMSC transplantation mitigates osteoarthritis.
Materials and Methods
All procedures in this study were carried out in strict accordance with the Guidelines on the Care and Use of Laboratory Animals issued by the Chinese Council on Animal Research and the Guidelines of Animal Care and received approval from the Ethical and Welfare Committee for Animal Experiments of Sichuan University (Approval No: SCTK-IRB-2024-034). The sample sizes for all experiments in this study were determined by referencing previous similar studies, without conducting a power analysis.
Preparation and Culture of BMSCs and ACs
BMSCs used in this study were isolated following a previously established protocol. 17 Briefly, bone marrow was aseptically collected from the tibia and femur condyles of anesthetized male New Zealand rabbits (0.75 kg, 10 weeks old). The collected bone marrow was centrifuged at 1000 r/min for 5 minutes, and the supernatant was discarded. The cell pellets were washed with phosphate-buffered saline (PBS; Solarbio, Beijing, China) and centrifuged again. The resulting cell pellets were resuspended in a complete medium consisting of 89% alpha minimal essential medium (α-MEM; HyClone, Logan, UT), 10% fetal bovine serum (FBS; Gibco, CA, USA), and 1% penicillin/streptomycin (Sangon, Shanghai, China), and incubated at 37°C with 5% CO2. The medium was refreshed every 2 days. When the BMSCs reached 80% to 90% confluence, they were digested with 0.25% trypsin (Gibco, CA, USA) and subcultured at a 1:2 ratio.
ACs were isolated following another previously described method. 18 In brief, full-thickness articular cartilage was dissected from the femoral condyles and tibial plateaus of the rabbit knee joints. ACs were then isolated from the cartilage tissues through sequential enzymatic digestions with 14 U/ml pronase (Sigma, Missouri, USA) in Dulbecco’s Modified Eagle Medium-F12 (DMEM-F12; HyClone, Beijing, China) at 37°C for 90 minutes, followed by digestion with 31.25 U/ml collagenase-2 (Sigma, Missouri, USA) in DMEM-F12 at 37°C overnight. The isolated chondrocytes were cultured in 6-well plates (Nest, Wuxi, China) in DMEM-F12 medium with 10% FBS at 37°C and 5% CO2. The DMEM-F12 medium was refreshed every 2 days. Once reaching 80% to 90% confluence, the ACs were digested with 0.25% trypsin and subcultured at a 1:2 ratio.
Identification of BMSCs and ACs
All BMSCs and ACs used in the experiments were in passages 3 to 5. The morphologies of the BMSCs and ACs were observed daily and captured using an inverted microscope (Leica, Wetzlar, Germany). The expression of surface markers CD11b, CD34, CD45, CD73, CD90, and CD105 (Zen-bio, Chengdu, China) in the BMSCs was analyzed using flow cytometry. Specifically, the third passage BMSCs were incubated with the corresponding antibodies at room temperature for 15 minutes, washed once, and then analyzed using a flow cytometer (ThermoFisher, Shanghai, China). According to previously established criteria, 19 a cell population can be considered high-purity BMSCs if it shows more than 95% positivity for CD73, CD90, and CD105, and less than 2% negativity for CD11b, CD34, and CD45. The multipotent differentiation potentials of BMSCs, including osteogenesis, adipogenesis, and chondrogenesis, were identified using Alizarin red S (ARS) staining, Oil Red O staining, and Alcian Blue staining, respectively, after induction with osteogenesis, lipid formation, or cartilage induction medium (Cyagen Biosciences, CA, USA).
In addition, immunofluorescence was also employed to assess the expression level of CD73 and CD90 in BMSCs. To prepare, BMSCs were plated on sterile coverslips in 24-well plates at a concentration of 5 × 104 cells/ml, with each well receiving 1 ml of cell suspension. The cells adhered to the coverslips were fixed using formaldehyde (Life iLab Bio, Shanghai, China) at −20°C for 10 minutes. Following fixation, the cells were permeabilized with 0.1% Triton X-100 (Solarbio, Beijing, China) for 10 minutes at room temperature, then blocked with 10g/l bovine serum albumin (Solarbio, Beijing, China) for another 10 minutes. They were subsequently incubated with the primary antibodies overnight at 4°C, including CD73 (Abcam, Cambridge, UK; Cat No: ab317426) and CD90 (Abcam, Cambridge, UK; Cat No: ab307736). After washing 3 times with PBS, the secondary antibody (Goat Anti-Rabbit IgG H&L; ZenBio, Chengdu, China; Cat No: 550037) was applied and allowed to incubate at room temperature for 4 hours. The cell nuclei were stained with 4',6-diamidino-2-phenylindole (DAPI; Solarbio, Beijing, China) for 5 minutes at room temperature. Finally, cells were then observed and digitally captured using a fluorescence microscope (Olympus, Tokyo, Japan).
The expression level of type II collagen in ACs was evaluated using immunocytochemical analysis according to a previous study. 20 Briefly, ACs were seeded on sterile coverslips in 24-well plates at a concentration of 5 × 104 cells/ml, with each well receiving 1 ml of cell suspension. The cells adhered to the coverslips were fixed using formaldehyde (Life iLab Bio, Shanghai, China) at −20°C for 10 minutes. After fixation, the coverslips were washed 3 times with PBS to remove the fixative. Subsequently, the coverslips were blocked with 10g/l bovine serum albumin (Solarbio, Beijing, China) for 30 minutes to minimize nonspecific binding. They were then incubated with type II collagen antibody (1: 200; ZenBio, Chengdu, China) overnight at 4°C. After washing thrice with PBS, a secondary antibody (1:1000; Goat Anti-Rabbit IgG H&L; ZenBio, Chengdu, China; Cat No: 550037) was applied and allowed to incubate at room temperature for 4 hours. The stained BMSCs and ACs were observed and imaged using an inverted microscope, and the images were analyzed using ImageJ software.
Experimental Design and Conditioned Medium Preparation
After identification, 1 ml of suspension containing primary BMSCs and ACs were seeded into 6-well plates at the density of 2 × 105 cells/ml and incubated. When the cells reached approximately 80% confluence (24 hours), the medium was removed, and the cells were washed 3 times with PBS. Then, 1.5 ml of medium containing 0 ng/ml (Group A, serves as negative control), 5 ng/ml (Group B), 10 ng/ml (Group C), 20 ng/ml (Group D), and 40 ng/ml (Group E) TNF-α (Cat No: SRP3177; Merk, NJ, USA) was added to each well and incubated for 6, 12, 24, and 48 hours. The concentrations of TNF-α were determined based on previous studies.13,21 Afterward, the medium was removed again, and the cells were washed another 3 times. Next, 1.5 ml of fresh medium was added for another 12 hours of incubation. Afterward, the corresponding supernatant (labeled as BMSCCM-1 and ACCM-1, respectively) was collected for subsequent experiments.
In addition, other primary BMSCs and ACs were seeded into 6-well plates at the same density and incubated. When these cells reached approximately 80% confluence (24 hours), the medium was removed, and the cells were washed 3 times with PBS. The collected BMSCCM-1 and ACCM-1 were then added to the ACs and BMSCs, respectively, and incubated for 6, 12, 24, and 48 hours. Similarly, the medium was removed again, and the cells were washed another 3 times. Next, 1.5 ml of fresh medium was added for another 12 hours of incubation. Afterward, the BMSCs (labeled as BMSC-2) and ACs (labeled as AC-2) and their corresponding supernatant (labeled as BMSCCM-2 and ACCM-2, respectively) were collected for subsequent analysis. Figure 1 shows the sketch of the design of this experiment.

Sketch of the experimental design. Different colored rectangles indicate culture with different mediums. BMSCs = bone marrow mesenchymal stem cells; ACs = articular chondrocytes; CM = conditioned medium.
Cell Viability Analysis
The viabilities of the BMSCs and ACs in each group were assessed using the Cell Count Kit-8 (CCK8) assay kit (Solarbio, Beijing, China). Specifically, 100 μl of fresh medium containing 1 × 105 BMSCs or ACs was seeded into a 96-well plate, with 3 replicates for each group. After 24 hours of incubation, the medium was removed, and the cells were washed 3 times with PBS. Then, 100 μl of the collected BMSCCM-1 and ACCM-1 were added to the ACs and BMSCs, respectively, and incubated for 6, 12, 24, and 48 hours. Afterward, the medium was again removed, and the cells were washed another 3 times. Next, 100 μl of fresh medium was added for an additional 12 hours of incubation. The supernatant in each well was then discarded, and 100 μl of fresh complete medium containing 10% CCK-8 solution was added. After 2 hours of incubation, the optical density at 450 nm (OD450) was measured using a microplate reader (ThermoFisher, Shanghai, China).
Reverse Transcription-Quantitative Polymerase Chain Reaction (RT-qPCR) Analysis
In brief, the total RNA of collected BMSCs-2 and ACs-2 was extracted from cells using Trizol Reagent (TransGen, Beijing, China) following the manufacturer’s protocol. The concentration and purity of RNA were assessed with a spectrophotometer (ThermoFisher, NC, USA) and 1% agarose gel electrophoresis. The total RNA samples were then reverse-transcribed into complementary DNA (cDNA) using the RevertAid First Stand cDNA Synthesis Kit (ThermoFisher, Shanghai, China) according to the manufacturer’s instructions. Subsequently, the relative expression levels of IL6 (encoding IL-6), IL17A (encoding IL-17), TNF (encoding TNF-α), IL10 (encoding IL-10), Runx2 (encoding Runt-related transcription factor 2, RUNX2), and Alp (encoding alkaline phosphatase, ALP) genes to the GAPDH (encoding GAPDH) gene were determined. The resulting Ct data were analyzed using the 2−ΔΔC method in Excel software. The sequences of the gene primers are presented in Table 1.
Sequences of Primers Used in the Reverse Transcription-Quantitative Polymerase Chain Reaction Experiment.

Total Protein Extraction and Immunoblotting Analysis
Proteins were extracted from the collected BMSCs using RIPA lysis buffer (Solarbio, Beijing). Protein concentrations were measured with a BCA kit (Beyond, Shanghai, China). The proteins were then denatured by heating at 100°C for 10 minutes. Equal amounts of protein from each sample were loaded onto a 12% sodium dodecyl sulfate-polyacrylamide gel for electrophoresis. After running the gel, the proteins were transferred to a polyvinylidene fluoride membrane. This membrane was then blocked with 5% skim milk in TBST for 2 hours. Following the blocking step, the membranes were incubated overnight at 4°C with primary antibodies, including RUNX2 (HuaBio, Hangzhou, China; Cat No: ET1612-47) and ALP (HuaBio, Hangzhou, China; Cat No: ET7111-23). The next day, the membranes were treated with a secondary antibody (Goat Anti-Rabbit IgG H&L; ZenBio, Chengdu, China; Cat No: 511203) and enhanced chemiluminescent reagents to visualize the protein bands. The intensity of these protein bands was analyzed using ImageJ software.
Enzyme-Linked Immunosorbent Assay (ELISA)
The concentrations of cytokines IL-6, IL-17, TNF-α, and IL-10 in the collected conditioned medium (BMSCCM-2 and ACCM-2) were assessed using corresponding commercialized ELISA kits (Nanjing Jiancheng, Nanjing, China) following their instructions. Briefly, 100 μl of supernatant was added to each well, and the reaction plate was sealed with a sealing membrane and incubated at room temperature for 2 hours. Subsequently, biotinylated antibody, horseradish peroxidase-labeled streptavidin, and tetramethylbenzidine solution were added to each well, and the plate was further incubated in the dark at room temperature overnight. Finally, 50 μl of stop solution was added to each well, and the OD450 value in each well was determined using a microplate reader.
Statistical Analysis
Data are presented as mean ± standard error and analyzed using SPSS 29 (IBM, NY, USA). Graphs were generated using OriginPro 2024b (Originlab, MA, USA), Adobe Illustrator 2022, and Adobe Photoshop 2022 (Adobe, CA, USA) unless stated otherwise. Statistical differences were determined using a 1-way analysis of variance, and the Turkey method was used for post hoc tests unless specified otherwise. For repeated-measures data, a repeated-measures analysis of variance (ANOVA) method was applied to assess differences across groups and time points. A P value of less than .05 was considered statistically significant.
Results
BMSCs and ACs Isolation and Identification
The isolated BMSCs exhibited a fibroblast-like morphology, characterized by elongated, spindle-shaped cells that were evenly distributed across the field and in a confluent monolayer. The density of the BMSCs indicated active proliferation, with the cells displaying a uniform and healthy appearance (
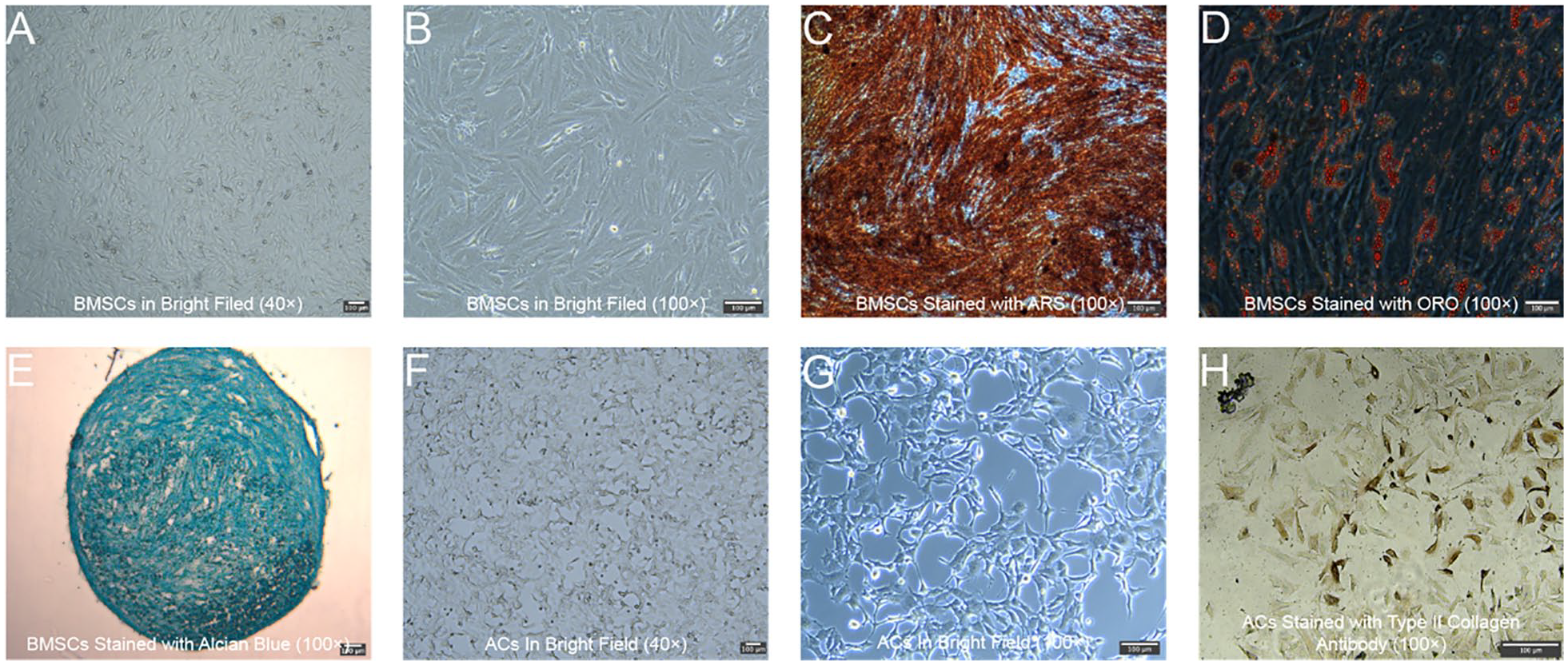
Isolation and Identification of BMSCs and ACs. (
The result of the immunofluorescence analysis indicated that nearly all of the isolated BMSCs expressed the surface markers CD73 and CD90 (
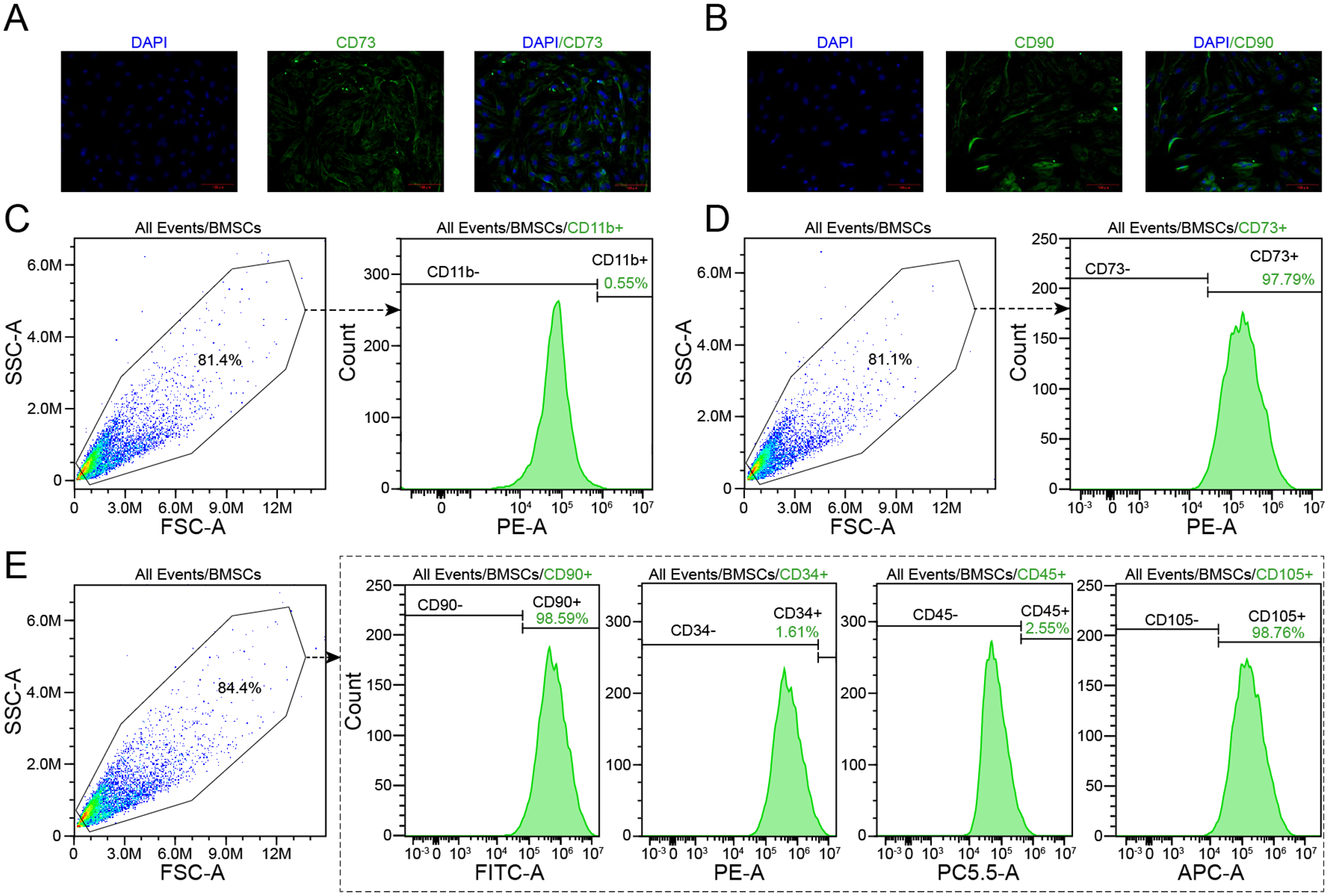
Immunofluorescence and flow cytometry analyses of the surface markers of BMSCs. (
Effects of Conditioned Medium Derived from ACs that Treated with Varying Concentrations of TNF-α on the Proliferation, Osteogenic Differentiation, and Cytokines Secretion of BMSCs
First, we evaluated the proliferation capacity of BMSCs treated with conditioned medium derived from ACs exposed to 0, 5, 10, 20, and 40 ng/ml of TNF-α for 6, 12, 24, and 48 hours. As
Figure 4A
and
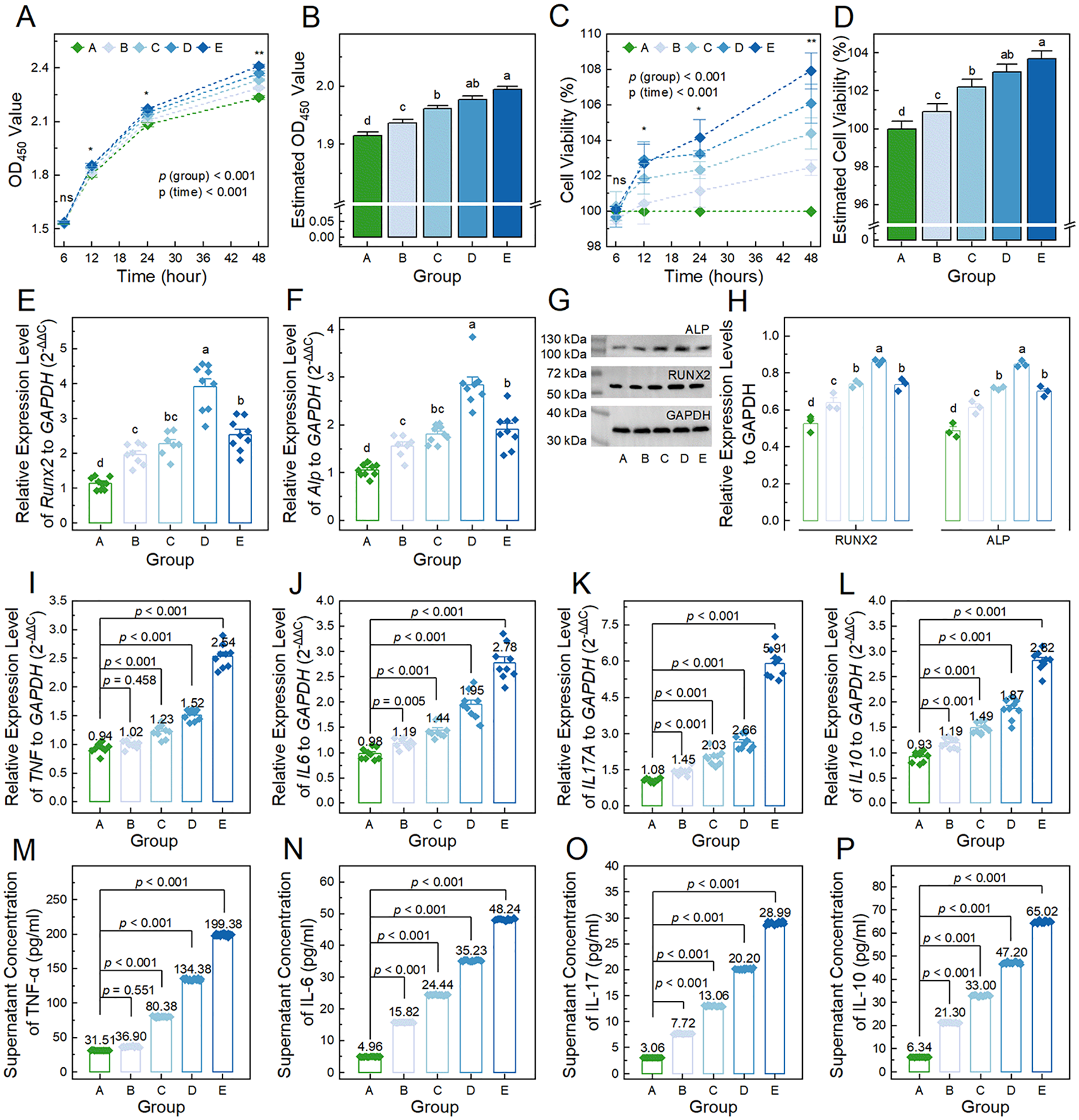
Effects of conditioned medium derived from ACs treated with varying concentrations of TNF-α on the proliferation capacity, osteogenic differentiation, and cytokine secretion of BMSCs. (
Next, we assessed the osteogenic differentiation of BMSCs in each group by measuring the relative expression levels of the indicators RUNX2 and ALP at both the gene and protein levels. As shown in
Figure 4E
and
Meanwhile, we also evaluated the cytokine secretion levels of BMSCs in each group. The relative expression levels of the TNF gene in BMSCs at baseline (0.94 ± 0.09) were comparable to those in group B (1.02 ± 0.08; p = 0.458), but significantly lower than those in groups C-E (1.22 ± 0.11, 1.52 ± 0.10, and 2.54 ± 0.20, respectively) (p < 0.001;
Effects of Conditioned Medium Derived from BMSCs That Treated with Varying Concentrations of TNF-α on the Proliferation Capacity and Cytokines Secretion of ACs
We evaluated the proliferation capacity of ACs treated with conditioned medium derived from BMSCs exposed to 0, 5, 10, 20, and 40 ng/ml of TNF-α for 6, 12, 24, and 48 hours. As
Figure 5A
and
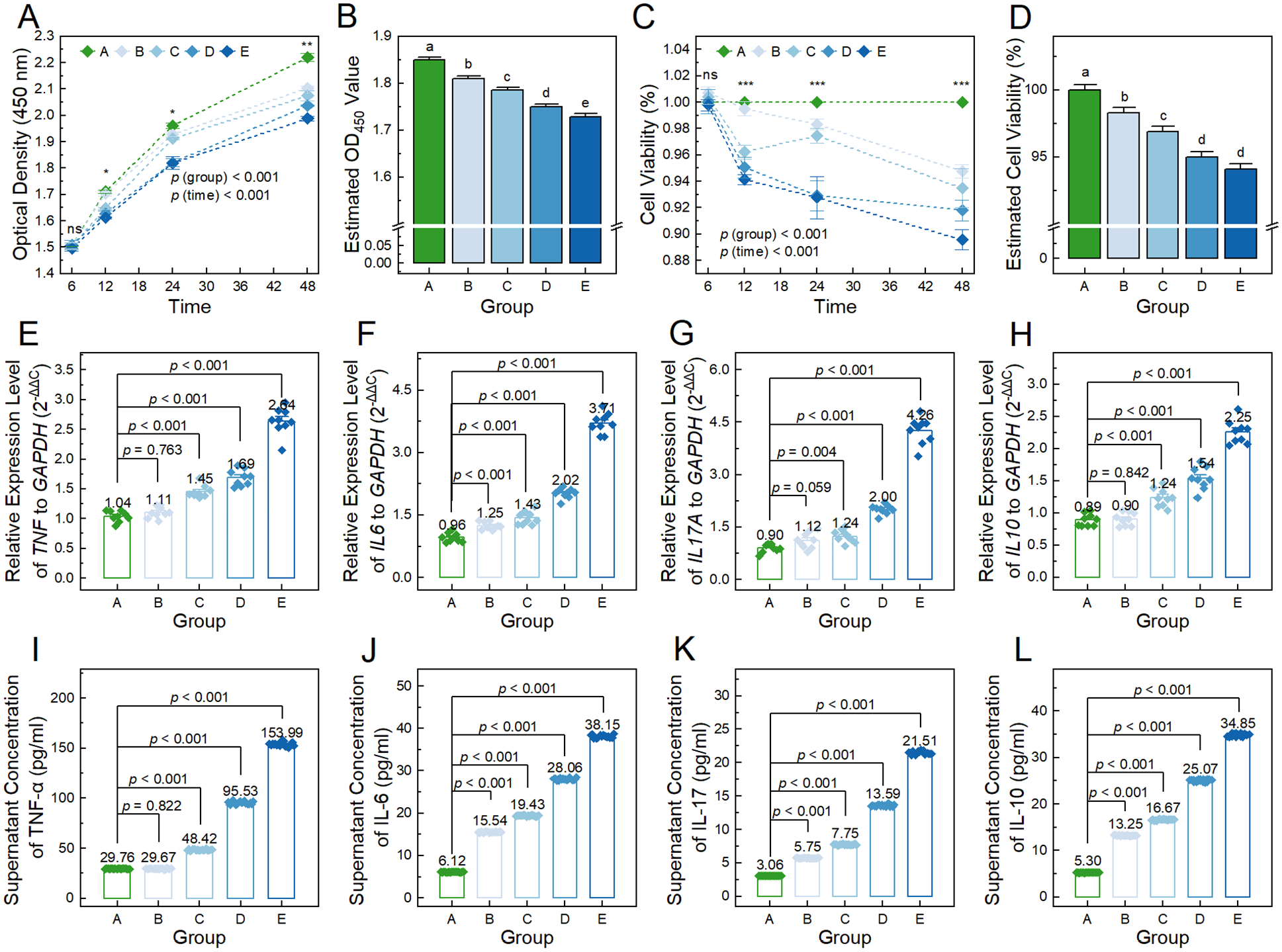
Effects of conditioned medium derived from BMSCs treated with varying concentrations of TNF-α on the proliferation capacity and cytokine secretion of ACs. (
Afterward, we evaluated the cytokine secretion levels of ACs treated with a conditioned medium derived from BMSCs treated with 0, 5, 10, 20, and 40 ng/ml of TNF-α. The relative expression level of the TNF gene in ACs at baseline condition (1.04 ± 0.03) was comparable to those in group B (1.11 ± 0.03; p = 0.763), but significantly lower than those in groups C-E (1.45 ± 0.03, 1.69 ± 0.05, and 2.64 ± 0.08, respectively) (p < 0.001;
Discussion
ACs play a crucial role in maintaining the integrity and function of articular, a specialized connective tissue that covers synovial joints, by secreting a variety of extracellular matrix components. 22 The degeneration of ACs inevitably leads to the breakdown of articular cartilage, which is the primary pathological change associated with osteoarthritis, a debilitating condition affecting millions of people worldwide.23,24 In light of this, therapies that target the modulation of the AC microenvironment, such as BMSC-based treatments, have gained attention for their potential to repair damaged cartilage and alleviate OA symptoms.25,26 Recent studies suggest that BMSCs secrete bioactive factors that can support cartilage regeneration by modulating AC activity. Moreover, emerging evidence hints at a bidirectional interaction between ACs and BMSCs, where AC secretions may also influence BMSC function. 12 However, this interaction remains underexplored. In this study, we sought to investigate this interaction by evaluating how BMSCs and ACs affect each other’s cytokine secretion in response to varying TNF-α concentrations, thus providing new insights into their potential crosstalk in the context of OA therapy.
To explore the bidirectional interaction between BMSCs and ACs, we isolated primary BMSCs and ACs from the bone marrow, femoral condyles, and tibial plateaus of rabbits (
Among the factors affecting the functions of ACs, inflammatory cytokines are considered to significantly contribute to the pathogenesis and progression of osteoarthritis.
33
Any factors that result in the infiltration and activation of local immune cells can lead to inflammatory conditions of cartilage, such as allogeneic cartilage transplantation, gout, infection, and obesity can also lead to inflammatory responses in the joints.
34
Hence, to simulate the inflammatory microenvironment in articular cartilage under varying degrees of osteoarthritis, we added 0, 5, 10, 20, and 40 ng/ml TNF-α, an important cytokine for both BMSCs and ACs,13,16,21 to the medium of BMSCs and ACs (
It is well established that secretions derived from BMSCs are capable of modulating the function of ACs,
11
and vice versa.
12
In our study, we found that the conditioned medium derived from BMSCs exposed to TNF-α significantly decreased the viability of ACs. Conversely, the conditioned medium derived from ACs exposed to TNF-α significantly elevated the viability of BMSCs (
We observed that the conditioned medium derived from ACs treated with TNF-α dose-dependently increased the relative expression levels of TNF, IL6, IL17A, and IL10 genes in BMSCs, along with corresponding increases in their protein concentrations in the medium of BMSCs (
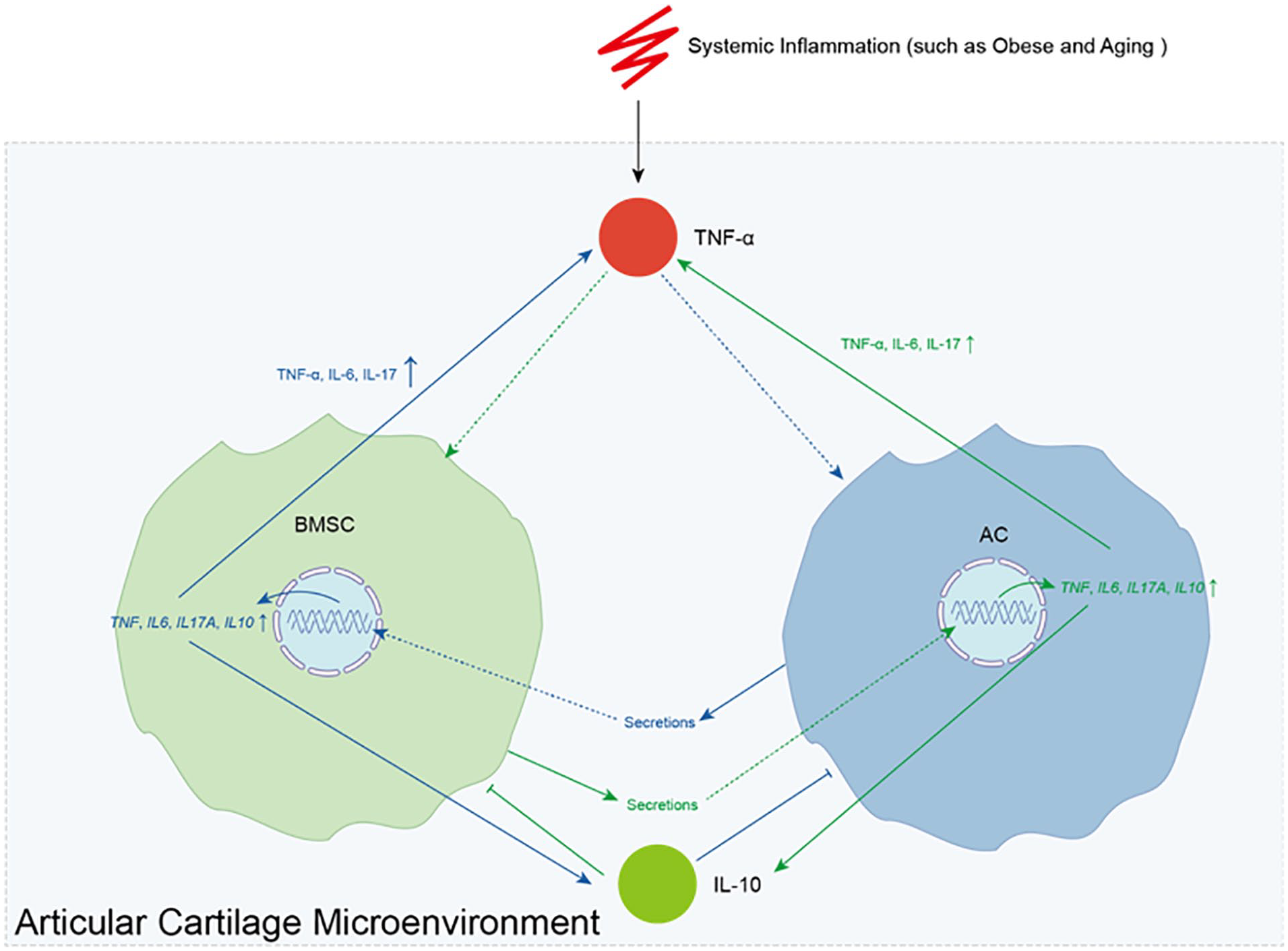
Sketch illustrating the TNF-α regulated bidirectional interaction network between BMSCs and ACs. System inflammation, such as that caused by obesity and aging, results in high levels of TNF-α in the articular cartilage microenvironment. The TNF-α in the articular cartilage microenvironment simulates BMSCs (green line) and ACs (blue line) secrete factors, which further results in elevated expression levels of TNF, IL6, IL10, and IL17A genes in ACs and BMSCs, and consequently increase the secretion of pro-inflammatory cytokines TNF-α, IL-6, and IL-17, as well as the anti-inflammatory cytokine IL-10. The secreted TNF-α promotes this circle, while IL-10 inhibits this it.
In addition, recent reports suggest local immune modulation plays a critical role in joint repair. Ilya Klabukov et al. found that xenogeneic tissue-engineered cartilage implantation induces immune cell infiltration and pro-inflammatory cytokines release, 43 while Robert Ossendorff’s findings highlight the superior outcomes of autografted cartilage in comparison to allografts. 44 These findings reinforce the hypothesis that controlling the local immune microenvironment is crucial in joint repair strategies. Therefore, in using BMSC therapies for inflammatory joint diseases, minimizing the local immune response by using autologous cells and reducing cellular stimulation should be prioritized.
In addition, it should also be noted that in our study, approximately 30 pg/ml of TNF-α and 2-6 pg/ml of IL-16, IL-10, and IL-17 were observed in the supernatant of both ACs and BMSCs treated with conditioned medium derived from BMSCs or ACs without TNF-α treatment (
This study has several limitations. First, we used rabbit BMSCs and ACs, which may not fully replicate human cell behavior, limiting direct translatability. Future research should incorporate human cells to validate our findings. Additionally, we used a traditional submerged culture, which doesn’t mimic the complex joint microenvironment. Developing 3D organoid models could improve physiological relevance. Moreover, we used only TNF-α to model inflammation, while other cytokines and enzymes like IL-1β and MMPs also play key roles in osteoarthritis. Future studies could use synovial fluid from OA patients for a more comprehensive inflammatory model. Finally, in vivo experiments using animal models (rats or rabbits) are necessary to further validate our findings in a more complex biological system.
Conclusion
In summary, our results confirmed that TNF-α in the articular cartilage microenvironment simulates BMSCs and ACs to secrete certain factors. This leads to elevated expression of TNF, IL6, IL17A, and IL10 genes, and consequently increases the secretion of pro-inflammatory cytokines TNF-α, IL-6, and IL-17, which promotes this pro-inflammatory vicious circle, as well as the anti-inflammatory cytokine IL-10, which inhibits it. Our study revealed a TNF-α regulated bidirectional interaction between BMSCs and ACs, deepening our understanding of the pathogenesis of osteoarthritis and aiding in the prevention and treatment of osteoarthritis. However, further experiments are warranted to validate this phenotype and explore its underlying mechanisms.
Footnotes
Acknowledgments and Funding
Not Applicable. The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This study was supported by the Sichuan Taikang Hospital Intramural Research Program (grant no. SCTK202308).
Author Contributions
Y.H.: Conceptualization, Data curation, Formal analysis, Investigation, Methodology, Visualization, Writing-original draft, and Writing-review & editing; F.Z.: Writing-Review & Editing; J.W.: Methodology, Validation, Investigation, and Visualization; S.W.: Supervision, Project administration, and Writing-Review & Editing. The manuscript was thoroughly examined by all authors and subsequently approved in its final version.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Ethical Approval and Consent to Participate
This study was carried out in strict accordance with the Guidelines on the Care and Use of Laboratory Animals issued by the Chinese Council on Animal Research and the Guidelines of Animal Care and received approval from the Ethical and Welfare Committee for Animal Experiments of Sichuan University (approval no: SCTK-IRB-2024-034).
Data Availability
All data and materials can be accessed from the corresponding author once requested.