
Abstract
Objective
Meanwhile, the association between osteoarthritis (OA) and type 2 diabetes mellitus (T2DM) is well known. However, it remains unclear whether vitamin K2 (vit.K2) could exert chondroprotective effects. Hence, this study investigates the interrelation between OA and T2DM under the influence of vit.K2 in chondrocytes.
Methods
Using an in vitro OA/T2DM model, articular chondrocytes were harvested from adult male Zucker diabetic fatty (ZDF) Leptfa/Crl rats, categorized as non-diabetic (heterozygous: fa/+) or diabetic (homozygous: fa/fa). Based on vit.K2 supplementation of the rats, four groups arose: control without or with vit.K2 and diabetic without or with vit.K2 supplementation. Inflammatory conditions simulating OA were induced by exposing chondrocytes to tumor necrosis factor alpha (TNFα) and C5a. Chondrocyte response was analyzed using proliferation, metabolic and wound healing assays, immunolabeling, as well as gene expression analyses.
Results
The proliferation of chondrocytes from control rats with vit.K2 supplementation was significantly higher than those without vit.K2 feeding, under both normoglycemic (NG) and hyperglycemic (HG) conditions. The wound closure ability of chondrocytes was significantly higher in the non-diabetic compared with the diabetic chondrocyte donor group. TNFα and C5a exerted catabolic effects under HG conditions by significantly inducing Tnfα gene activity in chondrocytes of control rats without vit.K2 supplementation and a significant reduction of collagen type 2 gene expression in those cells of control rats with vit.K2 supplementation.
Conclusions
The response of chondrocytes derived from non-diabetic and diabetic donors differed. The vit.K2 supply of chondrocyte donor rats exerted anabolic effects on chondrocytes.
(A) Adult healthy heterozygous (fa/+, control [Co]) and (b) diabetic homozygous (fa/fa T2DM) ZDF rats received a diet supplemented with vitamin K2 (vit.K2) or not. Chondrocytes were isolated (c) from knee joint cartilage and examined in vitro (d and e) after stimulating with normo- (f, NG) or hyperglycemic (g, HG) media and/or inflammatory cytokines (tumor necrosis factor [TNF]α, C5a). The following analyses were performed (H): metabolic and proliferative activity, cell migration, and expression of genes indicating cellular stress or differentiated chondrocytes. fa/+: heterozygous rats which bear the missense mutation in the leptin receptor gene on one allele, the other represents the wild type gene. fa/fa: homozygous rats which possess the missense mutation in the leptin receptor gene on both alleles. Scale bar = 50 µm. The images and photos were drawn and taken by the authors.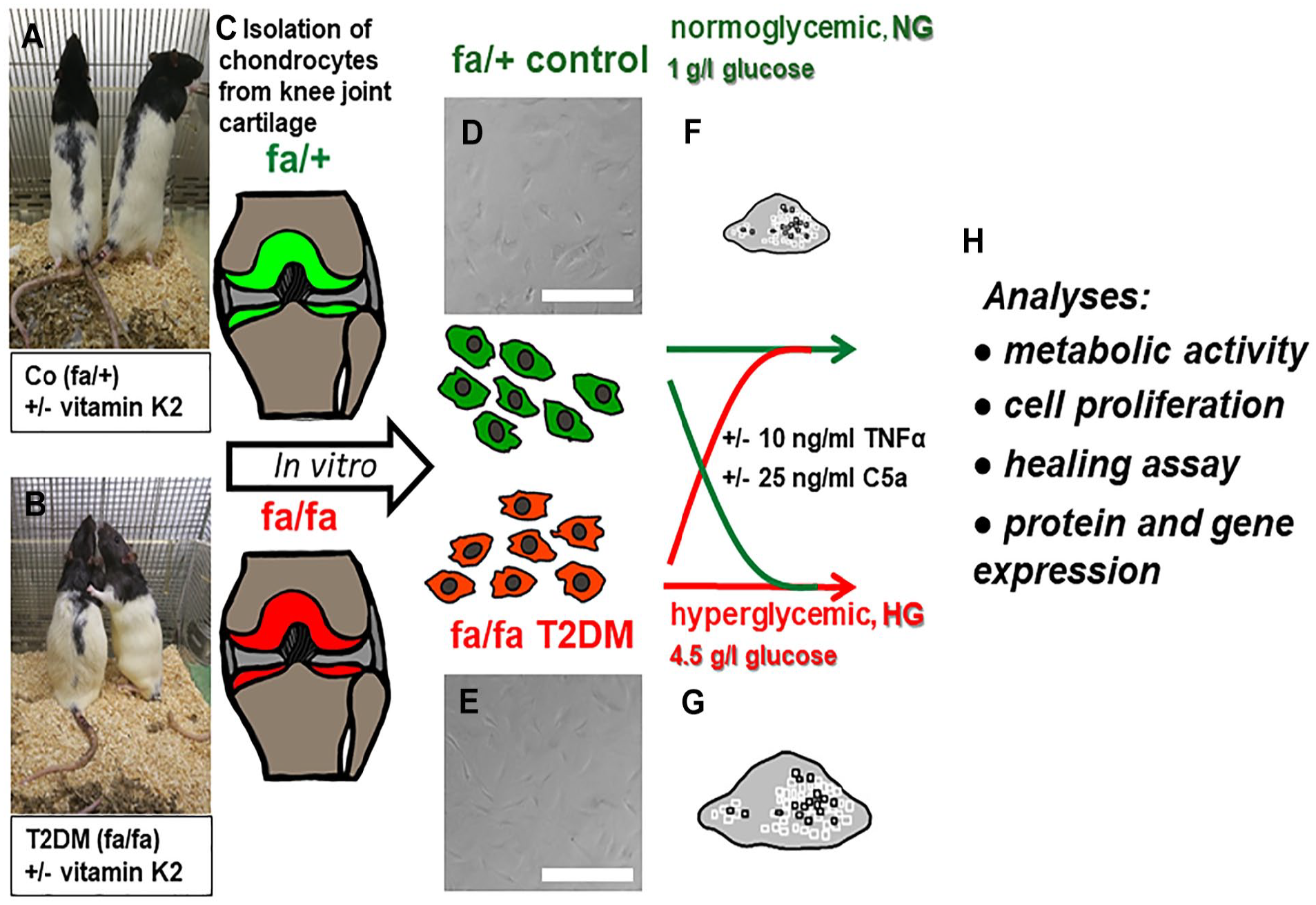
Introduction
The understanding of osteoarthritis (OA) to be a “wear and tear” disease has shifted to a more detailed understanding of the complex pathophysiology involving an intermittent local and systemic inflammatory state.1,2 Through different pathways inflammatory mediators such as Interleukin 1β and tumor necrosis factor alpha (TNFα) lead to the downregulation of extracellular matrix (ECM) synthesis, the destruction of ECM, and inhibition of proliferation of chondrocytes. 3 Meanwhile, different phenotypes of OA have been distinguished. 4
Due to high prevalence and shared risk factors such as obesity5,6 and aging, 6 OA and type 2 diabetes mellitus (T2DM) often coexist. T2DM is a metabolic disorder with an expected worldwide prevalence of 12.2% by 2045 for the population aged 20-79 years, imposing a growing economic burden on individuals and society. 7 However, many aspects of the interrelation of T2DM and OA remain unclear. This knowledge is needed to develop future therapeutic strategies in T2DM. Suitable models are required to elucidate the pathogenesis of T2DM-associated OA. Rodent models are commonly used in T2DM research. The Zucker diabetic fatty (ZDF) rat is a valid genetic model of T2DM reflecting typical features of T2DM comparable to human disease conditions.8,9 The ZDF rat has a leptin receptor defect which causes hyperphagia and obesity. 10 Rats homozygous for this mutation and exposed to a high protein diet develop T2DM, whereas heterozygous rats remain non-diabetic under similar conditions.8,11
Although there is no well-established link, there are reports that T2DM contributes to OA development through different pathways: proinflammatory cytokines, advanced glycation end products (AGEs), and oxidative or ER stress.12,13 TNFα also contributes to reduced angiogenesis in T2DM during fracture healing which could explain delayed restoration. 14 Meanwhile, also unrestrained complement activation has been implicated in OA pathogenesis.15,16 Particularly, the cleavage of the complement components C3 and C5 which is associated with the release of proinflammatory C3a and C5a as so-called anaphylatoxins has been attributed to OA.17,18
Vitamin K2 (vit.K2) comprises a group of menaquinone derivates (MK4-MK13), 19 first identified in 1935. 20 It is widely accepted that K vitamins are essential for the regulation of blood coagulation factors. 21 Emerging research suggests extensive involvement of vit.K2 in various systems of the body, influencing physiological and pathological functions, 21 particularly in regulating calcium metabolism. 22 Vit.K2 serves as a carboxylation cofactor for vit.K-dependent proteins (VKDP) such as osteocalcin (OC) and matrix Gla protein. 22 The carboxylated matrix Gla protein inhibits tissue calcification,23,24 and the carboxylated matrix Gla protein is lesser expressed in OA cartilage. 25 Cartilage calcification is an unwanted feature in OA. 17 Vit.K influences the balance between osteoblasts and osteoclasts leading to low bone density in cases of vit.K deficiency. 26 OA, especially in the knee, has been linked to vit.K deficiency by Misra and Azuma et al.27,28 Notably, a vit.K2 analog, inhibits matrix metalloproteinases (MMPs) in a tumor cell line 29 responsible for matrix degradation in OA.2,3 Moreover, vit.K2 (MK-7) is enhancing glycemic homeostasis and insulin sensitivity. 30 However, data on the effects of vit.K on chondrocytes remain rudimentary. The influence of vit.K1 on mouse cartilage tissue was tested by Shea et al. 31 indicating that an insufficient vit.K1 (phyllochinone) supply led to a loss of proteoglycans. In a recently published combined in vitro/in vivo study vit.K2 exerted the capacity to counteract features of OA by reducing chondrocyte ferroptosis and ECM degradation. 32
Therefore, we investigated the impact of MK7 on articular chondrocytes using a combined T2DM-OA model aiming to evaluate its putative therapeutical potential. The hypothesis of the present study was that vit.K2 feeding of diabetic rats which served as chondrocyte donors might reduce catabolic effects in a subsequent chondrocyte-based in vitro OA model.
Materials and Methods
Study Design
In this research project, adult male ZDF Leprfa/Crl rats were used as cell donors with a mean body weight of 400.7±23.4 g at finalization. These rats, either with a heterozygous (fa/+) or homozygous (fa/fa) genotype (Charles River Laboratories Inc., Chatillon-sur-Chalaronne, France), carry a leptin receptor gene mutation leading to hyperphagia and obesity. 10 T2DM was induced in fa/fa rats by feeding them a diabetogenic diet (Purina 5008, SSniff Spezialdiäten GmbH, Soest, Germany). Heterozygous rats served as controls, since they do not develop T2DM (Supplementary Table 1). Cartilage tissue was harvested post mortem. The 20 donor animals were finalized in another animal project conducted in accordance with the Federation of European Laboratory Animal Science Associations (FELASA) guidelines. This project was also approved by the institutional internal review board and local review board (Regierung Unterfranken, Bavaria, RUF 55.2.2-2532-2-729-17). The rats were introduced into the facility at the age of 71-82 days with 2 or 3 individuals per cage, randomized into groups, and allowed to acclimatize for 2 weeks. The rats were housed under standardized conditions (55% humidity, 21±2°C, Memmert GmbH and co.KG, Schwabach, Germany). Rats were checked daily by a veterinarian. Subsequently, they were divided into four groups two with fa/fa and two with fa/+ rats (Supplementary Table 1, n = 3-8), all fed with Purina 5008 diet. One group of each genotype was supplemented with 100 mg/kg vit.K2 (MK7) in the diet, generously provided by Kappa Bioscience AS (Oslo, Norway). Supplementation started when the rats were 14-15 weeks old and continued until sacrifice at week 26-27. Blood glucose levels were monitored using AlphaTrak 2 glucometers (Zoetis, Parsippany-Troy Hills Township, NJ, USA).
Chondrocyte Isolation
After euthanasia, the knee joint cartilage of each animal was scraped off, rinsed with phosphate buffered saline (PBS, PAN-Biotech GmbH, Aidenbach, Germany) and cartilage slices were treated overnight with collagenase (1 mg/ml, Nordmark, Uetersen Germany) suspended in culture medium (CM) composed of Dulbecco’s modified Eagle’s medium (DMEM)/F12 with 3.15 g/l glucose, supplemented with 50 IU/ml amphotericin B, and 0.5% (v/v) 100× non-essential amino acid mix, 50 IU/ml penicillin/streptomycin (all: PAN-Biotech GmbH), and 25 mg/ml ascorbic acid (Sigma-Aldrich, St. Louis, MO, USA). 33 The cartilage/collagenase mixture was agitated on a rotary shaker within an incubator set at 37°C and 5% CO2. The next day, the mixture was filtered through a 100-µm pore size cell sieve (TPP, Trasadingen, Switzerland). The isolated cells were washed once with PBS and transferred into T25 cell culture flasks (CellPlus, Sarstedt AG, Nümbrecht, Germany) containing 5 ml CM supplemented with 10% fetal bovine serum (FBS, PAN-Biotech GmbH). The CM was replaced three times a week and cells were allowed to grow until 80% confluence. At this point, cells were either trypsinized and cryopreserved in cryoprotection medium (10% dimethylsulfoxide [DMSO] in FBS) in liquid nitrogen for future use or expanded in T175 cell culture flasks.
For all following experiments, cells were seeded into the corresponding flasks or well plates for 24 hours in CM with 10% FBS. The CM was discarded, and cells were washed once with PBS before being starved for 1 hour using CM (DMEM/F12) with 1% FBS either as normoglycemic (NG) medium with 1 g/l glucose or hyperglycemic (HG) medium with 4.5 g/l glucose. The latter should simulate T2DM conditions. Following starvation, the NG or HG medium was removed, and NG and HG medium with 1% FBS or NG and HG stimulation media containing 1% FBS, 10 ng/ml TNFα (Peprotech GmbH, Hamburg, Germany), 25 ng/ml C5a (R&D Systems, Minneapolis, MN, USA), or a combination of both were added for 24 hours if not otherwise stated. TNFα and the complement split fragment anaphylatoxin C5a were added to simulate inflammatory conditions of OA in vitro.
Metabolic alamarBlue™ Assay
A total of 10,000 chondrocytes/well (34,483 cells/cm2) were seeded into 96-well plates (Sarstedt AG). The medium volume in each well was 100 µl during stimulation, and an additional 25 µl of alamarBlueTM solution (ThermoFisher Scientific Inc., Waltham, USA) was added. The metabolic activity was measured after 6 hours at 600 nm for oxidized and 570 nm for reduced resazurin using a plate reader (Tecan Austria GmbH, Grödig, Austria). All samples were measured in triplicates, and three to five independent experiments were conducted for each experimental group with chondrocytes of different donor rats as indicated in each figure legend.
Measurement of Cell Proliferation Using CyQUANT Cytotoxicity Assay
Initially, 5,000 chondrocytes/well (17,241 cells/cm2) were seeded into 96-well plates and in 100 µl of CM as described above. Then, the cells were incubated with the respective stimulation media for 48 hours, and the DNA content was measured following the CyQUANT Cytotoxicity Assay (ThermoFisher Scientific Inc.) protocol. The fluorescence was measured using λEx of 485 nm and λEm of 530 nm in a TECAN plate reader (Tecan Austria GmbH). All samples and each concentration of the standard were tested and measured in triplicates.
Wound Healing Assay
A total of 300,000 chondrocytes/well (33,076 cells/cm2) were seeded into 6-well plates (Sarstedt AG). After 24 hours of incubation, a scratch was made in the cell monolayer using a 1 ml Eppendorf pipet tip (Sarstedt AG). All wells were rinsed with PBS, and CM with 10% FBS was added immediately. Cell migration was monitored using a Cellwatcher M microscope (PHIO scientific GmbH, Munich, Germany), placed inside the incubator, capturing one picture of each well every 30 minutes over a 24-hour period. Wound closure and cell motility were analyzed using PHIOme Data Management and Analysis Platform Software Add-Ons Version 1.4.2 (PHIO scientific GmbH). The scratch width was measured automatically by the PHIOme software add-on “Wound healing.” The average velocity of wound closure was determined automatically using the PHIOme software add-on “Motility.”
Scratch Assay Using the Confocal Laser Scanning Microscope
A total of 40,000 cells/well (21,978 cells/cm2) were seeded into 24-well plates (Sarstedt AG) and cultured in 1 ml CM as described above. A scratch was made on the cell monolayers using a 1 ml pipet tip (Sarstedt AG). CM was removed, and cells were washed with PBS and starved during this time, scratches were marked and photographed at three stable positions using a confocal laser scanning microscope (SPEII DMi8 Leica, Wetzlar, Germany). After the starvation period, cells were stimulated as mentioned above. Following 24 hours of stimulation at 37°C with 5% CO2, scratches were photographed again in their original three stable positions. Differences in the scratch area after 24 hours were calculated using ImageJ Software 1.52d (National Institute of Health, MD, USA).
Immunofluorescence Staining
A total of 20,000 chondrocytes were seeded onto a 12 mm cover slip (17,683 cells/cm2) (VWR International GmbH, Darmstadt, Germany) in a 24-well plate as described above. After stimulation, the cover slips were fixed using 4% paraformaldehyde (PFA, Santa-Cruz Biotechnology, Inc. Dallas, TX, USA) in PBS. For immunolabeling, the cover slips were rinsed three times with tris-buffered saline (TBS) and incubated for 20 minutes with blocking solution (5% donkey serum, 0.1% Triton X-100 in TBS) to reduce nonspecific background fluorescence. Blocking solution was replaced by antibodies against collagen type 1 alpha1 chain (goat anti-human, Southern Biotech, Birmingham, AL, USA) and collagen type 2 (rabbit anti-human, Origene, Rockville, Maryland, USA) diluted 1:50 in blocking solution. After a 24-hour incubation period, antibodies were washed off three times using TBS, and secondary antibodies (Cyanine [cy]3-dye conjugated donkey anti-goat, and AlexaFluor488-dye conjugated donkey anti-rabbit [both: Invitrogen, Carlsbad, CA, USA]) and 4,’6-diamidine-2-phenylindole-dihydrochloride (DAPI [Roche, Mannheim, Germany]) were applied in 1:200 dilutions in blocking solution. After 1 hour, the secondary antibodies were washed off three times using TBS, and cover slips were mounted using Fluoromount (ThermoFisher Scientific Inc.). Pictures were taken with the confocal laser scanning microscope (SPEII with DMi8, Leica Microsystems, Wetzlar, Germany). Image analysis was performed with ImageJ Software 1.52d from three representative photos per cover slip. Therefore, the “mean” intensity (for red and green channels separately) of the picture was divided by the number of cells in the picture (as indicated by DAPI nuclei staining).
Gene Expression Analysis Using Real-Time Detection Polymerase Chain reaction (PCR)
A total of 250,000 chondrocytes were seeded in T25 tissue culture flasks (10,000 cells/cm2) containing 3 ml of CM as described above. After the stimulation period (4 or 24 hours), cells were washed with PBS and lysed using RLT-Buffer (Qiagen, Venlo, Netherlands) containing 1% mercaptoethanol (Sigma-Aldrich, St. Louis, MO, USA). RNA was purified and then transcribed into complementary DNA (cDNA) following the instructions of the manufacturer of the Qiagen miniprep kit. The resulting cDNA, at a concentration of 11.7 ng/µl, was subjected to amplification using a StepOnePlus Real-Time PCR System (ThermoFisher Scientific Inc.) and specific primer pairs (ThermoFisher Scientific Inc.) targeting the genes for tumor necrosis factor (Tnf)α, interleukin (IL) 6, Suppressor of cytokine signaling (Socs)1 and Socs3, collagen type 1 alpha1 chain (Col1a1), collagen type 2 alpha1 chain (Col2a1), as well as heme oxygenase 1 (Hmox1), which were tested against hypoxanthine phosphoribosyltransferase 1 (Hprt1) as an endogenous control (see Supplementary Table 2).
Statistical Analysis
For statistical analysis, means of triplicates were analyzed in GraphPad (GraphPad Software Inc., San Diego, CA, USA). For metabolic and proliferative activity, immunolabeling, endpoint scratch assay, and gene expression analysis, a two-way analysis of variance (ANOVA) with Tukey’s multiple comparison as a post hoc test was used. For migration and motility an unpaired t-test was performed. Each assay was performed using at least three independent experiments. Displayed in the graphs are always mean values ± standard deviation. The symbols indicate the single measured values of the independent experiment. A p value of * ≤ 0.05 and a p value of *** ≤ 0.001 were set at statistical significance for ANOVA and a p value of # ≤ 0.05 for t-test.
Results
Metabolic Activity
The metabolic activity of chondrocytes derived from control and the T2DM rat groups did not significantly differ. The differences between the chondrocyte donor groups without and with vit.K2 supplementation did not reach the statistical significance level. This applies to both the control and the T2DM groups and the different glucose conditions of chondrocyte culturing (NG vs. HG). The stimulation of chondrocytes with both, TNFα and/or C5a had no statistically significant effect (
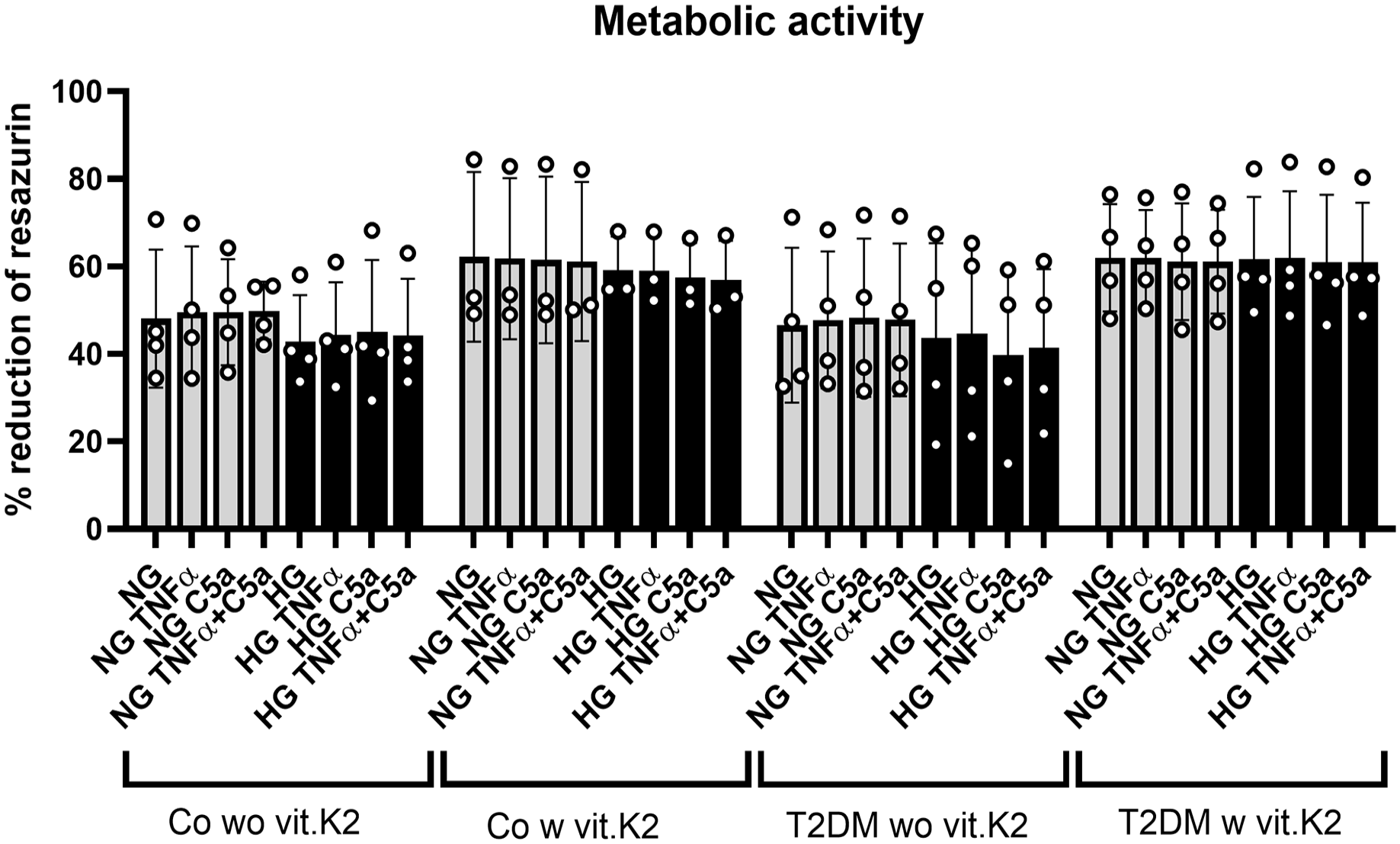
Metabolic activity of articular chondrocytes derived from non-diabetic and diabetic ZDF rats with or without vitamin (vit.) K2 supplementation measured by alamarBlueTM assay after exposure of 6 hours to normoglycemic (NG) or hyperglycemic (HG) conditions with or without tumor necrosis factor (TNF)α and/or the complement split fragment C5a. The metabolic activity detectable as resazurin reduction was measured 6 hours after resazurin addition. Gray bars represent media containing a glucose concentration of 1 g/l simulating NG conditions, while black bars represent media containing a glucose concentration of 4.5 g/l simulating HG T2DM conditions. The TNFα concentration was 10 ng/ml, the C5a concentration was 25 ng/ml. n = 3-4 biological chondrocyte donors (rats) were included in independent experiments (circles show results of independent experiments). Articular chondrocytes derived from non-diabetic ZDF rats without vit.K2 (Co wo vit.K2) and with vit.K2 (Co w vit.K2) and diabetic ZDF rats without vit.K2 (T2DM wo ut vit.K2 (T2DM wo vit.K2) as well as with vit.K2 (T2DM w vit.K2) supplementation. Rats “w vit.K2” received a diet which was supplemented with 100 mg/kg vit.K2 (menaquinone 7). Mean values and standard deviations are shown. No significant differences could be detected after two-way ANOVA multiple comparison.
Proliferative Activity Based on DNA Content
The proliferative activity of the chondrocytes deriving from non-diabetic rats fed with vit.K2 was significantly higher when comparing the proliferation of chondrocytes from rats of the control groups fed either a diet without or with vit.K2 supplementation (NG with vs. without vit.K2, HG [both p < 0.001], NG+C5a [p < 0.05],
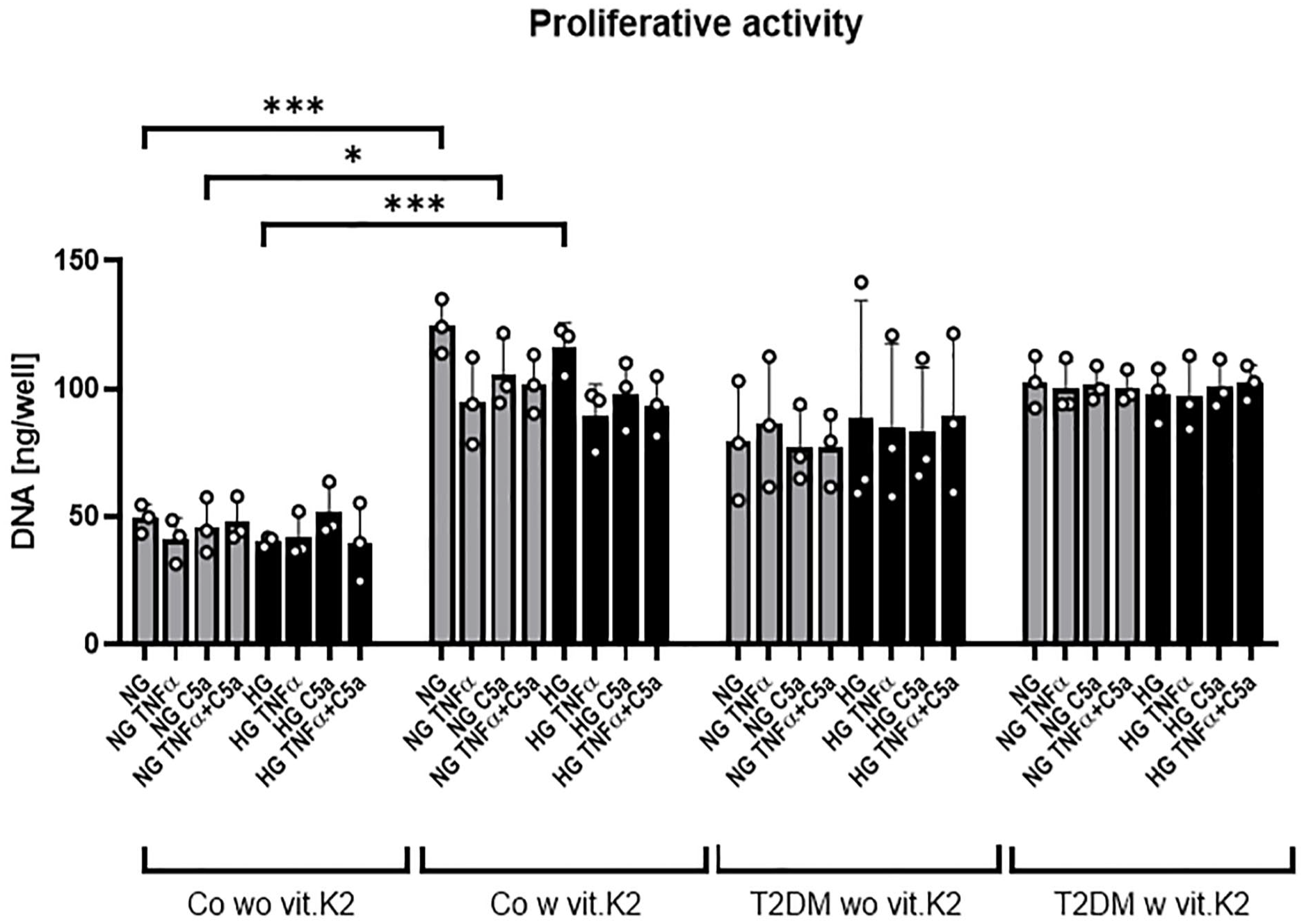
Proliferative activity of articular chondrocytes derived from non-diabetic and diabetic ZDF rats fed or not with vitamin (vit.) K2 measured after 48 hours exposure to normo- (NG) or hyperglycemic (HG) conditions and/or tumor necrosis factor TNFα and complement split fragment C5a using the CyQUANT Cytotoxicity Assay. Gray bars represent media containing a glucose concentration of 1 g/l simulating normoglycemic (NG) conditions while black bars represent media containing a glucose concentration of 4.5 g/l simulating T2DM HG conditions. TNFα concentration was 10 ng/ml, C5a concentration was 25 ng/ml. n = 3 chondrocytes of three different rats were included in independent experiments (circles show results of independent experiments). Articular chondrocytes derived from non-diabetic ZDF rats without vit.K2 (Co wo vit.K2) and with vit.K2 (Co w vit.K2) and diabetic ZDF rats without vit.K2 (T2DM wo vit.K2) and with vit.K2 (T2DM w vit.K2) supplementation. Chondrocyte donor rats (w vit.K2) received a diet which was supplemented with 100 mg/kg vit.K2 (menaquinone 7). Mean values and standard deviations are shown. Data were analyzed with two-way ANOVA multiple comparison with *p ≤ 0.05; ***p ≤ 0.001.
Cell Migration and Motility
Cell movement can be divided into two different types. The targeted movement or so-called migration was tested with the wound healing assay. Here, the cells migrate specifically in the direction of the injury inserted into the cell layer. As the cells were closing the introduced wound, it will be referred to as “closure.” The other type is the so-called motility, where the cells are moving randomly without a directed course.
The wound closure by chondrocytes derived from control rats was significantly faster than that of chondrocytes from T2DM rats. The scratch width difference after 20 hours of incubation was 0.46 ± 0.19 mm for the control groups (without/with vit.K2) compared with 0.28 ± 0.06 mm for the T2DM groups (without/with vit.K2) (p < 0.03). Chondrocytes deriving from rats supplemented with vit.K2 (Co/T2DM) were closing the wound faster (0.42 ± 0.19 mm) compared with those without vit.K2 (0.31 ± 0.08 mm after 20 hours), but the difference was not significant. The slope of the control without vit.K2 group was significantly higher (unpaired t-test; p = 0.0402) than the slope of the T2DM without vit.K2 group (
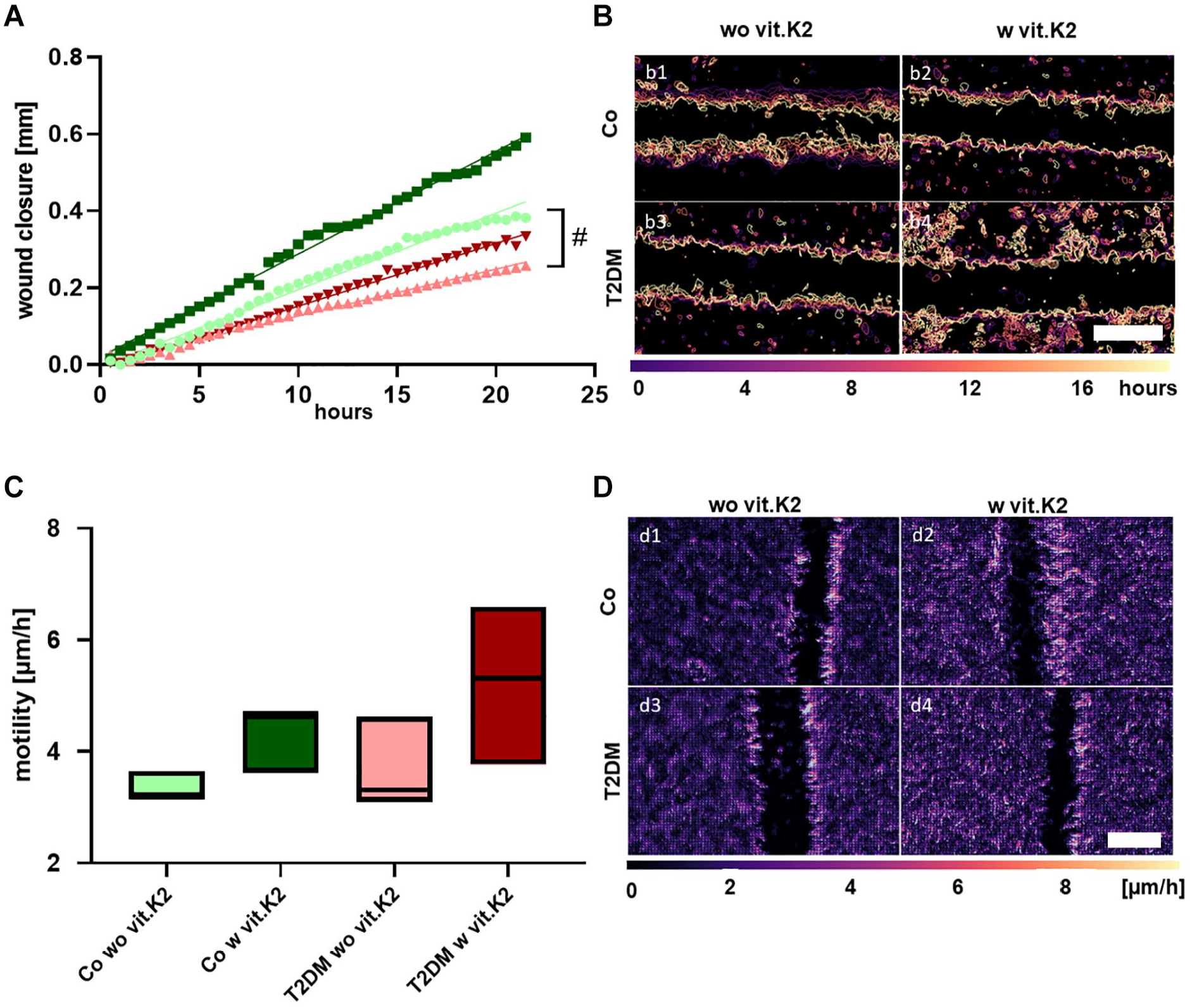
Time lapse microscopic monitoring of the scratch closure (
Immunofluorescence Staining
Chondrocytes were immunolabeled using antibodies against collagen type 1 alpha1 and collagen type 2 to show differences between the four groups in the immunoreactivity of collagens. Some chondrocytes showed mainly collagen type 2 immunoreactivity, while others displayed collagen type 1 synthesis (
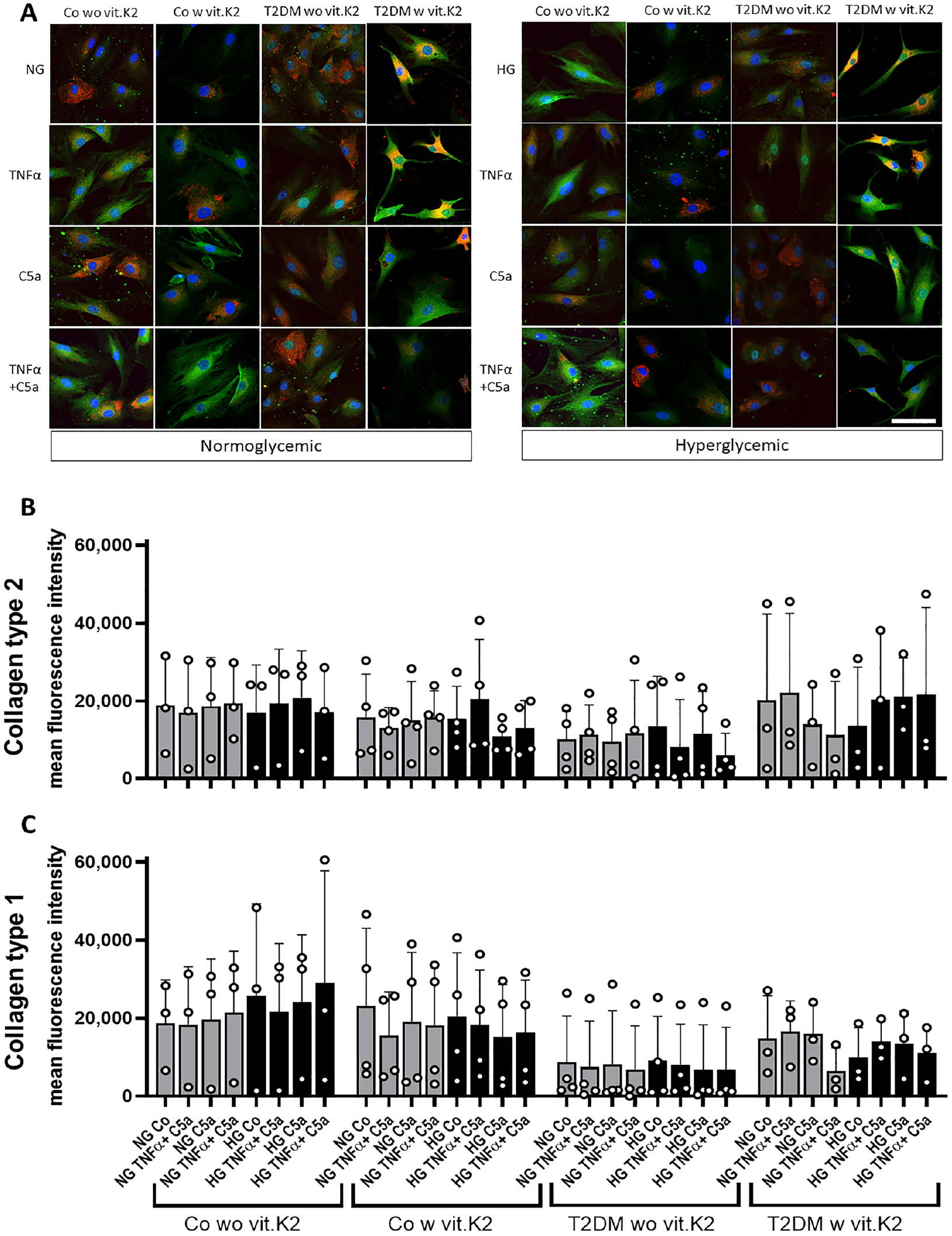
Representative images of immunolabeled chondrocytes (
Gene Expression
After 4 hours, Tnfα and IL6 gene expression was analyzed. There was a significant induction of Tnfα gene expression in chondrocytes of the non-diabetic control group without vit.K2 supplementation cultured under HG conditions when treated with TNFα and C5a. IL6 gene expression was significantly higher induced by C5a and TNFα under NG compared to HG conditions in chondrocytes of T2DM rats treated with vit.K2 (
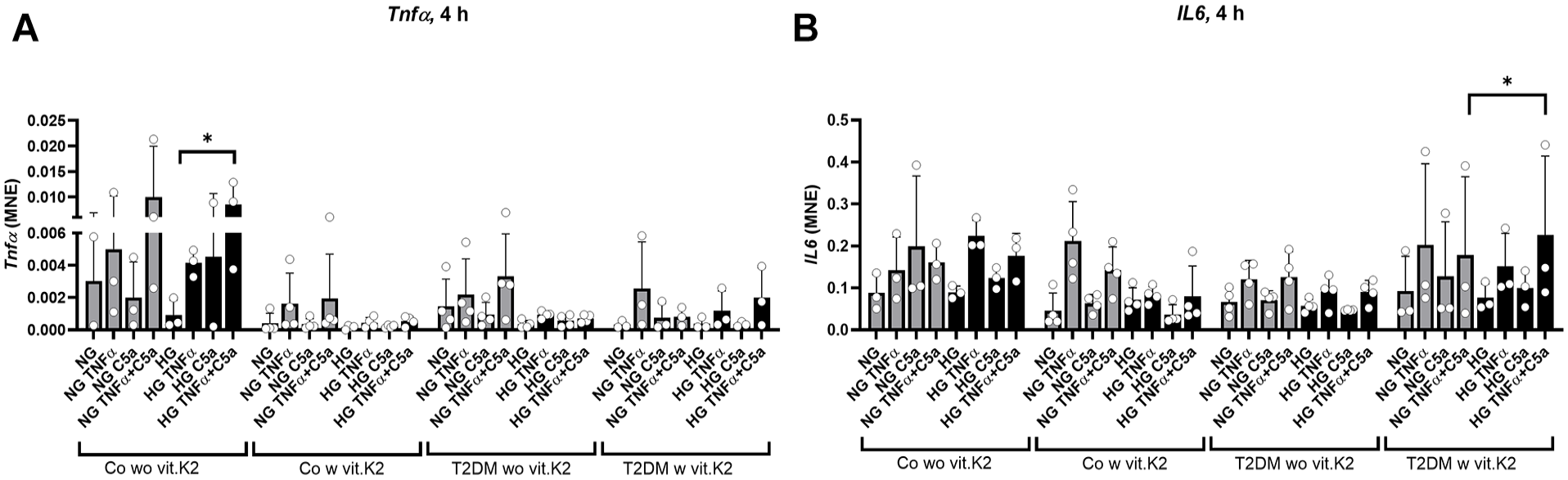
Mean normalized gene expression (MNE) of tumor necrosis factor (Tnf)α and interleukin (IL)6 normalized against hypoxanthine phosphoribosyltransferase 1 (Hprt1) after 4 hours of stimulating chondrocytes with 10 ng/ml TNFα, 25 ng/ml C5a or the combination of both in normoglycemic (NG) and hyperglycemic (HG) media. Shown are mean values with standard deviations (SD) of three to four independent experiments for each group (circles show results of independent experiments). (
At 4 and 24 hours of stimulation, the expression of Hmox1 was generally lower expressed in the chondrocytes derived from rats fed with vit.K2 in both—control and T2DM groups. At this time a difference was observed between the control without vit.K2 and the control with vit.K2 group even though it did not reach statistical significance. Additionally, there was a trend at the time of 24 hours in the groups without vit.K2 (Co and T2DM): TNFα increased the expression of Hmox1 in chondrocytes from animals without vit.K2 supplementation in both, NG and HG media, while this trend was barely detectable in samples of rats which received vit.K2 in their diet. The combination of TNFα and C5a elevated the Hmox1 expression only in chondrocytes of the non-diabetic controls without vit.K2. In the control group without vit.K2 the mean normalized expression (MNE) values in the HG stimulations were more than twofold elevated compared with the NG conditions (
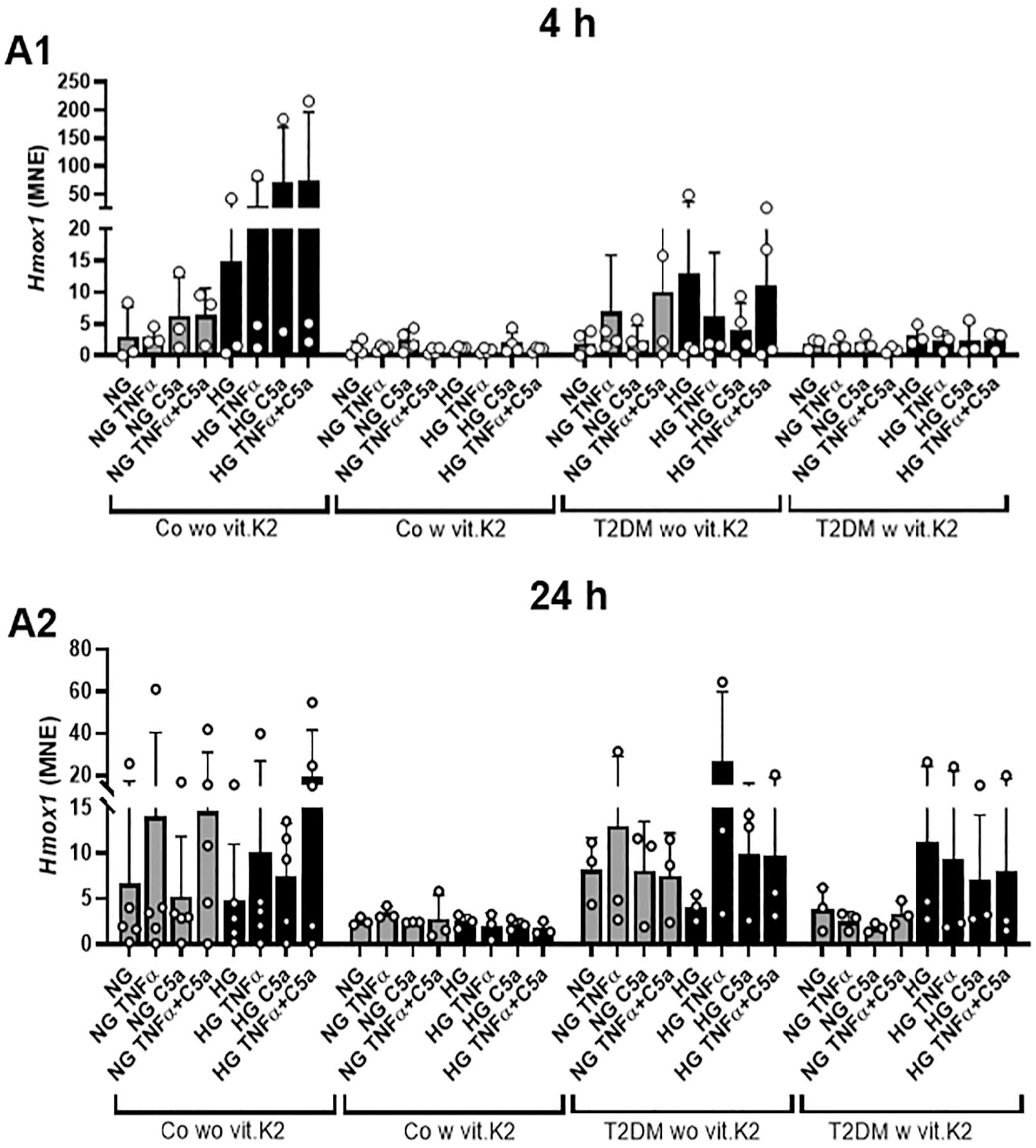
Mean normalized expression (MNE) of heme oxygenase (Hmox)1 normalized against hypoxanthine phosphoribosyltransferase 1 (Hprt1) after 4 hours (
Chondrocytes derived from non-diabetic and diabetic rats, fed with a vit.K2 supplemented diet showed a trend of lower expression of Socs1 compared with chondrocytes of both rat groups without vit.K2 supplementation at both time points of investigation (4 and 24 hours). There was no relevant difference between HG and NG groups, irrespective of stimulation with TNFα and/or C5a. TNFα, also in combination with C5a led to some induction in chondrocytes of rats which received no additional vit.K2 (4 hours).
Socs3 was mostly lesser expressed in chondrocytes derived from vit.K2 supplemented animals compared to those deriving from rats without vit.K2 supplementation at both investigation time points, although all of the HG-stimulated chondrocytes deriving from the control with vit.K2 animals expressed Socs3 at a level comparable to that of control without vit.K2 and T2DM without vit.K2 groups (Co without vit.K2 and T2DM without vit.K2 [
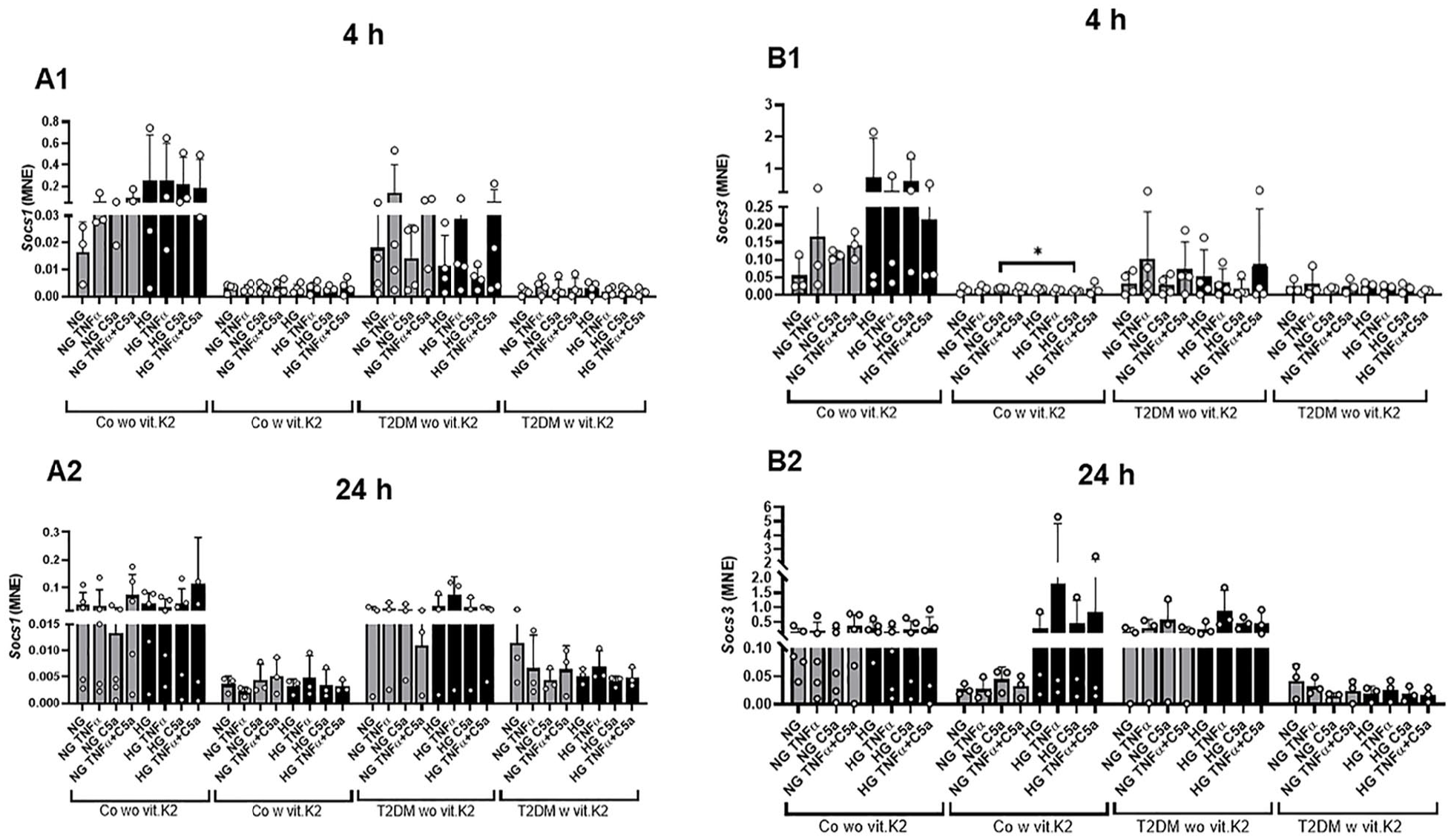
Mean normalized expression (MNE) of suppressors of cytokine signaling (Socs)1 (
Col2a1 expression was higher in the T2DM without vit.K2 group compared to the others even though statistically significant levels were not reached. Stimulation with TNFα or the combination of TNFα and C5a reduced Col2a1 expression compared with the corresponding NG and HG values. Significantly lower expression of Col2a1 was observed in NG TNFα compared to HG C5a and HG TNFα and C5a compared to HG CM stimulation in chondrocytes from control rats supplemented with vit.K2 (both p < 0.05). Col1a1 seemed to be slightly higher expressed in the control with vit.K2 group and the NG value in each group was the highest, often followed by the HG value, while the addition of TNFα and C5a led to reduced Col1a1 expression. Statistically significant suppression of Col1a1 (p < 0.05) was detected when comparing the NG TNFα or NG C5a with the NG control in chondrocytes of heterozygous, vit.K2 fed rats (
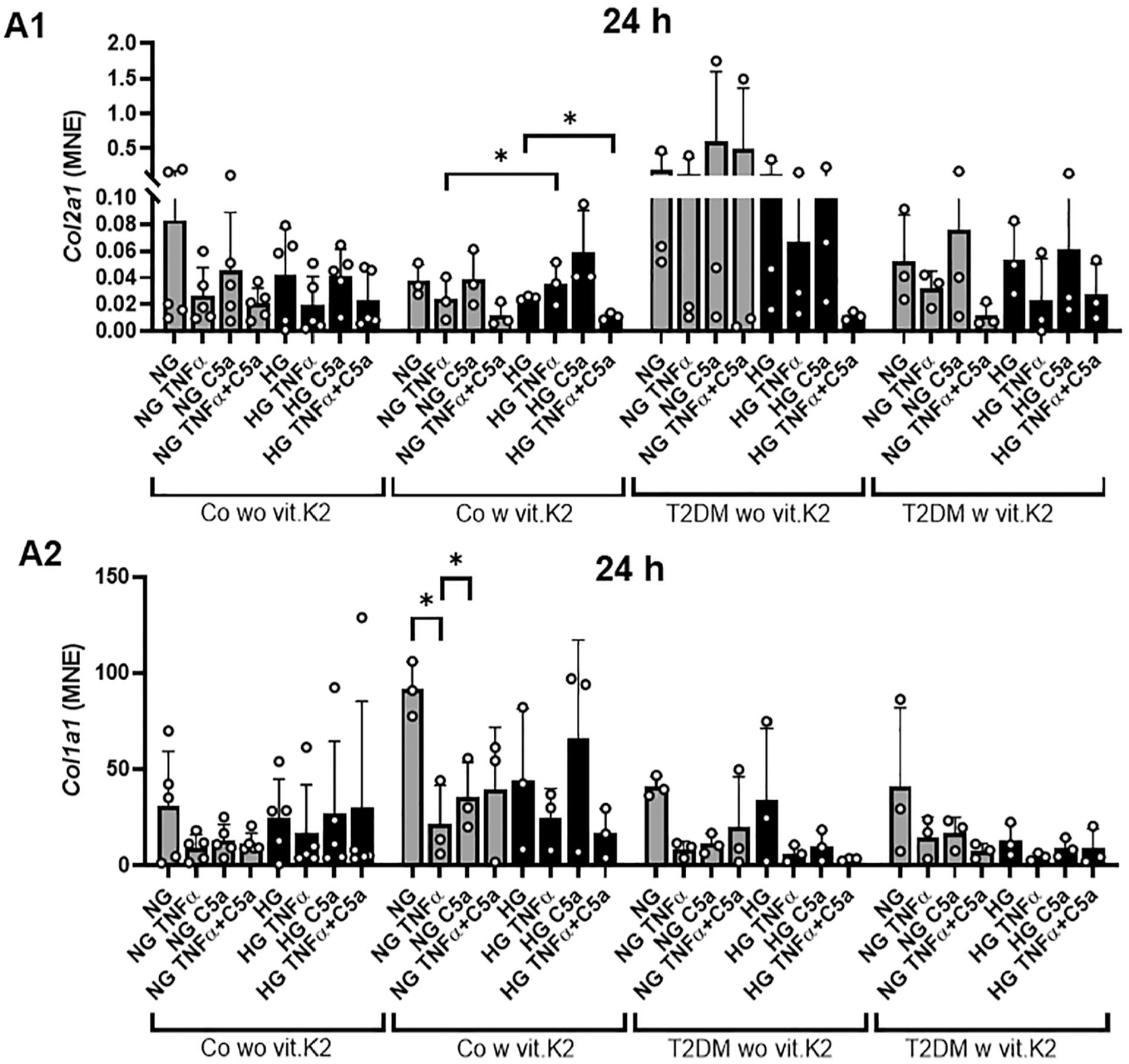
Mean normalized expression (MNE) of collagen type 2 (Col2a1,
Discussion
The main findings of the present study are (1) a statistically significant elevated proliferation in chondrocytes from non-diabetic rats induced by vit.K2 feeding. Moreover, a trend of increased proliferation in chondrocytes of diabetic rats compared with those of non-diabetic rats which showed no statistically significant further increase by vit.K2 supplementation of donor animals. (2) A significantly lower wound closure ability was detected in chondrocytes from diabetic compared with non-diabetic rats. (3) A suppression of Socs1 and Socs3 gene expression in chondrocytes of animals supplemented with vit.K2.
The observed effect of vit.K2 on chondrocyte proliferation is in agreement with the observation of a stimulatory effect of vit.K2 on mesenchymal stem cells and skeletal muscle satellite cell proliferation. 34 The proliferation could be enhanced by induction of vit.K-dependent matrix Gla protein expression since another study demonstrated the association of matrix Gla protein with chondrocyte proliferation. 35 A higher proliferation rate could also explain the slight trend of elevated metabolic activity observed in the chondrocytes from the rats supplemented with vit.K2. As the chondrocytes were allowed to adhere and grow for 24 hours before the stimulation and measurement of metabolic activity started, the chondrocytes from the with vit.K2 groups were—due to higher proliferative activity—probably already exceeding the chondrocytes from the groups of donor rats without vit.K2 supplementation in numbers. Other studies showed both, a significantly higher metabolic activity and a corresponding trend for higher proliferative activity in human chondrocytes in NG compared with HG media.33,36 This could be explained at least in parts by lower glucose uptake of cells exposed to increasing glucose concentration. 37 Like other cell types chondrocytes are able to migrate. 38 Since this property reflects repair capability chondrocyte wound healing assays were used by many other researchers in cartilage research.39 -42
The wound healing assay showed a significantly higher velocity of scratch closure by the chondrocytes from the control compared with the T2DM without the vit.K2 group. It is well known that healing processes are compromised by T2DM, for example, fracture healing requiring a balanced transition from cartilage to bone. 43 A strong trend of faster growth into the scratch was measured in those chondrocyte cultures derived from rat groups supplemented with vit.K2 compared with those which received no vit.K2 supplementation. A stimulatory effect on cell migration by vit.K2 was also described for endothelial 44 and bovine skeletal muscle cells. 34 As the chondrocytes from T2DM rats showed a trend toward a higher proliferative activity, the faster closure of the wound observed in the control groups could not be caused by higher proliferation but by directed migration.
The trend of overall higher undirected motility, that is seen in the chondrocytes of T2DM rats and the groups that were supplemented with vit.K2, could be caused by a higher proliferation in those groups, as the cells arising after mitosis from the dividing original cell need to rearrange with their neighboring cells and therefore, move around with no particular direction. In mouse vascular smooth muscle cells activated insulin receptor isotypes mediate migration. 45 The migration of bovine chondrocytes was stimulated by the insulin-like growth factor in a similar manner. 46 Chondrocytes from diabetic and obese ZDF rats might exert insulin resistance 47 and hence, the lacking efficacy of insulin might explain the impaired directed migration and enhanced proliferation. 47
Chondrocytes of the four donor groups differed also in terms of their gene expression of cytoprotective and cytokine regulatory factors. The transcriptional activity of anti-oxidative enzyme gene Hmox1 and the two cytokine negative feedback regulator Socs genes of chondrocytes varied more depending on whether they originate from donor rats with or without vit.K2 supplementation, compared with the genotype. T2DM increases oxidative stress 48 and the oxidative stress response gene Hmox1 is upregulated in individuals suffering from T2DM as a defense mechanism against this oxidative stress,49,50 but also in OA. 51 Accordingly, in our study TNFα stimulation correlated with inflammatory conditions since TNFα alone or in combination with C5a induced as a strong trend its own gene expression and that of IL6 as demonstrated after 4 hours. This induction of Tnfα gene activity by TNFα and C5a was significant for HG conditions in chondrocytes of non-diabetic control rats. TNFα upregulated Hmox1 expression in chondrocytes of control and T2DM rat groups without vit.K2 supplementation as observed after 24 hours. Notably, the Hmox1 gene was generally lower expressed in chondrocytes of vit.K2 treated rats compared with those derived from donors without vit.K2 supplementation at both investigated time points suggesting an early effect persisting over 24 hours. This is in agreement with the study of Wang et al. 52 who found that the induction of Hmox1 expression by lipopolysaccharide (LPS) was suppressed by vit.K2 in a mice lung injury model. Hence, the vit.K2 uptake may either alter chondrocyte response to inflammation or more probably chondrocytes experience lesser stress and inflammation due to the anti-oxidative effects of vit.K2 52 which does not require regulation by these cytokine and stress response regulators. A similar gene expression pattern was observed for Socs1, known to alleviate oxidative stress in diabetes. 53 Socs3, which is in a similar manner to Socs1 linked to insulin signaling mediating inflammation-induced insulin resistance (review [54]), was suppressed by vit.K2 in chondrocytes of both types of donor rats at both time points of investigation. No major difference in Socs1 gene expression was observed between control and T2DM groups without vit.K2. In regard to Socs1, Protein S and Gas69, as K vitamin-dependent ligands and parts of the TAM family (Tyro3, Axl, and Mer receptor tyrosine kinases) have been implicated in its regulation by vitamin K.55,56 In our experiment, stimulation with inflammatory mediators reflecting OA conditions played a minor role in the regulation of Socs gene activity. van de Loo et al. 57 showed an enhanced Socs3 expression in chondrocytes from human OA cartilage while Socs1 expression remained unaffected. de Andres et al. 58 reported a reduction of Socs2 expression, but no statistically significant changes in Socs1 and Socs3 in chondrocytes of human late OA cartilage (even though there was a trend toward a higher Socs3 expression).
Transcription of both collagen genes (type 2 and 1) was impaired by TNFα compared with the respective controls without TNFα, although this effect did not reach the significance level. This could be influenced by the vitamin C content of the chondrocyte growth medium which might interfere with the TNFα effect. This expected collagen suppression by TNFα is described by others. 59 T2DM is often associated with OA, 60 characterized by ECM remodeling toward lower collagen type 2 and higher collagen type 1 content. However, OA chondrocytes can also sometimes upregulate collagen type 2 61 and type 1, depending on the OA stage and cartilage zone. 62 The increase in collagen type 2 gene expression in T2DM without vit.K2 might suggest a deregulated transcriptional activity. Other authors suggested either an increased collagen degradation in T2DM without vit.K2 63 or post-transcriptional suppressive mechanisms. 64 This upregulation of collagen type 2 transcripts was not visible on the protein level and was absent in T2DM chondrocytes with vit.K2 supplementation at the gene level. However, calculating the ratio between collagen type 2 to type 1 gene expression still reflected this increase, in this case also for chondrocytes of vit.K2 supplemented diabetic rats.
Interestingly, feeding the living chondrocyte donor rats with vit.K2 clearly influenced chondrocyte responses in subsequent cell culture: anabolic effects such as a slight increase in metabolic activity, proliferation, wound closure, undirected motility and a modified gene expression profile with suppression of the activity of some stress response genes such as Hmox1, and cytokine feedback regulators Socs1 and Socs3 (with an exception for HG in control with vit.K2). The elevated serum levels of vit.K2 in these rats fed with vit.K2 were measured in another recent publication. 9 Particularly, the T2DM rats showed a high vit.K2 level. 9 The indirect approach of vit.K2 via feeding of donor animals might also explain the data variability in the groups with vit.K2 supplementation since the cell expansion for the experiments might led to different degrees of cellular vit.K2 loss. This observation that vit.K2 is stored in chondrocytes is highly surprising and raises the question of whether and how chondrocytes might take up vit.K2 and store it intracellularly explaining the maintenance of vit.K2 effects during several weeks of cell harvesting, freezing, and subsequent in vitro culture. This issue requires deeper analyses in the future.
The influence of the inflammatory mediators (TNFα and C5a) remained subordinate, for example, the induction of Hmox1 by TNFα in chondrocytes of rats which received no vit.K2 supplementation. Isolation of chondrocytes from adult and compromised diabetic rats is challenging due to the thin cartilage layer—most researchers use immature rats and hence, immature cartilage for chondrocyte isolation. For this reason, only limited cell numbers and not from all in the original animal experiment available donors could be harvested. The advantage of the rat model is that it allows to study untreated T2DM without any influence of therapeutic drugs.
Study Limitations
One limitation of this study is that ZDF rats suffer from a leptin receptor defect causing T2DM which might also influence metabolic responses of chondrocytes. Nevertheless, leptin resistance seems to play also a role in age-related obesity being associated with insulin resistance in humans. 65 The indirect approach by applying vit.K2 via feeding the chondrocyte donor animals might explain the high degree of standard deviation since food uptake, vit.K2 storage in tissues and chondrocytes, different chondrocyte culture times, and so on might influence cell response to vit.K2. As a limitation of this study, putative molecular interactions of anaphylatoxin C5a and TNFα with growth medium contents have to be considered, even under serum-reduced starving conditions. The vit.K2 feeding of the cell donor animals did not start directly after birth but high vit.K2 serum levels could be confirmed in the rats supplied with vit.K2. 9 Collagen was only assessed intracellularly by the immunostaining in a semiquantitative manner as a limitation of the experimental settings. Hence, only information about procollagen production is accessible and not of that collagen fraction released into the culture supernatant. The advantage of this methodological approach is to visualize simultaneously (pro)collagen type 1 and 2 syntheses in each cell. Moreover, our chondrocyte CM contains ascorbic acid which is known to stimulate collagen synthesis 66 an effect which might mask the influence of the treatment regime on collagen synthesis.
In vitro studies like our study are generally limited by their methodology and their transferability on in vivo conditions might be lower than expected due to systemic influencing factors.
Conclusion
This in vitro study with cultured chondrocytes showed that the combination of TNFα and C5a used to simulate in vitro inflammatory conditions of OA exerted indeed catabolic effects by significant induction of Tnfα gene activity under HG conditions in chondrocytes of control rats without vit.K2 supplementation and a significant reduction of collagen type 2 gene expression in those cells of control rats with vit.K2 supplementation under HG conditions. The response of articular chondrocytes derived from non-diabetic and diabetic donors differed: The chondrocytes of T2DM donors are irreversibly compromised as indicated by the significantly reduced wound healing capacity. The vit.K2 supply of the chondrocyte donor rats led to anabolic effects in these chondrocytes during in vitro culturing such as significantly enhanced proliferation in chondrocytes of non-diabetic rats with vit.K2 supplementation compared with those non-diabetic animals without vit.K2 feeding. The low expression of Hmox1 and Socs1,3 genes under the influence of vit.K2 might indicate that chondrocytes of non-diabetic and diabetic donors with a high intracellular vit.K2 status seem to be protected from an exhausting anti-oxidative and inflammatory response. This observation suggests that vit.K2 uptake could exert protective effects in T2DM-associated OA which needs further investigation in vivo.
Supplemental Material
sj-docx-1-car-10.1177_19476035251317091 – Supplemental material for Differential Responses of Articular Chondrocytes from Diabetic and Non-Diabetic Rats to Glucose Conditions and Inflammatory Stimuli: Influence of a Vitamin K2 Enriched Diet
Supplemental material, sj-docx-1-car-10.1177_19476035251317091 for Differential Responses of Articular Chondrocytes from Diabetic and Non-Diabetic Rats to Glucose Conditions and Inflammatory Stimuli: Influence of a Vitamin K2 Enriched Diet by Vivienne Rüdig, Tobias Braun, Nils Fleischmann, Alexander Reinhardt, Julius Wehrmann, Clemens Gögele, Maria Kokozidou, Christian Werner, Ingo Mrosewski and Gundula Schulze-Tanzil in CARTILAGE
Supplemental Material
sj-docx-2-car-10.1177_19476035251317091 – Supplemental material for Differential Responses of Articular Chondrocytes from Diabetic and Non-Diabetic Rats to Glucose Conditions and Inflammatory Stimuli: Influence of a Vitamin K2 Enriched Diet
Supplemental material, sj-docx-2-car-10.1177_19476035251317091 for Differential Responses of Articular Chondrocytes from Diabetic and Non-Diabetic Rats to Glucose Conditions and Inflammatory Stimuli: Influence of a Vitamin K2 Enriched Diet by Vivienne Rüdig, Tobias Braun, Nils Fleischmann, Alexander Reinhardt, Julius Wehrmann, Clemens Gögele, Maria Kokozidou, Christian Werner, Ingo Mrosewski and Gundula Schulze-Tanzil in CARTILAGE
Supplemental Material
sj-docx-3-car-10.1177_19476035251317091 – Supplemental material for Differential Responses of Articular Chondrocytes from Diabetic and Non-Diabetic Rats to Glucose Conditions and Inflammatory Stimuli: Influence of a Vitamin K2 Enriched Diet
Supplemental material, sj-docx-3-car-10.1177_19476035251317091 for Differential Responses of Articular Chondrocytes from Diabetic and Non-Diabetic Rats to Glucose Conditions and Inflammatory Stimuli: Influence of a Vitamin K2 Enriched Diet by Vivienne Rüdig, Tobias Braun, Nils Fleischmann, Alexander Reinhardt, Julius Wehrmann, Clemens Gögele, Maria Kokozidou, Christian Werner, Ingo Mrosewski and Gundula Schulze-Tanzil in CARTILAGE
Supplemental Material
sj-tif-4-car-10.1177_19476035251317091 – Supplemental material for Differential Responses of Articular Chondrocytes from Diabetic and Non-Diabetic Rats to Glucose Conditions and Inflammatory Stimuli: Influence of a Vitamin K2 Enriched Diet
Supplemental material, sj-tif-4-car-10.1177_19476035251317091 for Differential Responses of Articular Chondrocytes from Diabetic and Non-Diabetic Rats to Glucose Conditions and Inflammatory Stimuli: Influence of a Vitamin K2 Enriched Diet by Vivienne Rüdig, Tobias Braun, Nils Fleischmann, Alexander Reinhardt, Julius Wehrmann, Clemens Gögele, Maria Kokozidou, Christian Werner, Ingo Mrosewski and Gundula Schulze-Tanzil in CARTILAGE
Supplemental Material
sj-tif-5-car-10.1177_19476035251317091 – Supplemental material for Differential Responses of Articular Chondrocytes from Diabetic and Non-Diabetic Rats to Glucose Conditions and Inflammatory Stimuli: Influence of a Vitamin K2 Enriched Diet
Supplemental material, sj-tif-5-car-10.1177_19476035251317091 for Differential Responses of Articular Chondrocytes from Diabetic and Non-Diabetic Rats to Glucose Conditions and Inflammatory Stimuli: Influence of a Vitamin K2 Enriched Diet by Vivienne Rüdig, Tobias Braun, Nils Fleischmann, Alexander Reinhardt, Julius Wehrmann, Clemens Gögele, Maria Kokozidou, Christian Werner, Ingo Mrosewski and Gundula Schulze-Tanzil in CARTILAGE
Supplemental Material
sj-tif-6-car-10.1177_19476035251317091 – Supplemental material for Differential Responses of Articular Chondrocytes from Diabetic and Non-Diabetic Rats to Glucose Conditions and Inflammatory Stimuli: Influence of a Vitamin K2 Enriched Diet
Supplemental material, sj-tif-6-car-10.1177_19476035251317091 for Differential Responses of Articular Chondrocytes from Diabetic and Non-Diabetic Rats to Glucose Conditions and Inflammatory Stimuli: Influence of a Vitamin K2 Enriched Diet by Vivienne Rüdig, Tobias Braun, Nils Fleischmann, Alexander Reinhardt, Julius Wehrmann, Clemens Gögele, Maria Kokozidou, Christian Werner, Ingo Mrosewski and Gundula Schulze-Tanzil in CARTILAGE
Footnotes
Acknowledgment and Funding
The authors would like to thank Benjamin Kohl, Teresa Schotte, and Jingjian Shi for their support and acknowledge the funding by Stiftung Endoprothetik (costs for in vitro analyses), by OPED (for publication fees), PHIO scientific GmbH for support (time lapse microscopy), KappaBio (vit.K2 [MK-7]), and Kerscher’sche Stiftung (costs for animals).
Declaration of Conflicting Interests
The author(s) declared the following potential conflicts of interest with respect to the research, authorship, and/or publication of this article: Declaration of Generative AI and AI-assisted technologies in the writing process: Statement: During the final adaption of this work, the authors used ChatGPT 3.5 in order to improve readability and language. After using this tool, the authors reviewed and edited the content again and take full responsibility for the content of this manuscript.
Ethical Approval
This project was also approved by the institutional internal review board and local review board (Regierung Unterfranken, Bavaria, RUF 55.2.2-2532-2-729-17).
Data Availability
Study data can be provided on request.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.