
Abstract
Objective
Osteoarthritis (OA) is a degenerative joint disease characterized by cartilage degradation, causing severe pain and disability. Recent studies suggest that miR-450a-5p may regulate inflammatory pathways in OA. This study aimed to elucidate the role of miR-450a-5p in OA, providing a potential therapeutic target for the clinical treatment.
Methods
Cartilage tissues were collected from OA patients undergoing knee replacement surgery, and CHON-001 cells were treated with interleukin (IL)-1β to induce an OA model in vitro. Real-time quantitative polymerase chain reaction was used to detect the miR-450a-5p expression, and Western blot determined the lipopolysaccharide-induced tumor necrosis factor (TNF)-α factor (LITAF) expression. The targeting relationship between LITAF and miR-450a-5p was verified by dual-luciferase reporter assay. Cell proliferation and apoptosis were assessed using the Cell Counting Kit-8 assay and flow cytometry, respectively. Levels of IL-6, IL-10, and TNF-α were measured via enzyme-linked immunosorbent assay. In addition, Western blot was employed to detect the expressions of matrix metalloproteinase-3 (MMP-3), collagen III, and aggrecan in extracellular matrix (ECM).
Results
MiR-450a-5p expression was significantly down-regulated in OA tissues and IL-1β-induced CHON-001 cells (~60%), while LITAF expression was markedly increased (~1.8-fold). There was a negative correlation between miR-450a-5p and LITAF in OA tissues (r = −0.596, P < 0.01). MiR-450a-5p directly targeted and inhibited LITAF expression. Its overexpression promoted chondrocyte proliferation, reduced apoptosis and inflammatory cytokines, and mitigated ECM degradation.
Conclusions
MiR-450a-5p inhibited LITAF expression, thereby attenuating apoptosis, inflammation, and ECM degradation in chondrocytes. It may serve as a promising therapeutic target for OA.
Introduction
Osteoarthritis (OA) is the most common joint disease in the elderly, leading to chronic pain, stiffness, and disability. 1 As reported previously, OA affected 12% of the population over 25 years old and 50% of the population over 65 years old. 2 The main characteristics of OA include chronic inflammation, progressive destruction of articular cartilage, and subchondral sclerosis. 3 The development of OA has been demonstrated to be related to age, previous joint injury, obesity, genetics, gender, and joint shape. 4 However, there are still no effective disease-modifying therapies due to the limited understanding of OA pathogenesis. Currently, joint replacement is the main clinical treatment for advanced OA. 5
Chondrocytes, the only resident cells in joint cartilage, play a crucial role in maintaining the dynamic balance between anabolism and catabolism in ECM. Risk factors such as abnormal mechanical stress and pro-inflammatory cytokines (like interleukin [IL]-1β) have been proved to reduce chondrocytes and degrade the ECM in cartilage, as evidenced by increased levels of matrix metalloproteinases (MMPs) and decreased levels of collagen III and aggrecan. 6 Despite efforts to understand the pathological process of OA have increased, the molecular mechanisms remain elusive. Therefore, identifying novel drug targets to develop more effective treatment options is of high medical necessity.
Recently, microRNAs (miRNAs) have been recognized as one of the epigenetic mechanisms in gene expression regulation. 7 Innovative targets of OA, such as genes involved in OA development, have been uncovered through genetic networks, epigenetics, and miRNA-based approaches. 8 The miRNAs are a group of conserved small RNAs, consisting of 20 to 22 nucleotides, which regulate the post-transcriptional expression of mRNAs by binding to the 3’-untranslated region, leading to mRNA denaturation and inhibition of mRNA translation. 9 Several miRNAs have been identified to be associated with OA pathogenesis. For example, miR-27a promotes the autophagy and apoptosis in IL-1β-treated chondrocytes via the phosphoinositide 3-kinase/protein kinase B/mammalian target of rapamycin signaling pathway. 10 In addition, miR-29b-3p promotes chondrocyte apoptosis by targeting progranulin, thereby promoting OA development. 11 Another study demonstrated that miR-140-5p affects chondrocyte proliferation, apoptosis, and inflammation by targeting high mobility group box 1 in OA. 12
MiR-450a-5p regulates a variety of biological responses and has been implicated in the pathogenesis of various diseases. It is often abnormally expressed in malignancies, where it influences processes such as cell proliferation, apoptosis, autophagy, and angiogenesis.13,14 For instance, miR-450a-5p was shown to modulate autophagy by targeting the estimated glomerular filtration rate, enhancing the sensitivity of glioma cells to gefitinib in chemotherapy. 15 Low expression of miR-450a-5p has been reported to be associated with poor prognosis in colorectal cancer and other diseases, indicating its potential regulatory effect on cancer stem cell properties and angiogenesis. 16 Moreover, recent studies have highlighted its role in regulating inflammatory signaling pathways and cellular stress responses. 17 Given that chronic inflammation, matrix degradation, and cell death are central to the pathophysiology of OA, it is conceivable that miR-450a-5p may influence OA progression by modulating similar mechanisms in chondrocytes. However, its specific role in OA remains unclear. Therefore, this study aimed to elucidate the biological function and regulatory targets of miR-450a-5p in OA, thereby providing a new therapeutic target for clinical treatment.
Lipopolysaccharide (LPS)-induced tumor necrosis factor (TNF)-α factor (LITAF) is an important transcription factor that activates the transcription of inflammatory cytokines induced by LPS, such as TNF-α. 18 Briefly, LITAF interacts with the signal transducer and activator of transcription 6 (STAT6), forming a complex that translocates into the nucleus, where it binds to the promoter regions of cytokines and mediates the expression of inflammatory cytokines (such as TNF-α, IL-6, and IL-10). 19 A recent study has shown that LITAF, an endolysosomal protein, is not only active against bacterial toxins and Gasdermin D (a terminal effector of pyroptosis) but also mediates cell resistance to pore-forming toxins, leading to cell death. 20 Importantly, LITAF has been implicated in the regulation of inflammatory responses in various diseases such as arthritis. Dysregulation of LITAF expression can lead to an exacerbated inflammatory response, contributing to the degradation of cartilage and progression of OA.18,21
To investigate the potential role of miR-450a-5p and LITAF in OA, we collected cartilage tissues from OA patients and established an IL-1β-induced chondrocyte injury model in CHON-001 cells. The purpose of this study was to explore the regulatory roles of miR-450a-5p and LITAF in OA progression, providing a theoretical basis for the treatment of OA.
Methods
Clinical Sample Collection
Cartilage tissues, used for RNA extraction and subsequent experiments to analyze miR-450a-5p and LITAF expression, were collected from 2 distinct groups of participants at the Affiliated Wuxi Fifth Hospital of Jiangnan University between January 2020 and March 2021. The OA group consisted of 20 patients undergoing knee replacement surgery due to OA, diagnosed based on clinical symptoms and radiographic evidence. The normal group comprised 20 individuals without OA who underwent traumatic amputation surgery and had no history of joint diseases. All participants provided informed consent voluntarily, and the study procedures were approved by the research ethics committee of Affiliated Wuxi Fifth Hospital of Jiangnan University (Ethics Review No. 20191201). Demographic details, including age, gender distribution, and any relevant clinical characteristics, were recorded for both groups to ensure comparability and minimize potential confounders (Supplementary Table 1).
Cell Culture and Transfection
Human chondrocyte cell line CHON-001 (ATCC) was cultured in a complete Dulbecco’s Modified Eagle Medium (DMEM, Solarbio, Beijing) containing 10% fetal bovine serum (FBS, Gibco, USA) and 1% penicillin-streptomycin solution (Beyotime, Guangzhou) in an incubator at 37°C with 5% CO2. An in vitro OA cell model was constructed by treating CHON-001 cells for 12 hours with IL-1β (10 ng/ml, PeproTech, USA). 22
Cells at passages 3 to 5 were used for experiments. When the cell confluence reached 70% to 80%, negative control (NC) mimics, miR-450a-5p mimics, NC plasmid, and LITAF overexpression plasmid were transfected into the cells using Lipofectamine 2000 (Invitrogen, USA). After transfection, the cells were collected for subsequent experiments.
Real-Time Quantitative Polymerase Chain Reaction
Total RNA from cells or tissues was extracted using Trizol reagent (Thermo Fisher Scientific) and quantified with the NanoDrop 8000 spectrophotometer (Thermo Fisher Scientific, USA). Then, the extracted RNA was reverse transcribed into cDNA by PrimeScript RT Master Mix (Thermo Fisher Scientific) according to the manufacturer’s protocol. Real-Time Quantitative Polymerase Chain Reaction (RT-qPCR) was performed using the SYBR Green kit (Applied Biosystems, USA) on an ABI 7500 Real-Time PCR System (Applied Biosystems). Glyceraldehyde-3-phosphate dehydrogenase (GAPDH) was used as the internal reference. Each experiment was performed in 6 replicates. The procedures were shown as follows: pre-denaturation at 95°C for 10 minutes, followed by 40 cycles of denaturation at 95°C for 15 seconds, annealing at 60°C for 1 minute, and extension at 72°C for 30 seconds. The relative expression of the target genes was calculated using the 2−ΔΔ Ct method. 23 The primer sequences are shown in Table 1.
The Primer’s Information.
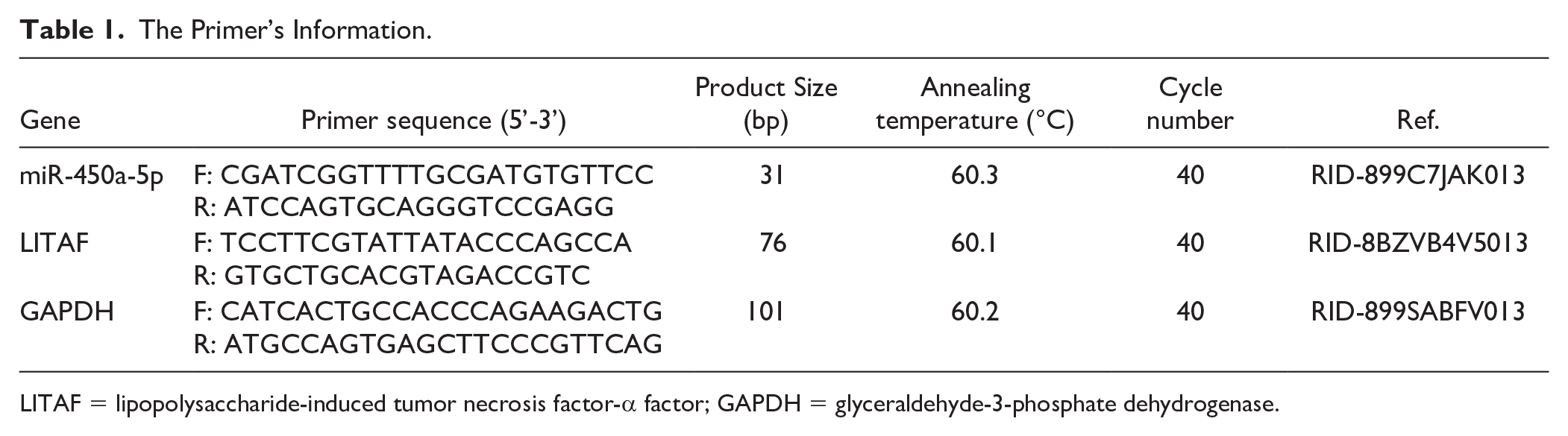
LITAF = lipopolysaccharide-induced tumor necrosis factor-α factor; GAPDH = glyceraldehyde-3-phosphate dehydrogenase.
CCK-8 Assay
Cell proliferation was determined using CCK-8 regent (Beyotime, #C0037) based on the manufacturer’s instructions. 24 Briefly, cells (2 × 104 cells/well) were seeded in a 96-well plate and subjected to the following treatments. The cells in the control group received no additional treatment. The cells in the IL-1β group were treated with IL-1β (10 ng/ml) for 12 hours. The cells in the IL-1β + NC mimics group were transfected with NC mimics and treated with IL-1β (10 ng/ml) for 12 hours. The cells in the IL-1β + miR-450a-5p group were transfected with miR-450a-5p mimics and treated with IL-1β for 12 hours. Subsequently, 10 µl of CCK-8 solution was added to each well on days 0, 1, 2, and 3 after the indicated treatment, and the cells were incubated at 37°C for 2 hours. The absorbance at 450 nm was measured using a microplate reader. All experiments were biologically repeated at least 3 times.
Flow Cytometry
The treated CHON-001 cells were harvested using trypsin and washed twice with pre-cooled sterile phosphate-buffered saline (PBS). The cell concentration was adjusted to 5 × 105 cells/ml. Then, 200 μl of cell suspension was incubated with 10 μl of Annexin V-FITC (Beyotime, #C1062M) and 10 μl of propidium iodide solution (20 mg/l) for 10 minutes at room temperature in the dark. Upon adding 500 μl of PBS, the cell apoptosis was determined via flow cytometry (BD Biosciences, LSRFortessa, Beijing). The apoptosis index was calculated as follows: Apoptosis index = number of apoptotic cells / (number of apoptotic cells + number of normal cells) × 100%. 25
Enzyme-Linked Immunosorbent Assay
The cultured cells were centrifuged for 10 minutes to collect the supernatants. Then, the levels of TNF-α (#ab181421), IL-6 (#ab178013), and IL-18 (#ab215539) were measured using corresponding enzyme-linked immunosorbent assay (ELISA) kits (Abcam, Cambridge, UK), following the manufacturer’s protocol. Finally, the optical density at 450 nm (OD450) was determined. 24
Western Blot Analysis
Cells were collected, and the total protein was extracted using cell lysis buffer. 24 The protein concentration was measured with the BCA Protein Assay Kit (Beyotime, #P0012). Equal amounts of protein (20 μg) were mixed with 5× sample loading buffer, boiled, and denatured. Upon separation by sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE), the proteins were transferred onto the polyvinylidene fluoride (PVDF) membranes (Millipore, USA). The membranes were blocked with 5% skimmed milk for 1 hour, incubated with primary antibodies overnight at 4°C. After washing 3 times, the membranes were incubated with the secondary antibodies at room temperature for 1 hour. Following washing, the membranes were supplemented with chemiluminescence reagent and visualized using a gel imaging system. Image J software was used to analyze the signals of protein bands, and GAPDH was used as an internal reference. The primary and secondary antibodies, all purchased from Abcam, were used as follows: MMP-3 (1:1000, #ab52915), collagen III (1:1000, #ab7778), aggrecan (1:1000, #ab3778), LITAF (1:1000, #ab187533), GAPDH (1:2500, #ab9485), and goat anti-rabbit IgG H&L (HRP) (1:5000, #ab6721).
Dual Luciferase-Reporter Assay
The interaction between miR-450a-5p and LITAF was assessed by luciferase analysis. 26 Specifically, the cells were transfected with dual-luciferase reporter vectors containing wild-type (WT) or mutant-type (MUT) binding sites of LITAF and miR-450a-5p, as well as miR-450a-5p mimics, using Lipofectamine 2000 (Invitrogen). After 24 hours of transfection, the luciferase signal was detected using the dual-luciferase reporter assay kit (Beyotime, #RG027).
Data Analysis
SPSS 26.0 was used for 1-way analysis of variance and independent samples t-test analysis, and GraphPad Prism 9.0 was employed for plotting. An independent t-test was used for comparison between 2 groups, and a 1-way analysis of variance for comparison among multiple groups. The results were represented as mean ± standard deviation, the Pearson correlation coefficient was used to analyze the expression correlation, and P < 0.05 was considered statistically significant.
Results
Reduced Expression of miR-450a-5p in Cartilage Tissues and Interleukin-1β-Induced CHON-001 Cells
The expression level of miR-450a-5p was significantly reduced in the cartilage tissues obtained from OA patients compared to normal controls (
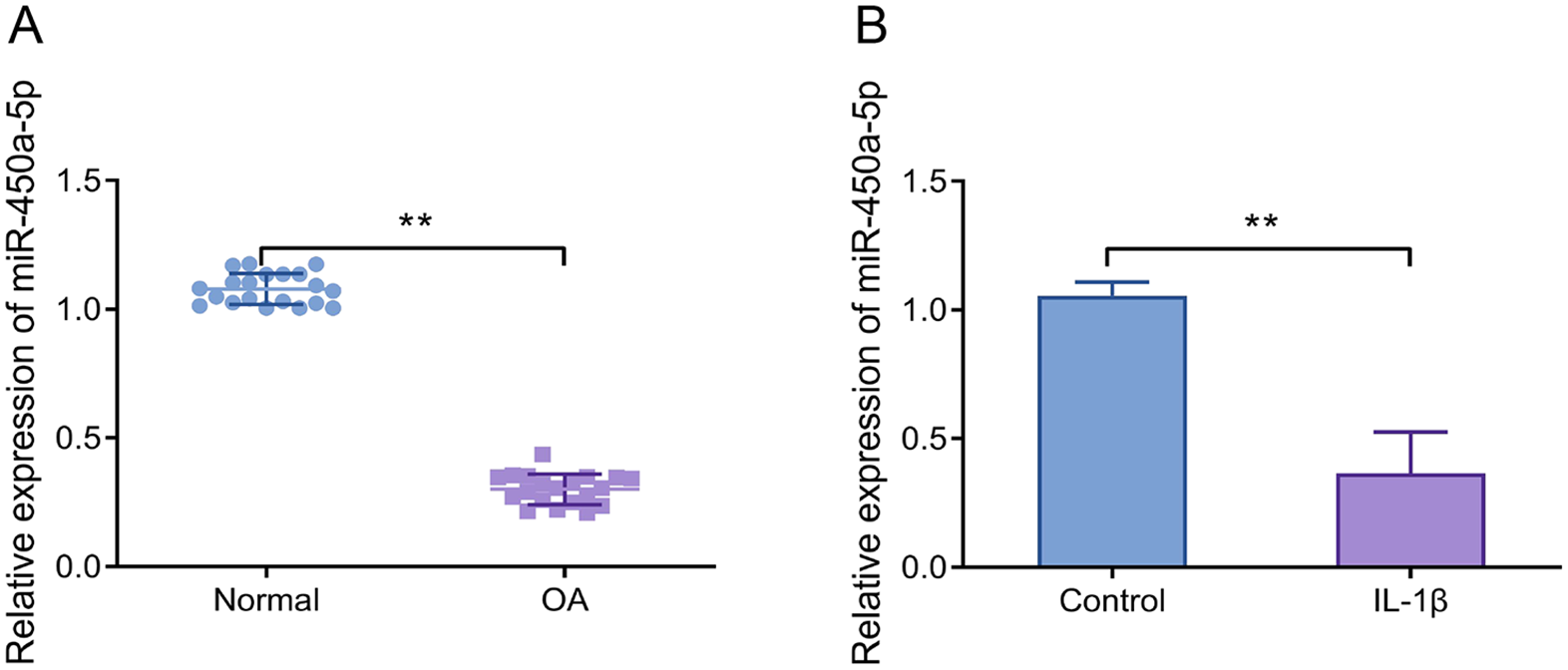
MiR-450a-5p expression is significantly elevated in cartilage tissues and IL-1β-induced chondrocytes. (
MiR-450a-5p Inhibits Apoptosis, Inflammation, and Extracellular Matrix Degradation in Interleukin-1β-Induced CHON-001 Cells
To verify the function of miR-450a-5p, CHON-001 cells were transfected with miR-450a-5p mimics. RT-qPCR results confirmed successful up-regulation of miR-450a-5p (
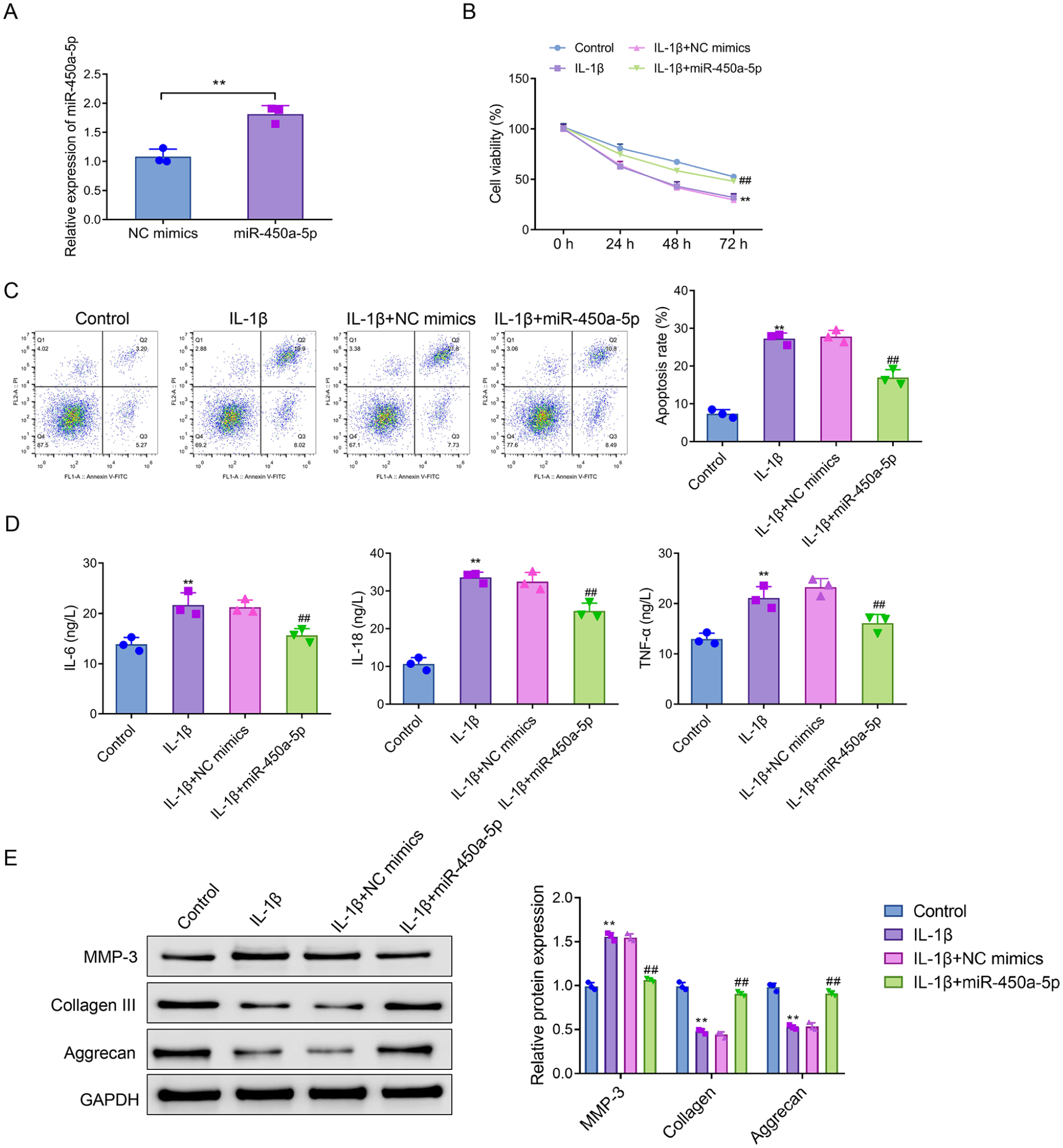
MiR-450a-5p inhibits IL-1β-induced apoptosis, inflammation, and extracellular matrix degradation in chondrocytes. (
In addition, up-regulation of miR-450a-5p level could moderate the release of cellular inflammatory factors. In contrast to the control group, the levels of IL-6, IL-18, and TNF-α were considerably raised in the IL-1β group. Overexpression of miR-450a-5p inhibited the increase of these cytokines induced by IL-1β (
Lipopolysaccharide-Induced Tumor Necrosis Factor-α Factor Acts as a Target Gene of miR-450a-5p
The interaction site between miR-450a-5p and LITAF was predicted by bioinformatics tools (
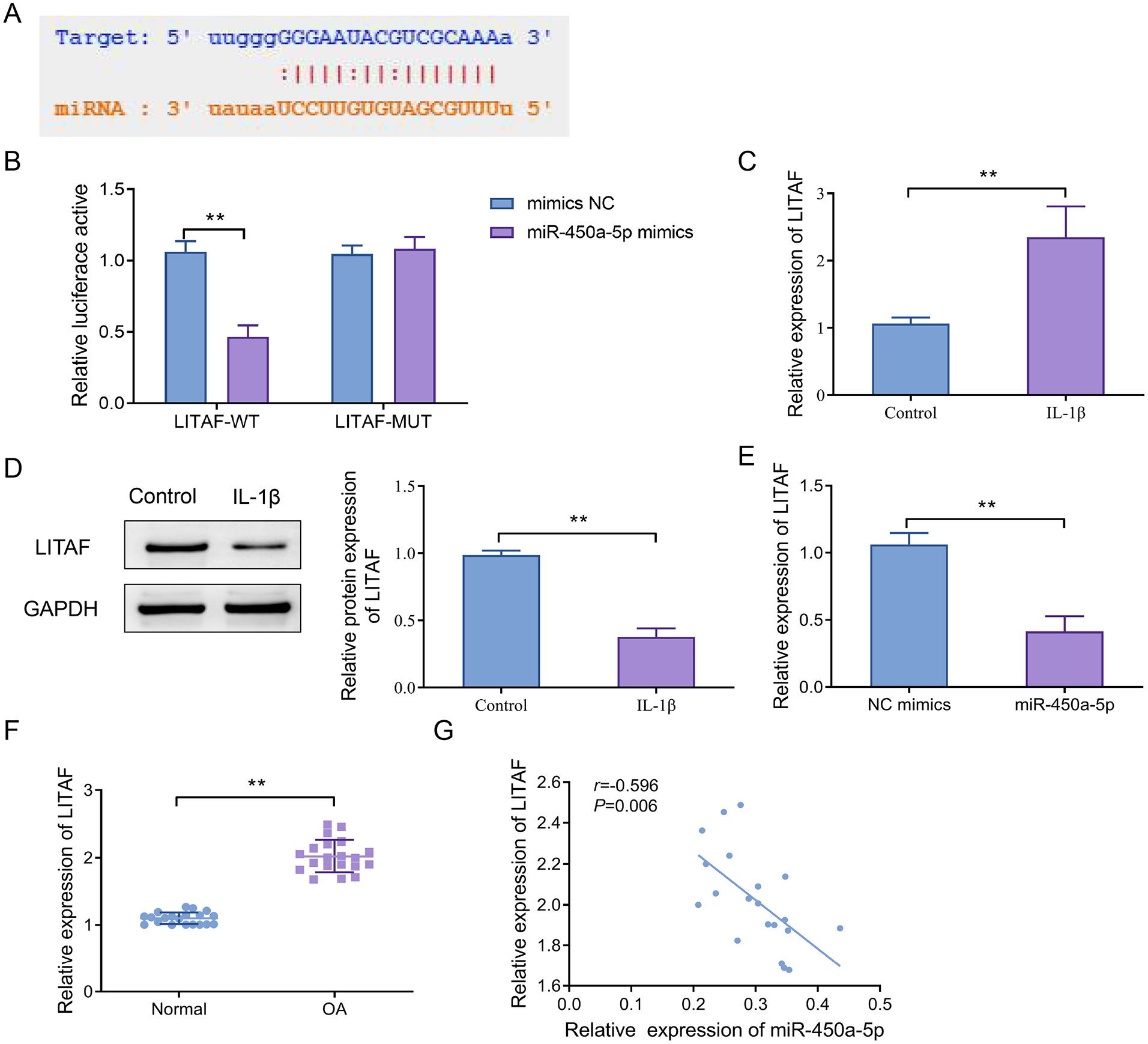
LITAF serves as a direct target of miR-450a-5p. (
Lipopolysaccharide-Induced Tumor Necrosis Factor-α Factor Reduces the Protective Effect of miR-450a-5p on Interleukin-1β-Induced CHON-001 cells
To determine whether miR-450a-5p regulated IL-1β-induced apoptosis, inflammation, and ECM degradation by inhibiting LITAF, we co-overexpressed miR-450a-5p and LITAF in CHON-001 cells. Western blot analysis showed that LITAF protein levels were significantly increased (~1.8-fold) in the miR-450a-5p + LITAF group compared to the miR-450a-5p + vector group (
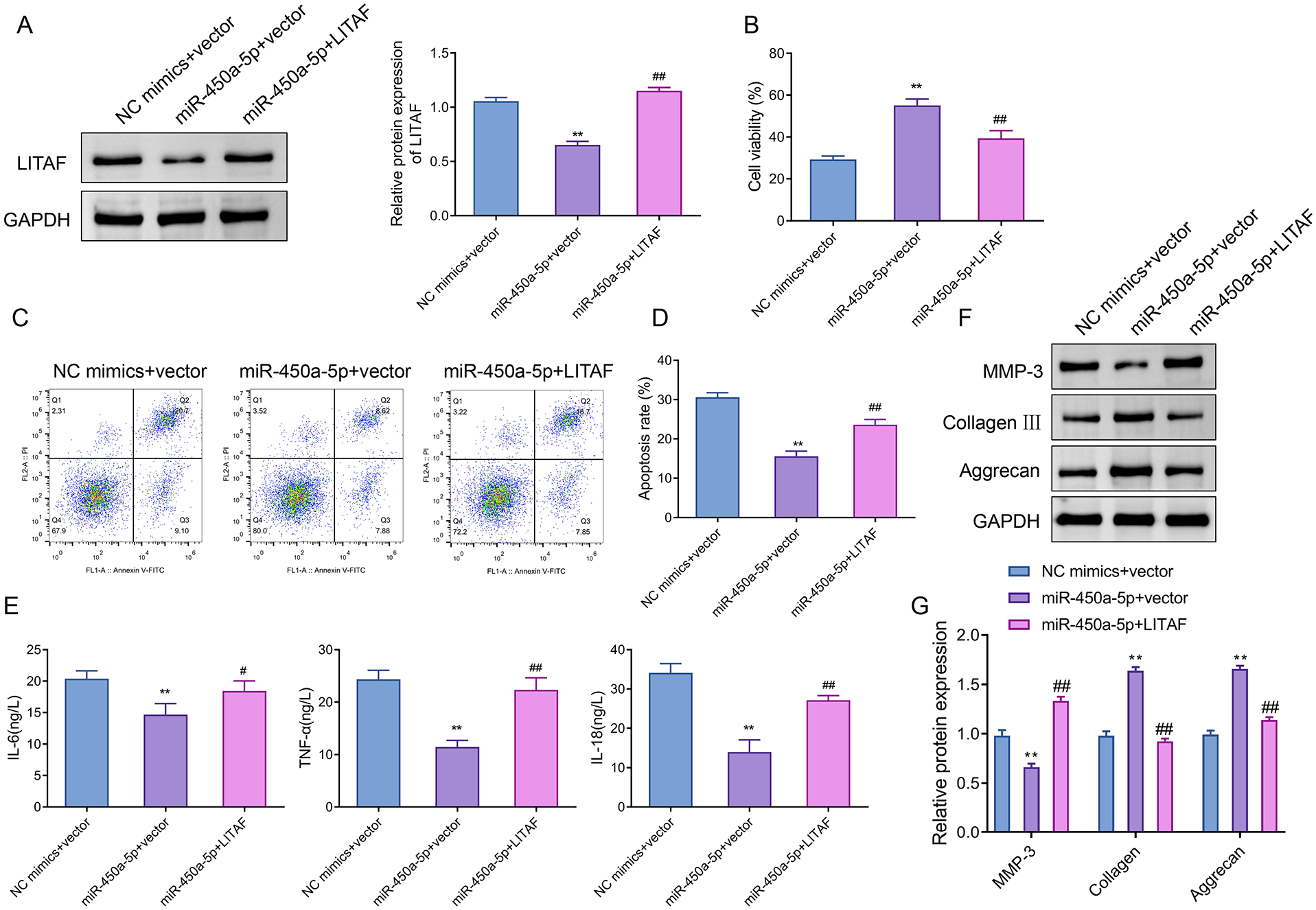
LITAF reduces the protective effect of miR-450a-5p on IL-1β-induced chondrocytes. (
ELISA results revealed that levels of IL-6, IL-18, and TNF-α were considerably decreased in the miR-450a-5p + vector group compared to the NC mimics + vector group, while these levels were significantly increased in the miR-450a-5p + LITAF group in comparison with the miR-450a-5p + vector group (
Overall, the above results collectively suggested that overexpression of LITAF reversed the ameliorative effects of miR-450a-5p on IL-1β-induced apoptosis, inflammation, and ECM degradation in CHON-001 cells.
Discussion
Abnormal regulation of miRNA level is closely associated with a variety of diseases, including OA. 27 Various miRNAs have been reported to regulate the development of OA. For example, microRNA-142-3p inhibits chondrocyte death and inflammation in OA by inhibiting the nuclear factor kappa B (NF-κB) pathway regulated by HMGB1, 28 and miR-210 also inhibits this pathway through targeting death receptor 6. 29 The findings underscore the critical role of miRNA-regulated signaling networks in OA. In this study, we found that miR-450a-5p expression was significantly reduced in OA cartilage and IL-1β-treated chondrocytes in contrast to the normal controls, suggesting its role in inhibiting OA progression. Given the multifaceted roles of miRNAs in cellular processes, we speculated that miR-450a-5p could interact with other miRNAs that target similar pathways or genes involved in OA. For instance, miR-140 is another miRNA known to be involved in cartilage homeostasis and OA. 30 It targets ADAMTS-5, a key enzyme responsible for cartilage degradation. It is conceivable that miR-450a-5p may work in conjunction with miR-140 to modulate cartilage matrix components and inflammation. Similarly, miR-34a, which targets the SIRT1 gene and is involved in apoptosis and senescence of chondrocytes, might interact with miR-450a-5p in regulating these processes. 31 Therefore, further research is required to elucidate the precise mechanisms and interactions between miR-450a-5p and other miRNAs.
IL-1β is known to play a key role in inducing cartilage degeneration. 32 It affects the ECM of articular cartilage, which mediates various biological responses for cartilage recovery and homeostasis. 33 Persistent ECM damage can impair joint function and accelerate OA development. 34 IL-1β modulates ECM composition by reducing levels of structural proteins like collagen III and aggrecan, 35 while promoting the production of MMPs such as MMP-1, MMP-3, and MMP-13, leading to cartilage degradation. 36 In our study, overexpression of miR-450a-5p in IL-1β-stimulated CHON-001 cells reversed these deleterious effects by enhancing cell proliferation, reducing apoptosis, and regulating cytokine secretion and ECM degradation markers such as IL-6, IL-18, TNF-α, MMP-3, collagen III, and aggrecan. This highlights the therapeutic potential of miR-450a-5p in OA. Modulation of miR-450a-5p expression, either via gene therapy or small molecule delivery, may alleviate inflammation and protect cartilage. Thus, future studies should focus on the role of miR-450a-5p in different OA models and investigate its interaction with other miRNAs and signaling cascades. However, despite its therapeutic potential, miRNA-based strategies face several translational challenges, including limited in vivo stability, off-target effects, and the difficulty of achieving sustained and targeted delivery to articular cartilage.
TNF-α, an important cytokine mediator of immunomodulation and inflammation, plays a pro-inflammatory and pro-apoptotic role in a variety of cell types. 37 LITAF, a transcription factor, enhances TNF-α gene expression and has been linked to the transcription of various inflammatory cytokines (such as TNF, monocyte chemoattractant protein 1 [MCP-1], and IL-10). 38 Dysregulation of LITAF expression contributes to chronic inflammation and has been implicated in cancers including pancreatic and gastric cancer. 39 For example, miR-1 has been shown to inhibit gastric cancer cell growth by targeting LITAF, 40 and miRNA-106 was reported to affect prostate cancer radioresistance via LITAF regulation. 41 In our study, we demonstrated that miR-450a-5p directly targets LITAF by binding its 3’UTR, thereby inhibiting its expression. This was validated by dual-luciferase reporter assay and supported by increased LITAF expression in OA tissues and IL-1β-treated chondrocytes. Moreover, simultaneous overexpression of miR-450a-5p and LITAF showed that LITAF could reverse the protective effects of miR-450a-5p. Mechanistically, LITAF has been shown to interact with STAT6, facilitating its nuclear translocation and transcriptional activation of inflammatory genes, including TNF-α. 42 In addition, LITAF may enhance NF-κB pathway activation, which is known to upregulate matrix-degrading enzymes such as MMP-3. 43 Therefore, knockdown of LITAF may downregulate both TNF-α and MMP-3 by attenuating these signaling cascades, thereby reducing inflammation and cartilage degradation in OA.
Our findings align with previous research highlighting the role of miRNAs in OA pathogenesis, such as miR-27 and miR-140, which modulate inflammatory pathways and cartilage degradation processes. 44 Like these miRNAs, miR-450a-5p appears to play a protective role against inflammation through its regulatory effects on LITAF. This discovery not only reinforces the importance of miR-450a-5p in OA but also distinguishes our findings by detailing the specific mechanisms through which miR-450a-5p acts. This could explain discrepancies with previous studies that may arise from different experimental conditions or focus on other miRNAs. Further validation of these results is necessary to consolidate miR-450a-5p’s role as a potential therapeutic target in OA.
However, we acknowledged several limitations in this study. First, our study was limited to a single human chondrocyte cell line and primarily relies on in vitro experiments. Therefore, in vivo animal experiments and more human chondrocyte cell lines should be taken into consideration in the future. Second, the sample size of this study was small, and future studies should include larger samples and validation in clinical samples. In addition, investigating the role of miR-450a-5p at different time points could provide a more comprehensive understanding.
In summary, our study provides an understanding of miRNA roles in OA, highlighting miR-450a-5p’s capacity to influence disease progression by targeting key molecular pathways. Future research should expand on the intricate network of miRNA interactions to fully elucidate their potential in OA treatment strategies.
Conclusion
To sum up, miR-450a-5p is significantly downregulated in OA tissues and has the ability to target and inhibit LITAF expression. This regulation helps suppress chondrocyte apoptosis, inflammation, and ECM degradation (
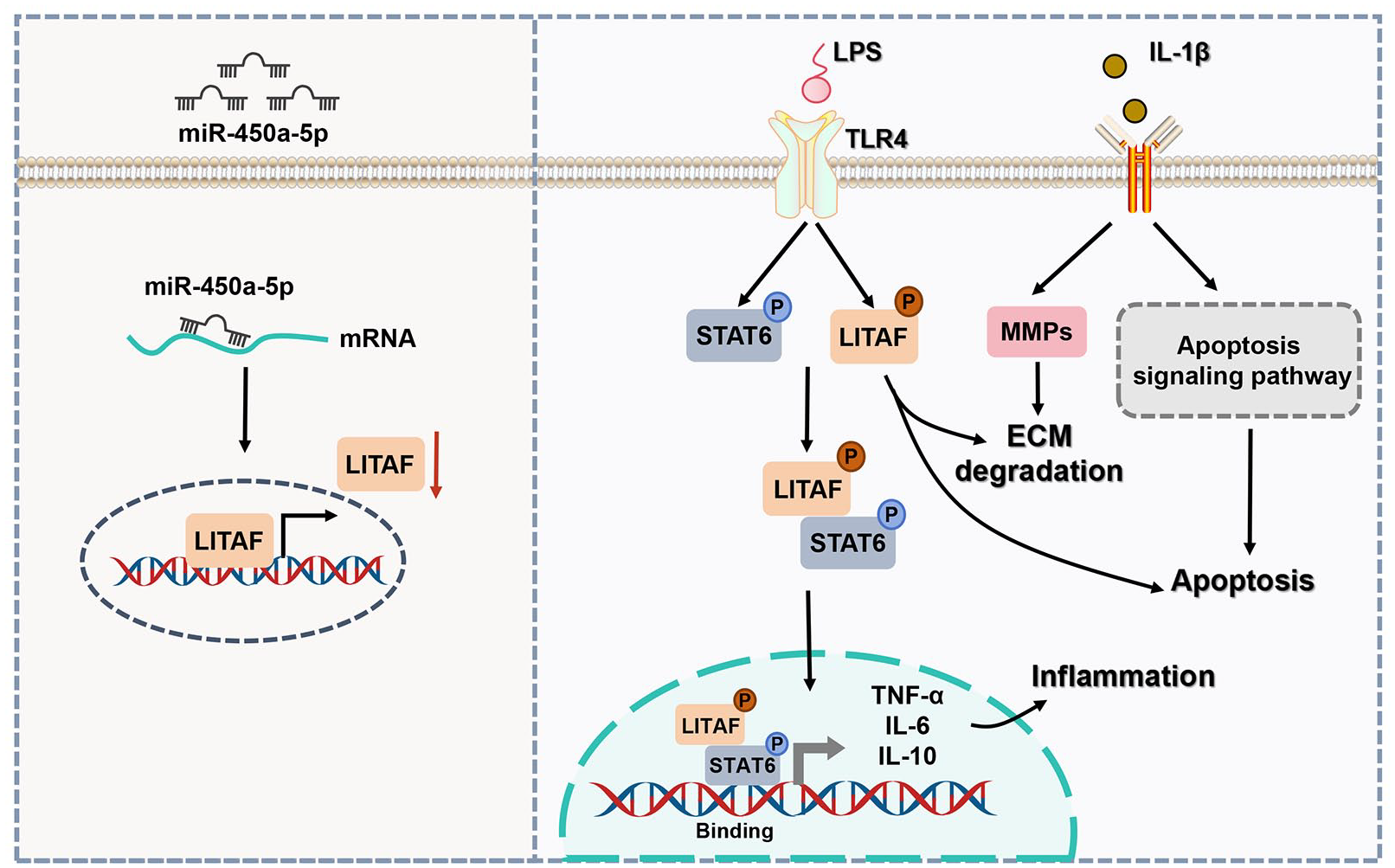
The diagram of potential mechanism by which miR-450a-5p targets and inhibits LITAF in OA.
Supplemental Material
sj-docx-1-car-10.1177_19476035251344478 – Supplemental material for Mir-450a-5p Ameliorates IL-1β-Induced Chondrocyte Apoptosis, Inflammation, and Extracellular Matrix Degradation by Down-Regulating LITAF
Supplemental material, sj-docx-1-car-10.1177_19476035251344478 for Mir-450a-5p Ameliorates IL-1β-Induced Chondrocyte Apoptosis, Inflammation, and Extracellular Matrix Degradation by Down-Regulating LITAF by Guo-feng Jia, Wei Tan and Xu Han in CARTILAGE
Footnotes
Acknowledgments and Funding
The authors would like to thank the patients for their support in publishing this article.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Author Contributions
G.-f.J., W.T., and X.H. contributed to concepts, design, data analysis, statistical analysis, manuscript preparation, data acquisition, and manuscript editing.
Ethics Approval and Consent to Participate
All participants were informed and voluntarily signed the informed consent. All study procedures were approved by the research ethics committee of Affiliated Wuxi Fifth Hospital of Jiangnan University.
Consent of Publication
Not applicable.
Availability of Data and Materials
The data sets used and/or analyzed during the current study are available from the corresponding author upon reasonable request.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.