
Abstract
Background
Osteoarthritis (OA) is a chronic disease that seriously affects human health. Although biomarkers are vital to the discovery and therapy of OA, current research on OA-specific biomarkers remains limited, indicating a need for further expansion of this field of study.
Methods
In this study, differential genes in OA patients and normal samples in Genomics Expression Omnibus (GEO) database were analyzed for signaling pathway enrichment. Then, Weighted Gene Co-expression Network Analysis (WGCNA) combined with Least Absolute Shrinkage and Selection Operator (LASSO) analysis was used to obtain key genes associated with OA diagnosis, including BCL6 co-repressor (BCOR), Coiled-Coil Domain Containing 59 (CCDC59), Jun Proto-Oncogene (JUN), Lysine Demethylase 3A (KDM3A), L3MBTL Histone Methyl-Lysine Binding Protein 4 (L3MBTL4) and Zinc Finger Protein 292 (ZNF292). Finally, the role of KDM3A in OA cell model was verified by constructing KDM3A overexpression and silencing cell lines.
Results
It was found that overexpression of KDM3A significantly downregulated β-catenin expression compared with the oe-NC group, thus affecting a series of biological processes in the OA cell model, specifically, increasing antioxidant capacity, reducing levels of inflammatory factors, and inhibiting extracellular matrix degradation.
Conclusion
This study not only provided six key target genes for OA but also revealed the important role of KDM3A in OA, providing a reference for gene targeted therapy for OA patients.
Background
Osteoarthritis (OA) represents a prevalent chronic degenerative condition, with its incidence progressively escalating in tandem with advancing age. 1 It predominately affects elderly populations, with a prevalence of 60% in individuals aged 50 and above, and increasing to 75% in those aged 80 and older. 2 The pathophysiological hallmarks of OA encompass cartilage degeneration, bone hyperplasia, synovial inflammation, and muscle atrophy. 3 Cartilage degeneration is one of the main characteristics of OA, which is manifested by the increase of chondrocytes, the degradation of matrix proteins and the thinning of cartilage layer, and eventually the complete disappearance or calcification of cartilage. 4 Currently, a variety of biomarkers, including those derived from serum, chondrocytes, and synovial cells, are being utilized for the early diagnosis of OA. 5 However, there are no effective biomarkers for prediction and treatment in OA.
The Wnt/β-catenin pathway plays a pivotal role in the initiation and progression of OA. At physiological levels, its activation stimulates chondrocyte proliferation and maintains their specialized characteristics, which in turn facilitates the repair and regeneration of articular cartilage. 6 However, excessive activation of this signaling pathway tends to trigger an increase in chondrodegradation-related enzymes, leading to accelerated cartilage matrix degradation. 7 In addition, the occurrence and progression of OA are accompanied by the release of inflammatory cytokines. 8 Research has proved an interplay between the Wnt/β-catenin pathway and inflammatory cytokines, notably interleukin-1β (IL-1β), where these cytokines have the capacity to either trigger or suppress the Wnt/β-catenin pathway, ultimately modulating the pathological mechanisms of OA. 9 Lysine Demethylase 3A (KDM3A) is a histone demethylase that affects the transcriptional activity of genes by removing methyl groups from histones, thereby regulating processes such as cell growth and differentiation. 10 Emerging evidence indicates KDM3A can promote pancreatic tumorigenesis by enhancing expression of genes pivotal to the cell cycle 11 ; moreover, it catalyzes hepatocarcinogenesis via the phosphatidylinositol 3-kinase (PI3K) pathway. 12 Given its significant involvement in these oncogenic processes, KDM3A has currently emerged as a promising therapeutic target for a wide range of diseases. 13 Relevant investigations have indicated a direct interaction between KDM3A and Wnt/β-catenin pathway, which significantly influences the oncogenesis of human colorectal cancer stem cells. 14 However, the role of KDM3A in osteoarthritis (OA) pathogenesis, particularly its interactions with β-catenin signaling, remains underexplored and warrants systematic investigation.
We conducted a differential gene expression analysis using data sets from the Gene Expression Omnibus (GEO) repositories, comparing data from OA patients to normal individuals. This analysis identified a panel of candidate genes differentially expressed in OA. Among these, KDM3A was selected for further investigation through biological experiments to explore its role in the OA cell model. This study provides a foundational rationale for investigating potential diagnostic markers and therapeutic targets in the context of OA.
Materials and Methods
Differential Expression Gene Screening
A total of 38, comprising 20 OA and 18 normal samples, were obtained from the GEO database (https://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE114007) for the analysis related to OA. We utilized the R package DESeq2 to identify DEGs, setting the screening criteria at P < 0.05 and |log2FC| > 1. The expression patterns of DEGs were graphically represented: volcano plots were crafted to depict the intensity and significance of gene expression changes, while heatmaps were employed to illustrate the clustering of DEGs. These visualizations were executed with the R packages “ggplot2” and “pheatmap,” respectively.
Functional Enrichment Analysis
We performed Gene Ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) analyses on the filtered DEGs using the “clusterProfiler” package in R language. Gene Set Enrichment Analysis (GSEA) is a computational method utilized to ascertain whether a predefined collection of genes exhibits statistically significant differences between two distinct biological conditions. Utilizing GSEA version 4.2.3, we conducted GSEA analysis on gene sets. The selection criteria were set at |NES| > 1, P < 0.05, FDR q-value < 0.25, to obtain the signaling pathway associated with OA-related genes.
Weighted Gene Co-Expression Network Analysis (WGCNA)
The “WGCNA” package was employed to cluster the samples, with parameters set as power = 7, minModuleSize = 80 and MEDissThres = 0.25. A weighted gene co-expression network was constructed by building Topological Overlap Matrix (TOM). Then, the dynamic tree cutting algorithm combined with hierarchical clustering technology was used to process TOM, and several gene modules with high internal correlation were successfully identified. Subsequently, gene modules were categorized on account of their correlation with specific traits. This classification was facilitated by examining the Module Eigengene (ME) for each module and calculating the Pearson correlation coefficient with the OA trait, along with its corresponding p-value. Consequently, gene modules with a strong association with OA were identified.
Establishment of Least Absolute Shrinkage and Selection Operator (LASSO) Regression Model
To affirm and predict relevant diagnostic biomarkers, we employed machine learning algorithms for OA prediction. The LASSO regression algorithm, deployed via the “glmnet” package in R, was employed for selecting genes that markedly differentiate OA patients from normal samples. Efficacy of these genes in distinguishing OA was assessed by the area under the receiver operating characteristic (ROC) curve (AUC value).
Cellular Material
The SW1353 cells (CL-0447, Wuhan Procell Biotechnology Co., Ltd.) were cultured in DMEM-H medium supplemented with 10% fetal bovine serum (FBS) and 1% penicillin/streptomycin (P/S). 15 The KDM3A gene was cloned into the pLVX-IRES-puro vector to obtain the oe-KDM3A recombinant plasmid. Subsequently, the oe-KDM3A and empty pLVX-IRES-puro plasmids were transfected into SW1353 cells to establish the oe-KDM3A and oe-NC (negative control) cell lines, respectively. In addition, si-NC and si-KDM3A plasmids (with the target sequences for knockdown listed in Table 1 ) were constructed and transfected into SW1353 cells to produce the si-NC and si-KDM3A cell lines. After 48 hours, the SW1353 cells were treated with IL-1β at a concentration of 10 ng/mL for 24 hours to induce an OA model.
Sequence of Interference Target Sites.

Quantitative Real-Time Polymerize Chain Reaction (PCR)
The RNA was extracted using isopropyl alcohol precipitation, 16 first-strand cDNA was synthesized using Goldenstar RT6 cDNA Synthesis Kit Ver.2 (TSK30, QingKe, Beijing). q-PCR was performed on a Bio-Rad IQ5 with 2x T5 Fast qPCR Mix (SYBR Green I) (TSE20, QingKe, Beijing). The primers employed for this detection are listed in Table 2 .
Primer Sequence.

Western Bolt
Take 500 μg of cell protein and mix it with 5x SDS loading buffer (8015011, Dakewei, Beijing, China) at 4:1. The mixture was then heated to 100°C for 6 min in a metal bath to denature the proteins, then 60 μg was taken for immunoblotting. The primary antibodies we used were anti-KDM3A (A2322, Abclonal, Wuhan, China), anti-CTNNB1 (bs-1165R, Bioss, Beijing, China), and anti-GAPDH (A19056, Abclonal, Wuhan, China). The secondary antibody used was HRP-conjugated Goat anti-Rabbit IgG (H+L) (AS014, Abclonal, Wuhan, China).
Antioxidant Activity Assay
The activities of total antioxidant capacity (TAC), superoxide dismutase (SOD), and catalase (CAT) enzyme activities were assayed using a TAC detection kit (KTB1500, Abbkine, Wuhan China), a total SOD detection kit (S0101S, Beyotime, Shanghai, China), and a CAT colorimetric assay kit (E-BC-K031-M, Elabscience, Wuhan, China), respectively.
Enzyme-Linked Immunosorbent Assay (ELISA)
The levels of MMP-1, NO, PGE2, IL-1β, and IL-6 inflammatory cytokines were detected using ELISA kits: specifically, Human Matrix Metalloproteinase-1 (MMP-1) (E-EL-H6073, Elabscience, Wuhan, China), Human Nitric Oxide (NO) (NLH7371, NewLife Biosciences, Wuhan, China), Prostaglandin E2 (PGE2) (E-EL-0034, Elabscience, Wuhan, China), Human Interleukin-1β (IL-1β) (E-EL-H0149, Elabscience, Wuhan, China), and Human Interleukin-6 (IL-6) (E-EL-H6156, Elabscience, Wuhan, China).
TUNEL Staining
Cell apoptosis was determined using TUNEL apoptosis detection kit (G1501, Servicebio, Wuhan, China). Nuclei were counterstained with DAPI staining solution (C1005, Beyotime, Shanghai, China). Specimens were then mounted with Anti-Fade Mounting Medium containing FITC (P0126, Beyotime, Shanghai, China) to preserve fluorescence.
Immunofluorescence Detection
Collagen II and Aggrecan protein expression were detected by immunofluorescence using the following reagents: Collagen II (A19308, ABclonal, Wuhan, China), Aggrecan Rabbit Polyclonal Antibody (A12045, ABclonal, Wuhan, China), FITC-labeled Goat Anti-Rabbit IgG (GB22303, Servicebio, Wuhan, China), Goat Serum (C0265, Beyotime, Shanghai, China), DAPI Staining Solution (C1005, Beyotime, Shanghai, China), and Anti-Fade Mounting Medium (P0126, Beyotime, Shanghai, China). After capturing images using a fluorescence microscope, the images were imported into ImageJ software. The threshold was adjusted, and appropriate regions of interest (ROI) and thresholding algorithms were selected. Within the designated ROI, the “Measure” function was utilized to quantify the fluorescence signal intensity, and the “Mean gray value” was chosen as the quantitative indicator for the average fluorescence intensity.
Statistical Analysis
All statistical analyses pertinent to bioinformatics were executed utilizing R software (version 4.1.0). Comparisons among continuous variables were stratified by group, with the t test applied to normally distributed data and the Mann-Whitney U test to nonnormally distributed data. LASSO regression analysis was conducted using the “glmnet” library. ROC curve analysis was utilized to assess the diagnostic accuracy of the included biomarkers. Statistical significance was set at P < 0.05.
Experimental data are presented as the mean ± standard deviation. Group comparisons were conducted using the t test, with statistical analysis and graphical representation performed by GraphPad Prism version 8. Differences between groups were considered statistically significant at P < 0.05.
Results
Differential Gene Screening and Pathway Enrichment
Using the GSE114007 data set, we identified 2,585 DEGs by comparing transcriptomic profiles of OA patients with those of healthy controls. Visual analysis through heatmaps and volcano plots revealed that a total of 1,319 genes were upregulated, whereas 1,266 genes exhibited downregulation (
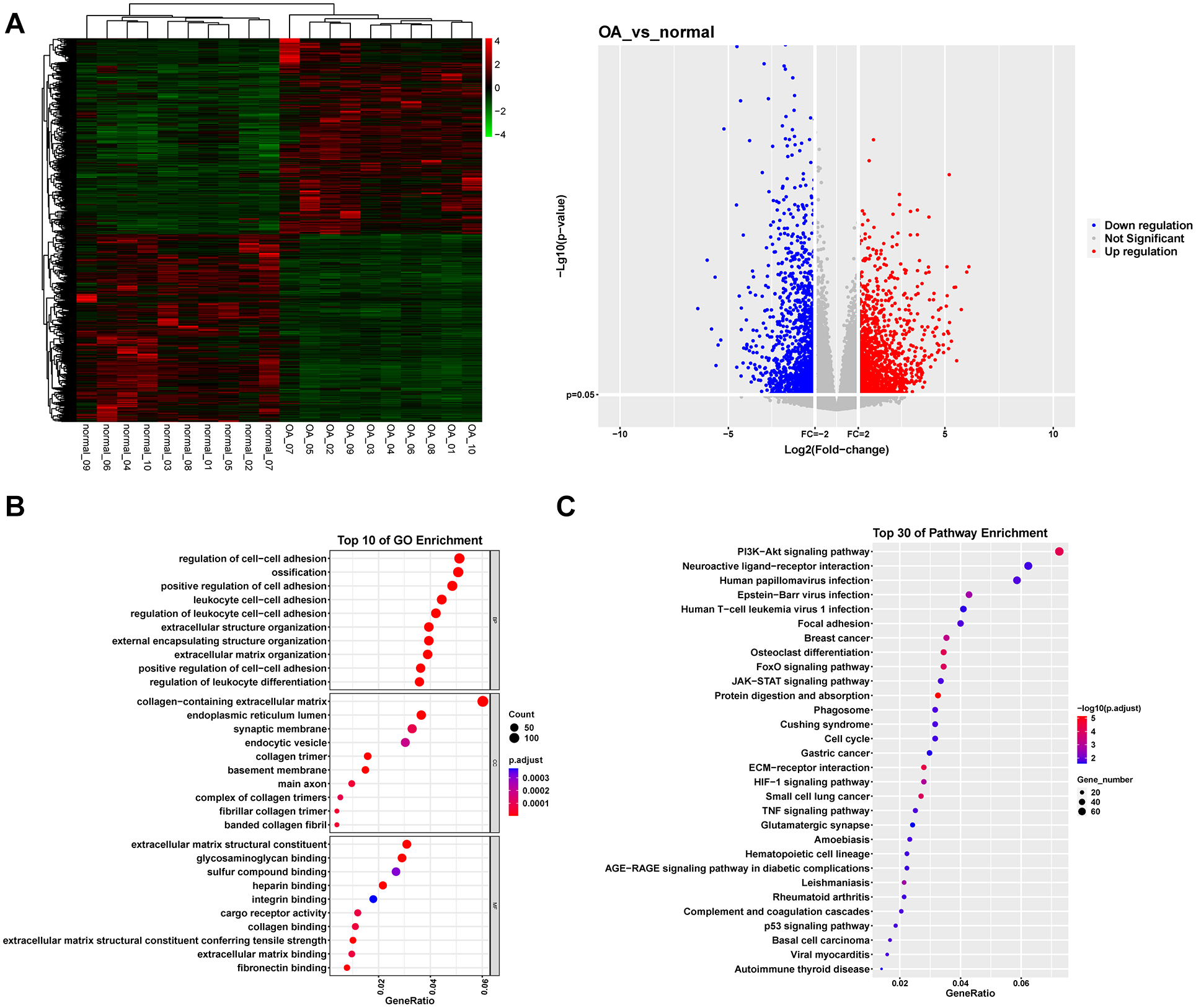
Differential gene screening and enrichment analysis between OA and normal individuals. (
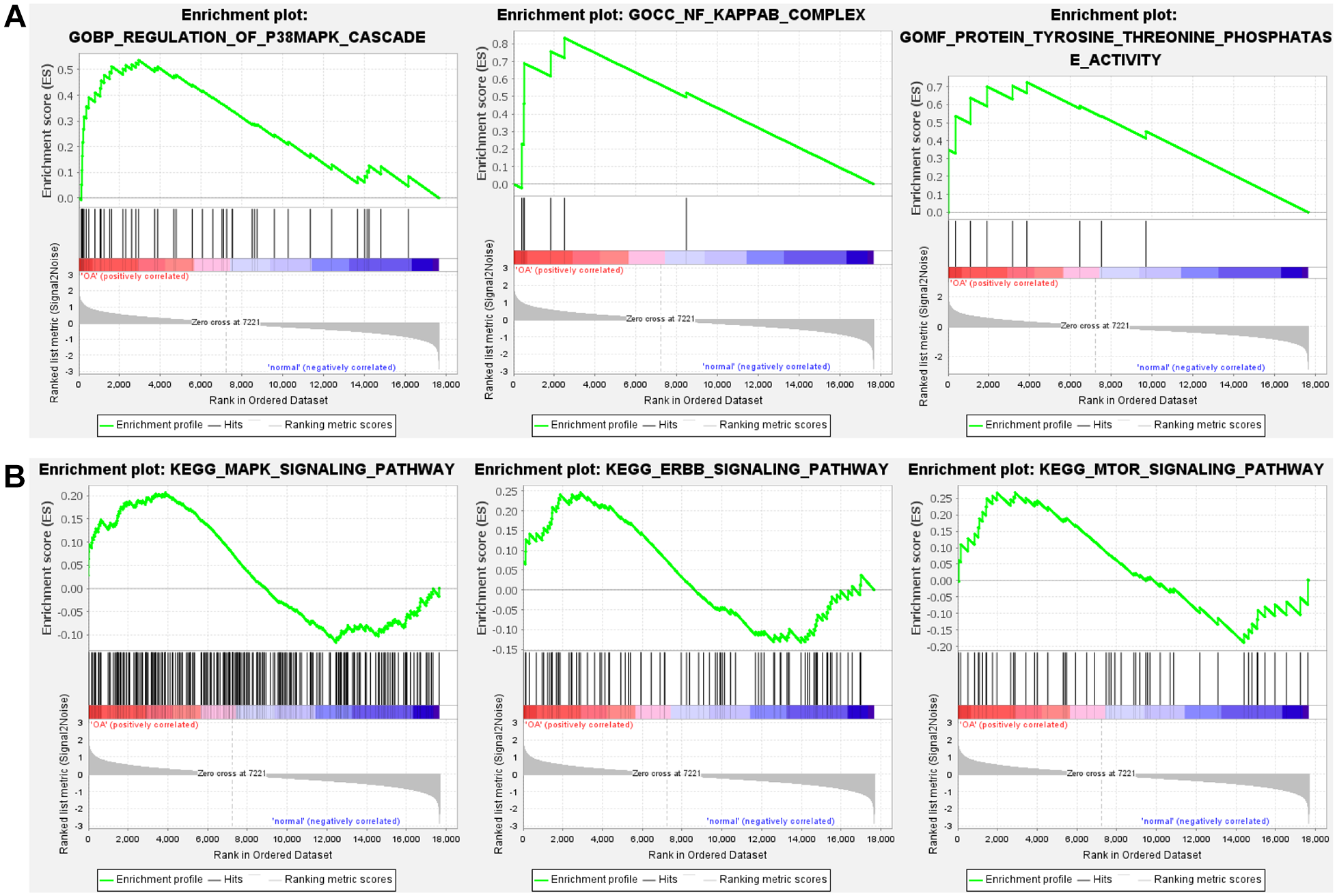
GSEA enrichment analysis. (
WGCN Analysis
Subsequently, the selected genes were clustered based on samples using R software. To ensure that the network followed to a scale-free distribution, we employed the pick Soft Threshold function from the WGCNA package for the computation of the soft thresholding power, and a soft threshold power of 7 was chosen for constructing the co-expression network (
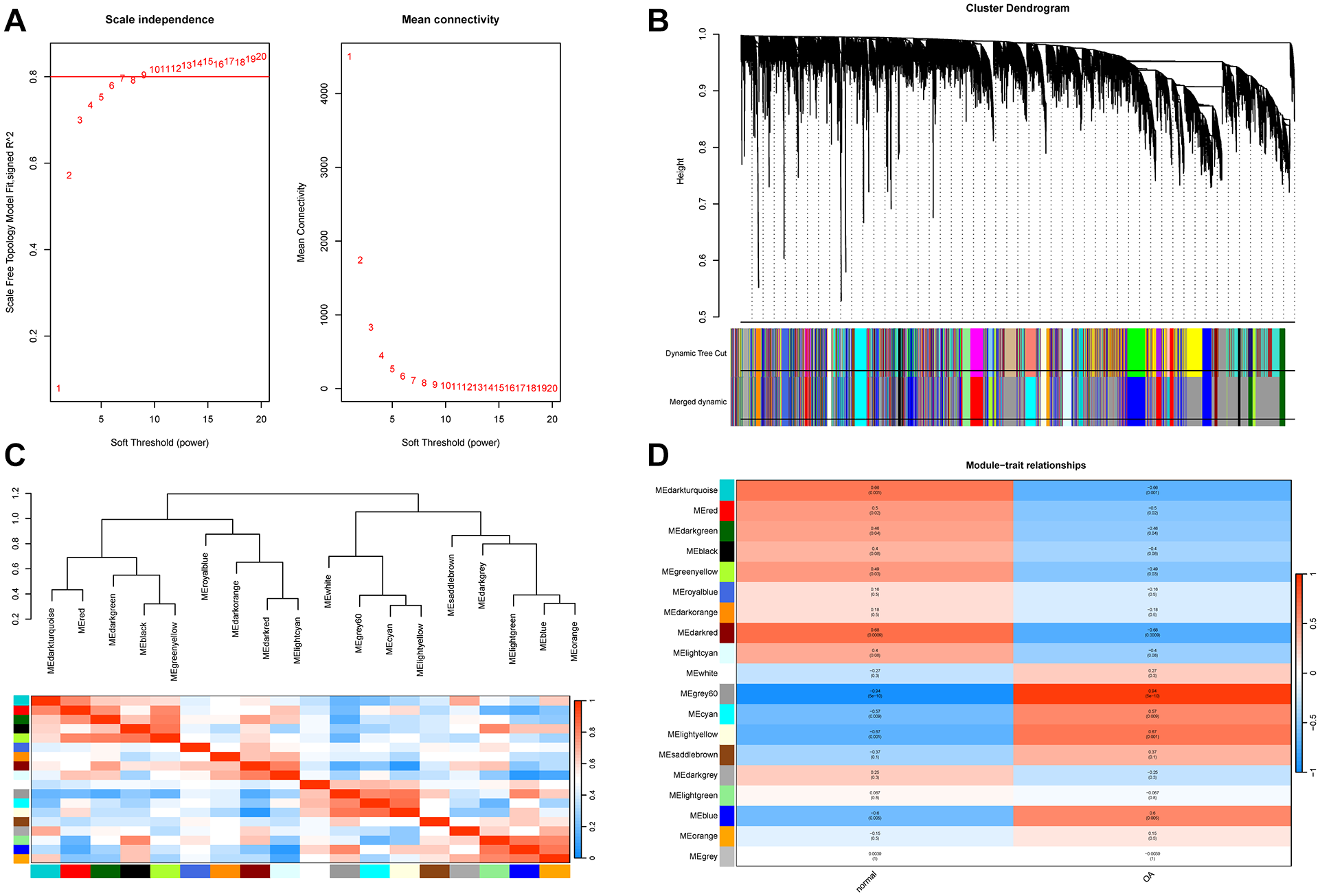
WGCNA analysis. (
Establishment of LASSO Regression Model
The genes from the MEgrey60 module identified by WGCNA were selected to establish a LASSO regression model. To ascertain the optimal regularization strength parameter λ, a 10-fold cross-validation strategy was implemented. A cross-validation curve was then generated, depicting the 10-fold cross-validation error associated with various values of λ. Ultimately, it was determined that the optimal regularization strength parameter λ was 6 (
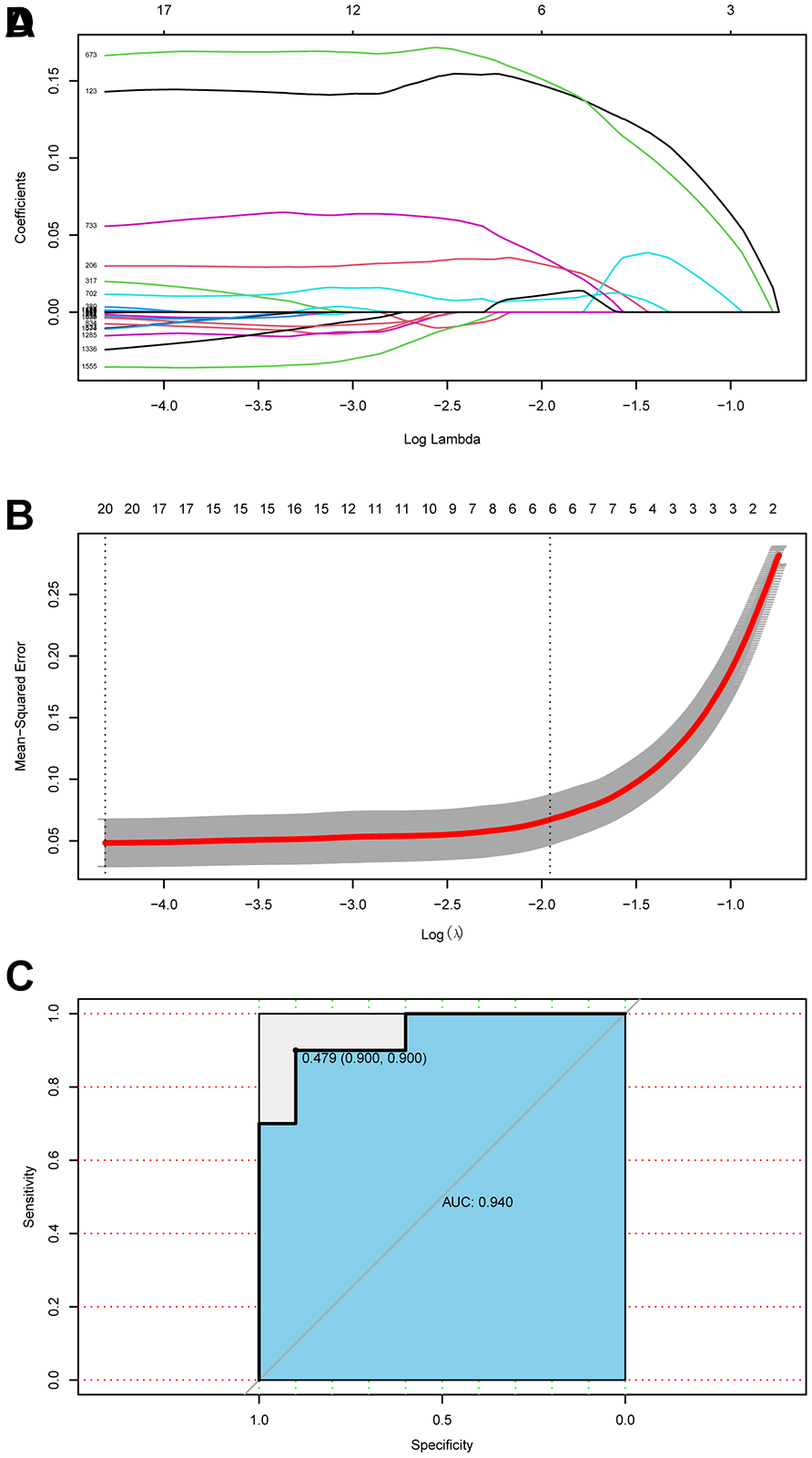
LASSO regression model. (
Among the six modeling genes, studies have indicated that JUN is highly associated with OA. 17 Moreover, KDM3A interacts with β-catenin, significantly affecting the development of cancer stem cells via the Wnt/β-catenin pathway. 14 Notably, the Wnt/β-catenin pathway plays a pivotal role in the initiation and progression of OA. When this pathway is suppressed, a mitigation of OA symptoms is observed. In contrast, its overactivation leads to the induction of chondrocyte aging.18,19 However, the intricate mechanism through which KDM3A directly regulates this pathway to influence OA remains elusive.
Overexpression of KDM3A Affects OA Cell Models by Down-Regulating β-Catenin
To further explore the mechanism of KDM3A affecting OA, we first overexpressed KDM3A in OA model cells. The results of q-PCR and Western blot (WB) indicated that, in the oe-KDM3A group, both the gene and protein expression levels of KDM3A were notably elevated relative to the oe-NC group, while the CTNNB1 (Code for β-catenin) level was notably decreased (

KDM3A overexpression impacts oxidative stress, inflammation, apoptosis, and ECM degradation via Wnt/β-catenin pathway. (
Silencing of KDM3A Affects OA Cell Models by Upregulating β-Catenin
To verify the reliability of the aforementioned results, we further transfected si-KDM3A and si-NC into OA model cells. Initially, q-PCR and WB assays revealed that in contrast to the si-NC group, the si-KDM3A group exhibited significantly decreased gene and protein expression levels of KDM3A, whereas gene and protein expression of CTNNB1 were substantially elevated (
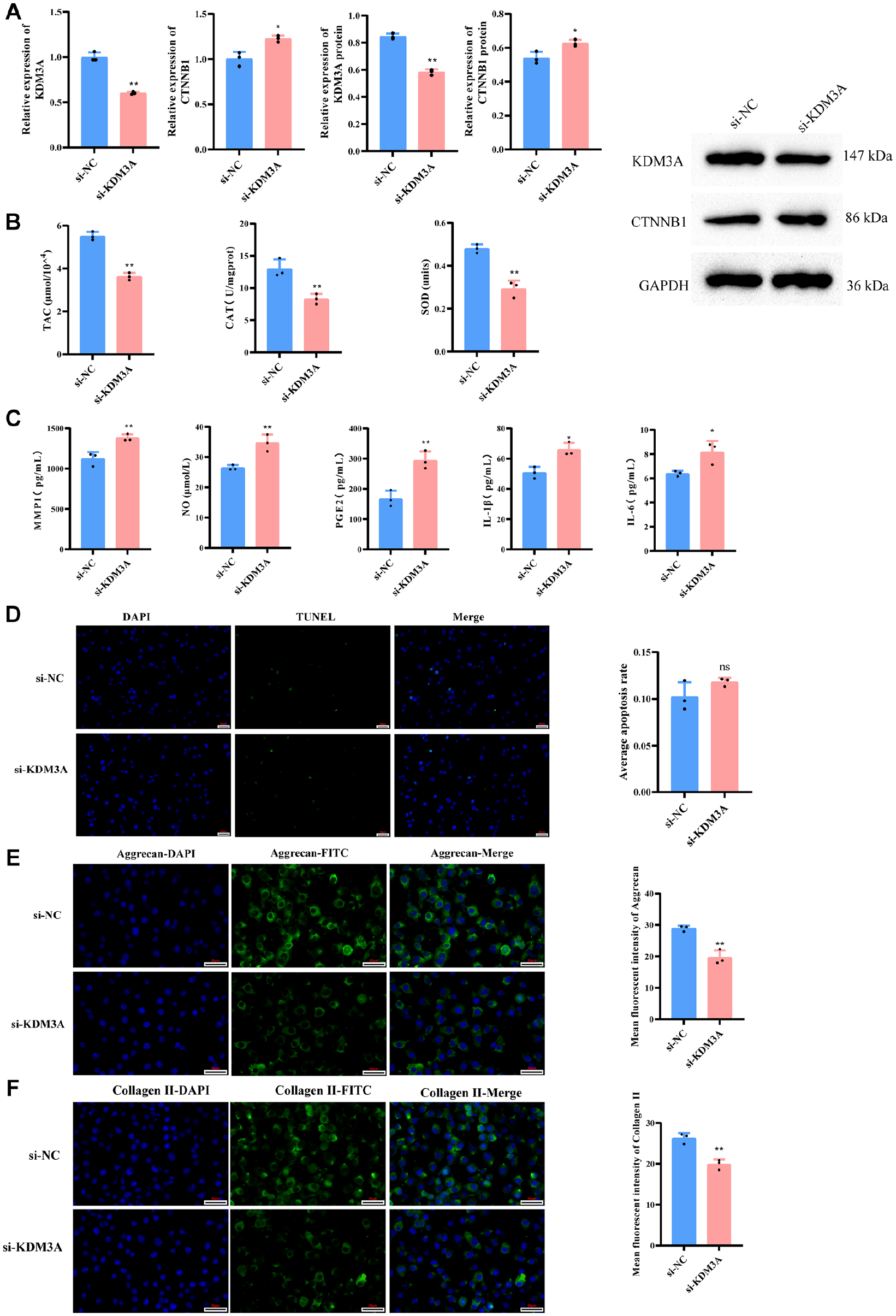
KDM3A silencing impacts oxidative stress, inflammation, apoptosis, and ECM degradation via Wnt/β-catenin pathway. (
Discussion
Osteoarthritis is a prevalent global condition, impacting approximately 300 million individuals worldwide as estimated. 20 Therefore, there is an urgent need to identify more reliable biomarkers for the early detection and effective treatment of OA. In our research, we employed a bioinformatics-driven screening approach and successfully pinpointed six candidate genes that hold potential for predicting OA. Among these, KDM3A was identified for its capacity to interact with β-catenin and its involvement in Wnt/β-catenin pathway. Our findings suggest that KDM3A may play a role in the progression of OA by modulating β-catenin levels in response to antioxidants, inflammatory responses, and extracellular matrix degradation.
The GO analysis indicated that differentially expressed genes (DGEs) are predominantly present in the pathways associated with ECM and collagen binding. Collagen constitutes a principal component of the ECM, serving to protect and repair damaged articular cartilage through its network-like structure. 21 Research has indicated that administration of collagen injections can mitigate OA symptoms. 22 Analysis utilizing the KEGG pathway database indicated that the majority of the DGEs were prominently associated with ECM-receptor interactions, which supports our analysis results from the GO pathways. Furthermore, GSEA analysis revealed that genes with a tendency to change were enriched in the GO pathways of related to regulation of p38MAPK cascade and NF-κB complex. Relevant research has demonstrated that suppression of the p38MAPK and NF-κB pathway markedly curbs human chondrocyte apoptosis and cartilage deterioration.23,24 Furthermore, GSEA analysis revealed that genes with a propensity to mutate were also notably enriched in the MAPK and ErbB signaling pathways, as categorized in the KEGG database. ErbB1, a member of the ErbB receptor family, has been implicated in the degeneration of articular cartilage. 25 It orchestrates the breakdown of ECM through activation of the MAPK pathway. 26 To identify valuable biomarkers, we used WGCNA and LASSO analyses. As a result, a total of six key genes were identified: BCOR, CCDC59, JUN, L3MBTL4, ZNF292, and KDM3A. BCOR functions as a transcriptional repressor and is integral to various biological processes, including embryonic development, cell differentiation, and tumorigenesis. 27 However, no studies have yet demonstrated the involvement of BCOR in the onset of OA. CCDC59 is a protein containing a coiled-coil domain. Overexpression of the CCDC59 protein can significantly protect mice from pulmonary inflammatory responses and improve lung function. 28 JUN is involved in the regulation of multiple gene expressions, exerting crucial influences in cellular stress responses, tumorigenesis, and immune responses. 29 Meanwhile, JUN serves as a biomarker and therapeutic target for OA. Research findings indicate that JUN is significantly downregulated in OA rats and aged chondrocytes. 30 L3MBTL4 is involved in chromatin remodeling and gene expression regulation. It may play a crucial role in cell differentiation and development, and has been identified as a key factor in the development of hypertension. 31 ZNF292 is a tumor suppressor gene, and studies have shown that ZNF292 is mutated in various cancers, suggesting its potential role in cell proliferation and differentiation. 32
Based on the above information and the role of KDM3A in the Wnt/β-catenin signaling pathway, we selected KDM3A as the focus of subsequent research. The expression level of CTNNB1 (Code for β-catenin) was remarkably decreased in oe-KDM3A and significantly increased in the si-KDM3A, suggesting that KDM3A affects the expression of β-catenin. Oxidative stress precipitates cartilage erosion and further exacerbates disintegration of the chondrocyte ECM. 33 Our study revealed that upon silencing the KDM3A gene, there was an upregulation of β-catenin expression accompanied by a significant decline in antioxidant capacity. Research has demonstrated that activation of the Wnt/β-catenin pathway elicits the production of reactive oxygen species (ROS). 34 Hence, the decreased antioxidant capacity in the si-KDM3A group may be due to enhanced β-catenin activity, potentially disrupting antioxidant signaling pathways or reducing antioxidant enzyme expression and activity. Inflammatory cytokines such as IL-1β and IL-6 are highly upregulated in OA patients, triggering the formation of ROS, which further exacerbates cartilage damage. 35 Moreover, the levels of MMP1 and NO are significantly enhanced in OA.36,37 Consistent with this, our findings indicated a significant reduction in the levels of MMP1, NO, PGE2, IL-1β, and IL-6 in the oe-KDM3A group when compared with the oe-NC group. Antioxidant enzymes neutralize radicals, mitigating oxidative damage and impeding the onset of inflammation. 38 As a result, an upsurge in KDM3A levels down-regulates β-catenin, thereby intensifying antioxidant enzyme efficacy, consequently resulting in a reduction in the levels of inflammatory factors in the OA cell model.
Apoptosis is a pivotal factor in OA pathogenesis, with aberrant chondrocyte apoptosis hastening the disease’s progression. 39 In addition, degradation of the ECM is a critical aspect in the pathological process of OA, involving various enzymes such as matrix metalloproteinases (MMPs). 40 It has been demonstrated that resveratrol inhibitors can suppress the expression of matrix metalloproteinases (MMP1, MMP3, MMP13) and β-catenin, thus curtailing chondrocyte apoptosis and ECM degradation. 41 However, in this study, TUNEL staining revealed no statistically significant differences among the oe-KDM3A, si-KDM3A, and control groups. We speculate that this might be because although KDM3A can improve the cellular survival environment and reduce oxidative stress and inflammatory responses, it does not necessarily directly influence the decision-making process of cell apoptosis. Alternatively, the impact of KDM3A on the Wnt/β-catenin pathway may not be sufficient to trigger significant changes in cell apoptosis. Furthermore, Collagen II and Aggrecan proteins are important proteins in articular cartilage, and their levels are closely related to ECM degradation. 42 Notably, immunofluorescence showed that overexpression of KDM3A led to an upregulation of Collagen II and Aggrecan. This finding suggests that KDM3A might delay OA by reducing β-catenin, thereby boosting Collagen II and Aggrecan expression in OA cells.
In conclusion, this research reveals that KDM3A interacts with the Wnt/β-catenin pathway through β-catenin, affecting essential biological processes in OA models, such as oxidative stress, inflammation, and extracellular matrix degradation. This finding not only deepens our comprehension of the regulatory mechanisms underlying OA but also offers a novel perspective for the screening and application of OA biomarkers. However, it is important to acknowledge that this study has certain limitations. Specifically, SW1353 cells were utilized in this research, and there are differences in their physiological characteristics and responsiveness to stimuli compared with primary human chondrocytes. In the future, we will consider expanding our investigations to include primary human chondrocytes, to more comprehensively assess the potential therapeutic value of the KDM3A-Wnt/β-catenin signaling pathway in OA.
Conclusion
In this study, we discovered that KDM3A exerts a significant influence in OA model cells by downregulating the expression of β-catenin. Moreover, KDM3A shows promise as a potential key biomarker for OA. Future investigations could further explore the specific mechanisms of action of KDM3A in OA, as well as its application potential in clinical diagnosis and treatment.
Footnotes
Acknowledgments and Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This study was supported by the Chongqing Traditional Chinese Medicine scientific research project (Joint project of Chongqing Health Commission and Science and Technology Bureau) (No.2024ZYZD006), the Science and Technology Research Program of Chongqing Municipal Education Commission (Grant No.KJQN202215120), and Natural Science Foundation of Chongqing, China (No.cstc2021jcyj-msxmX0853).
Author Contributions
Y.F., S.H.Y., Q.P., H.J., and J.Z. designed and wrote the original manuscript, performed the experiments and wrote the original manuscript, administered, and coordinated the whole study project. All authors have read and agreed to the published version of the manuscript.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Ethics Approval
Not applicable.
Consent for Publication
Not applicable.
Clinical Trial Number
Not applicable.
Availability of Data and Materials
The original contributions presented in the study are included in the article.