
Abstract
Objective
Damage to the joint-surface repair itself as fibrocartilage, a mixture of cartilaginous and fibrous tissues, which leads to debilitating conditions with persistent joint pain. The regulatory mechanisms that control the formation of these tissues are poorly understood.
Methods
We analyzed single-cell RNA-sequencing data from the repaired tissue formed in a chondral defect model of the monkey knee joint to identify genes specifically expressed in the repaired tissue. We used primary chondrocytes from semaphorin 7A (Sema7A) knockout mice to analyze the function of Sema7A in the dedifferentiation of chondrocytes in vitro. We introduced articular cartilage defect model in the knee joints in Sema7A knockout mice.
Results
Single-cell RNA-sequencing analysis revealed that Sema7A is specifically expressed in the cartilaginous tissue in the repaired tissue formed in the articular cartilage defect. In vitro analysis showed that Sema7A autonomously induced the dedifferentiation of chondrocytes at passage 2, which is assumed to correspond to cartilaginous tissue cells, toward a fibroblastic state. In addition, Sema7A heteronomously suppressed the re-differentiation of passage 8 fibroblastic cells, which were assumed to correspond to fibrous tissue cells. Addition of anti-integrin β1 neutralizing antibody abolished Sema7A-induced suppression of Col2a1 and Col9a1 expression, suggesting that integrin β1 mediates Sema7A function. Sema7A knockout mice showed significantly improved healing in an articular surface defect model of the knee joints. Sema7A deletion increased cartilaginous tissue formation and decreased fibrous tissue formation in joint-surface defects.
Conclusions
Sema7A regulates the balance between cartilaginous and fibrous tissues during articular cartilage damage repair.
Introduction
Articular cartilage is an avascular tissue with a limited ability to repair itself. Once damaged, it does not repair itself as healthy hyaline cartilage, but as fibrocartilage, a mixture of cartilaginous and fibrous tissue, 1 which can lead to cartilage degeneration and joint pain. Currently, no curative drugs are available for damaged articular cartilage. Treatments include bone marrow stimulation and autologous cultured chondrocyte implantation. However, in all of these treatments, the repaired cartilage still regenerates as a mixture of cartilaginous and fibrous tissue.2,3 This mixture of cartilaginous and fibrous tissue is functionally inferior to hyaline cartilage, and progressive degeneration of the tissue leads to secondary osteoarthritis. Understanding the molecular mechanisms that regulate the repair process of articular cartilage damage is necessary to develop therapies that heal articular damage in hyaline cartilage. However, the underlying molecular mechanism remains largely unknown, partly because of the absence of appropriate in vitro experimental models for fibrocartilage formation. Primary chondrocytes have been known to undergo dedifferentiation into fibroblastic cells when serially passaged in a monolayer expansion culture in a dish.4,5 Chondrocytes passaged multiple times lose the expression of chondrocyte markers, express fibrosis-related genes, 6 and have similar properties to chondrocytes in osteoarthritis. 7 Thus, in vitro dedifferentiated chondrocytes could be a candidate for modeling fibrocartilage in vivo.
Semaphorin is a general term for ligands that have a sema domain; they consist of a family of eight subclasses that are either secreted or anchored proteins. 8 Semaphorin was initially reported as a guiding factor for nerve axons. 9 It has since been reported to be involved in various physiological processes, such as angiogenesis,10,11 tumor invasion,12,13 the immune system,14 -16 and bone metabolism.17,18 Semaphorin 4D was shown to be involved in the inflammatory process in chondrocytes in a mouse model of rheumatoid arthritis. 19 Semaphorin 7A (Sema7A) is the only glycosylphosphatidylinositol (GPI)-anchored protein in the Semaphorin family. 20 Its primary functions include promoting the elongation of nerve axons 21 and acting as an immune regulator in lymphocytes and myelocytes. 22 These effects are mediated by the interaction between Sema7A and its two specific receptors, PlexinC123,24 and Integrinβ1.14,25 In addition, it has been reported that in rheumatoid arthritis, cells in synovium secrete Sema7A, which strongly stimulates monocytes, inducing the secretion of inflammatory cytokines, thereby exacerbating rheumatoid arthritis. 26 Sema7A is also involved in fibrosis in the lungs and liver.27,28 However, there have been no reports on the involvement of Sema7A in the physiology and pathology of articular cartilage.
To elucidate the molecular mechanisms responsible for the formation of a mixture of cartilaginous and fibrous tissues in the repair process after damage to articular cartilage, we searched for genes that were highly expressed in repaired tissue using single-cell RNA-sequencing (scRNA-seq) data that we previously reported. 29 We found enrichment of Sema7A expression in some cells of the repaired tissue formed from a knee articular cartilage defect in a macaque monkey model. Therefore, we hypothesized that Sema7A is involved in the formation of the mixture of cartilaginous and fibrous tissues. To test this hypothesis, we conducted experiments using a chondrocyte dedifferentiation model in vitro and Sema7A knockout mice to investigate the function of Semaphorin7a in the formation of a mixture of cartilaginous and fibrous tissues after articular cartilage damage.
Material and Methods
Ethics Statement
All methods were performed in accordance with the relevant guidelines and regulations. All animal experiments were approved by the Institutional Animal Care and Use Committee of Osaka University (approval no. 03-044-026). The experiments using recombinant DNA were approved by the Recombinant DNA Experiment Safety Committee of Osaka University (No. 04794).
scRNA-seq Analysis
We used scRNA-seq data (GSE206120) in which samples were collected from healthy hyaline articular cartilage (cyAC5) and from repaired tissue spontaneously formed 4 months after the creation of a chondral defect in articular cartilage (cyFC17w) in knee joints using a macaque monkey model. 29
Single cells were clustered using Seurat v4.0.348 in R 4.1.0. as previously described. 29 After filtering the cells, we normalized the expression data using the NormalizeData function (scale factor = 1,000,000, following the analytical parameters used by Muris49). We identified highly variable genes with the FindVariableFeatures function [selection.method = “vst,” nfeatures = 5,000, mean.cutoff = c(0.1, Inf), and dispersion.cutoff = c(0.5, Inf)]. After scaling the data, principal component analysis was performed, and the top 42 principal components were selected for dimensional reduction using UMAP (uniform manifold approximation and projection). The cluster resolution of the values was then determined, and the DEGs (differentially expressed genes) of each cluster were obtained using the FindMarkers function between the two groups (i.e., cell clusters). Gene set enrichment analysis (GSEA) was performed using fgsea package (v1.19.0) and C8: cell type signature gene sets in molecular signatures database (MSigDB).
Repaired Tissue in the Chondral Defects in the Knee Joints of Cynomolgus Monkey
Samples obtained in a previously reported study 29 were used. Briefly, cynomolgus monkeys (3-4 years old) were purchased from Ina Research (Nagano, Japan). The skin and joint capsules of the right knee were opened under general anesthesia. Chondral defects (1 mm diameter and 0.5 mm depth) were created at the trochlea of the distal femur. The joint capsule and the skin were closed. The monkeys were euthanized after 17 weeks. Knee joints were harvested and subjected to immunohistochemical analysis for Sema7A.
Sema7a Knockout Mice
Sema7a-/- mice on the C57BL/6 background were prepared and genotyped as described previously. 21
Primary Chondrocyte Culture
Primary chondrocytes were isolated from the epiphyseal cartilage of Sema7a-/- mice 5 days after birth, as described previously. 30 The isolated chondrocytes were stored in liquid nitrogen until further use.
Frozen stored chondrocytes were thawed, seeded at 1 × 106 cells per 10-cm dish (cat #3020-100, Iwaki, Tokyo, Japan), and cultured in Dulbecco’s modified Eagle’s medium supplemented with 10% fetal bovine serum and 1% penicillin-streptomycin at 5% CO2 in humidified air (passage 0, P0). When the culture reached sub-confluency, the cells were trypsinized and re-plated at 1 × 105 cells per 6-cm dish (cat #3961-035, Iwaki) (P1). RNA samples were collected from cells at passages 0, 2, 4, 6, and 8, when the culture reached sub-confluency, and were subjected to Sema7a mRNA expression analysis.
The co-culture assay was performed using a cell culture insert (cat #353103; Falcone). Wild-type primary chondrocytes at P8 were seeded at a density of 5 × 104 cells/well in a 12-well plate. A cell culture insert was placed on top of each well, and wild-type primary chondrocytes at P2 or Sema7a-/- primary chondrocytes at P2 were seeded at 5×104 cells/insert. RNA samples were collected from the cells at the bottom of the 12-well plate after 48 hours of co-culture.
Reverse Transcription Quantitative Polymerase Chain Reaction
Total RNA was extracted using an RNeasy kit (Qiagen, Hilden, Germany). For quantitative reverse transcription polymerase chain reaction (PCR), total RNA was reverse-transcribed into first-strand cDNA using ReverTra Ace (Toyobo, Osaka, Japan) and oligo(dT)20 primers. PCR amplification was conducted using the KAPA SYBR FAST qPCR Master Mix ABI Prism kit (KAPA Biosystems, Wilmington, DE) and the StepOnePlus Real-Time PCR System (Thermo Fisher Scientific, Waltham, MA). Sequences of the PCR primers used are listed in
Neutralization Assay of Integrin β1
Anti-integrin β1 neutralizing antibody solution 31 (4.3 mg/ml, cat #BE0232, BioXCell, Lebanon, NH) was purchased. Wild-type primary chondrocytes at P2 or Sema7a-/- chondrocytes at P2 were seeded at 5 × 104 cells/well in a 12-well plate. Anti-integrin β1 antibody solution was added to the culture medium at a final concentration of 10 μg/ml, and total RNA was collected 48 hours later.
Plasmids and Transfection Assay
The mouse Sema7A expression vector, Sema7a-FC-His, was obtained from Addgene (#72175; Watertown, MA). An Egfp expression vector under the control of the CMV promoter (pCMVb-gw/Egfp) was used as a negative control. Wild-type primary chondrocytes at passage 2 were seeded into six-well tissue culture plates (Corning, Corning, NY) at a density of 1 × 105 cells/well. Cells at 50% confluence were used for plasmid transfection assays in accordance with the manufacturer’s protocol using the Lipofectamine 2000 reagent (Invitrogen, Waltham, MA). The medium was replaced 6 h after transfection, and total RNA or protein was collected 48 hours later.
Western Blot Analysis
The cells were lysed using RIPA lysis buffer (Thermo Fisher Scientific) to collect protein samples. Samples were then added to the sample buffer (Invitrogen) and reducing agent (Invitrogen) and heated at 95°C for 10 minutes. The samples were applied to a 4-12% gel (Invitrogen) to separate the proteins, which were then transferred to a nitrocellulose membrane (Invitrogen). The membrane was incubated overnight with anti-β-actin antibody (#4967, Cell Signaling Technology, Danvers, MA) and Sema7A antibody (ab16066, Abcam, Cambridge, UK), followed by incubation with mouse anti-rabbit IgG-HRP (sc-2357, Santa Cruz Biotechnology, Dallas, TX) or m-IgG2a BP-HRP (sc-542731, Santa Cruz Biotechnology). The immunoreactive bands were visualized using SuperSignal™ West Dura Extended Duration Substrate (Thermo Fisher Scientific) and Fusion SOLO.7 S.EDGE (Vilber-Lourmat, Marne La Vallee, France), and quantified using Evolution Capt (Vilber-Lourmat).
Introduction of Articular Cartilage Repair Model into the Sema7a−/− Mouse Knee Joints
The number of animals to be used was estimated from the previously reported experiment
32
and our pilot study. Six male Sema7a−/− mice and Six male Sema7a+/− mice at 8-week old were anesthetized using dexmedetomidine hydrochloride (0.75 mg/kg), midazolam (4 mg/kg), and butorphanol tartrate (5 mg/kg). To control bleeding, a local injection of 1% xylocaine was administered before making an incision in the joint capsule. The medial parapatellar approach was used to expose the femoral patellar groove by dislocating the patella. Articular cartilage defects were created in the patellar groove using a surgical scalpel (Feather Disposable Scalpel #10; Feather, Osaka, Japan), following a previously described method
32
with modifications (
The samples were fixed with 4% paraformaldehyde and decalcified using ethylenediaminetetraacetic acid. They were then embedded in paraffin, and the femoral axis was placed perpendicular to the sectioning plane. The histological sections were obtained at 200 μm proximal to the intercondylar notch (
Introduction of Osteoarthritis Model Into Sema7a−/− Mouse Knee Joints
The right knee joints of the Sema7a+/− and Sema7a−/− mice were subjected to destabilized medial meniscus surgery 34 at 8-week old. The mice were sacrificed 4 weeks after the operation, and the knee joints were subjected to histological analysis.
Sagittal sections were prepared and stained with Safranin O-fast green iron hematoxylin. The cartilage destruction was scored using the OARSI (Osteoarthritis Research Society International) scoring system. 35 The OARSI scoring was performed by two independent scorers in a blinded manner. The femur and tibia scores were summed and designated as the OARSI score for each knee.
Histological Analysis
Semi-serial sections were immunostained with a DAB kit (cat #K1497, Dako). The primary antibodies used were anti-Sema7A (cat #LS-C406765-20, LSBio, 1:100), anti-type I collagen (cat #1310-01, SouthernBiotech, 1:1,000), and anti-collagen type2 (cat #MS-235-P0, Thermo scientific, 1:1,000), and the secondary antibodies used were rabbit polyclonal antibodies (cat #K4003; Dako, Glostrup, Denmark), goat polyclonal antibodies (cat #6420-05; SouthernBiotech, Birmingham, AL), mouse polyclonal antibodies (cat #K4001, Dako). Images of the histological sections were analyzed using the BZ-X800 analyzer software (Keyence Corp., Japan).
Cartilage repair was scored using the Wakitani histological scoring system, with reference to Safranin O staining.33,36 The maximum score in this system was 14, and a lower score indicated that the cartilage repair was closer to normal articular cartilage. Two evaluators assessed the histological sections in a blinded manner.
Data Analysis
Number of animals used per experiment is detailed in the corresponding figure legend. No inclusion/exclusion criteria used. Data are presented as mean ± standard error. We used a two-tailed Welch’s t test and Tukey’s multiple comparison test to analyze the statistical significance. Statistical significance was set at P < 0.05.
Results
Identification of Sema7A Specifically Expressed in the Cartilaginous Tissue Formed After Articular Cartilage Damage
To identify genes that are highly expressed in tissues that spontaneously form in defects in articular damage, we used data from an scRNA-seq analysis (GSE206120).
29
In that analysis, a chondral defect was created in the articular cartilage of the knee joint of a cynomolgus monkey, and the repaired tissue that spontaneously formed in the defect 4 months later (cyFT.17w.2) was harvested and subjected to scRNA-seq analysis. The intact articular cartilage (cyAC5) was harvested and analyzed as a control. After reducing the number of cells to 196 in each sample using a subset function, we performed principal component analysis, data clustering, dimension reduction using UMAP, and two-dimensional projection using Seurat. The resolution parameter was set to 0.8, which revealed four cell clusters
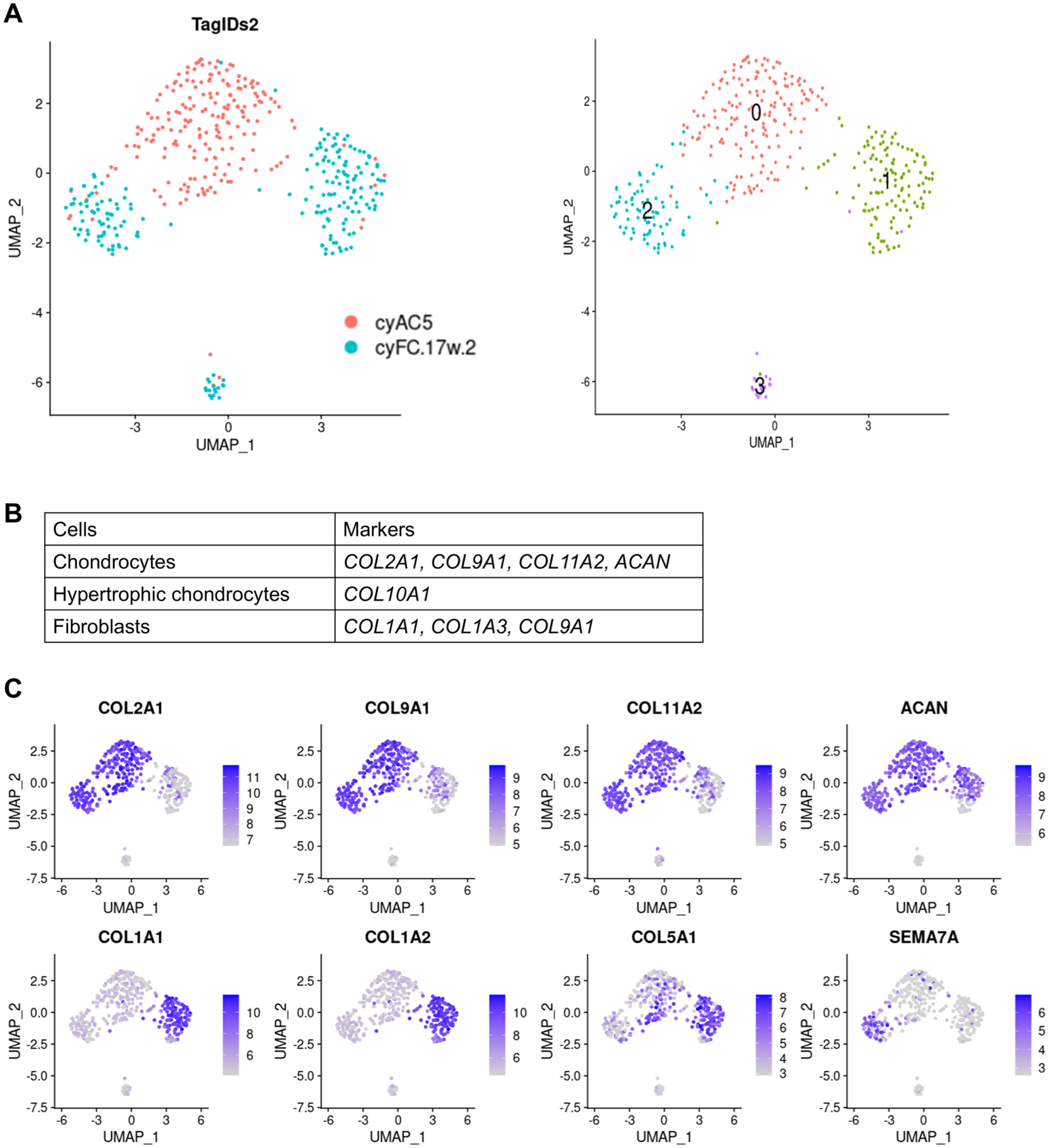
scRNA-seq analysis of intact articular cartilage and repaired tissue formed in chondral defect in articular cartilage in a macaque monkey model.
The feature plot function of marker genes (
To analyze the spatial existence of these two types of cells in the repaired tissue, we performed immunohistochemical analysis on repaired tissue that was formed 4 weeks after the creation of a chondral defect in the joint surface of the knees of mice. The repaired tissue contained two distinct areas: one area highly expressing Col2 and the other area highly expressing Col1
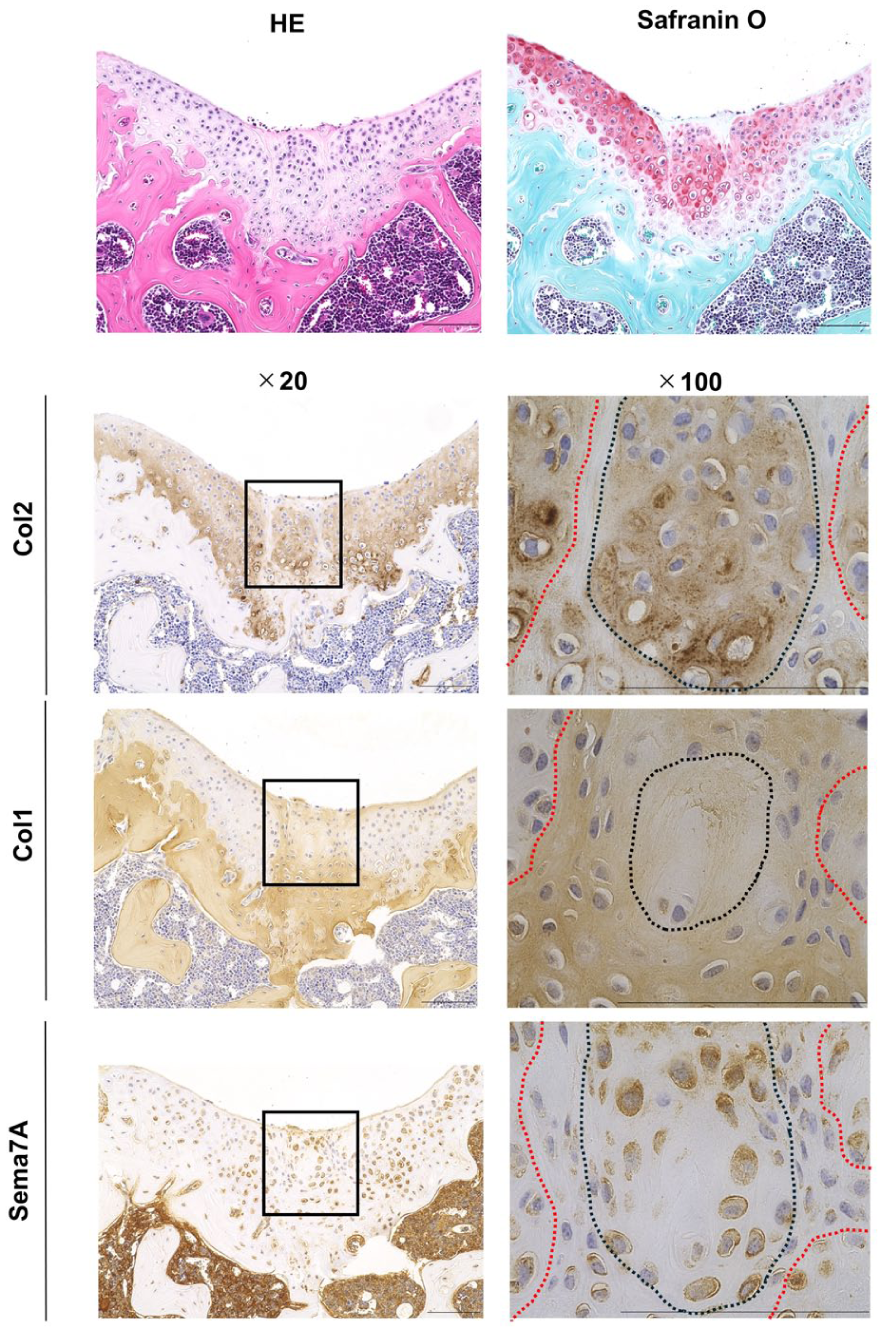
Histological analysis of repaired tissue formed in chondral defect in articular cartilage in wild-type mice. Cartilage defects were created in 8-week-old wild-type mice, and tissue sections were prepared 4 weeks after surgery. Semi-serial tissue sections were stained with hematoxylin-eosin and Safranin O-fast green-hematoxylin and immunostained for Semaphorin7a (Sema7A), type I collagen (Col1), and type II collagen (Col2). The rectangles in the left panels show the repaired cartilage area, which is enlarged in the right panels. The dotted black line indicates the boundary between strongly and weakly stained areas in the repaired tissue. The dotted red line denotes the boundary between the repaired tissue and the native articular cartilage. Data are representative of n = 6 mice.
To search for molecules that are involved in the formation of repaired tissue in articular cartilage defects, we used the FindMarkers function in Seurat to identify DEGs between cartilaginous tissue cluster 2 and the other clusters (0, 1, and 3) (
Sema7A is a GPI-anchored membrane-bound protein that is shed by the action of ADAM17 and acts as a secreted protein. 26 Therefore, Sema7A produced by cartilaginous tissue can act on both cartilaginous tissue autonomously and fibrous tissue heteronomously. To elucidate whether Sema7A affects the formation of cartilaginous or fibrous tissues in articular cartilage defects, we performed further experiments.
Sema7A Autonomously Induced Dedifferentiation of P2 Chondrocytes In Vitro
To analyze the function of Sema7A expressed in cartilaginous repaired tissue, we first employed a chondrocyte dedifferentiation model in vitro. Primary chondrocytes undergo dedifferentiation into fibroblastic cells when passaged in a monolayer expansion culture in a dish.4,5 Morphological changes that occurred due to dedifferentiation during the passage of murine primary chondrocytes were confirmed under a microscope (
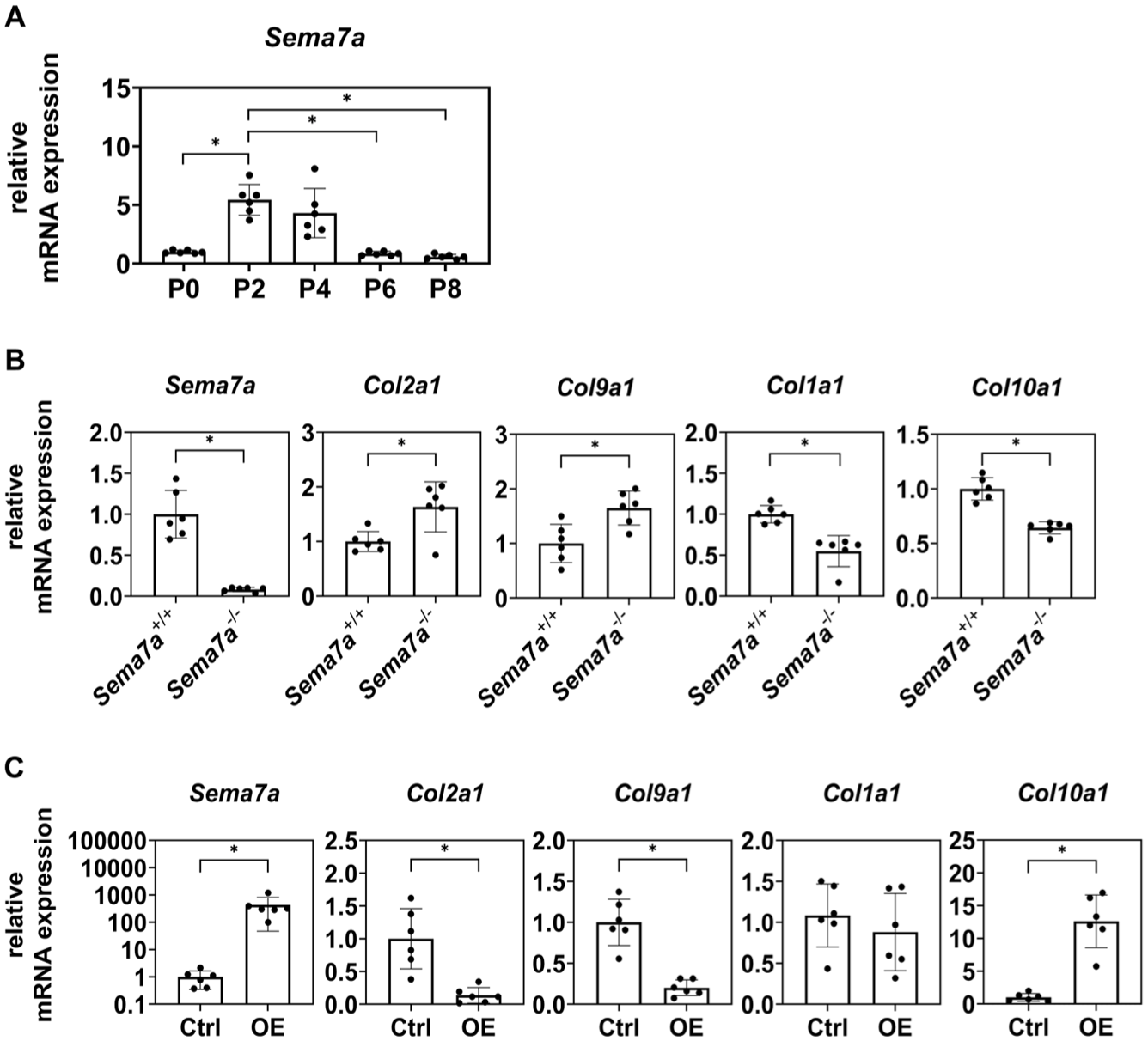
Sema7A downregulated chondrocytic differentiation of P2 chondrocytes in vitro.
To analyze the function of Sema7A, we prepared primary chondrocytes from Sema7A knockout mice (Sema7a-/-) and passaged them in a monolayer expansion culture. P2 Sema7a-/- chondrocytes expressed Col2a1 and Col9a1 at higher levels and Col1a1 and Col10a1 at lower levels than P2 Sema7a+/+ chondrocytes (
Sema7A Deletion Induced Redifferentiation of Dedifferentiated P8 Cells Toward the Chondrocytic State
The presence of cartilaginous and fibrous tissues in close proximity within the repaired tissue suggests that these two tissues interact and regulate each other. To test this hypothesis, we co-cultured P2 Sema7a-/- or Sema7a+/+ chondrocytes with P8 Sema7a+/+ chondrocytes (
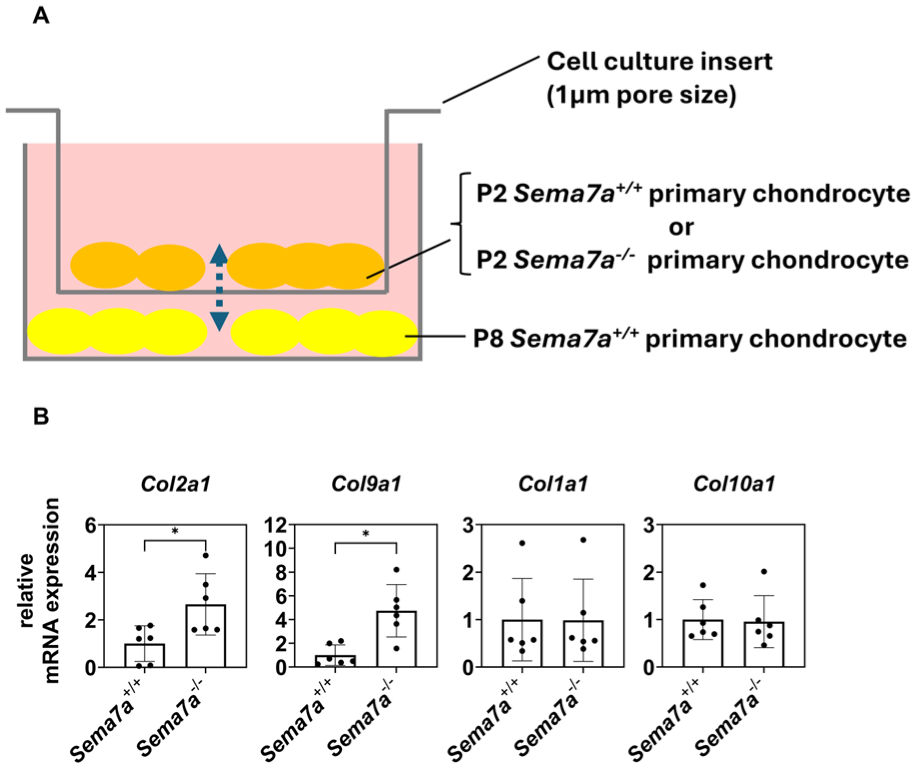
Sema7A secreted from P2 chondrocytes suppresses chondrocytic redifferentiation of P8 chondrocytes in a heteronomous manner.
Integrin β1 (ITGB1) Partly Transduced Sema7A Signal in Chondrocytes
It has been reported that Sema7A transduces intracellular signaling through ITGB1 and plexin C1 (PLXNC1).
37
FeaturePlot analysis of the scRNA-seq data on the repaired tissue in monkeys indicated that ITGB1 was ubiquitously expressed in that tissue, as well as in the articular cartilage (
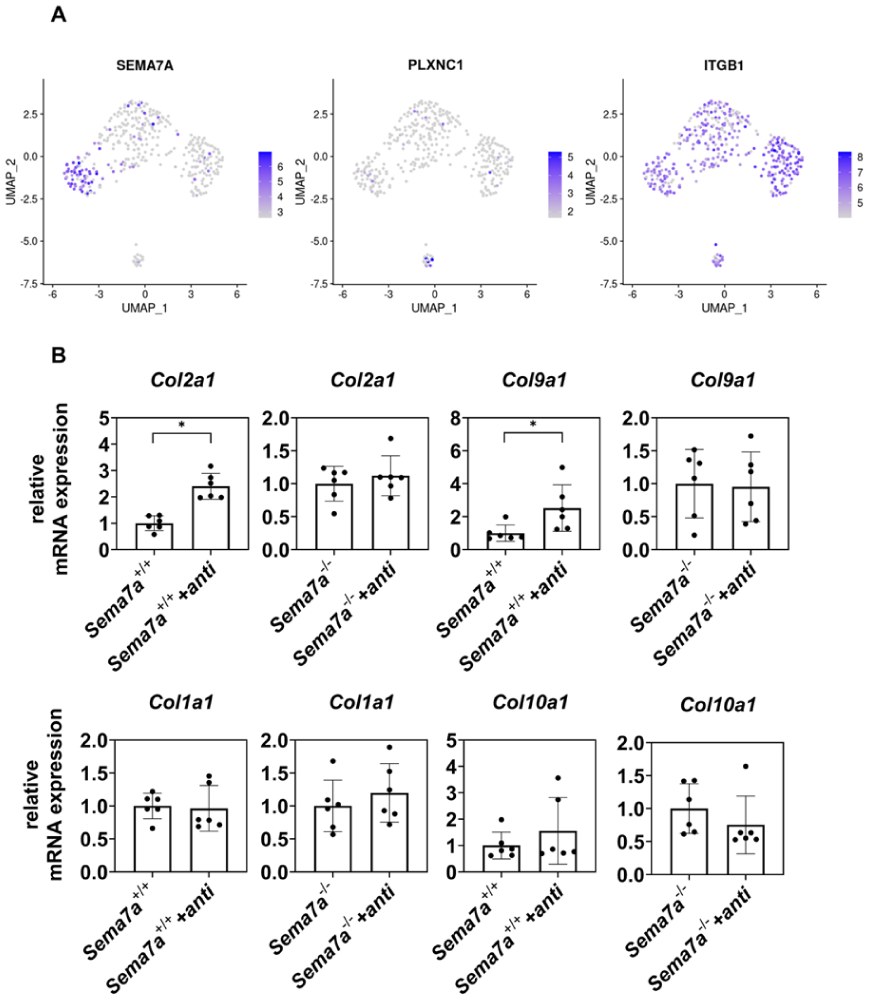
Integrin β1 (ITGB1) transduces Sema7A signal in chondrocytes.
To analyze whether ITGB1 is involved in the function of Sema7A in repaired tissue formation, we added a neutralizing anti-ITGB1 antibody to P2 Sema7a+/+ and P2 Sema7a-/- chondrocyte cultures. Addition of this neutralizing anti-ITGB1 antibody increased Col2a1 and Col9a1 expression in P2 Sema7a+/+ chondrocyte cultures, but did not affect their expression in Sema7a-/- chondrocyte cultures (
Sema7A Deletion Improved Cartilage Regeneration in Chondral Defects in the Knee Joints in Mice
Our results raised the hypothesis that Sema7A could be a target molecule for improving the regeneration of damaged articular cartilage. To test this hypothesis, we created chondral defects in distal femoral grooves of Sema7a+/- mice and Sema7a−/− mice and sacrificed them 4 weeks later. There were no significant differences in body length or weight between the Sema7a+/− and Sema7a−/− mice, either at the time of creation of the chondral defects or at the time of sacrifice (
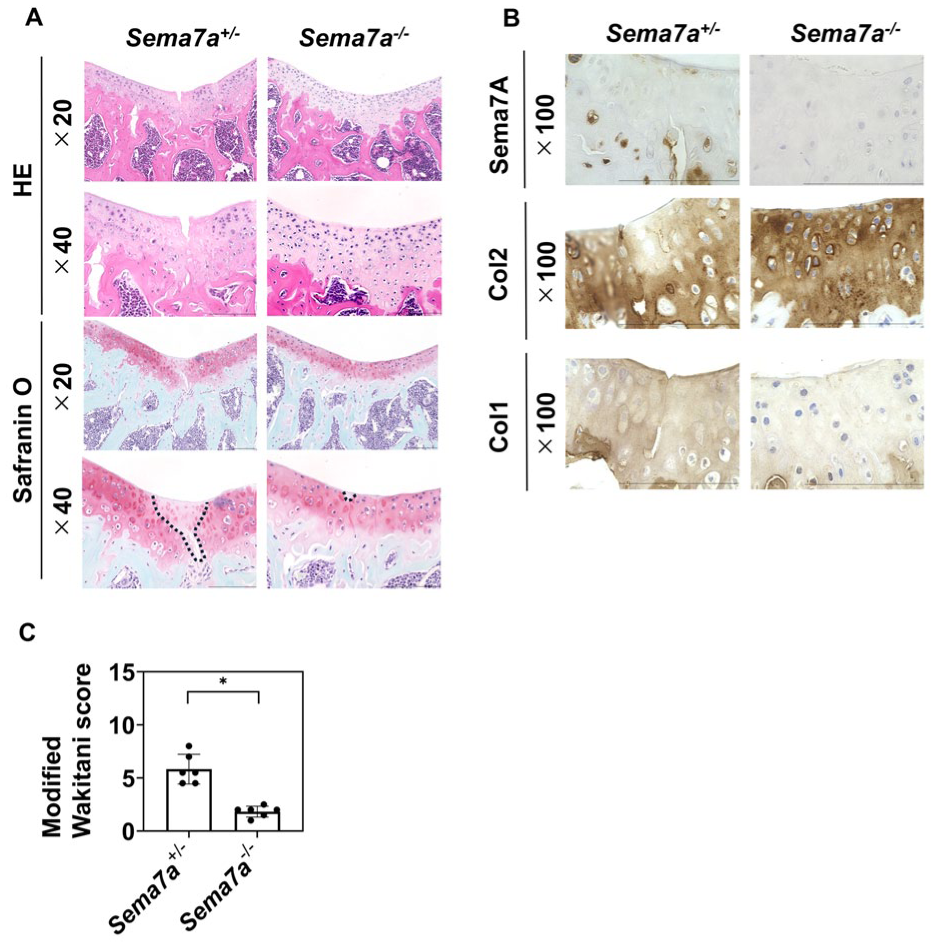
Effects of Sema7a deletion on cartilage regeneration of chondral defects in knee joins in mice. Chondral defects were created in the distal femoral grooves of Sema7a+/- and Sema7a−/− mice, which were sacrificed 4 weeks later. The samples were harvested and subjected to histological analysis.
Sema7A Deletion Did Not Affect the Development of Osteoarthritis in the Knee Joints of Mice
Next, we examined whether Sema7A deletion affects the development of osteoarthritis in adult mice. We subjected the knees of Sema7a+/- and Sema7a−/−mice to destabilized medial meniscus surgery at 8-week old and then histologically analyzed them 4 weeks after the operation. There were no significant differences in cartilage degradation between the Sema7a+/- mice and Sema7a−/− mice (
Discussion
Because of its limited capacity for self-repair, articular cartilage damage does not heal spontaneously. The repaired tissue formed in articular cartilage defects is a mixture of cartilaginous and fibrous tissues. If the balance between cartilaginous and fibrous tissues in repaired tissues could be altered, a promising therapeutic strategy would be to increase the amount of cartilaginous and decrease the amount of fibrous tissue. However, the mechanisms regulating this balance are unknown.
It has been known that primary chondrocytes lose chondrocytic characters, undergo dedifferentiation, and become fibroblastic when they are expanded in monolayer culture.5,38 -40 In contrast, dedifferentiated fibroblastic chondrocytes can be redifferentiated into chondrocytes to some extent by changes in cellular signaling molecules, such as histone deacetylase, cathepsin K, and ROCK.41 -44 Our in vitro experiments indicated that Sema7A autonomously turns P2 chondrocytes into a fibroblastic state. In contrast, Sema7A deletion heteronomously induced the redifferentiation of P8 dedifferentiated fibroblastic cells. In other words, Sema7A suppressed the redifferentiation of fibroblastic cells.
Under the assumption that P2 chondrocytic cells and P8 fibroblastic cells correspond to cartilaginous and fibrous tissue cells in repaired tissues in chondral defects, the effects of Sema7A led us to propose a model in which the balance between cartilaginous and fibrous tissues is regulated by Sema7A, which is specifically produced by cartilaginous tissue. When large cartilaginous tissue is formed, increased Sema7A expression causes the autonomous dedifferentiation of cartilaginous and heteronomous suppression of the redifferentiation of fibrous tissue (
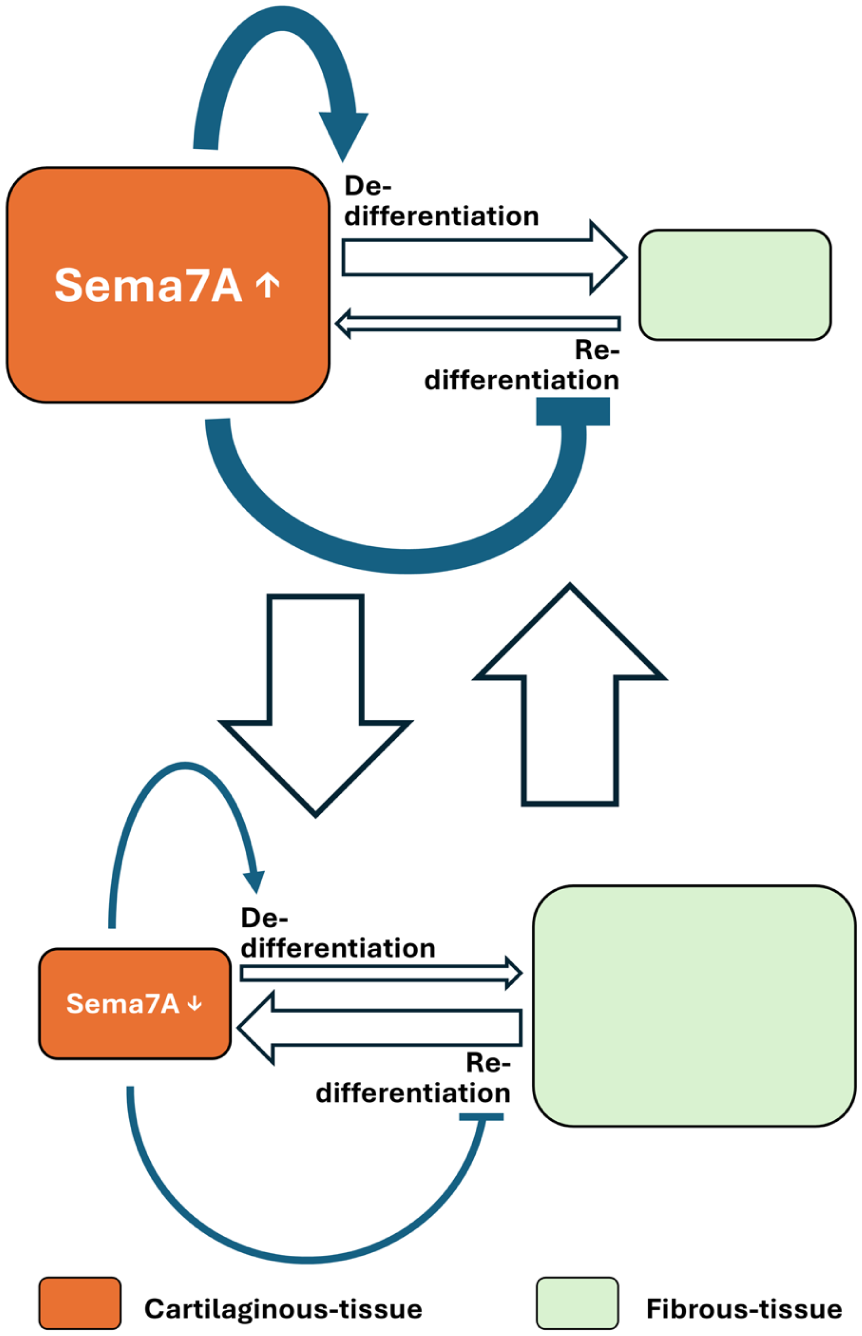
A model of how the balance between cartilaginous and fibrous tissue formations is controlled through expression of Sema7A. When a large cartilaginous tissue is formed, increased Sema7A expression causes the autonomous dedifferentiation of the cartilaginous and the heteronomous suppression of the redifferentiation of fibrous tissue (top row), leading to a reduction in cartilaginous and expansion of fibrous tissue (downward arrow). As the cartilaginous tissue reduces Sema7A expression, it decelerates the autonomous dedifferentiation of cartilaginous and heteronomous inhibition of fibrous tissue redifferentiation (bottom row), reverting to the expansion of cartilaginous and reduction of fibrous tissue (upward arrow).
The factors involved in the formation of fibrocartilage are not limited to Sema7A, and changes in the environment surrounding the chondrocytes, such as the extracellular matrix, have a significant impact. 45 Our findings suggest that Sema7A may be a therapeutic target for the regeneration of damaged articular cartilage. In contrast, Sema7A deletion did not affect the course of osteoarthritis development in mice. This discrepancy can be explained by the difference in the expression pattern of Sema7A in these two lesions; Sema7A was expressed in repaired tissue in chondral defects, but was not expressed in osteoarthritic cartilage. Thus, Sema7A plays an important role in the repair of chondral defects but is absent in the degeneration process of osteoarthritis. Chondral defects occur when the joints suffer from acute injury or trauma, whereas osteoarthritis occurs more chronically. The mechanisms regulating Sema7A expression during acute or chronic healing processes remain to be elucidated. This study will contribute to the precise identification of indications for future therapies targeting Sema7A.
When we extrapolated in vivo situations from our in vitro experimental results, we assumed that P2 and P8 cultured chondrocytes corresponded to cartilaginous- and fibrous-tissue cells, respectively, in the repaired tissue. However, this assumption has not been proven; thus, it is a limitation of the present study. Further studies are needed to confirm whether the repair process of chondral defects in joints is controlled by Sema7A as we proposed in the model.
We found that integrin β1 at least partly mediates the Sema7A function of dedifferentiating chondrocytes. Although downstream pathways of integrin β1 in repair tissues in chondral defects remains to be analyzed, previous studies of other tissues and organs suggest that the following mechanisms are involved: integrin β1 plays an important role in mediating the adherence of fibroblasts to the extracellular matrix during repair process of tissues;46,47 excess activities of fibroblasts lead to the development of scarring and fibrotic diseases;48,49 and studies have revealed that integrin β1 contributes to the formation and maintenance of these conditions in other tissues including the lung.50 -52,53
In summary, this study shows that Sema7A promotes fibrocartilage formation in chondral defects. Conventional treatments for cartilage damage have not been able to prevent the formation of fibrocartilage in repaired cartilage. The results of this study suggest that Sema7A can be a target for developing treatments to increase cartilaginous and decrease fibrous tissue in repaired tissue, leading to the healing of hyaline cartilage and improving long-term clinical outcomes.
Supplemental Material
sj-pdf-1-car-10.1177_19476035261418126 – Supplemental material for Semaphorin 7A Regulates the Balance Between Cartilaginous and Fibrous Tissues in the Repair Process of Articular Cartilage Damage
Supplemental material, sj-pdf-1-car-10.1177_19476035261418126 for Semaphorin 7A Regulates the Balance Between Cartilaginous and Fibrous Tissues in the Repair Process of Articular Cartilage Damage by Ryosuke Hatada, Kengo Abe, Akihiro Yamashita, Nanao Horike, Sho Obata, Atsushi Kumanogoh, Shuichi Matsuda and Noriyuki Tsumaki in CARTILAGE
Supplemental Material
sj-xlsx-2-car-10.1177_19476035261418126 – Supplemental material for Semaphorin 7A Regulates the Balance Between Cartilaginous and Fibrous Tissues in the Repair Process of Articular Cartilage Damage
Supplemental material, sj-xlsx-2-car-10.1177_19476035261418126 for Semaphorin 7A Regulates the Balance Between Cartilaginous and Fibrous Tissues in the Repair Process of Articular Cartilage Damage by Ryosuke Hatada, Kengo Abe, Akihiro Yamashita, Nanao Horike, Sho Obata, Atsushi Kumanogoh, Shuichi Matsuda and Noriyuki Tsumaki in CARTILAGE
Footnotes
Acknowledgements
The authors would like to thank Tomoko Kato for the preparation of the histological sections and Masayuki Nishide for the preparation of the Sema7a+/− mice.
Ethical Considerations
All methods were performed in accordance with the relevant guidelines and regulations. All animal experiments were approved by the Institutional Animal Care and Use Committee of Osaka University (approval no. 03-044-026). The experiments using recombinant DNA were approved by the Recombinant DNA Experiment Safety Committee of Osaka University (No. 04794).
Author Contributions
R.H. and N.T. are responsible for conceptualization; R.H., K.A., A.Y., and N.H. are responsible for investigation; S.O. and A.K. contributed to resources; S.M., and N.T. are responsible for supervision; and R.H. and N.T. are responsible for writing.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This study was supported by KAKENHI from the Japan Society for the Promotion of Science (JSPS) (Grant Number 23K27720); AMED under Grant Numbers 25bm1223026 (joints) and JP25bm1223014 (nucleus pulposus); and KSAC-GAP under Grant Number KSAC2025_44. This work was also supported by the World Premier International Research Center Initiative (WPI), MEXT, Japan.
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Data Availability Statement
All data needed to evaluate the conclusions in the paper are present in the paper and/or the Supplementary Materials. All the original data are available from the authors upon request.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.