
Abstract
The current research sheds light on a physiological mechanism potentially underlying confrontational responses to infidelity. Findings suggest that responses to infidelity threats in adulthood are shaped by hormonally mediated masculinization of the brain in utero. 2D:4D digit ratio (widely regarded as an index of prenatal testosterone exposure) moderated behavioral and endocrinological responses to infidelity threat. After an infidelity prime (but not a control prime), lower (more masculine) 2D:4D was associated with a greater tendency to approach attractive same-sex targets (intrasexual rivals) and with heightened increases in circulating testosterone, a hormone related to a variety of aggressive and confrontational behaviors.
On the evening of March 10, 2007, Sean Powell sat in his car, waiting for his lover, Erin McLean. With little warning, Erin’s husband Eric approached the car with a rifle, took aim, and killed Powell (ABC News, 2007). Similarly violent reactions to infidelity can be found worldwide, throughout history, and across species (Buss, 1988; Daly & Wilson, 1988; Rilling, Winslow, & Kilts, 2004). However, violence is by no means the only response to infidelity. What factors, then, underlie confrontational or even violent responses to infidelity?
The Evolutionary Psychology of Mate Guarding
Evolutionary perspectives suggest that people possess mechanisms designed to protect romantic partners from intrasexual rivals. People can suffer immense reproductive and social costs from a partner’s infidelity (Buss, 2002). Consequently, concerns about infidelity evoke a cascade of affective, cognitive, and behavioral processes aimed at guarding one’s partner from same-sex rivals (Maner, Miller, Rouby, & Gailliot, 2009). Some of those responses involve confrontation and violence. Confrontational responses may reduce the likelihood of a partner’s future indiscretion, ward off mate poachers, and help one regain social status that might have been lost due to infidelity (Buss, 1988; Buss & Shackelford, 1997; Daly & Wilson, 1988; Vandello & Cohen, 2003). However, confrontation can also come with costs: It may lead to injury, and an unsuccessful confrontation may further decrease one’s social standing. Consequently, there is likely to be variation in the extent to which infidelity prompts confrontational responses.
To understand confrontational responses to romantic rivals, it is important to examine implicit physiological and behavioral processes that may underlie those responses. Many studies have examined explicit emotional and behavioral reactions to infidelity (Buss & Shackelford, 1997). Far fewer studies have examined implicit reactions. From an evolutionary perspective, adaptive social behavior is rooted in basic, lower-order physiological and cognitive processes. Nevertheless, few studies have directly examined how responses to infidelity involve such processes (for exceptions, see Fussell, Rowe, & Park, 2011; Maner et al., 2009; Takahashi et al., 2006).
The current research helps fill this gap in the literature by investigating implicit psychological and physiological responses to the threat of infidelity. We focus on the extent to which 2D:4D digit ratio—a biomarker associated with exposure to prenatal hormones—moderates two mechanisms potentially underlying confrontational responses to infidelity. We examine a physiological mechanism (release of testosterone) and an implicit behavioral mechanism (approach-oriented action tendencies). Moreover, we examine these processes within the context of an experimental design that directly manipulates whether people are threatened by thoughts of infidelity. This approach substantially extends the literature, which has tended to employ correlational methods.
Prenatal Hormones and Responses to Infidelity
From an evolutionary perspective, variation among adult phenotypes partially reflects adaptive plasticity in response to environmental cues during early development. Much of this adaptive plasticity occurs during the first several years of childhood and into adolescence. Recent epigenetics research, for example, suggests that early life stress can interact with preexisting genetic vulnerabilities to affect long-term psychological outcomes (Canli et al., 2006). Hormonal processes play a particularly strong role in development. For example, increased release of steroid hormones (e.g., testosterone) during puberty dramatically alters nervous system structure which, in turn, influences a range of psychological and behavioral processes in adulthood (Sisk & Zehr, 2005). Understanding the nature and timing of these developmental processes provides important information about the source of individual differences later in life.
Even prior to developmental processes occurring during the first few years of life and adolescence, however, adaptive plasticity can begin occurring in utero. For example, ecological information about resource scarcity and energetic and social demands may be conveyed from mother to fetus, in turn influencing the way in which the fetus develops (Kuzawa, 2005). This influence on fetal development shapes the organization of the brain as a way of preparing the organism for similar environmental conditions likely to be encountered in the future (Bateson, 2001; Gluckman & Hanson, 2004; Kuzawa & Quinn, 2009). Indeed, abundant evidence suggests that early developmental experiences predict life history and reproductive strategies later in life (e.g., Kuzawa, McDade, Adair, & Lee, 2010).
One way in which this developmental preparation occurs is via changes in prenatal exposure to hormones. Prenatal testosterone, for example, has organizing effects on brain development, leading to permanent changes in brain structure. Modulation of phenotypes by maternal testosterone is thought to reflect an adaptive tuning to environmental conditions (Sachser, Hennessy, & Kasier, 2011). For example, in some species, maternal stress has been shown to increase fetal testosterone levels (Grant & Irwin, 2009; Guibert et al., 2011). Fetal testosterone, in turn, has been shown to speed early development, to promote increases in birth weight and the incidence of male births (Eising, Eikenaar, Schwabl, & Groothuis, 2001; Helle, Laaksonen, Adamsson, & Huitu, 2008), and to produce more competitive and dominant offspring (Schwabl, 1996). For example, in guinea pigs, daughters of mothers who lived in unstable social conditions during pregnancy displayed greater masculinized behaviors (Sachser & Kaiser, 1996), and that masculinization is linked to increases in androgen receptor functioning and serum testosterone levels (Kaiser, Kruijver, Swaab, & Sachser, 2003); such masculinization is thought to make daughters more robust and competitive in an unstable environment.
Masculinizing effects of prenatal testosterone are consistent with the initiation of a fast life history strategy. Fast life history strategies are known to result from relatively stressful and unpredictable environments (Del Giudice, Ellis, & Shirtcliff, 2011; Ellis et al., 2012). Across many species, fast life history strategies are characterized by relatively early and direct investment in reproduction. Given an unpredictable environment, investing early and directly in reproduction reflects an adaptive strategy aimed at immediately maximizing the number of one’s offspring, rather than delaying reproduction and risking the possibility of lower reproductive success in the future (Brumbach, Figueredo, & Ellis, 2009). In addition to early investment in reproduction, fast life history strategies are characterized by risk taking, impulsivity, aggression, and dominance, behaviors aimed at facilitating one’s ability to compete directly with intrasexual rivals to ensure immediate reproductive access to potential mates (Ellis et al., 2012; Griskevicius, Tybur, Delton, & Robertson, 2011; Kaplan & Gangestad, 2005; McDonald, Donnellan, & Navarrate, 2012). Thus, early developmental experiences can shape future reproductive strategies, including the extent to which people engage in direct intrasexual competition over mates. Based on this literature, one might predict that exposure to prenatal testosterone could potentiate a heightened propensity to respond competitively and aggressively toward possible rivals, particularly when the threat of infidelity is salient.
Prenatal Hormones and 2D:4D Ratio
Because prenatal testosterone exposure cannot be measured directly in adults, research has relied upon a putative indicator of prenatal testosterone—the ratio of the length of the second (index) finger to the fourth (ring) finger, otherwise known as 2D:4D. On average, men have lower 2D:4D than women—a sex difference observed as early as the first trimester of gestation (Malas, Dogan, Evcil, & Desdicioglu, 2006). Additionally, both male and female rats injected with testosterone during fetal development display shorter second and longer fourth digits during adulthood than control animals (Talarovičová, Kršková, Blažeková, 2009). There remains some question as to precisely what hormonal processes give rise to individual differences in 2D:4D. For example, one study suggested that, in men and women, 2D:4D was associated with the ratio of testosterone to estradiol (Lutchmaya, Baron-Cohen, Raggat, Knickmeyer, & Manning, 2004). Although the literature is not definitive regarding what specific hormonal processes predict 2D:4D, several lines of research suggest that lower 2D:4D reflects heightened exposure and sensitivity to fetal androgens (Breedlove, 2010; Brown, Hines, Fane, & Breedlove, 2002; Manner, Bundred, Newton, & Flanagan, 2003; Manning, 2002; Manning, Bundred, Newton, & Flanagan, 2003). There is consensus, therefore, that 2D:4D is associated with masculinizing effects of hormones in utero.
Consistent with the hypothesis that prenatal androgens shape psychological processes later in life, research demonstrates that lower 2D:4D is associated with greater adulthood dominance and masculinity (Manning & Fink, 2008; Neave, Laing, Fink, & Manning, 2003) and risk taking (Garbarino, Slonim, & Sydnor, 2011) in both men and women. The link between 2D:4D and dominance has been demonstrated in other primates, as well (e.g., rhesus macaques; Nelson, Hoffman, Gerald, & Schultz, 2010). The link between 2D:4D and dominance seems specific to aggressive forms of dominance, rather than more prosocial forms (van der Meij, Alemla, Buunk, Dubbs, & Salvador, 2011). Moreover, although there is little to no correlation between 2D:4D and baseline levels of aggression (Honekopp & Watson, 2011; Voracek & Stieger, 2009), lower 2D:4D is associated with increased aggression after provocation (Millet & Dewitte, 2007; Ronay & Galinsky, 2011).
With respect to mate guarding, the link between 2D:4D and reactive aggression is noteworthy because aggression is a common reaction to infidelity (Daly & Wilson, 1988; Miller & Maner, 2008). Men, in particular, tend to report feeling homicidal and violent after a partner’s infidelity (Shackelford, LeBlanc, & Drass, 2000), and male sexual jealousy often precipitates violent behavior (Puente & Cohen, 2003; Shackelford, Goetz, Buss, Euler, & Hoier, 2005). Violent reactions to infidelity are particularly common among individuals concerned about dominance and status (Vandello & Cohen, 2003).
Because low 2D:4D is associated with masculinity and dominance—both factors associated with infidelity-related aggression—confrontational responses to infidelity may be expressed particularly among individuals low in 2D:4D. Indeed, previous research indicates that, among men, lower 2D:4D is correlated with a greater self-reported tendency to threaten rivals who showed interest in one’s romantic partner (Cousins, Fugère, & Franklin, 2009).
However, previous research falls short of telling the whole story. Self-reports may reflect people’s conscious intentions, without necessarily tapping into more fundamental processes that presumably underlie behavior. In the current research, we predicted that low 2D:4D would be associated with two implicit processes reflecting people’s disposition toward confrontation. Moreover, whereas previous studies of 2D:4D and responses to infidelity have relied largely on correlational designs, we examine this link experimentally. This allowed us to assess the extent to which 2D:4D predicts responses that are specific to the threat of infidelity.
Lower-Order Responses to the Threat of Infidelity
Mechanisms designed to protect against relationship threats should be observed across a range of processes operating at relatively early stages of physiology and social perception (Maner, Gailliot, Rouby, & Miller, 2007; Maner, Miller, Rouby, & Gailliot, 2009). Indeed, those lower-order processes provide a foundation for higher-order forms of cognition and behavior. Here we focused on two lower-order processes—one behavioral and one physiological—potentially indicative of a confrontational mate-guarding strategy: implicit approach-biased action tendencies and increases in circulating testosterone.
Approaching Intrasexual Rivals
People possess fundamental approach and avoidance systems that are embodied within simple motor movements. Studies suggest that muscle extension (e.g., pushing a lever) is mentally represented as avoidance, whereas muscle flexion (e.g., pulling a lever) is mentally represented as approach (Cacioppo, Priester, & Bernston, 1993; Chen & Bargh, 1999; Förster, Friedman, Özelsel, & Denzler, 2006). The facilitation of simple motor movements of the hand and arm reflect the body’s implicit preparation for physically interacting with stimuli in the immediate environment (Cesario, Plaks, & Higgins, 2006; Cesario et al., 2010; Eder & Rothermund, 2008; Wilkowski & Meier, 2010). Avoidance-biased action tendencies help people evade threats by promoting escape and withdrawal. Approach-biased action tendencies, in contrast, may help people overcome social challenges, in part by promoting oppositional and confrontational behaviors (Fishbach & Shah, 2006). Pulling a joystick has been validated in many studies as an action tendency associated with approach behavior and approach-oriented patterns of neural activation (Cunningham, van Bavel, Arbuckle, Packer, & Waggoner, 2012; Ernst et al., 2013; Roelfs, Minelli, Mars, van Peer, & Toni, 2009; Voncken, Rinck, Deckers, & Lange, 2012). In the current study, we expected that, in response to infidelity threat, masculine 2D:4D would be associated with greater approach-biased action tendencies toward same-sex rivals, as indicated with a joystick task.
In the current study, we also specified the type of rival that people should find most threatening. Both men and women tend to place a premium on physical attractiveness when seeking out short-term romantic partners (Greiling & Buss, 2000; Haselton & Gangestad, 2006; Li & Kenrick, 2006; Scheib, 2001). As a consequence, infidelity concerns tend to promote vigilance toward and negative evaluations of same-sex rivals who are high in physical attractiveness (Maner et al., 2009). Thus, when threatened by infidelity, approach-biased action tendencies may be most pronounced in response to same-sex individuals who are physically attractive. Therefore, we hypothesized that, after an infidelity prime (imagining a situation in which one’s partner is found cheating), more masculine 2D:4D would be correlated with greater displays of approach (pulling a joystick) relative to avoidance (pushing a joystick), specifically in response to physically attractive, same-sex targets.
Increases in Circulating Testosterone
In addition to its organizational effects on brain development, testosterone has activational effects via current circulating levels. These activational effects influence a range of behavioral processes. In particular, increases in circulating testosterone are accompanied by increases in expressions of dominance and aggression (Geniole, Carré, & McCormick, 2011). Within the domain of mate guarding, nonhuman animal studies suggest that increases in testosterone are linked to aggressive responses to relationship threats (Wingfield, Hegner, Dufty, & Ball, 1990). For example, among male rhesus macaques, increases in testosterone are associated with aggressive behavior toward romantic interlopers (Rilling et al., 2004). Indeed, several lines of research suggest that increases in circulating testosterone prompt confrontation and aggression (Mazur & Booth, 1998). Because low 2D:4D individuals are expected to display confrontational responses to infidelity threats, we predicted that lower 2D:4D would be correlated with heightened increases in testosterone after an infidelity prime.
Overview of Hypotheses
The current study evaluated moderating effects of 2D:4D on behavioral and endocrinological indices of intrasexual confrontation in response to infidelity. We predicted that, in response to an infidelity prime (but not a control prime), lower (more masculine) 2D:4D would be associated with a greater tendency to approach attractive, same-sex targets (but not other targets), as indicated by an implicit approach-avoidance joystick task. Lower 2D:4D was also expected to correlate with increases in circulating testosterone after an infidelity prime (but not a control prime). Finally, because comparative research suggests that changes in testosterone are associated with confrontational responses to romantic interlopers (Rilling et al., 2004), we predicted that increases in testosterone after an infidelity threat would be correlated with approach-biased action tendencies in response to attractive same-sex targets.
Method
Participants
Fifty-eight undergraduates (ages 18–23; 27 women, 31 men) participated. Participants refrained from activities known to affect hormone levels: eating food or drinking caffeinated beverages or alcohol for 2 hr prior to testing; exercising for 12 hr prior to testing; smoking for 6 hr prior to testing. Seventeen additional people participated, but due to photocopier malfunction, their 2D:4D could not be calculated.
Procedure
To reduce diurnal testosterone variability, all participants arrived between noon and 5:00 p.m. Participants provided a baseline saliva sample and then were randomly assigned to undergo a procedure designed to prime concerns about infidelity or a control state (Maner et al., 2007, 2009). Participants in the infidelity condition thought of their current romantic partner or someone toward whom they had strong romantic feelings. Participants imagined a scenario in which that person was observed flirting with and being intimate with another person at a party. Participants paused at four points in the scenario to write about their visualization (see Miller & Maner, 2008, for content analyses of responses to this visualization task). In the control condition, participants imagined a distress-producing scenario in which they took and failed an important academic exam. This control condition has been shown to produce an unpleasant, high arousal state equivalent in valence and arousal to the infidelity condition (Maner et al., 2009).
Participants then completed the approach-avoidance joystick task (see below for details). Participants then completed a set of filler questionnaires prior to providing a second saliva sample. Changes in testosterone typically require 12-15 min before being fully detectable in saliva (Schultheiss & Stanton, 2009). The time spent completing questionnaires allowed for sufficient time lag between the priming manipulation and the second saliva sample. After providing the second sample, participants’ hands were photocopied to assess 2D:4D and participants were debriefed.
Measures
2D:4D
Image-based measurements of 2D:4D tend to be more reliable than direct measurements (Voracek & Offenmüller, 2007). We measured the lengths of the second and fourth digits from photocopies of the ventral surface of both hands. Measurements were made from the midpoint of the finger crease proximal to the palm to the tip of the finger using a digital caliper measuring to 0.01 mm (Manning, 2002). All measurements were performed in duplicate by two research assistants blind to condition and participant sex. Absolute-agreement intraclass correlation coefficients (ICCs) revealed excellent interrater reliability for all digits (all ICCs > .975, ps < .001). Average finger length measurements were used to calculate 2D:4D.
Testosterone Measurement
Saliva samples were frozen at −20°C. To precipitate mucins, samples were thawed and centrifuged at 4,000 rpm for 10 min. Supernatant was stored at −20°C until assayed. Commercially available solid-phase Coat-A-Count125I radioimmunoassay kits from Siemens Medical Solutions Diagnostics (Los Angeles, CA) were used to measure testosterone. Standards were diluted with distilled water (analytical range: 5–400 pg/ml). Tubes were incubated overnight after adding radio-labeled tracer. Samples were processed in duplicate using a high throughput, automated gamma counter. The intra- and inter-assay coefficients of variation were 7.8% and 10.8%, respectively.
Approach-Avoidance Task
After completing a practice block of 16 trials, participants completed two experimental blocks. Stimuli consisted of 40 facial photographs displaying neutral expressions used in previous research (Maner et al., 2009) as well as 40 neutral objects (e.g., cups, baskets, and bowls). Facial stimuli consisted of 10 attractive women, 10 attractive men, 10 average-looking women, and 10 average-looking men (for additional stimulus details and ratings of attractiveness, see Maner et al., 2009).
Participants performed two trial blocks in which they quickly categorized faces and objects by pushing a joystick forward (away from themselves, reflecting avoidance) and pulling it backward (toward themselves, reflecting approach; Förster et al., 2006). During one block, participants pushed the joystick away from themselves in response to faces and pulled the joystick toward themselves in response to objects. During the other block, participants did the reverse: pulled for faces and pushed for objects. Each block included 40 trials (20 faces and 20 objects). On each trial, a fixation cross-appeared for 2,500 ms, followed by a target stimulus. The stimulus remained visible until a joystick response was recorded. Each stimulus was shown once and appeared in random order. Order of blocks was counterbalanced.
Consistent with previous research (Cacioppo et al., 1993; Priester, Cacioppo, & Petty, 1996), we calculated a measure of relative approach by subtracting mean push reaction times from mean pull reaction times for each face type (attractive same-sex, attractive opposite-sex, average same-sex, average-opposite sex). Higher scores indicated relative facilitation of avoidance action tendencies; lower values indicated relative facilitation of approach action tendencies.
Results
Approach-Avoidance
Trials on which participants moved the joystick in the incorrect direction and trials on which a participant’s latency was greater than three standard deviations (SDs) away from his or her mean latency were excluded. On average, for each participant, 3.5% (SD = 1.6%) of trials were excluded. Two participants had unusually high latencies (mean latency was greater than three SDs above the sample mean; M = 649 ms, SD = 84 ms); their data were excluded. Preliminary analyses revealed no significant main effects or interactions associated with participant sex.
Because men, on average, have lower 2D:4D than women (see Table 1), 2D:4D values were centered within each sex. Using mixed-model analysis of covariance (ANCOVA), approach-avoidance responses were predicted from priming condition (infidelity vs. control; between-subjects), target type (attractive same-sex vs. others; within-subjects), right-hand 2D:4D (centered; between-subjects), and all interactions. 1 To account for practice effects, block order was included as a covariate in all analyses.
Mean (SD) 2D:4D by Participant Sex.

Note. SD = standard deviation.
As predicted, we observed a significant three-way interaction between priming condition, target type, and 2D:4D, F(1, 51) = 7.32, p = .009, partial η2 = .13. Follow-up analyses focused on effects of target type and 2D:4D within the infidelity and control conditions. Within the control condition, there were no significant effects. Although the target by 2D:4D interaction approached significance, F(1, 51) = 3.25, p = .08, partial η2 = .06, none of the simple effects approached significance (all ps > .13).
Within the infidelity condition, however, there was an interaction between target type and 2D:4D, F(1, 51) = 4.50, p = .039, partial η2 = .08. This was due to a significant effect of 2D:4D on approach-avoidance responses to attractive same-sex targets, F(1, 51) = 4.55, p = .038, partial η2 = .08. Among participants primed with infidelity, lower 2D:4D was associated with stronger approach-biased action tendencies toward attractive same-sex targets (Figure 1). We observed no effects of 2D:4D on responses to any of the other target types, all ps > .30.
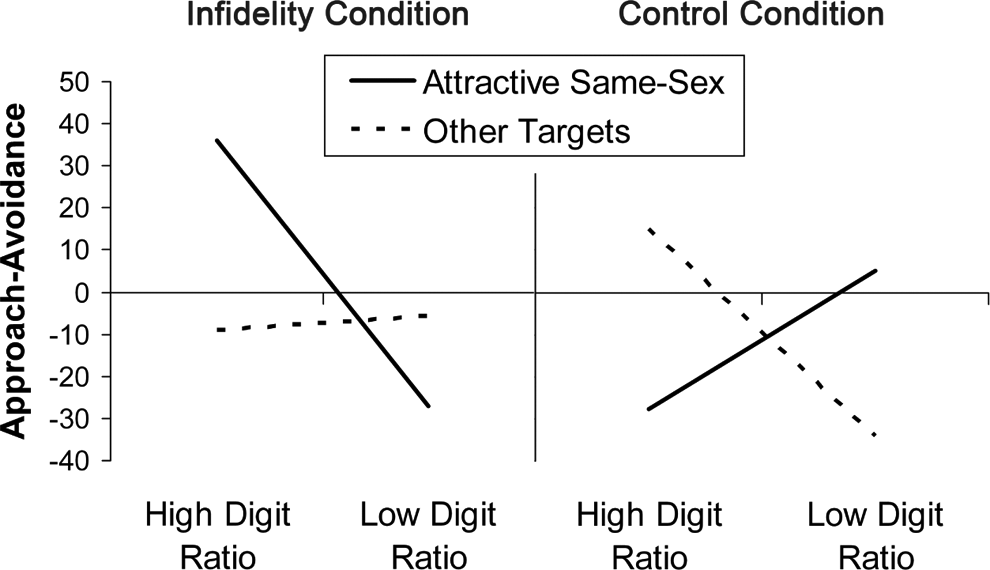
Among participants primed with infidelity, lower (more masculine) 2D:4D was associated with approach-biased responses to attractive same-sex targets. More negative values on the approach-avoidance measure correspond to more approach.
Testosterone
Testosterone concentrations were not calculated for 17 participants due to participants not providing enough saliva for adequate analysis, participants indicating that they did not refrain from confounding activities (e.g., eating, smoking), or experimenter error (failing to collect one of the saliva samples). Forty-one participants remained (23 men and 18 women). Consistent with previous research (Maner, Miller, Schmidt, & Eckel, 2008), to account for differences in testosterone variability in men versus women, changes in testosterone (post-manipulation testosterone minus baseline testosterone) were standardized within each sex (women: M = 1.2 pg/ml, SD = 5.0; men: M = 2.1 pg/ml, SD = 14.9).
Using ANCOVA, standardized change in testosterone was predicted from priming condition (infidelity vs. control), 2D:4D, and their interaction. Analyses revealed a significant interaction, F(1, 37) = 5.02, p = .031, partial η2 = .12. Among participants in the control condition, there was no effect of 2D:4D on change in testosterone, F < 1. Among participants primed with infidelity, however, there was a significant effect of 2D:4D, F(1, 37) = 4.75, p = .036, partial η2 = .11. Among participants primed with infidelity, lower 2D:4D was associated with greater increases in testosterone (Figure 2). There were no significant main effects or interactions involving participant sex.

Among participants primed with infidelity, lower (more masculine) 2D:4D was associated with greater increases in testosterone. Testosterone change scores are standardized within sex.
Ancillary Analyses
Additional analyses examined the relationship between change in testosterone and approach-avoidance responses. Among participants primed with infidelity, there was a medium-to-large (marginally significant) negative correlation between approach-avoidance responses to attractive same-sex targets and changes in testosterone, r = −.44, p = .06; increases in testosterone were associated with increases in relative approach toward attractive same-sex targets after infidelity priming. That is, when primed with infidelity, those participants who demonstrated the greatest approach-biased action tendencies toward attractive same-sex targets also tended to display the largest increases in testosterone. There was no relationship between change in testosterone and approach-avoidance responses to other targets in the infidelity condition or control condition (see Table 2).
Correlations Between Change in Testosterone and Approach-Avoidance Responses. Lower Values on the Approach-Avoidance Measure Indicate Greater Approach.

We also conducted ancillary analyses to assess whether any of the current effects were moderated by participants’ relationship status (in a committed relationship vs. single). For the approach-avoidance task, in addition to a three-way interaction between target type, 2D:4D, and priming condition, F(1, 47) = 10.71, p = .002, partial η2 = .19, we observed a four-way interaction with relationship status, F(1, 47) = 6.27, p = .016, partial η2 = .12. Follow-up analyses showed that the three-way interaction effect was significant for committed participants, F(1, 47) = 13.65, p = .001, partial η2 = .23, but not single participants, F(1, 47) = .38, p = .54, partial η2 = .01. One possible explanation is that infidelity threat is especially salient for those with a current long-term partner. Another is that the priming manipulation was more effective for committed individuals, because their priming visualization involved a current partner. However, moderation by relationship status should be interpreted cautiously, as it did not replicate for analyses involving testosterone: We observed only the aforementioned three-way interaction between priming condition and 2D:4D; testosterone reactivity was not moderated by relationship status (p = .93).
Discussion
The current findings suggest that responses to the threat of infidelity are shaped by the organizing effects of androgens in utero. 2D:4D, which reflects hormonally mediated prenatal masculinization of the brain, was associated with behavioral and endocrinological responses to infidelity priming. The responses we observed are consistent with an orientation toward confrontation and aggression.
When primed with infidelity, low 2D:4D participants were more likely than high 2D:4D participants to approach images of attractive intrasexual rivals. This effect was demonstrated using a task that assesses implicit action tendencies. Those action tendencies are thought to reflect an individual’s preparation for approaching or avoiding stimuli in the immediate environment. The approach-biased responses observed among low 2D:4D participants, therefore, are consistent with an orientation toward confronting intrasexual rivals.
Consistent with this conclusion, low 2D:4D was also related to heightened increases in testosterone after infidelity priming. Moreover, increases in testosterone were associated with greater tendencies to approach attractive same-sex rivals. This fits with primate research demonstrating a relationship between testosterone and aggression toward romantic interlopers (Rilling et al., 2004). Indeed, testosterone is tied to confrontational behaviors, competitiveness, and aggression (Geniole et al., 2011; Mazur & Booth, 1998). Thus, the overall pattern of results across two different measures suggests that masculinization of the brain in utero (as indexed by 2D:4D) may potentiate confrontational responses to reproductive threats later in life.
These findings fit with evidence for functional plasticity in prenatal development. Variation in prenatal testosterone exposure may partially reflect a maternal process through which offspring are prepared to develop and behave optimally given prevailing environmental conditions (Dufty, Clobert, & Moller, 2002). Prenatal testosterone may be associated with confrontational responses to threatening intrasexual rivals as an adaptation to environmental instability. Environmental instability during early development may incline offspring toward a faster life history strategy, which entails heightened intrasexual competition, aggression, and risk taking (Kaplan & Gangestad, 2005). High prenatal testosterone may prepare individuals to respond confrontationally to reproductive threats, and this propensity may persist into adulthood.
Some have questioned associations between 2D:4D and psychological phenomena. Although some studies report significant correlations between 2D:4D and aggression, others have failed to find such a link (Honekopp & Watson, 2011; Voracek & Stieger, 2009). One potential explanation for these inconsistencies involves the distinction between baseline behavior versus reactivity to environmental triggers. Much of the 2D:4D literature presumes correlations between prenatal testosterone and baseline behavioral processes. Consequently, researchers typically have assessed relationships between 2D:4D and psychological phenomena without examining environmental triggers. However, there are reasons to suspect that prenatal endocrine effects on brain organization potentiate behaviors in response to environmental cues (Kemme, Kaiser, & Sachser, 2007). By using an experimental design in which we manipulated threat of infidelity, we were able to identify a pattern of reactivity to threat among individuals with masculine 2D:4D. Only when primed with infidelity did 2D:4D predict approach action tendencies and testosterone reactivity. Heightened prenatal testosterone exposure may predispose individuals toward confrontation and aggression primarily when people are threatened (Millet & Dewitte, 2007; Ronay & Galinsky, 2011).
Limitations and Future Directions
Limitations provide valuable opportunities for further investigation. One limitation is that, although this research may have implications for understanding aggressive or confrontational behavior, we did not measure those behaviors. The current research serves as a useful springboard from which to further investigate the role of prenatal testosterone in shaping violent and adversarial responses to infidelity.
Another limitation pertains to the fact that we focused only on responses to intrasexual rivals. Romantic partners are also common targets of infidelity-related aggression, particularly among individuals concerned about dominance and status (Shackelford et al., 2005; Vandello & Cohen, 2003). A useful avenue for future research would be to examine the role of 2D:4D in intimate partner violence.
Another useful avenue for future research involves participant sex. We found no effects of participant sex in this research. This is consistent with prior research demonstrating similar effects of 2D:4D on vengeful responses to provocation in men and women (Ronay & Galinsky, 2011). However, other research suggests sexually differentiated associations between 2D:4D and social behavior. Cousins, Fugère, and Franklin (2009) found a negative correlation between 2D:4D and self-reported tendencies to threaten intrasexual rivals among men, but not women. One possibility is that men and women may respond to infidelity primes in a similar fashion at early stages of social perception (see Maner et al., 2009), but those responses may facilitate different behaviors in the two sexes. Future research would benefit from identifying where in the stream of processing the sexes differ in responses to infidelity.
Conclusion
Confrontational and violent reactions to infidelity have been observed throughout history, across cultures, and among a variety of species. The current findings offer novel insight into a physiological mechanism potentially underlying confrontational responses to infidelity. We provide evidence that prenatal androgen processes potentiate implicit behavioral and endocrinological infidelity responses that could underlie overt aggressive and confrontational behaviors. The current research grounds our understanding of responses to relationship threat in physiological processes shaped by natural selection. Carefully considering evolutionary and biological perspectives—as well as directly assessing the proximate physiological processes underlying social behavior—has powerful implications for furthering our understanding of social relationships.
Footnotes
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: Preparation of this article was partially supported by National Science Foundation grant BCS-1122785 awarded to the first author.