
Abstract
Recent research has demonstrated that ascribing minds to humanlike stimuli is a product of both their perceptual similarity to human faces and whether they engaged configural face processing. We present the findings of two experiments in which we both manipulate the amount of humanlike features in faces (in a doll-to-human morph continuum) and manipulate perceivers’ ability to employ configural face processing (via face inversion) while measuring explicit ratings of mind ascription (Study 1) and the spontaneous activation of humanlike concepts (Study 2). In both studies, we find novel evidence that ascribing minds to entities is an interactive product of both having strong perceptual similarity to human faces and being processed using configural processing mechanisms typical of normal face perception. In short, ascribing mind to others is bounded jointly by the featural cues of the target and by processes employed by the perceiver.
Identifying other minds is a crucial aspect of social cognition (Dennet, 1996). Indeed, ascribing another entity a mind seems to be a necessary condition for social cognition to be engaged. Put simply, before we turn on social processing, we need to know who or what requires it in the first place. The signal that another entity has a mind can come from a variety of sources (Wegner & Gray, 2016); however, one commonly used signal for mind is the presence of a face. In the current work, we investigate how and under what conditions the perception of humanlike faces translates into the ascription of humanlike minds.
Past research has reliably shown that the human perceptual system appears hardwired to attune to human faces (e.g., Langton, Law, Burton, & Schweinberger, 2008; Ro, Russell, & Lavie, 2001; see Hugenberg & Wilson, 2013), and this attunement is so potent that even the presence of humanlike facial features, such as eyes, can make nonbiological stimuli appear animate (Gao, McCarthy, & Scholl, 2010). Intuitively, we know immediately when we see human faces that they harbor human minds. They are animate (i.e., alive) and have sophisticated faculties that allow for both sensation and the ability to cogitate and act on the world (i.e., experience and agency; see Gray, Gray, & Wegner, 2007). 1 Despite this immediate experiential understanding that fellow human faces harbor fellow human minds, only limited research has investigated this face perception-to-mind inference cascade to clarify how and under what conditions apparent human faces signal actual human minds (Koldewyn, Hanus, & Balas, 2014; Looser & Wheatley, 2010). In the current research, we seek to address these questions directly. We provide evidence across two studies that mind is perceived in a face only when the features of the face are perceptually similar enough to humans to produce a sense of animacy and the features of the face can be processed configurally.
Face Perception and Mind Perception
Research has reliably demonstrated that mind ascription can be influenced via both top-down motives and bottom-up perceptual cues. The majority of the top-down effects can be summarized by noting that the ascription of mind to others seems to occur most often for selfish reasons, such as to help fulfill motives for social connection (e.g., Epley, Akalis, Waytz, & Cacioppo, 2008; Powers, Worsham, Freeman, Wheatley, & Heatherton, 2014), the need to prepare for an impending interaction (Khalid, Deska, & Hugenberg, in press), effectance motives (Epley, Waytz, & Cacioppo, 2007), and for intergroup motives (e.g., Castano & Giner-Sorolla, 2006; Hackel, Looser, & Van Bavel, 2014; Haslam, 2014). Although these motivated, top-down triggers of mind perception are important and well established in the literature, the current work investigates how faces can trigger mind perception via bottom-up processes. Comparatively, the possibility that perceptual processes influence mind ascription has received relatively short shrift in most models of mind perception (e.g., Gray et al., 2007). Indeed, only recently have scientists begun to investigate how faces can trigger the experience that targets have minds (Koldewyn et al., 2014; Looser & Wheatley, 2010). Specifically, extant evidence indicates that both facial features and facial configurations can signal mind.
Human faces have both features typical of humans, such as the presence and regular proportions of the eyes, nose, and mouth, as well as a regular configuration of those features (an eyes-over-nose-over-mouth configuration). Further, both features and configurations can signal a humanlike mind. For example, in their seminal work on this topic, Looser and Wheatley (2010) had participants judge whether targets that varied monotonically in their perceptual similarity to humans were animate using a scale ranging from definitely alive to definitely not alive. Their stimuli consisted of facial stimuli that were morphed between a doll face (i.e., definitely inanimate) and a human face (i.e., definitely animate). Consistent with their predictions, as the stimuli included a greater proportion of human relative to doll features, participants were more likely to perceive the targets as alive. Importantly, however, these effects were nonlinear. Perceivers’ animacy judgments fit a sigmoidal curve that was shifted toward the human end point of the continuum. This suggests that the perception of animacy is both categorical, in that targets were perceived to be either alive or not alive, and relatively conservative, in that morphed stimuli needed to be objectively more than 50% human to be subjectively perceived as animate. Nearly identical (r = .92) results occurred when participants rated the same stimuli on how likely it was that the target possessed a mind, suggesting that judgments of mind and animacy are essentially synonymous when evaluating human faces (see also Gao et al., 2010). To have a humanlike face is to have a humanlike mind.
Looser and Wheatley (2010) provided further evidence for the categorical nature of animacy by having participants complete a same–different categorization task. Participants were shown pairs of faces that were both below the animacy threshold, both above the animacy threshold, or that straddled the animacy threshold. Results demonstrated that participants were more sensitive to differences in faces that straddled the animacy threshold than they were to differences in faces that were both on one side of the threshold, even holding the objective difference between stimuli constant. These findings demonstrate that there is a “tipping point” at which the objective signal of humanness present in targets’ features gives rise to subjective perceptions of animacy. Notably, this threshold is sensitive to perceiver motives. For example, using the morph paradigm developed by Looser and Wheatley (2010), Hackel, Looser, and Van Bavel (2014) demonstrated that stimuli required greater perceptual similarity to humans to be ascribed mind when the target was believed to be an outgroup member relative to when it was believed to be an in-group member, especially among highly identified perceivers. Replicating the original findings, the bottom-up influence of the humanlike facial features on the perception of mind was robust and consistent, even in the presence of top-down motives. Thus, research consistently demonstrates that facial features can give rise to the perception of mind.
The configuration of facial features also serves as a bottom-up signal of mind. Indeed, whereas most stimuli are processed via a piecemeal integration of their features, people use configural processing to process human faces as an integrated Gestalt (Farah, Wilson, Drain, & Tanaka, 1998; see Maurer, Le Grand, & Mondloch, 2002, for a review). Configural processing appears to help explain humans’ great facility in processing faces, but not a vast variety of other nonface stimuli (e.g., airplanes and houses). Germane to the current work, recent research has demonstrated that processing faces configurally can lead to the perceptions of mind such that faces encoded using configural processing are ascribed more mind than are faces for which configural face processing is disrupted, even when visual information in the faces is otherwise identical. Hugenberg and colleagues (2016) demonstrate that disrupting perceivers’ ability to process faces in a configural manner (i.e., by inverting them; see Yin, 1969) undermines the face-to-mind cascade. In their first study, participants completed a lexical decision task (LDT) in which they were exposed to upright and inverted faces and asked to categorize letter strings as words or nonwords. Participants were quicker to categorize words related to humans (e.g., soul) after being exposed to upright faces compared to inverted faces, demonstrating that disrupting configural face processing inhibits the activation of concepts related to humanness. In a subsequent study, Hugenberg and colleagues (2016) showed that, compared to upright human faces, inverted human faces were ascribed lower levels of mental characteristics seen as unique to humans (e.g., thoughtfulness, empathy, and creativity). In other words, disrupting configural face processing caused perceivers to infer that others lacked sophisticated, uniquely human mental faculties (see also Fincher & Tetlock, 2016). Thus, evidence exists to suggest that the perception of mind in others is spontaneously generated when configural face processing occurs.
Current Work
The current work relies on the dissociation of face and mind afforded by comparing faces of stimuli that clearly have minds (i.e., humans) with face-like stimuli that clearly do not possess minds (i.e., dolls) to help understand how faces can signal minds. Such a comparison allows us to disentangle the visual information from the social information; both dolls and humans have faces, yet only the human face harbors a mind. Whereas doll faces may activate the face detection system, dolls are neither ascribed minds nor do they generate the sustained processing typical of actual faces (Wheatley, Weinberg, Looser, Moran, & Hajcak, 2011).
In the current work, we propose that the perception of mind arises out of the unique combination of the featural and configural signals that exist in faces. In other words, perceivers infer that others have minds when they share sufficient perceptual similarity with humans so as to be judged animate and they evoke configural processing, processing typically reserved for human faces. There is a good reason to believe that it may be the unique combination of these two processes may interact to generate ascriptions of mind.
Although recent research has demonstrated that configural processing and ascriptions of mind are linked (i.e., disrupting configural processing disrupts ascription of mind), not all stimuli that are processed configurally are spontaneously perceived to have minds. Indeed, with sufficient expertise, even clearly inanimate stimuli can be processed configurally. For example, fluent readers of English process Roman letters configurally (Wong et al., 2011). And yet, the letters on this page likely have not sprung to life in the minds of most readers. Although configural processing appears to influence ascriptions of mind (Hugenberg et al., 2016), it likely only does so when characteristics of the stimulus suggest it is alive. Work by Crookes and colleagues (2015) shows that perceivers have worse memory for computer-generated faces than real faces, suggesting that the perception of mind may be critical for important face perception processes (e.g., encoding identity) to engage.
From our perspective, manipulations of configural processing should influence ascriptions of mind, but only when they appear to be animate as a result of their perceptual similarity to humans. In other words, disrupting facial configurations (e.g., via face inversion) should only influence ascriptions of mind for faces that are plausibly human (i.e., include a large proportion of humanlike features). We conducted two studies in which we monotonically manipulated the objective perceptual similarity of stimuli to humans using Looser and Wheatley’s (2010) doll-to-human morph stimuli (see also Balas & Horski, 2012; Cheetham, Suter, & Jäncke, 2011; Hackel et al., 2014; Powers et al., 2014). 2 Separately, we manipulated participants’ ability to process stimuli configurally. To do so, we relied on face inversion (Yin, 1969). Face inversion is the gold-standard manipulation of configural face processing because it disrupts the eyes-over-nose-over-mouth configuration of the face, and therefore the ability to use configural processing, while still maintaining the objective features of the face and resembling a face than can appear in vivo (Maurer et al., 2002).
We hypothesized that increasing signals of humanlike features (i.e., more human morphs) in configurally processed upright faces would be associated with increases in mind perception, replicating past research. Of interest was whether the link between configural face processing and mind ascription was only observed for stimuli with high levels of human features. Specifically, we predicted that for faces that had nonhuman (i.e., highly doll-like) features, even humanlike configurations (i.e., upright) of those features would be insufficient to produce mind ascription. Instead, only when faces had human features and a humanlike configuration should perceivers ascribe mind to them. In Study 1, we measured explicit mind ascriptions of the upright and inverted morph stimuli, closely replicating the procedure from Looser and Wheatley (2010). In Study 2, we measured the extent to which the upright and inverted human and doll stimuli led to spontaneous activation of humanlike concepts in a LDT (see Hugenberg et al., 2016).
Study 1
Study 1 was designed to provide initial evidence that ascriptions of mind are bounded by both perceptual similarities to humans and configural face processing. To investigate this, we used a method adapted from Looser and Wheatley (2010, Study 1b). Specifically, we showed participants face-like stimuli that monotonically varied along a continuum from nonhuman to human. Critically, half of the targets were inverted, thus disrupting the ability for perceivers to use configural face processing. Participants were asked to rate the extent to which they believed each target had a mind. Because configural face processing is typically reserved for human faces, we hypothesized that inverted targets would be judged less likely to possess mind than upright targets, but only when targets emitted a sufficiently strong signal of humanness.
Method
Participants
We conducted a power analysis using G*Power 3.1, (Faul, Erdfelder, Lang, & Buchner, 2007) for a 2 × 11 interaction, assuming a small effect (ηp 2 = .05) and a .8 correlation between measures. This suggested a target N of 44 for 95% power. Forty-four participants (M age = 38.44, SD = 14.78) were recruited via Amazon’s Mechanical Turk and were remunerated US$0.40 for their participation. Most participants identified as White (81.8%) and female (54.5%). No participants were excluded from analysis; analyses were conducted only after data collection was complete.
Materials
Our stimuli were comprised of a subset of the morphed images used in past research (Hackel et al., 2014; Looser & Wheatley, 2010). Eight human faces served as base stimuli. These eight images were each morphed with a nonhuman face-like stimulus (i.e., a structurally similar doll or statue face; see Figure 1). Each face/doll pair was morphed in 10% increments along the morph continuum; thus, we had 11 images for each face identity (i.e., 0% human/100% doll, 10% human/90% doll, etc.). We further manipulated face orientation such that each set of images had upright and inverted versions. In total, this generated 176 stimuli.

Six sample morph stimuli (of 11) for two face identities. Participants saw morphs between human and humanlike faces along 11 equidistant points, presented upright and inverted.
Procedure
After providing informed consent, participants were informed that they would see a series of faces that varied in how human they seemed. Their task was to rate the extent to which they believed each target possessed a mind. Following Hackel and colleagues (2014), we defined mind by asking our participants to consider how a human mind is different from that of an animal or robot. Participants were then shown, one at a time, all 11 upright faces from 4 of the face identities and all 11 inverted faces from the remaining 4 face identities. Presentation order of face identities was independently randomized for each participant. The set of faces participants saw upright versus inverted was counterbalanced between subjects. Counterbalancing had no effects. Concurrent with the presentation of each stimulus, participants were asked to rate the extent to which the face has a mind on a 7-point scale, ranging from 1 (definitely does not have a mind) to 7 (definitely does have a mind).
After viewing all stimuli, participants were asked to provide demographic information (i.e., sex, age, race), were fully debriefed, and were provided their completion code to receive compensation. Compensation was not withheld for any participants.
Results and Discussion
Of primary interest was the extent to which face inversion interacted with the objective signal of humanness (i.e., morph level) in attributions of mind. To investigate this, we computed mean values for the mind attribution item separately for upright and inverted targets at each level of face morph. These values were submitted to a 2 (face orientation: upright vs. inverted) × 11 (morph level) repeated-measures analysis of variance (ANOVA). This analysis yielded a main effect of morph percentage, F(10, 430) = 238.22, p < .001, ηp 2 = .85. Replicating past research, stimuli that contain an objectively stronger signal of humanity were perceived as more likely to have a mind. Additionally, the average point of subjective equality (PSE)—the point at which a face was deemed equally likely to definitely have a mind or definitely not have a mind—was shifted past the midpoint for both the upright (M = .61, 95% CI [.55, .67], SD = .20), t(43) = 3.57, p = .001, d = 1.09, and inverted (M = .64, 95% CI [.57, .71], SD = .22), t(43) = 4.28, p < .001, d = 1.31 faces. The PSE was marginally further from the midpoint for inverted as compared to upright faces, t(43) = 1.85, p = .07, d = .28.
The ANOVA also yielded the predicted interaction between face orientation and morph percentage, F(10, 430) = 2.93, p = .001, ηp 2 = .06. To better understand the nature of this interaction, pairwise comparisons were computed at each of the 11 morph levels. We expected to see inversion influence mind ascription, but only when targets emitted a sufficiently strong signal of humanness, such that they were on the human side of the PSE. Indeed, for stimuli between 0% and 60% human, inversion did not influence mind attributions (ps > .31, ds < .16). However, stimuli that were past the upright PSE (i.e., between 70% and 100% human) were influenced by inversion 70%, t(43) = −2.73, p = .009, d = −.41; 80%, t(43) = −2.15, p = .037, d = −.32; 90%, t(43) = −2.81, p = .007, d = −.42; 100%, t(43) = −2.32, p = .025, d = −.35. As seen in Figure 2, inverted targets that were sufficiently human (i.e., past the upright PSE) were rated as less likely to possess mind than their upright counterparts. Moreover, a comparison of effect sizes (ds = −.41, −.32, −.42, and −.35, for each 70%, 80%, 90%, and 100% human morph percentage, respectively) suggests that once targets were perceived as alive—passing Looser and Wheatley’s (2010) “tipping point of animacy”—the inversion manipulation had a relatively stable effect on ascriptions of mind.
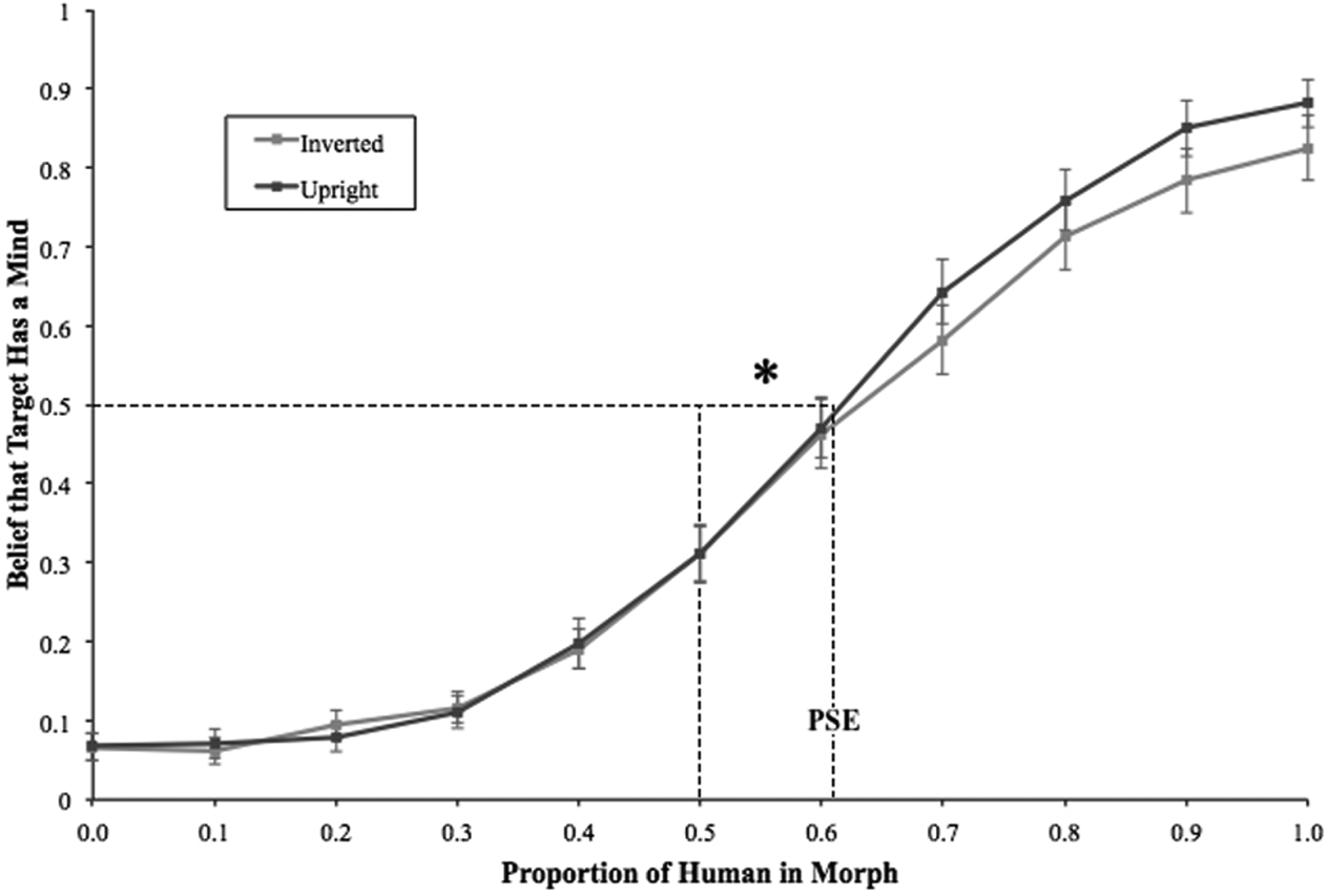
Participants’ beliefs about target’s minds in Study 1. Participants were more likely to believe that upright targets had mind compared to inverted targets, but only when targets had an objectively strong signal of humanness (i.e., they were past the point of subjective equality). Error bars represent standard error of the mean.
These results suggest that ascriptions of mind can emerge as the interactive product of the perceptual similarity of targets to humans and the processing style used to encode them. Specifically, we replicated the work of Looser and Wheatley (2010) by demonstrating that there is a clear categorical boundary of mind ascription, and that this threshold is only surpassed with a sufficiently strong signal of humanness from the features themselves. Additionally, we replicate the work of Hugenberg and colleagues (2016) by demonstrating that inversion biases mind ascription. Most importantly, we demonstrate that these effects are interactive—configural processing only has its effects on mind ascription in the presence of strongly humanlike features.
Study 2
Study 1 demonstrated that explicit ascriptions of mind arise from the interaction of featural similarities to human faces and configural face processing. However, the mere possession of a mind does not necessarily imply a fully humanlike mind. In Study 1, participants specified only that a target has a mind, not what faculties that mind possesses. In Study 2, we extend the findings of Study 1 beyond categorical ascriptions of mind to the spontaneous activation of concepts related to humanness. Indeed, recent research linking configural face processing to the perceptions of humanness has demonstrated that upright but not inverted human faces spontaneously activate concepts related to humanness (e.g., personality, soul; see Hugenberg et al., 2016).
Specifically, we hypothesized that humanlike concepts would be activated by the joint presence of strongly humanlike (but not doll-like) features and upright (but not inverted) faces. Conversely, we hypothesized that the ability to process a face configurally would not activate concepts related to humanness for inanimate targets (i.e., humanlike dolls), regardless of whether stimuli were presented upright or inverted. To test this hypothesis, we employed a method similar to that of Hugenberg and colleagues (2016, Study 1). Here, participants completed an LDT in which they were asked to categorize letter strings as either words or nonwords. The presentation of letter strings was preceded by the appearance of either an upright or inverted face that was either unambiguously human or nonhuman. Within-subjects, words were manipulated to be related to humans or machines. We predicted that upright human faces would facilitate recognition of human-related but not machine-related words, whereas nonhuman faces would facilitate recognition of neither human nor machine words, regardless of orientation.
Method
Participants
The design of Study 2 was adapted from Hugenberg and colleagues (2016), which employed 54 participants; however, given that we could not estimate whether the current effects would be larger or smaller than those in Hugenberg and colleagues (2016), we targeted at least 75 participants. In actuality, 84 undergraduate students (M age = 18.81, SD = 1.37) participated for partial course credit. The majority of our sample identified as White (85.7%) and female (69.0%). No participants were excluded from analysis; analyses were conducted only after data collection was complete.
Materials
Twelve base images served as primes. Specifically, six base images were taken from the fully human face set of stimuli used in Study 1; the other six base images were the fully nonhuman counterparts of those. Each base stimulus had an upright and inverted version, netting 24 total primes. Twelve words were used as targets for the LDT. We used words adapted from past research (Hugenberg et al., 2016) pretested to be related to humanness (i.e., human, person, individual, soul, personality, people) and machines (i.e., machine, computer, robot, device, engine, locomotive). We also used 12 pronounceable nonwords as foils. Presentation of faces and words was independently randomized for each participant.
Procedure
Participants completed a computerized LDT. After providing informed consent, participants were informed that we were interested in their ability to categorize letter strings as either words or nonwords. Participants were additionally informed that we were interested in their ability to complete this task while distracted; thus, they would be shown pictures of faces before each letter string. Participants were told to ignore the faces and focus on the word categorization task. Their task was to simply categorize each letter string as a word or nonword as quickly and as accurately as possible.
This LDT consisted of 192 trials. Every trial began with a fixation cross (1,000 ms) that was followed by a face prime (100 ms). After the face prime, a letter string appeared on screen where it remained until participants categorized it, via keystroke, as a word or nonword. Prime type (i.e., human face vs. nonhuman face) and orientation (i.e., inverted vs. upright) were manipulated within subjects, such that 96 trials contained upright faces and 96 trials contained inverted faces, of which half were human faces and half were nonhuman faces. On the critical trials that contained words, half of the words were related to humans and the other half were related to machines.
After completing the LDT, demographic information was requested and participants were fully debriefed.
Results and Discussion
Of primary interest was the extent to which human-related concepts were differentially activated by upright and inverted human and nonhuman face stimuli. In accordance with past research (Boucher & Rydell, 2012; Hugenberg et al., 2016), we first eliminated incorrect responses and responses with latencies faster than 300 ms or slower than 1,500 ms (9.02% of trials). 3 The remaining response latencies were averaged for each trial type of the 2 (prime orientation: upright vs. inverted) × 2 (prime type: human vs. nonhuman) × 3 (word type: human vs. machine vs. nonword) design. These values were then submitted to a repeated measures ANOVA. This analysis yielded the predicted three-way interaction, F(2, 162) = 3.55, p = .031, ηp 2 = .04, observed power = .72.
We decomposed this three-way interaction into the two-way interactions of prime orientation and prime type, separately at each word type. We turn first to human words. As shown in Figure 3, this analysis yielded a significant interaction, F(1, 81) = 3.38, p = .04, ηp 2 = .05, observed power = .54. Pairwise comparisons revealed that, as hypothesized, participants were quicker to recognize human words when they were primed with an upright human face (M = 553.09, 95% CI [536.63, 569.54], SD = 74.90) relative to either an upright nonhuman face (M = 577.71, 95% CI [559.65, 596.78], SD = 82.22), t(81) = −3.60, p = .001, d = −.40, or an inverted human face (M = 582.00, 95% CI [562.73, 601.27], SD = 87.69), t(81) = −4.50, p < .001 d = −.50. No other pairwise comparisons were significant (ps > .33).
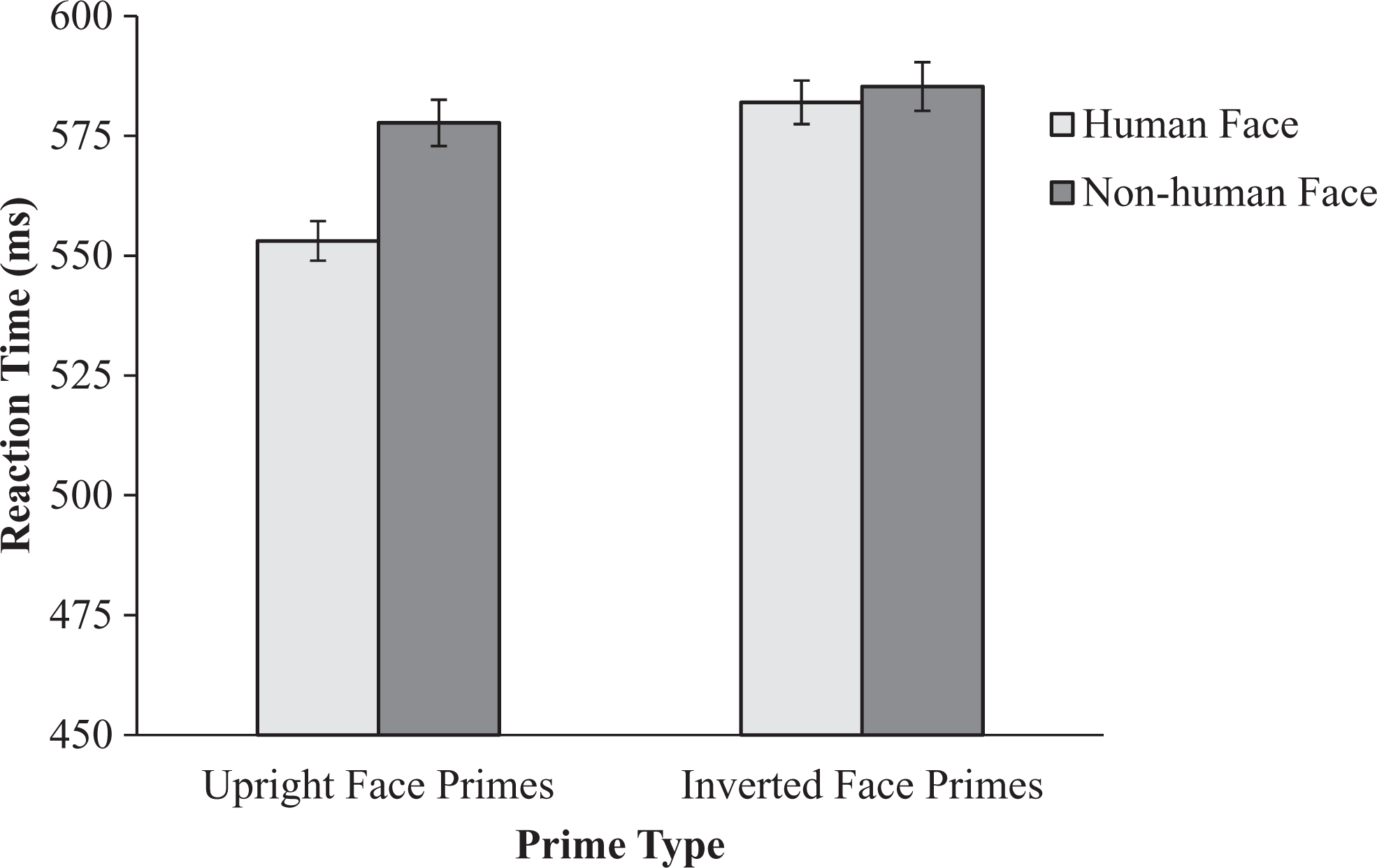
Lexical decision task (LDT) latencies for human words from Study 2. Upright human faces, but not upright doll faces, activated concepts related to humanness. Error bars represent standard error of the mean.
Next, we examined the interaction of prime orientation and prime type for machine words. Due to their similarity to humanoid robots, it may be intuitive to think that nonhuman faces might facilitate machine words. However, this analysis did not yield a significant interaction, F(1, 81) = 1.23, p = .27, ηp 2 = .02. Finally, we examined this interaction for nonwords. This analysis also yielded a nonsignificant interaction, F(1, 81) = .48, p = .49, ηp 2 = .01.
The results of Study 2 demonstrate that inverting faces inhibits the activation of concepts related to humanness, but only when primes were human faces. This effect was not observed for inverted compared to upright nonhuman faces. When primes were nonhuman or when the ability to process faces configurally was disrupted, concepts related to humanness were not activated. Moreover, words related to machines were not facilitated by any stimulus type or orientation, underscoring the unique link between perceptual similarities to humans and configural processing in ascribing humanlike minds, but not other types of minds.
General Discussion
Not all human-like stimuli possess a mind; therefore, parsing the human from the humanoid is an important act of social cognition. In the current research, we replicated past work demonstrating that both perceptual similarity to human faces (e.g., Looser & Wheatley, 2010) and configural face processing (e.g., Hugenberg et al., 2016) are important in generating judgments of whether targets appear to have a mind. However, we also demonstrated that these two characteristics interact in facilitating the activation of concepts related to humanness.
These findings have consequences for our understanding of a number of phenomena, ranging from predicting the uncanny valley to understanding autism. Research on the uncanny valley (Mori, 1970) has routinely demonstrated that as nonhuman entities become increasingly similar to humans they are perceived more favorably, up until a certain point at which they become a little too lifelike, and are perceived as unsettling or aversive. Indeed, the sense of unease we feel when viewing realistic simulations of humans (e.g., wax mannequins) can be attributed to the uncanny valley. One possibility that emerges from our data is that the sense of unease may be attributed to a confusion of the signals the brain is receiving. If, for example, a stimulus elicits configural processing, but has features signaling that it is nonhuman, the brain is receiving two potentially conflicting signals. These conflicting signals could explain the feeling of unease.
Autism’s theory of mind impairments are also linked to abnormal face processing. Indeed, people with autism typically struggle on tasks that involve face processing (Teunisse & de Gelder, 2003) and seem relatively less able to configurally process faces (Behrmann et al., 2006). To our knowledge, no work has investigated the extent to which people with autism are able to detect the objective signal of humanity in others, although Gray, Jenkins, Heberlein, and Wegner (2011) demonstrated that people scoring highly on a measure of autism-spectrum disorders tended to have reduced perceptions of adult human’s agency. New and colleagues (2010) have also demonstrated that people with autism do not show deficits in their sensitivity to animacy—they were equally able to detect the difference in scenes regardless of whether critical stimuli were alive. Perhaps the theory of mind deficits observed in autism may be more related to problems with configural face processing than with featural perceptions of animacy. This indicates it may be important to consider how both animacy and mind are jointly conveyed by faces for our understanding of autism. Nevertheless, considering the growing evidence suggesting that bottom-up perceptual processes are implicated in mind ascription, it may not be happenstance that individuals with chronic difficulties simulating the inner lives of others tend to process human faces like nonface objects.
Future work might explore the joint interaction of perceptual similarity and processing style on acts of dehumanization. For example, whereas people often actively minimize the humanness of outgroups (e.g., Castano & Giner-Sorolla, 2006) or minority groups (e.g., Goff, Eberhardt, Williams, & Jackson, 2008), the extent to which feature-based and configural processing effects act jointly to determine ascriptions of mind in the faces of outgroup members is not yet known. Similarly, the extent to which active motives for dehumanization can feed into how faces are processed is also poorly understood.
Finally, the current work used face inversion as its manipulation of configural processing. We chose this manipulation because it has been shown to reliably disrupt the ability to process stimuli configurally while retaining facial features and maintaining a sense of realism (i.e., inverted faces can be encountered in vivo). Nevertheless, it will be important in future work to extend these findings to other manipulations of configural processing (e.g., feature scrambling, composite face task). Thus, although we see the current results as clearly implicating the joint role of facial features and face processing in mind ascription, there are a number of important questions that remain unanswered.
Concluding Remarks
Whereas both featural and configural cues have been implicated in the perception of minds from faces, the current work helps to specify this face-to-mind cascade. Converging evidence using multiple methods demonstrates that ascribing humanlike minds to faces is bounded both by the features of the target and by processes employed by the perceiver.
Footnotes
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This research was supported by the NSF grant BCS-1423765.