
Abstract
Background
Recent advancements in cancer therapy still lack complementary treatments that substantially enhance the effectiveness of existing anti-cancer agents. A promising emerging strategy is the fine-tuning of nutrients, with a combinatory approach likely achieving superior results.
Objective
Here, the study aimed to evaluate the efficacy of a novel nutritional regimen that combines the ketogenic diet (KD) and high-dose vitamin C (VitC) as an adjuvant cancer therapy.
Methods
Syngrafts were established using triple-negative breast cancer EMT-6 cells in BALB/c mice. Five experimental groups were created: one received doxorubicin (DOX) alone, while the others received DOX with KD, high-dose VitC, or both. Treatments were administered over 14 days.
Results
Both KD and high-dose VitC individually enhanced the response to DOX, as reflected by favorable changes in cancer-associated markers, while their combination did not provide additional benefit. Specifically, KD positively influenced tumor proliferative signaling, angiogenesis, hypoxic response, anti-tumor immunity, and glucose metabolism, evidenced by reductions in insulin-like growth factor-1, vascular endothelial growth factor, hypoxia-inducible factor-1α, and programmed death-ligand 1, as well as decreased activity of pyruvate kinase and glyceraldehyde-3-phosphate dehydrogenase. High-dose VitC primarily targeted tumor replicative potential by reducing telomerase reverse transcriptase levels and may have inhibited metastasis by increasing tumor collagen content. However, neither the individual use of KD and high-dose VitC nor their combination enhanced DOX efficacy in reducing tumor volume or weight.
Conclusion
The combined nutritional approach, comprising KD and high-dose VitC, did not show significant advantages over the individual use of each in enhancing the tumor-inhibitory effects of DOX or modulating the tested cancer-associated markers. These findings highlight the need for further research to optimize dosing, treatment schedules, and mechanistic understanding of the combined therapy.
Introduction
Cancer is a leading cause of death worldwide. 1 Despite advances in treatment, conventional therapies such as chemotherapy and radiotherapy, as well as more recent approaches including targeted therapy and immunotherapy, remain limited by toxicity, treatment resistance, and modest long-term survival benefits in many solid tumors.2,3 Metastasis, responsible for about 90% of cancer-related deaths, 4 further underscores the urgent need for novel complementary strategies that selectively enhance therapeutic efficacy and overcome resistance without adding toxicity.
One emerging area of interest is nutritional modulation during cancer therapy, which aims to exploit tumor-specific metabolic dependencies. Cancer cells rely heavily on nutrients such as glucose, amino acids, and fatty acids for energy production, macromolecules biosynthesis, and redox balance. 5 Manipulating nutrient availability or exploiting nutrient-derived metabolites can disrupt these processes while potentially sparing normal cells. 6 Given the metabolic heterogeneity of cancer,7,8 combinations of nutritional interventions may offer greater benefits than single approaches by simultaneously targeting multiple hallmarks of cancer, including altered energetics, inflammation, angiogenesis, and immune evasion. 9
Within this context, the ketogenic diet (KD) and high-dose vitamin C (VitC) represent two promising strategies. The KD, a high-fat, low-carbohydrate diet, exerts antitumor effects by lowering systemic glucose, elevating ketone bodies (mainly β-hydroxybutyrate (BHB)), modulating oncogenic signaling pathways, triggering oxidative stress in cancer cells, and attenuating inflammation while supporting immune responses.10–22 High-dose VitC, on the other hand, selectively kills cancer cells through pro-oxidative mechanisms, generating reactive oxygen species and disrupting redox balance. 23 Additional mechanisms include modulation of the epigenome, oxygen-sensing, immunomodulatory functions, control of epithelial-to-mesenchymal transition, and regulation of kinase activity. 24
While both KD and VitC have individually demonstrated synergy with anticancer agents,24,25 their combined use has not yet been evaluated in a pre-clinical setting. Importantly, their mechanisms suggest strong potential complementarity. KD-induced reductions in circulating glucose not only deprive cancer cells of their primary fuel and lower nicotinamide adenine dinucleotide phosphate (NADPH)-dependent antioxidant defenses but also reduce competition at glucose transporters (GLUTs), thereby enhancing VitC uptake.26,27 This increased intracellular VitC, in the context of KD-induced metabolic stress, can amplify oxidative damage and overwhelm tumor redox defenses. Together, these interactions provide a compelling rationale for testing the KD–VitC combination.
Therefore, this study seeks to evaluate the efficacy of combining the KD with high-dose VitC as an adjuvant cancer therapy in an in vivo model of TNBC. We hypothesize that a combinatory approach is likely to result in improved efficacy of either nutritional intervention and a better synergy with cancer therapy.
Material and methods
Cell line and culture conditions
The murine mammary carcinoma cell line EMT-6 was used in this study, generously provided by Professor Wamidh Talib from Applied Science Private University, Amman, Jordan. This cell line exhibits characteristics similar to those of triple-negative human breast cancer and takes advantage of the complete mouse immune system. The cells were cultured in Minimum Essential Medium (EuroClone, Italy) supplemented with 10% fetal bovine serum (EuroClone, Italy), 1% L-glutamine (EuroClone, Italy), and 1% penicillin-streptomycin solution (Eurobio, France). The cultures were maintained at 37 °C in a humidified atmosphere with 5% CO2.
Establishment of the orthotopic breast cancer tumor
Female BALB/c mice aged 10–12 weeks (∼26 g) were housed under environmentally controlled conditions with a constant temperature (20–24 °C) and humidity (45–65%). Mice were adapted to a normal 12 h light/dark cycle and had free access to water and standard laboratory chow. For the syngeneic tumor model, 1 × 105 EMT-6 cells were resuspended in 50 μL of complete Minimum Essential Medium (EuroClone, Italy) and then subcutaneously injected into the fourth mammary fat pad of each mouse.
Mouse treatments
KD
The American Institute of Nutrition (AIN)-93 diet for laboratory rodents 28 served as the standard diet (StD). The KD was prepared by modifying this diet, substituting fat for carbohydrate on a calorie-for-calorie basis to achieve a ketogenic ratio of 4:1. The KD was formulated to deliver approximately 7 kcal/g of energy, consisting of 92% fat, 2% carbohydrate, and 6% protein. On the other hand, the StD was formulated to deliver approximately 3.9 kcal/g of energy, consisting of 9% fat, 76% carbohydrate, and 15% protein. The detailed composition of both diets is presented in Table 1.
Nutritional content of the diets.

StD: standard diet; KD: ketogenic diet.
StD was based on the composition of the American Institute of Nutrition (AIN)-93 M diet for laboratory rodents (26).
Modified from StD by substituting fat for carbohydrate on a calorie-for-calorie basis.
Ketogenic ratio = (0.9 F + 0.46 P):(C + 0.58 P + 0.1 F), where F is grams of fat; P is grams of protein, and C is grams of carbohydrate.
High-dose VitC
Sodium ascorbate was purchased from Sigma-Aldrich (Cat. #: A4034, Germany) and was dissolved in sterile saline. Stock solutions of 400 mg/mL were freshly prepared each time before use to deliver VitC at a dose of 4 g/kg. This high dose has been demonstrated to achieve a peak concentration as high as 30 mM in mouse peripheral blood after 90–180 min of intraperitoneal (IP) injection. 29 This millimolar concentration is known to be responsible for the pro-oxidant and cytotoxic effect of VitC toward cancer cells. 23
Chemotherapy (DOX)
DOX was obtained as a commercial solution in a 25-ml vial with a concentration of 2 mg/ml (EBEWE Pharma, Austria). To mimic oncologic treatment conditions in human patients, DOX was administered intraperitoneally in repeated cycles at three-day intervals, totaling three cycles at a dose of 5 mg/kg, resulting in a cumulative dose of 15 mg/kg/mouse.
Experimental design
A total of 45 mice were used in this study. The sample size was selected based on previously published EMT-6 syngeneic breast cancer studies, in which groups of 8–10 animals were sufficient to detect biologically meaningful differences in tumor growth. Although a formal a priori power calculation was not performed, our sample size is consistent with established practice in preclinical oncology research.
When the average tumor volume reached 341 mm3, mice were randomly divided into five experimental groups, each consisting of nine mice: (1) control group (CONT); (2) DOX group (DOXO); (3) KD group (KETO); (4) high-dose VitC group (HDVC); and (5) KD and high-dose VitC combined group (COMBO). Mice in the CONT, DOXO, and HDVC groups were fed a StD, while mice in the KETO and COMBO groups were fed a KD ad libitum. The KD regimen started two days before the first DOX injection.
Doxorubicin was administered at a dose of 2 mg/kg via IP injection in all groups except the CONT group. The treatment consisted of three cycles of DOX, with each cycle separated by three-day intervals. Mice in the HDVC and COMBO groups received three high-dose VitC injections in each DOX cycle: one injection simultaneously with DOX, one injection before, and one after. VitC was administered at a dose of 4 g/kg via IP injection. Sterile saline was used as a vehicle control for both DOX and high-dose VitC. See Figure 1 for a schematic representation of the experimental design.
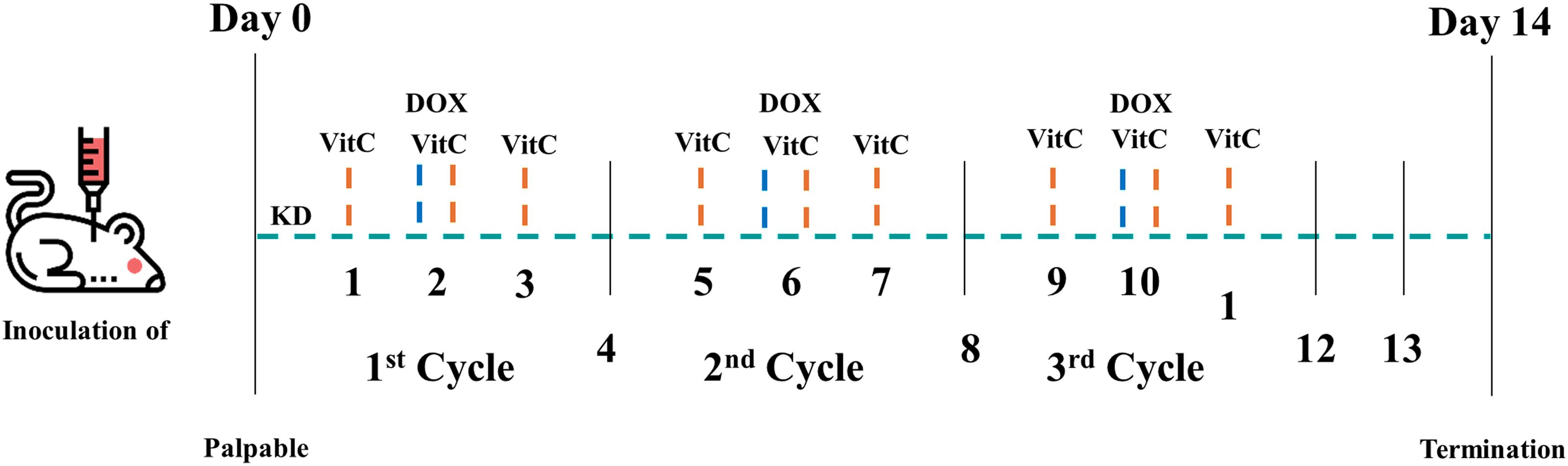
Basic schematic of the experimental design. DOX: doxorubicin; KD: ketogenic diet; VitC: vitamin C. This figure was created using images from Flaticon.com.
Tumor volume was measured every 3–4 days by a digital caliper and calculated using the following formula: tumor volume (mm3) = a × b2 × 0.5, where ‘a’ is the longest diameter, ‘b’ is the shortest diameter, and 0.5 is a constant to calculate the volume of an ellipsoid. Mice were monitored daily for pain and distress by observing changes in grooming, mobility, food intake, and body weight.
After the completion of a two-week treatment period, animals were killed with isoflurane. Blood was collected by cardiac puncture, and plasma samples were stored at −80 °C until analysis. The primary tumors were removed, rinsed with ice-cold phosphate-buffered saline (PBS), blotted, weighed, and cut in half. Subsequently, tumor tissues were either snap-frozen in liquid nitrogen or preserved in a 10% neutral buffered formalin solution for further analysis.
The study was not blinded during treatment administration or data analysis; however, all tumor measurements and subsequent analyses were conducted using standardized protocols to minimize potential bias.
Ketone bodies measurement
BHB levels were measured at baseline, prior to introducing the KD, and weekly throughout the study using the FreeStyle Optium Neo blood glucose and ketone monitoring system (Abbott Laboratories, USA) with compatible blood β-ketone test strips (Abbott Laboratories, USA). These test strips are designed to quantitatively measure blood BHB (mmol/L) from fresh whole blood samples.
Enzyme-linked immunosorbent assays (ELISAs)
Levels of hypoxia-inducible factor-1α (HIF-1α), vascular endothelial growth factor (VEGF), and cleaved caspase-3 in tumors were measured with the following ELISA kits according to the manufacturer's instructions: HIF-1α, cat# ab275103, Abcam, UK; VEGF, cat# ELM-VEGF-CL, RayBiotech, USA; and cleaved caspase-3, cat# PTE-CASP3-D175, RayBiotech, USA.
To measure total histone H3 acetylation in tumors, histone proteins were first extracted from the tumor tissues. Briefly, ∼50 mg of tumor samples were homogenized manually on ice in 500 µL of ice-cold lysis buffer (PBS plus 0.5% Triton X-100) supplemented with protease inhibitor cocktail (ReadyShield Protease Inhibitor Cocktail, cat# PIC0002, Sigma-Aldrich, Germany). The homogenates were then centrifuged at 10,000 rpm for 1 min, and the supernatants were discarded. The resulting pellets were resuspended in extraction buffer (0.5 N HCl plus 10% glycerol) and incubated on ice for 30 min, and then centrifuged at 12,000 rpm for 5 min. The supernatants were then transferred to new microcentrifuge tubes, and 150 µL of acetone were added per 50 mg of tissue and left overnight at −20 °C. Subsequently, the acetone mixture was centrifuged at 12,000 rpm for 5 min. The resulting pellets containing the histones were air-dried and dissolved in distilled water, forming the histone extracts. Total protein concentration was quantified using the BCA assay (Pierce BCA Protein Assay Kit, cat # 23227, Thermo Scientific, USA), and the total histone H3 acetylation was then analyzed with the Histone H3 Total Acetylation Detection Fast Kit (cat# ab115124, Abcam, UK) following the manufacturer's instructions.
To measure 5-hydroxymethylcytosine (5-hmC) in tumors, DNA was first extracted from the tumor tissues using the GeneJET Genomic DNA Purification Kit (cat# K0721, Thermo Scientific, USA). Subsequently, 100 ng of the DNA samples were used to perform the 5-hmC assay using the Quest 5-hmC DNA ELISA Kit (cat# D5425, Zymo Research, USA) following the manufacturer's instructions.
Plasma insulin-like growth factor-1 (IGF-1) concentrations were determined with the IGF1 Mouse ELISA Kit (cat# ab100695, Abcam, UK) following the manufacturer's instructions.
Colorimetric assays
Plasma glucose concentrations were determined by the glucose oxidase method using a commercial kit (GLUCOSE-LQ, cat# 41012, SPINREACT, Spain) following the manufacturer's instructions. In this method, glucose is oxidized by glucose oxidase, and the resulting hydrogen peroxide reacts with phenol and 4-aminophenazone in the presence of peroxidase to produce a red-violet dye. The intensity of the color is proportional to the glucose concentration and is measured spectrophotometrically.
Plasma ascorbic acid concentrations were determined using a commercial kit (EnzyChrom Ascorbic Acid Assay Kit, cat# EASC-100, BioAssay Systems, USA) following the manufacturer's instructions. In this assay, ascorbic acid is oxidized by ascorbate oxidase, resulting in the production of hydrogen peroxide, which reacts with a specific dye to form a pink-colored product. The color intensity is directly proportional to the ascorbic acid concentration in the sample.
Glyceraldehyde-3-phosphate dehydrogenase
Pyruvate kinase
Western blotting
Tumor tissues (∼20 mg) were homogenized and lysed in RIPA buffer containing protease and phosphatase inhibitors (cat# sc-24948A, Santa Cruz Biotechnology, USA). The lysates were centrifuged at 15,000 × g for 10 min, and protein concentration was determined using the DC-Protein Assay Kit (cat# 5000116, Bio-Rad, USA). Equal amounts of protein (40 µg) mixed with an equal volume of 4X Laemmli sample buffer were loaded onto a 7.5% SDS-PAGE gel, along with a molecular weight marker (cat# DM131-01, Biotrend, USA). After electrophoresis, proteins were transferred to a 0.45 µm-pore-sized nitrocellulose membrane (cat# 88018, Thermo Fisher Scientific, USA). Blots were blocked with 5% nonfat milk in Tris-buffered saline with 0.1% Tween 20 (TBST) for 2–4 h at room temperature. Subsequently, blots were probed with primary antibodies against nuclear factor-kappa B (NF-κB) (1:1000, cat#NBP2-67284, Novus Biologicals, USA), programmed cell death protein-1 (PD-L1) (1:500, cat# PA5-88105, Thermo Scientific, USA), telomerase reverse transcriptase (TERT) (1:500, cat# PA5-1116024, Thermo Scientific, USA), and β-Actin (1:200, cat# sc-47778, Santa Cruz Biotechnology, USA) for 16 h at 4 °C. Blots were washed with TBST and incubated with a goat anti-rabbit HRP-conjugated secondary antibody (1:5000, cat# ab97051, Abcam, UK) for 1 h at room temperature. The signal was developed using the western blotting luminol reagent for chemiluminescent HRP detection (cat# sc-2048, Santa Cruz Biotechnology, USA). Immunoreactive proteins were visualized using the ChemiDoc XRS
Histopathological analysis
After fixation in 10% neutral buffered formalin, tumor tissues underwent routine paraffin processing. The fixed tissues were then embedded in paraffin blocks, cut into 5 µm-thick sections, and stained with hematoxylin and eosin
Immunohistochemistry
For immunohistochemistry staining, tumor sections were prepared using the Rabbit HRP/DAB Detection IHC kit (cat# ab64261, Abcam, UK). Briefly, after routine paraffin processing and embedding, tumor tissues were cut into 5 μm-thick sections. Following dewaxing and rehydrating, sections were boiled in a sodium citrate buffer (10 mM sodium citrate, 0.05% Tween 20, pH 6.0) for 90 min at 85 °C in a water bath to retrieve antigens. Sections were then incubated with a hydrogen peroxide blocking buffer for 30 min to quench endogenous peroxidase. Next, tissues were permeabilized with PBS plus 0.3% Triton X-100 and blocked for non-specific binding with a protein block for 30 min. Anti-Ki-67 primary antibody (1:3000, cat# NB110-90592, Novus Biologicals, USA) was used to incubate the sections overnight at 4°C in a humidified chamber. Negative controls were run without primary antibodies. Afterwards, the sections were incubated with a biotinylated goat anti-rabbit secondary antibody for 30 min at room temperature, followed by streptavidin peroxidase for an additional 30 min. Signals were visualized using diaminobenzidine, and the nuclei were counter-stained with hematoxylin. Tissue sections were subsequently dehydrated, cleared, and mounted with DPX mounting media. Ki-67 staining in tumor tissues was manually quantified by determining the percentage of Ki-67-positive cells in three random high-power fields under a light microscope. Histopathology and immunohistochemistry slides were evaluated by two independent pathologists.
Statistical analysis
Data were analyzed using GraphPad Prism analytical software version 9.3.1 (GraphPad Software Inc., San Diego, CA, USA). Statistical significance was assessed by one-way analysis of variance (ANOVA) coupled with the least significant difference (LSD) post-hoc test for mean separation or the Kruskal-Wallis test. The results are expressed as the mean ± standard error of the mean (SEM)), and a P value < 0.05 was considered to indicate statistical significance.
Results
Body weight and energy intake
At the end of the study, body weight was significantly lower in all DOX-treated animals compared with their respective controls (Figure 2(a)). Additionally, the KD exacerbated the weight loss induced by DOX, as animals in both the KETO and COMBO groups displayed significantly lower body weight compared with those in the DOXO and HDVC groups (Figure 2(a)). Similar trends were observed in energy intake, which was reduced by DOX (Figure 2(b)), and this reduction was further intensified in the KETO and COMBO groups compared with the DOXO and HDVC groups (Figure 2(b)).
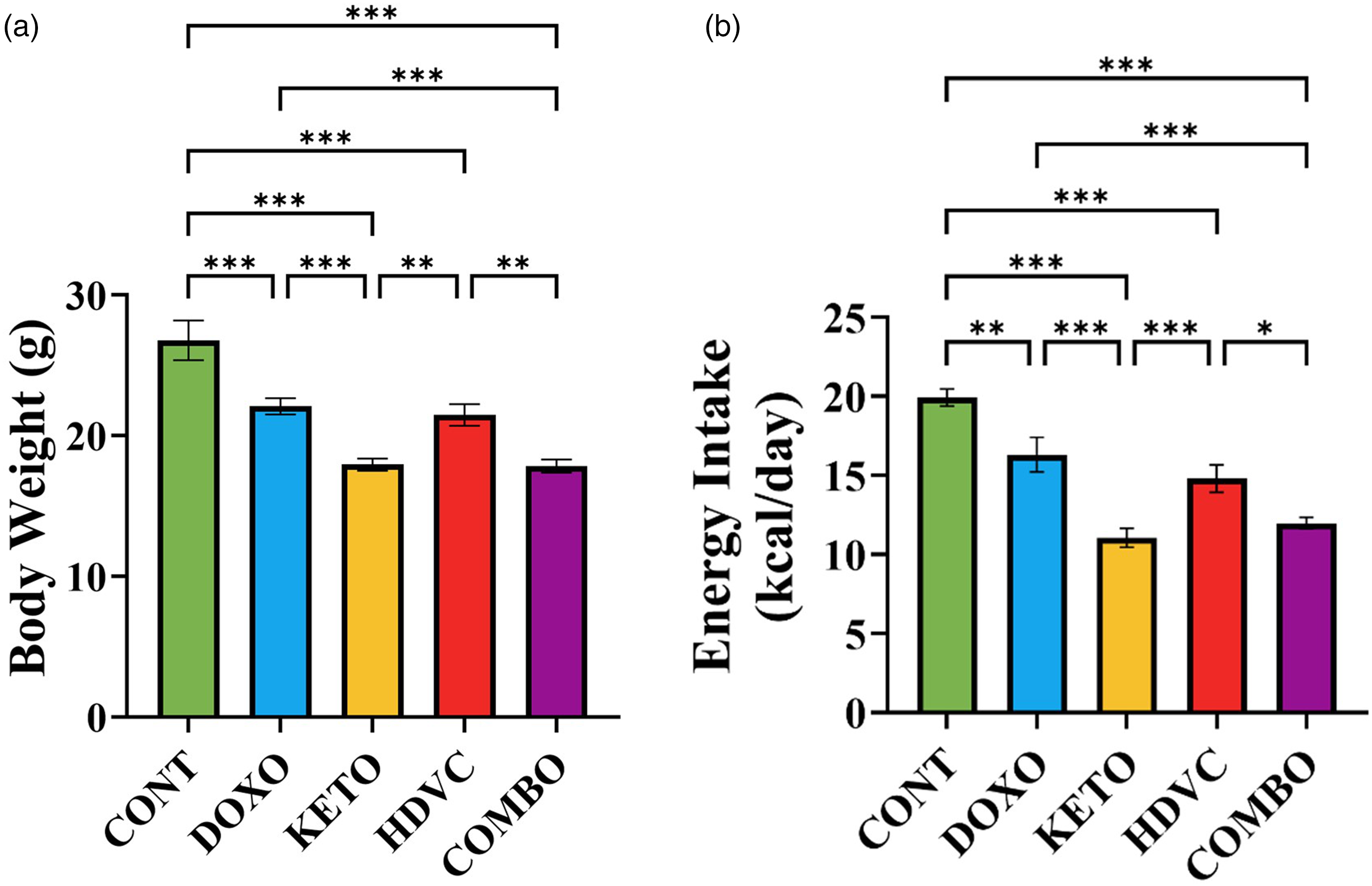
Effect of treatments used on body weight and energy intake of BALB/c mice bearing the EMT-6 TBNC tumors. (a) Body weight at the end of the study. (b) Average energy intake per day. Data were analyzed using one-way ANOVA followed by the LSD test (n = 9). * P ≤ 0.05, ** P ≤ 0.01, *** P ≤ 0.001. TNBC: triple-negative breast cancer.
Glucose and BHB levels
Glucose was measured once at the end of the study after a 2 h fasting. A highly significant difference in blood glucose levels was observed between groups consuming the KD and those consuming the StD (Figure 3). Notably, all DOX-treated groups, irrespective of the diet type (KD or StD), exhibited a significant drop in blood glucose levels compared with the controls (Figure 3).

Plasma levels of glucose at the end of the study. Data were analyzed using one-way ANOVA followed by the LSD test (n = 9). ** P ≤ 0.01, *** P ≤ 0.001.
Non-fasting BHB levels were repeatedly measured throughout the study. Mice in the groups consuming the KD achieved ketosis within 4 days after starting the diet, and their BHB levels remained continuously higher than those of the animals consuming the StD (Figure 4).
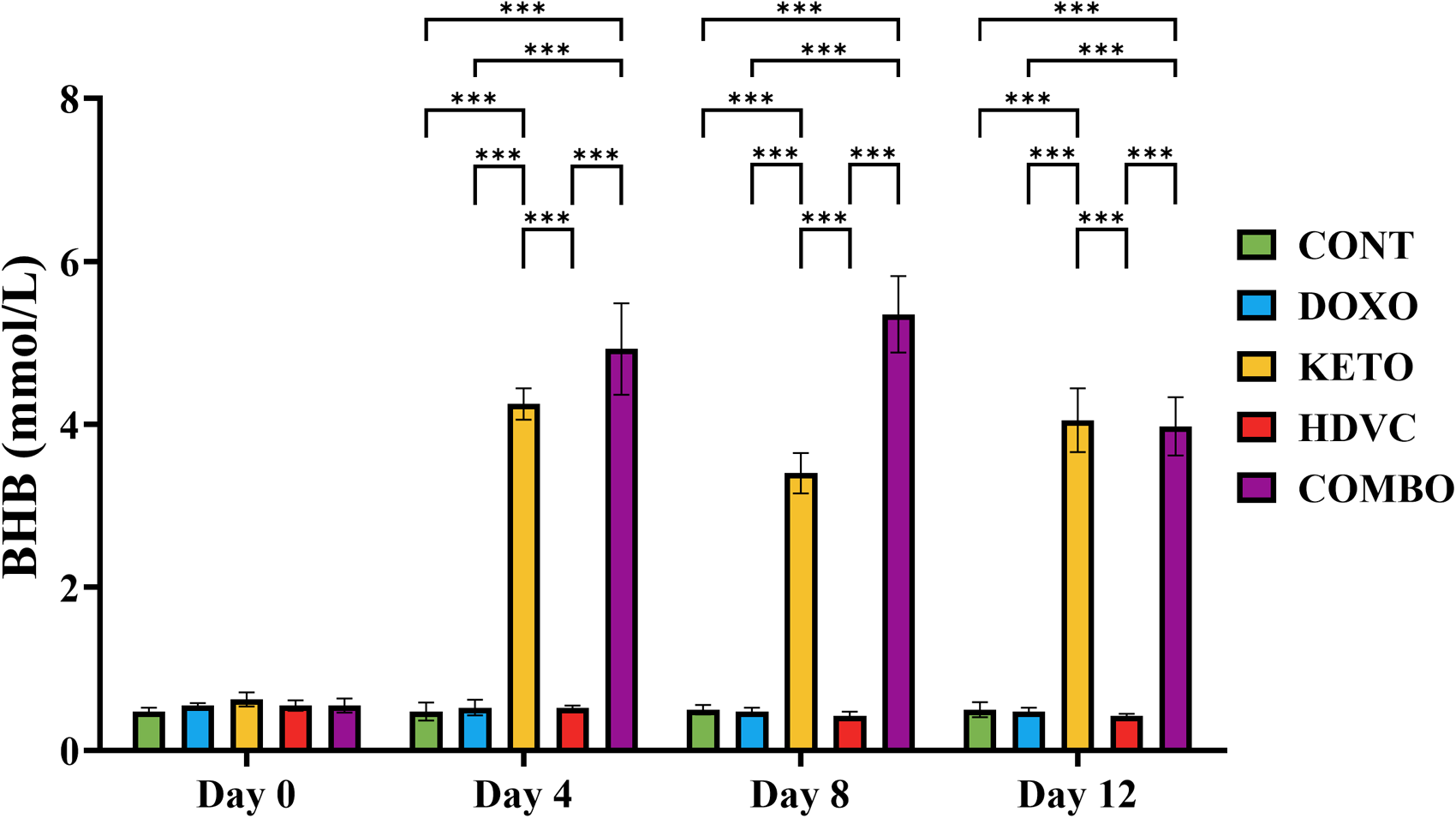
BHB levels at different time points of the study. BHB levels were measured twice a week. At baseline (day 0) before introducing the KD, BHB levels did not significantly differ between the experimental groups. Starting day 4 and throughout the study, BHB levels of the groups consuming the KD were significantly higher than those consuming the StD. Data were analyzed using one-way ANOVA followed by the LSD test (n = 4). ** P ≤ 0.01, *** P ≤ 0.001. BHB: β-hydroxybutyrate; KD: ketogenic diet; StD: standard diet.
Ascorbic acid levels
Ascorbic acid was assessed once at the end of the study, three days following the last VitC dose. The results showed that ascorbic acid levels were similar across all tumor-bearing animals, regardless of the treatment used (Figure 5). These levels are comparable with the physiological concentrations observed in non-tumor-bearing animals, 30 which indicates that neither the growing cancer, the use of DOX, nor the administration of high-dose VitC had an influence on the status of ascorbic acid in the tumor-bearing animals.

Plasma levels of ascorbic acid at the end of the study. There was no significant difference in ascorbic acid levels between the experimental groups assessed using one-way ANOVA (P > 0.05, n = 4).
Therapeutic efficacy against EMT-6 tumors in vivo
DOX treatment, whether administered alone or in conjunction with any of the nutritional adjuvants used, including the KD, high-dose VitC, or their combination, significantly reduced both tumor volume and weight in the EMT-6 tumor-bearing mice compared with the controls (Figure 6). Similarly, there was a notable improvement in the commonly used anti-tumor efficacy metrics, including the tumor growth inhibition (TGI) and the ratio of tumor volume in treated versus control mice (T/C), when DOX was administered either alone or in conjunction with any of the nutritional adjuvant treatments compared with the controls (Figure 7). However, the use of the KD, high-dose VitC, or their combination as nutritional adjuvants did not enhance DOX efficacy in reducing tumor volume or weight or in improving any of the anti-tumor efficacy metrics measured after three cycles of treatment (Figures 6 and 7).
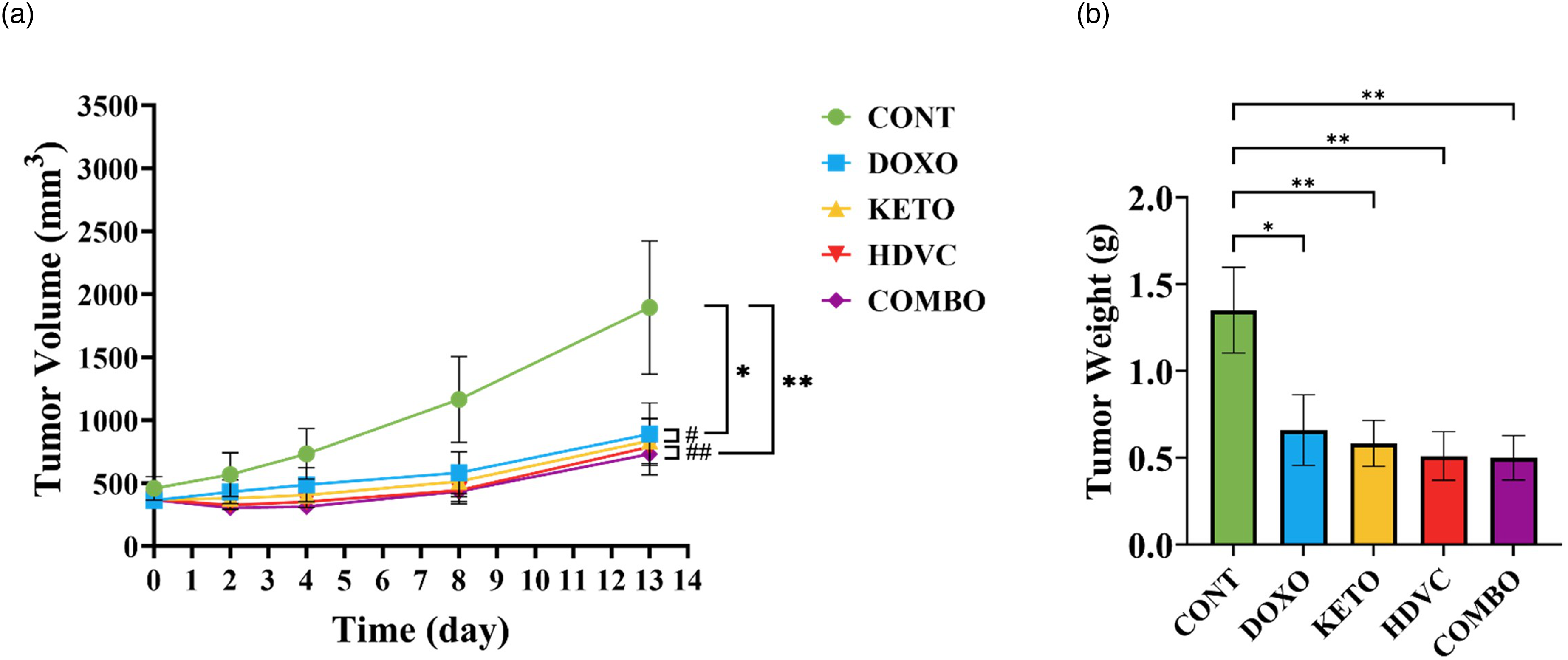
Tumor growth in vivo. (a) Average tumor volume throughout the study. (b) Average weight of excised tumors at termination. Data were analyzed using one-way ANOVA followed by the LSD test (n = 9). * P ≤ 0.05, ** P ≤ 0.01. # CONT vs DOXO/KETO, ## CONT vs HDVC/COMBO.
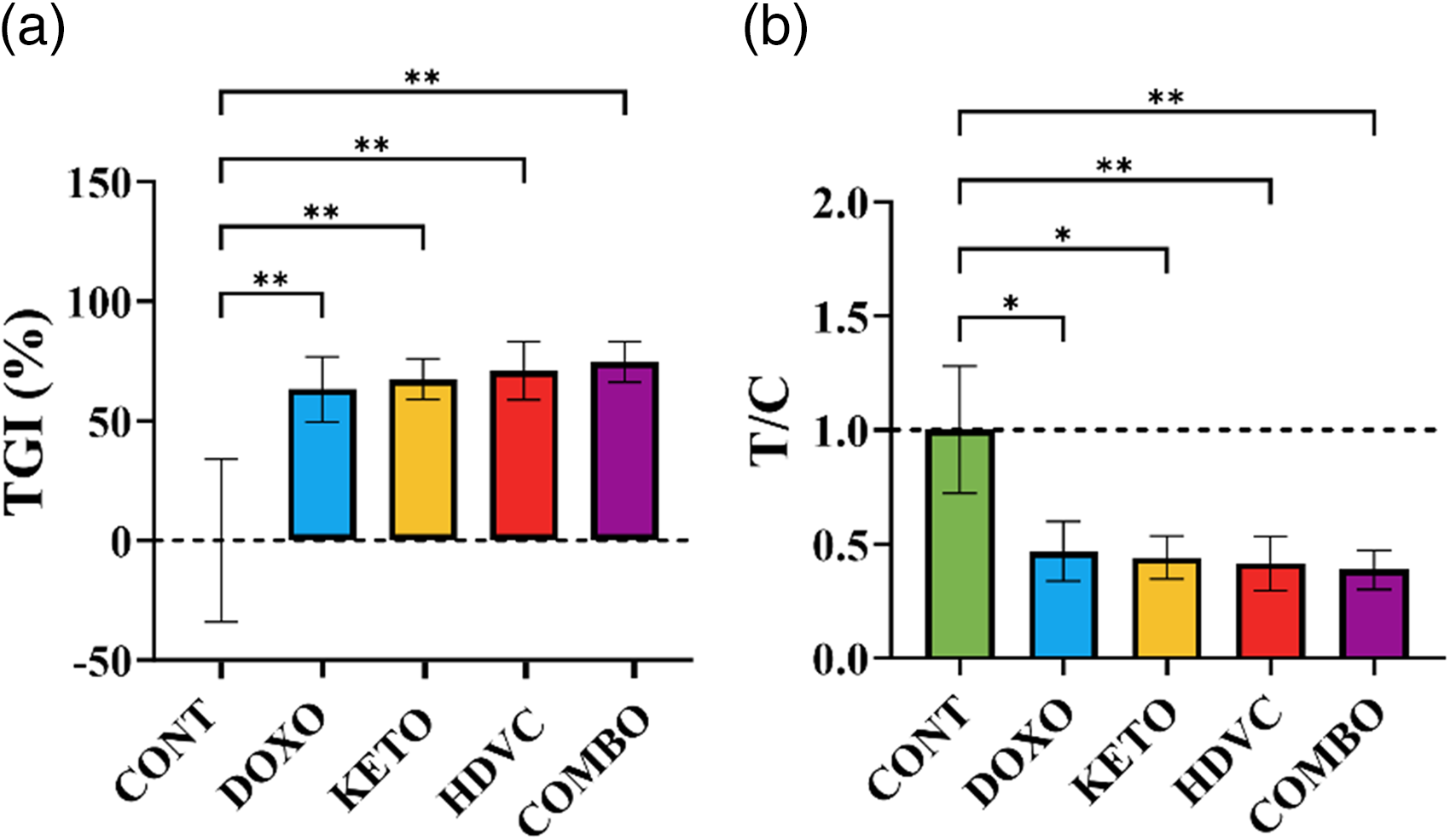
Anti-cancer activity scores in vivo. (a) Average TGI (%) at the end of the study (1 – (VT,e – VT,0) / (VC,e – VC,0) × 100) where VT,e and VT,0 represent the mean tumor volume of treatment group at the end of the study (e) and day 0 respectively, VC,e and VC,0 represent the mean tumor volume of the control group at the end of the study (e) and day 0 respectively. (b) T/C ratio at the end of the study (mean tumor volume in the treatment group/mean tumor volume in the control group at the end of the study). Data were analyzed using one-way ANOVA followed by the LSD test (n = 9). * P ≤ 0.05, ** P ≤ 0.01. T/C: the ratio between the tumor volume in the treatment group and in the control group
Ki-67 index
The proliferative activity of the EMT-6 tumor cells was assessed using the Ki-67 percentage score after exposure to different treatments (Figure 8). However, no significant differences were observed in the Ki-67 levels between any of the treatments used and the control. Additionally, the administration of the KD, high-dose VitC, or their combination as nutritional adjuvants did not exhibit an anti-proliferative effect on tumor cells when compared with DOX monotherapy.
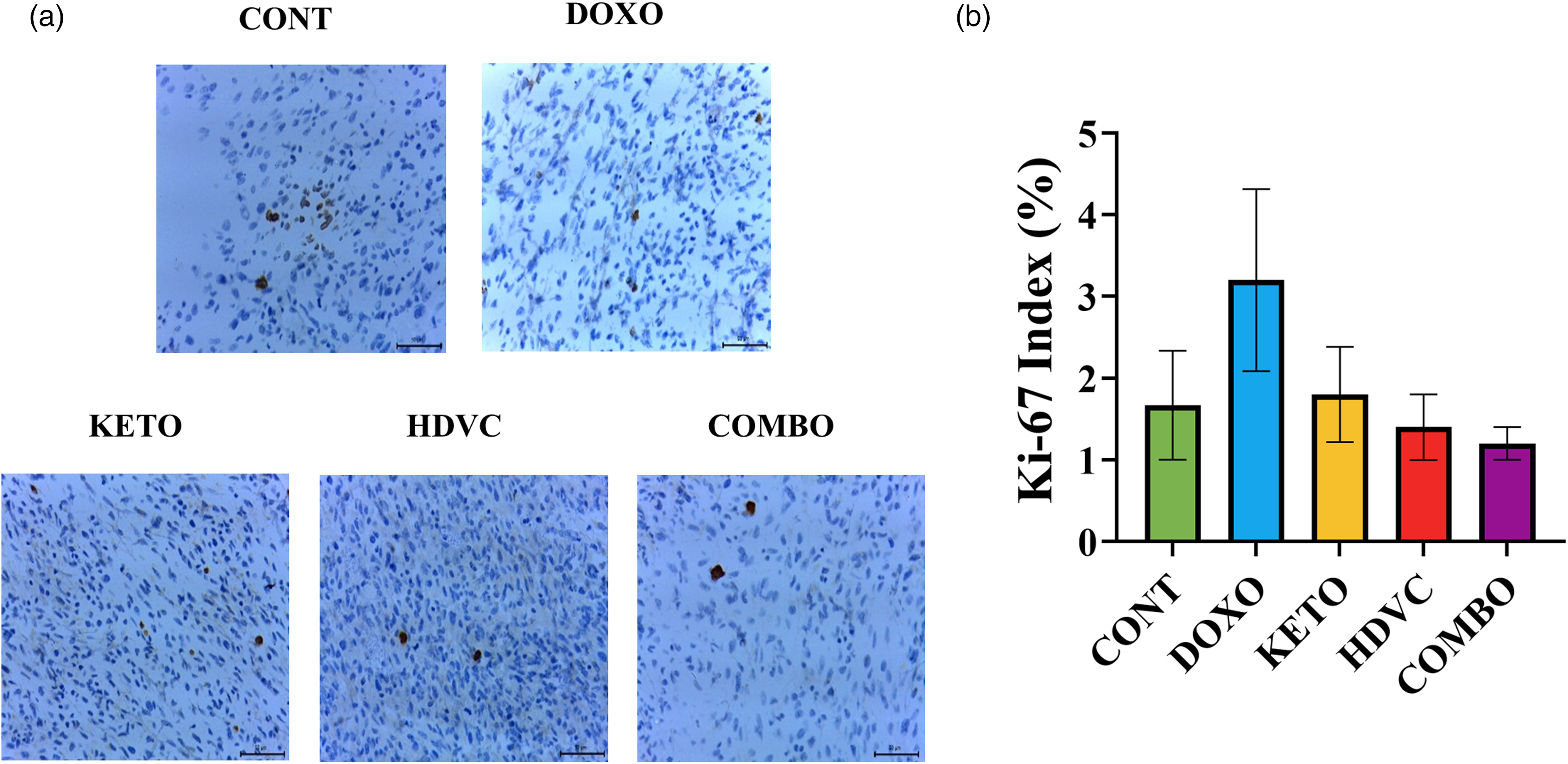
Immunohistochemistry staining of Ki-67 in TNBC tumors following treatment. (a) Representative images of Ki-67 immunostaining in tumor tissues. Nuclei stained brown are positive for Ki-67 (scale bar = 50 µm). (b) Quantification of Ki-67 in tumor tissues. Ki-67-positive cells were counted manually in 100 cells in three random high-power fields under a light microscope. Means were not significantly different (P > 0.05 using Kruskal-Wallis test, n = 3–5). TNBC: triple-negative breast cancer.
Cleaved caspase-3 levels
Apoptosis was detected and quantified by assessing the levels of cleaved caspase-3 in the EMT-6 tumor cells following treatment (Figure 9). Surprisingly, tumors from control animals showed significantly higher levels of cleaved caspase-3 in comparison with tumors from all DOX-treated animals. The administration of the KD, high-dose VitC, or a combination of both as nutritional adjuvants did not impact cleaved caspase-3 levels when compared with DOX treatment alone.
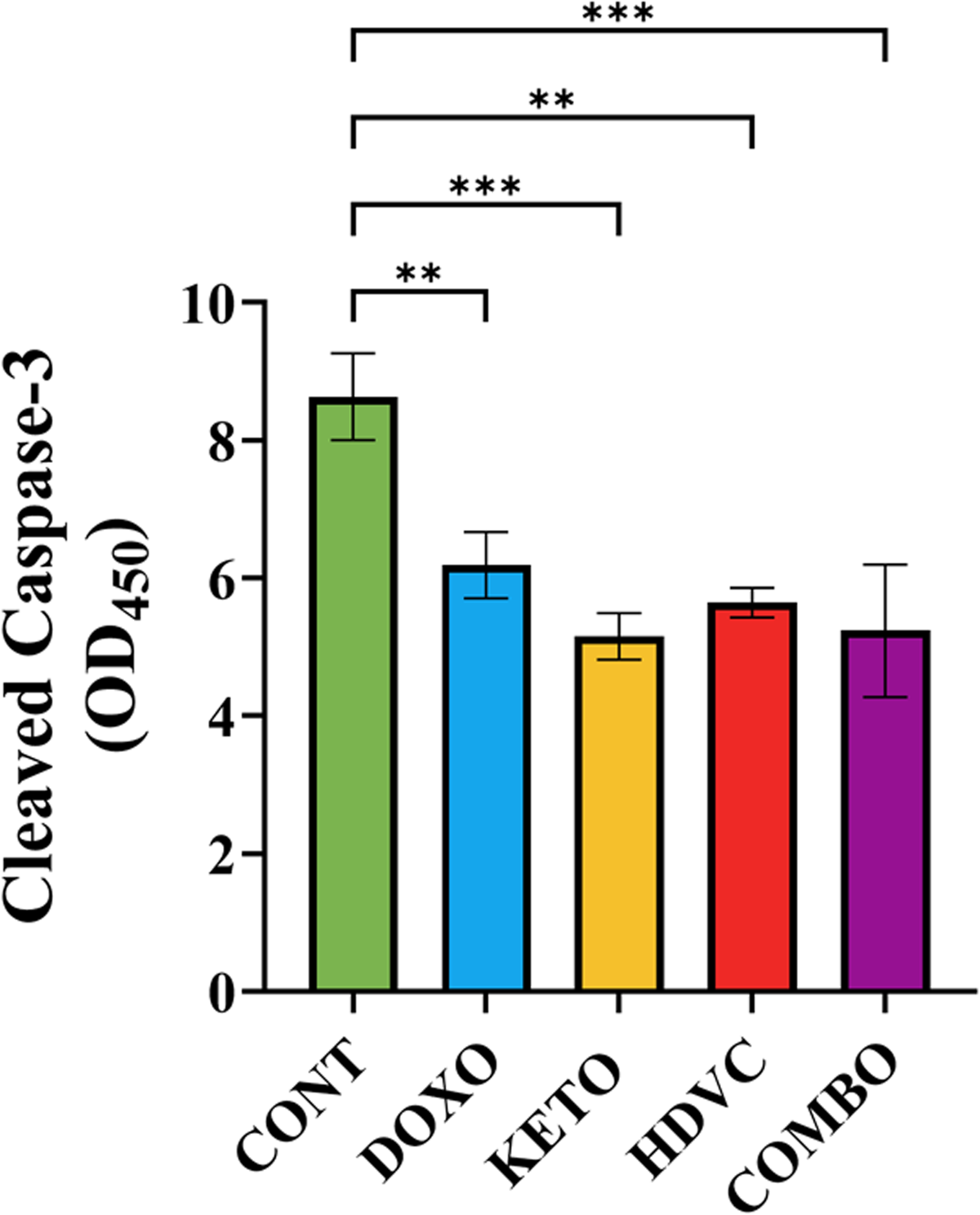
Levels of cleaved caspase-3 in TNBC tumors following treatment. Data were analyzed using one-way ANOVA followed by the LSD test (n = 6–7). ** P ≤ 0.01, *** P ≤ 0.001. TNBC: triple-negative breast cancer.
Histological examination of apoptosis and necrosis in EMT-6 tumors
Tumor sections were subjected to H&E staining and examined under a light microscope (Figure 10). Control tumors displayed minimal to no necrosis and apoptosis, whereas tumors from DOX-treated groups exhibited prominent necrotic regions with varying intensity, accompanied by reduced tumor cell density. Interestingly, necrosis occurred more frequently in tumors where the nutritional adjuvants, including the KD, high-dose VitC, or their combination were used, suggesting an enhancement of the necrotic action of DOX on the EMT-6 TNBC tumor cells.

Histological analysis of TNBC tumors subjected to different treatments. (a–e) Representative H&E staining of tumor sections, depicting (a) CONT group—no necrosis, (b) DOXO group—mild necrosis, (c) KETO group—moderate necrosis, (d) HDVC group—marked necrosis, and (e) COMBO group—mild necrosis. Necrosis levels were categorized as follows: Minimal (<10%), Mild (<30%), Moderate (30–50%), Marked (>50%). Arrows indicate regions of necrosis. For the HDVC group (d), no arrows were placed as necrosis was observed throughout the tissue section rather than localized to a specific region. Magnification = 400x, n = 5. H&E: hematoxylin and eosin; TNBC: triple-negative breast cancer.
TERT levels
TERT protein expression was evaluated using western blotting (Figure 11). The results indicated that DOX monotherapy did not induce a significant alteration in TERT levels. However, when high-dose VitC or its combination with KD were used as nutritional adjuvants along with DOX, a considerable reduction in TERT levels was observed compared with DOX monotherapy.
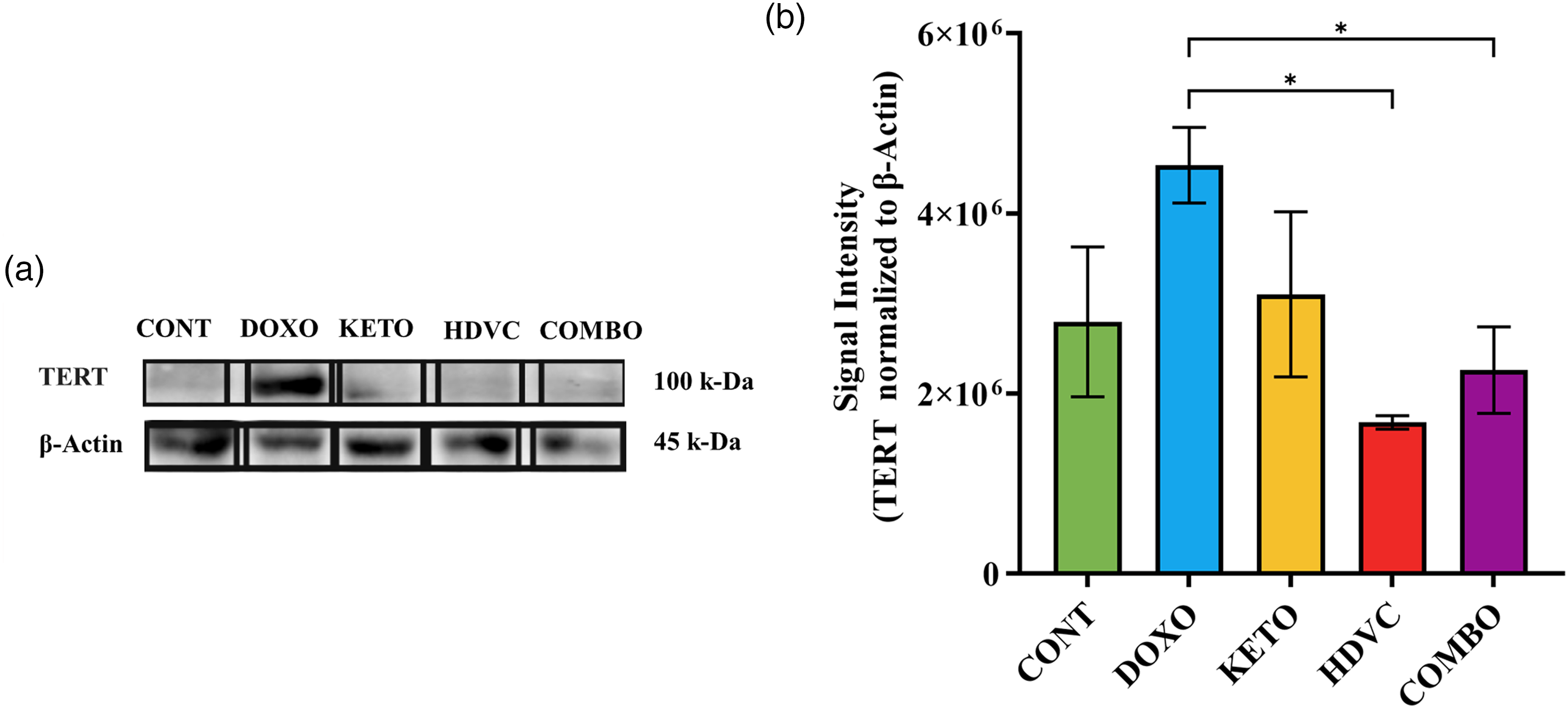
Levels of TERT in TNBC tumors subjected to different treatments. (a) Western blot bands were analyzed by densitometry, and the results were normalized to β-Actin. (b) Quantitative analysis of the western blot results. Data were analyzed using one-way ANOVA followed by the LSD test (n = 3–5). * P ≤ 0.05. TERT: telomerase reverse transcriptase; TNBC: triple-negative breast cancer.
IGF-1 levels
The levels of IGF-1 showed a significant decrease across all groups subjected to DOX treatment as opposed to the control (Figure 12). Furthermore, when the KD was administered as a nutritional adjuvant alongside DOX, either alone or in combination with high-dose VitC, there was a considerable drop (2.5-fold) in IGF-1 levels when compared with DOX monotherapy (Figure 12).
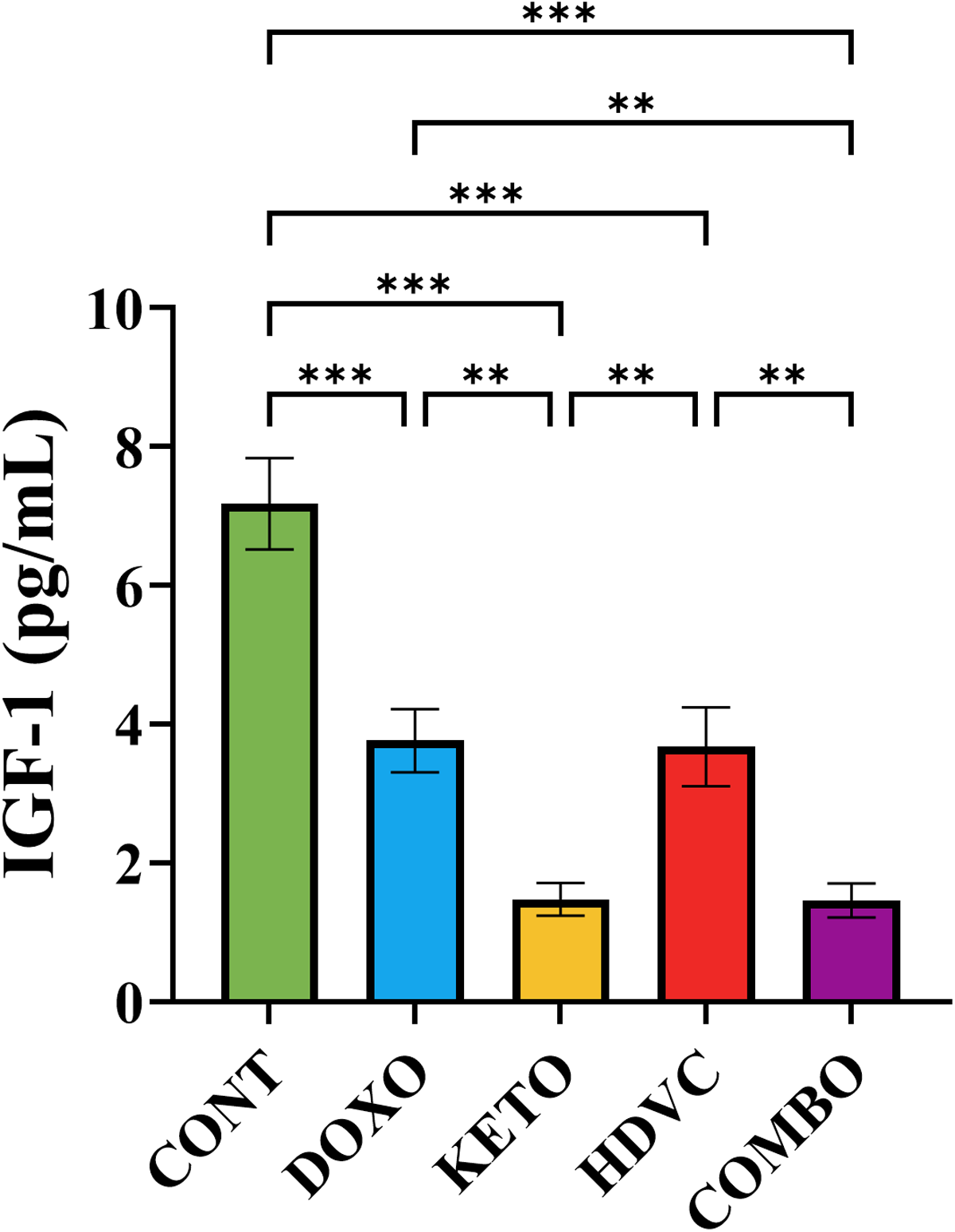
Plasma levels of IGF-1 following treatment. Data were analyzed using one-way ANOVA followed by the LSD test (n = 8). ** P ≤ 0.01, *** P ≤ 0.001. IGF-1: insulin-like growth factor-1.
HIF-1α levels
The levels of HIF-1α remained unchanged following DOX administration as a monotherapy (Figure 13). However, the introduction of the KD alongside DOX was the only nutritional adjuvant to elicit a prominent reduction in HIF-1α levels in comparison to the singular application of DOX (Figure 13). Intriguingly, even though combining high-dose VitC with DOX did not have any impact on HIF-1α levels compared to using DOX alone, it led to a noteworthy and statistically significant rise in HIF-1α levels when compared with the other nutritional adjuvants employed (Figure 13).
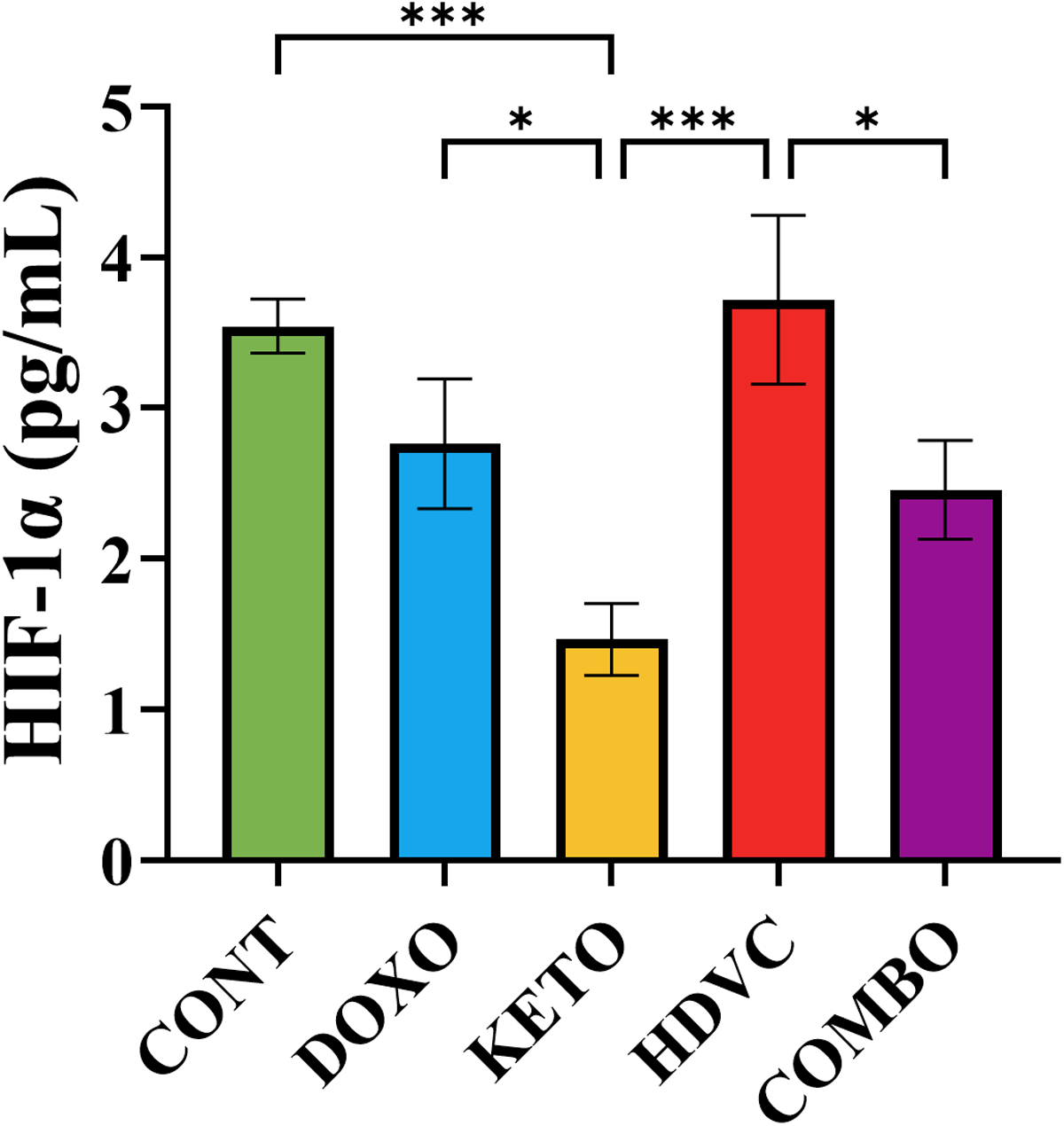
Levels of HIF-1α in TNBC tumors subjected to different treatments. Data were analyzed using one-way ANOVA followed by the LSD test (n = 5–6). * P ≤ 0.05, *** P ≤ 0.001. HIF-1α: hypoxia-inducible factor-1α levels; TNBC: triple-negative breast cancer.
VEGF levels
The use of DOX as a monotherapy exhibited no impact on VEGF levels (Figure 14). However, when the KD and its combination with high-dose VitC were used as nutritional adjuvants, they demonstrated a substantial decrease in VEGF levels compared with DOX monotherapy (Figure 14). The co-administration of high-dose VitC with DOX did not exert any influence on VEGF levels when compared to DOX monotherapy (Figure 14). Nevertheless, it did manifest a statistically significant elevation of VEGF levels when compared with the other nutritional adjuvants used.
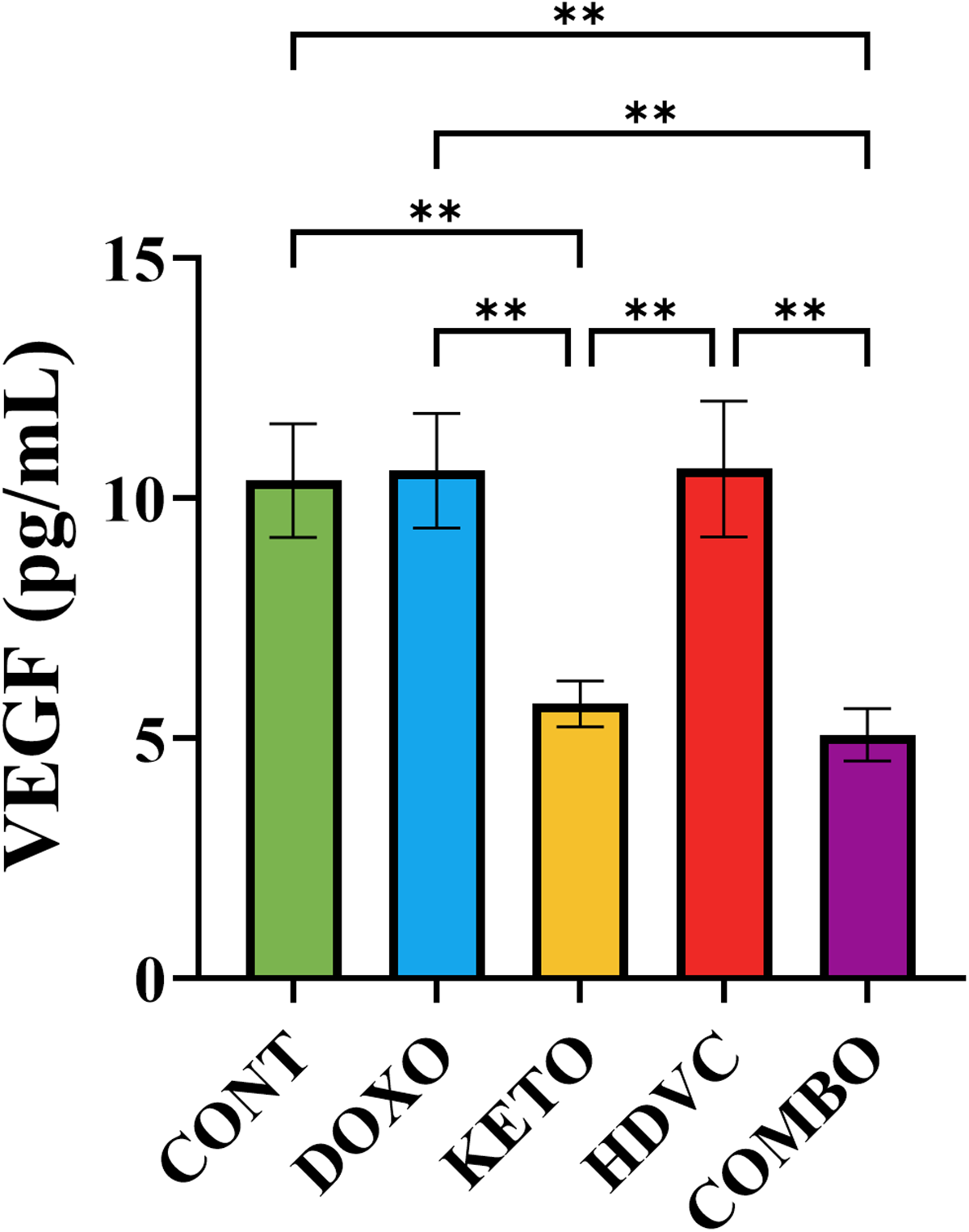
Levels of VEGF in TNBC tumors subjected to different treatments. Data were analyzed using one-way ANOVA followed by the LSD test (n = 5–7). ** P ≤ 0.01. VEGF: vascular endothelial growth factor.
Collagen content in tumors
Tumor sections underwent picrosirius red staining to facilitate the assessment of collagen accumulation (Figure 15). Tumors derived from mice treated with DOX as a single agent did not manifest a substantial change in collagen content relative to their controls. In contrast, the co-administration of high-dose VitC with DOX elicited a marked increase in tumor collagen content. This increase surpassed the levels observed with DOX monotherapy and exceeded the impact of the other nutritional adjuvants used.
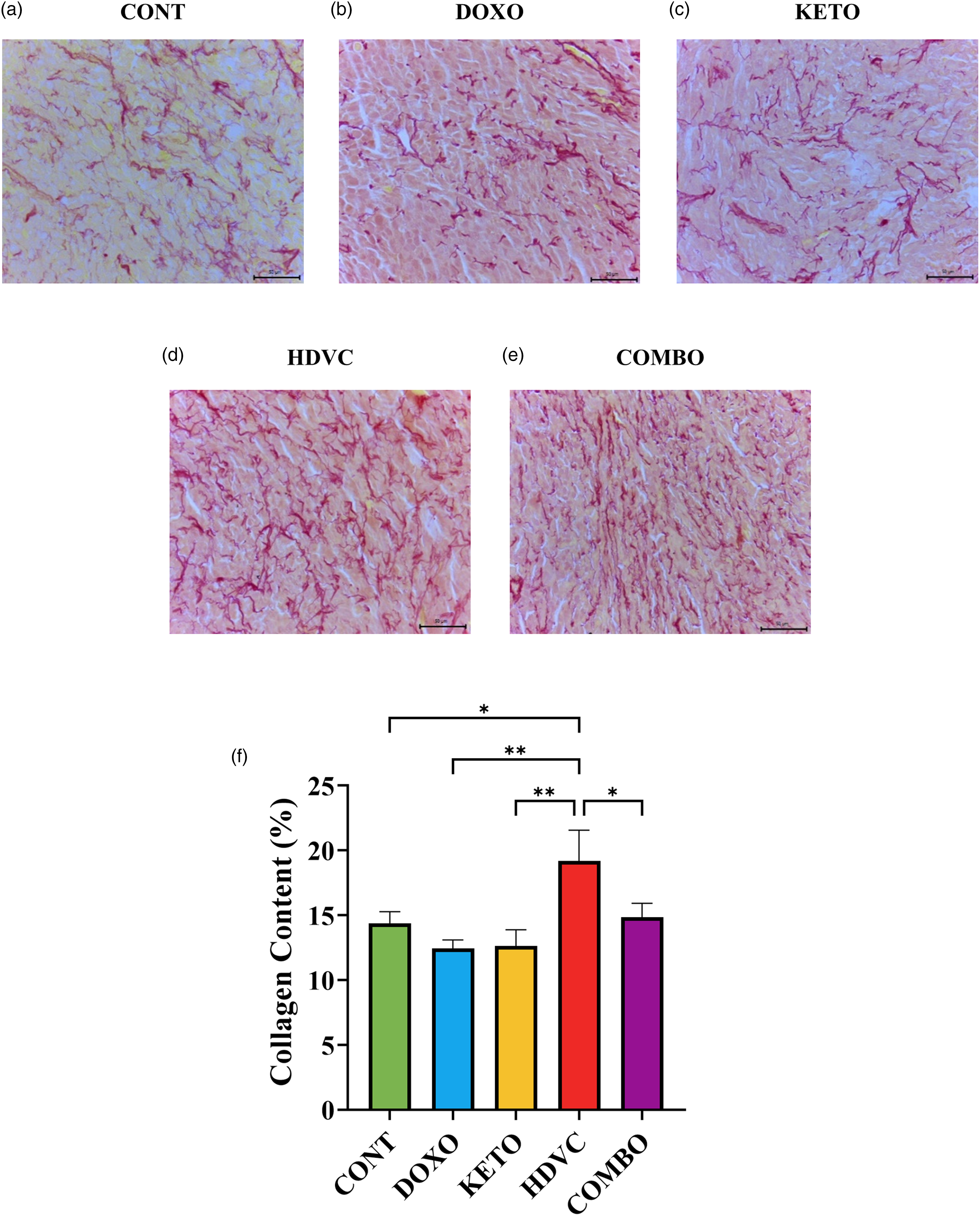
Collagen content of TNBC tumors subjected to different treatments. (a–e
NF-κB levels
NF-κB protein expression was assessed through western blotting (Figure 16). The use of DOX alone had no impact on NF-κB levels. Similarly, none of the nutritional adjuvants employed, including the KD, high-dose VitC, or their combination, induced any significant changes in NF-κB levels compared with DOX monotherapy.

Levels of NF-κB in TNBC tumors subjected to different treatments. (a) Western blot bands were analyzed by densitometry, and the results were normalized to β-Actin. (b) Quantitative analysis of the western blot results. Means were not significantly different (P > 0.05 using one-way ANOVA, n = 3–5). TNBC, triple-negative breast cancer.
PD-L1 levels
PD-L1 protein expression was evaluated via western blotting (Figure 17). The administration of DOX elicited a marked reduction in PD-L1 levels. Notably, the co-administration of the KD with DOX as a nutritional adjuvant, yielded a statistically significant and substantially more pronounced reduction in PD-L1 levels when compared with both DOX monotherapy and the other nutritional adjuvants employed.
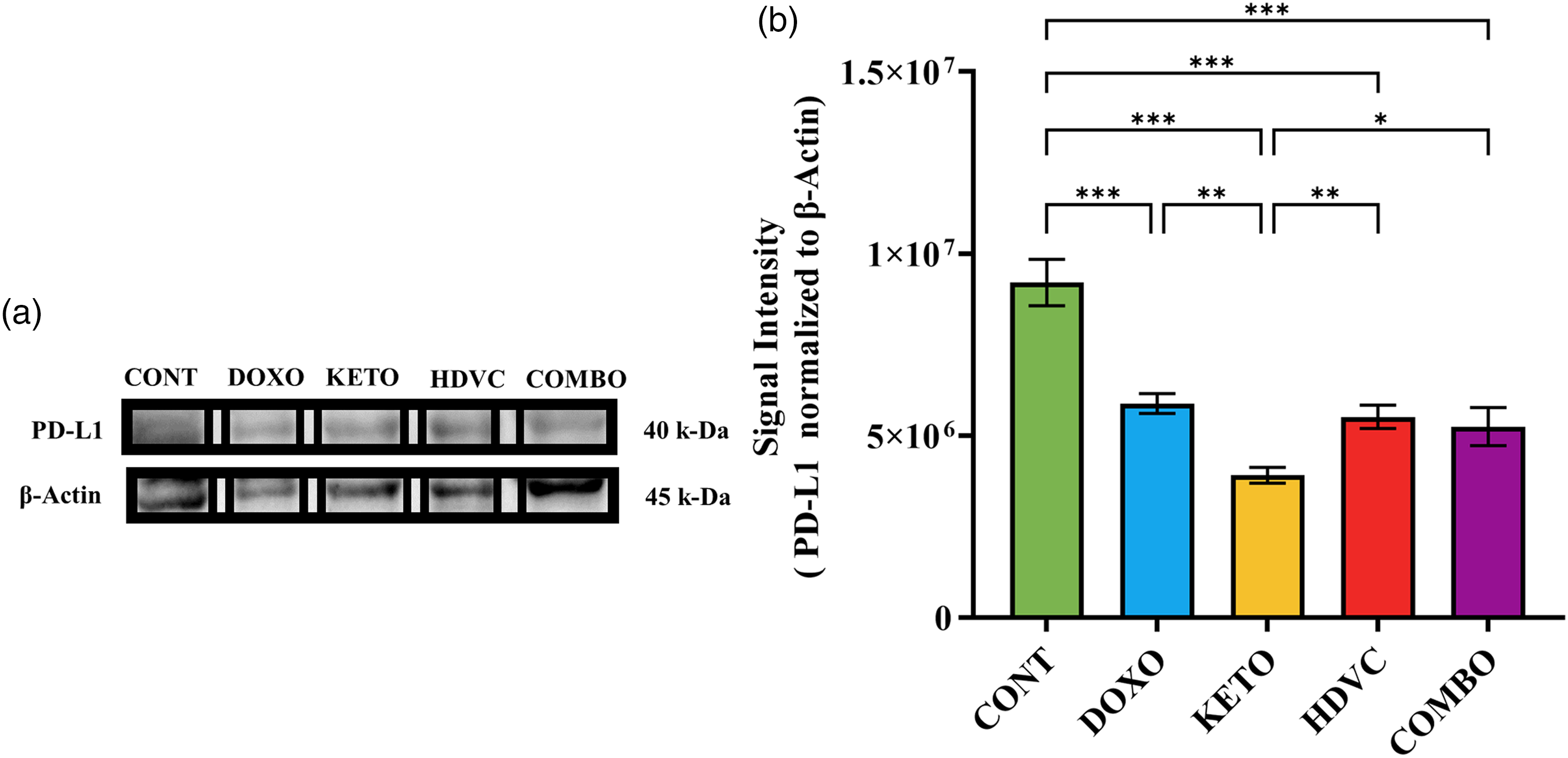
Levels of PD-L1 in TNBC tumors subjected to different treatments. (a) Western blot bands were analyzed by densitometry, and the results were normalized to β-Actin. (b) Quantitative analysis of the western blot results. Data were analyzed using one-way ANOVA followed by the LSD test (n = 2–5). * P ≤ 0.05, ** P ≤ 0.01, *** P ≤ 0.001. PD-L1: programmed death-ligand 1; TNBC: triple-negative breast cancer.
Glycolytic enzymes activity
The enzymatic activities of PK and GAPDH were quantified as illustrated in Figure 18. Following the administration of DOX, a pronounced decline in PK activity was observed, in contrast to GAPDH. When the KD was introduced as a nutritional adjuvant, either alone or in combination with high-dose VitC, it induced a greater reduction in both PK and GAPDH activity compared to DOX monotherapy. Conversely, the sole addition of high-dose VitC to DOX as a nutritional adjuvant did not impart any discernible modification to the activity levels of either enzyme in comparison to the use of DOX alone.
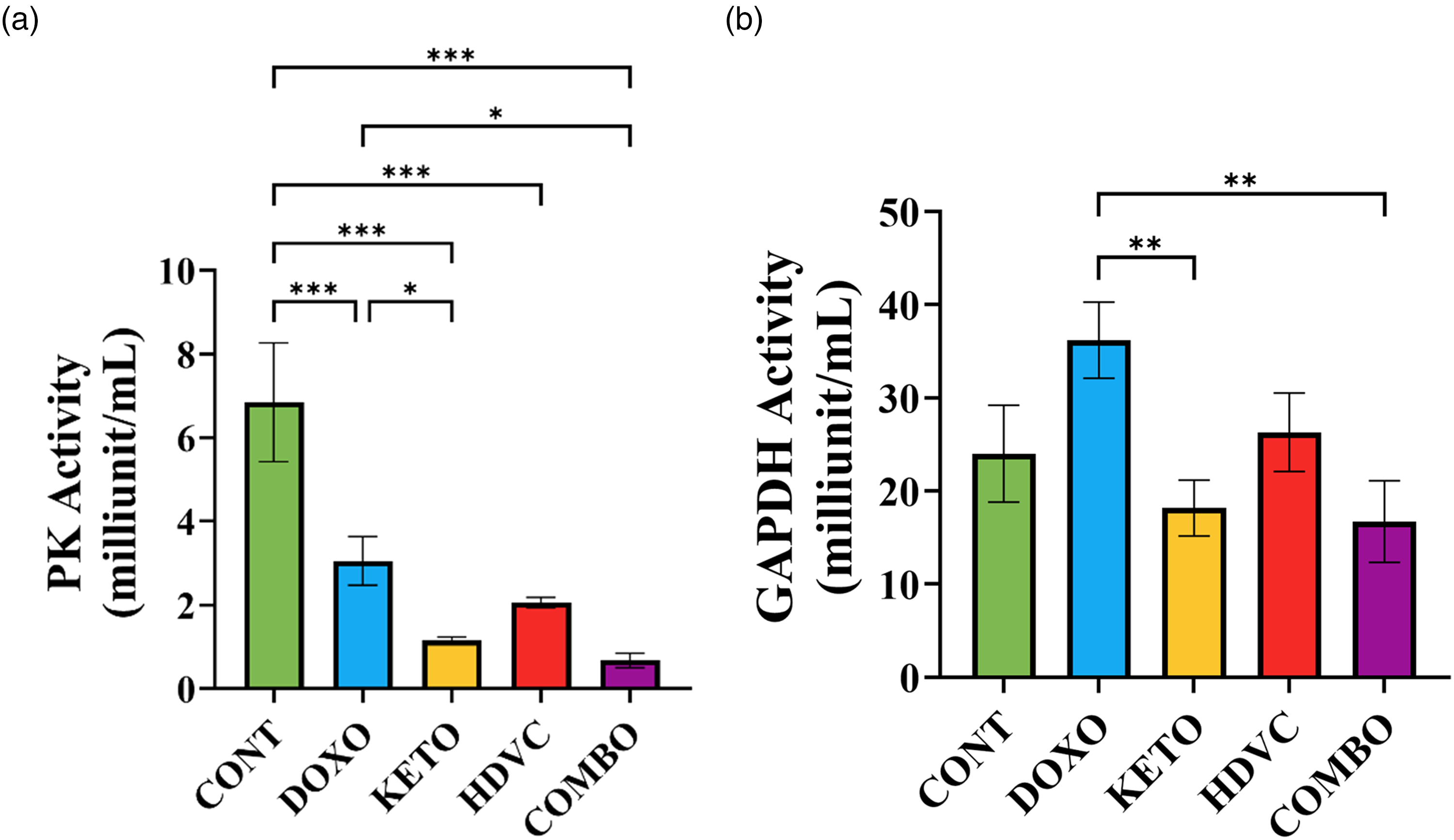
Pk and GAPDH activity in TNBC tumors subjected to different treatments
5-hmC levels
The presence of the 5-hmC modification in DNA was identified within tumors, as illustrated in Figure 19. The percentage of 5-hmC was consistent across all experimental groups, suggesting that neither DOX treatment nor the application of any of the nutritional adjuvants, including the KD, high-dose VitC, or their combination, had any influence on the global 5-hmC pattern.
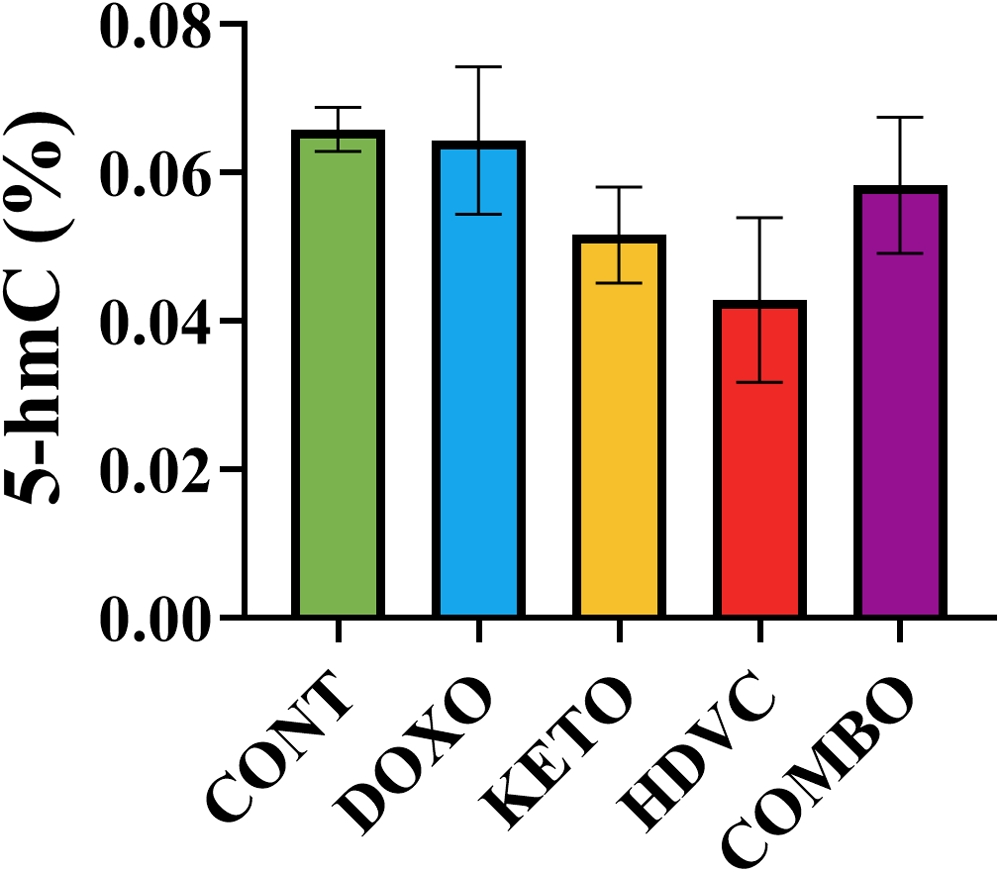
Levels of 5-hmC in TNBC tumors subjected to different treatments. Means were not significantly different (P > 0.05 using one-way ANOVA, n = 5–7). 5-mC: 5-hydroxymethylcytosine; TNBC: triple-negative breast cancer.
Total histone H3 acetylation
The variations in histone H3 acetylation profile were assessed within tumors, as illustrated in Figure 20. DOX administration as a single agent did not cause any significant alterations in the acetylation levels of histone H3 (Figure 20(a)). Moreover, the use of the KD, high-dose VitC, or a combination of both as nutritional adjuvants to DOX did not change how DOX affected histone H3 acetylation (Figure 20(b)).
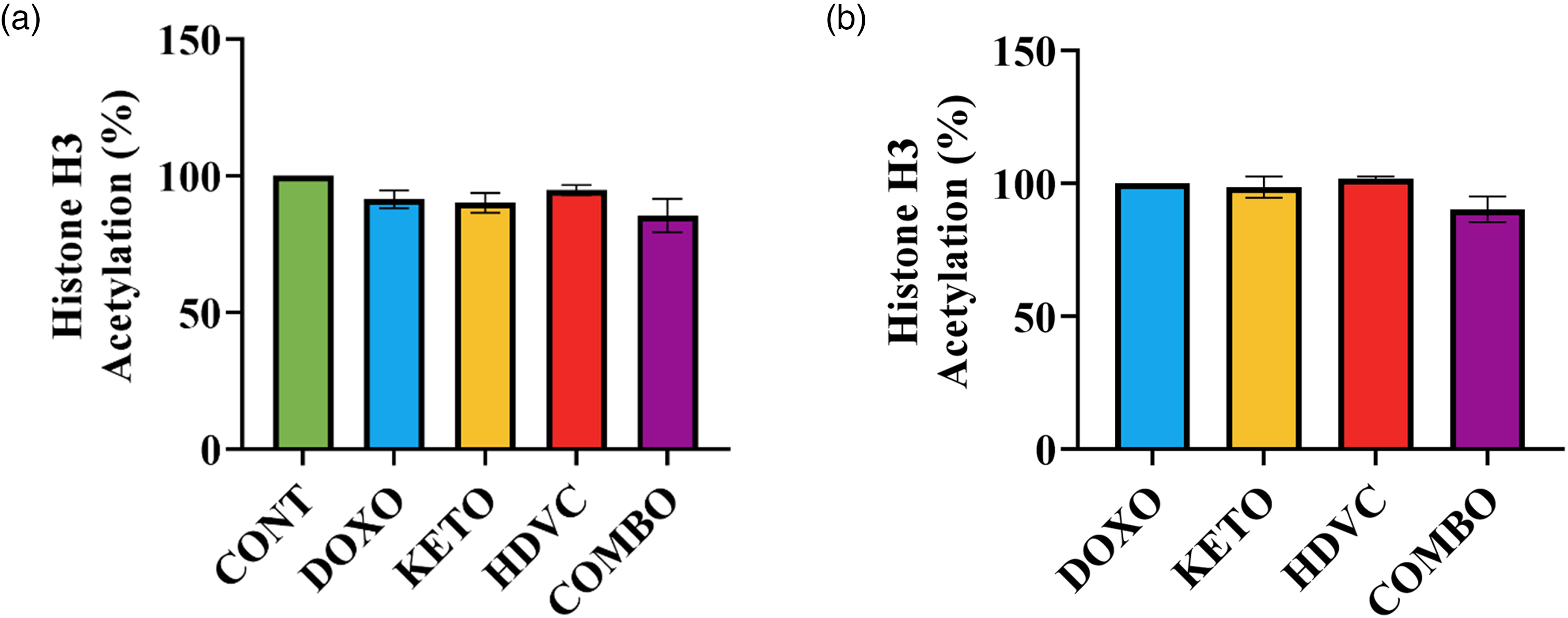
Levels of histone H3 acetylation in TNBC tumors subjected to different treatments. Levels of histone H3 acetylation did not differ significantly when compared with (a) CONT group (negative control) or (b) DOXO group (positive control) (P > 0.05 using one-way ANOVA, n = 5–6). TNBC: triple-negative breast cancer.
Discussion
In our study, we adopted a combined approach incorporating the KD and high-dose VitC as nutritional adjuvants to complement DOX, with the aim of augmenting the tumor-inhibiting action of this widely used chemotherapeutic drug. However, our results indicate that combining the KD and high-dose VitC with DOX did not lead to a further reduction in tumor growth compared with DOX alone or DOX combined with either KD or high-dose VitC.
These findings are contrary to our initial hypothesis, which proposed that a combination of nutritional interventions could yield superior therapeutic outcomes. We had anticipated that such a combined approach would be particularly effective by simultaneously targeting multiple processes and pathways involved in tumor proliferation and survival.
Despite this lack of a nutritionally enhanced tumor growth reduction, our findings indicate that the utilization of nutrition as an adjuvant cancer therapy has indeed facilitated an improved response to DOX. This improvement is evident through favorable changes detected in multiple markers linked to key cancer hallmarks. While these favorable changes did not result in a noteworthy decrease in the final tumor volume, they provide valuable insights into the intricate mechanisms governing tumor behavior and its response to therapeutic interventions. The final tumor volume remains the paramount therapeutic endpoint, but studying such markers is crucial for understanding the underlying biological processes.
Several of the examined cancer-associated markers showed positive responses to the KD, in contrast to the limited effect observed with high-dose VitC. This contradicts earlier findings that proposed potential benefits of high-dose VitC as an adjuvant cancer therapy. 24 This observation could also explain why implementing our integrated nutritional strategy did not provide notable benefits over the individual use of each nutritional adjuvant in enhancing the tumor-inhibitory effects of DOX or modulating these cancer-associated markers.
In fact, multiple factors, including treatment schedule, stability, and tumor characteristics, could have contributed to the observed discrepancy in VitC's impact in our study compared to previous reports.31,32 An essential aspect lies in VitC's short half-life of < 2 h in circulation, resulting from rapid renal clearance. 33 To maintain VitC levels within the tumor, daily administration has been reported as the most effective schedule. Many studies have shown that daily or twice-daily administration is necessary for enhanced co-treatment efficacy. 24 Despite this, we theorized that co-administering VitC with the KD, another proposed anti-cancer strategy, might increase cancer cell sensitivity to VitC, especially under the low plasma glucose levels caused by the KD. This reduction in glucose levels would result in less competition at the GLUTs, making cancer cells more susceptible to VitC. Our aim was to provide a treatment schedule (three days a week) that would be less burdensome for patients compared to daily administration.
Additionally, it has been demonstrated that sodium-dependent vitamin C transporter 2 (SVCT2) levels differ significantly among different breast cancer cell lines and are predictive of ascorbate uptake and cellular sensitivity to ascorbate cytotoxicity. 34 Unfortunately, we lack data regarding SVCT2 expression in the EMT-6 cells used in our study, which is an important factor to consider for assessing VitC efficacy.
Furthermore, differences in experimental models, including the choice of cancer cell lines, tumor type, and in vivo versus in vitro systems, can significantly influence responses to VitC. 24 Variations in dosage, treatment frequency, and duration further contribute to inconsistent outcomes. 24 Finally, metabolic context, including glucose availability, redox status, and antioxidant capacity, may modulate tumor response. 24
Considering these variables helps contextualize our findings, underscores the complexity of translating VitC effects across studies, and highlights the need for careful optimization of dosing regimens and model selection when evaluating VitC as a potential adjuvant therapy.
In our study, the only notable effect of high-dose VitC was seen in the positive modulation of TERT and collagen levels. This suggests potential anti-replicative and anti-metastatic properties for high-dose VitC. Despite these effects on tumor biology, high-dose VitC did not produce a measurable improvement in tumor volume reduction when combined with DOX.
It is well known that telomere maintenance is a common trait in cancer cells, enabling indefinite replication. 35 Targeting telomerase holds promise as an anti-cancer strategy due to its role in cancer cell replication, notably through TERT reactivation. Previous research aligns with our findings, showing that even low-dose VitC can reduce telomerase activity in cancer cells. 36 VitC may affect telomerase activity through various mechanisms, including provoking ROS production and reducing HIF-1α levels.37,38 However, our study's results do not support a reduction in HIF-1α levels as a primary mechanism for telomerase inhibition by high-dose VitC. Epigenetic changes, such as DNA methylation, also influence TERT expression and telomerase activity. 39 Some studies have shown that VitC can promote DNA demethylation by increasing ten-eleven translocation (TET)-mediated oxidation of 5-methylcytosine (5 mC) to 5 hmC. 40 While we didn't directly measure TERT promoter hypermethylation, our evaluation of global 5hmC levels found no significant impact from our treatments, including high-dose VitC.
Our study was designed to assess the collagen content of tumors as a biomarker for metastatic potential. Despite ongoing debate regarding the impact of increased collagen content on tumor progression, 41 several investigations have indicated an inverse relationship between elevated collagen deposition within the tumor stroma and reduced tumor invasion and metastasis.42–44 This phenomenon is attributed to the role of collagen as a physical barrier impeding the migratory capacity of cancer cells. 45
VitC functions as a critical cofactor for collagen prolyl and lysyl hydroxylases, facilitating the hydroxylation process at proline and lysine residues necessary for the formation of the correct triple-helical conformation of collagen. 46 Our investigation revealed a notable increase in tumor collagen levels subsequent to VitC administration, suggesting a potential anti-metastatic effect of high-dose VitC. The concept of leveraging VitC to counteract diminished collagen synthesis as a strategy to mitigate metastatic susceptibility was initially proposed decades ago and has since been extended in later research. 24
With regard to the KD, numerous investigations have highlighted the diet's efficacy in increasing the sensitivity of cancer cells to conventional chemotherapy or radiotherapy,18,47 as well as to targeted therapy 48 and immunotherapy. 49 However, certain studies have noted instances of either promoting tumor growth or inducing severe side effects in particular cancer models. 10
Our study showed that the KD demonstrated positive outcomes concerning many cancer-associated markers related to sustaining proliferative signaling, hypoxic response, inducing vasculature, avoiding immune destruction, and deregulating cellular energetics. Importantly, however, these molecular and microenvironmental changes did not translate into a further reduction in tumor volume beyond that achieved by DOX alone.
For example, we observed a profound reduction in plasma IGF-1 levels following the KD. Signal transduction through IGF-1/insulin-like growth factor-1 receptor (IGF-1R) can trigger various cellular effects, including promoting cellular growth, proliferation, survival, and resistance to therapy. 50 The KD might lower IGF-1 levels by inducing loss of appetite, reduced energy intake, and weight loss, as evidenced in our study.51,52 Additionally, the typical low-carbohydrate and low-protein content of the KD contribute to its effect on lowering IGF-1 levels. The KD used in this study had very low carbohydrate and protein content (2% and 5% of total energy, respectively). It is important to note that studies employing the KD use variable protein content, leading to variations in fat/protein ratios, potentially influencing the induction of ketosis and alterations in IGF-1 levels. Research has actually indicated a positive correlation between protein intake and serum IGF-1 levels. 53 Furthermore, the low blood glucose and insulin concentrations resulting from a low-carbohydrate diet, as observed in this study, contribute to the decrease in IGF-1 levels. 54 Evidence also shows an inverse relationship between BHB and IGF-1 concentrations, 10 all of which contribute to the KD's ability to lower IGF-1 levels and its downstream tumor-promoting pathways.
Our investigation revealed a reduction in HIF-1α levels solely with the KD treatment, whereas DOX monotherapy exhibited negligible impact, implying that a combined approach may confer superior efficacy compared to DOX alone. HIF-1α is the key transcriptional regulator of the hypoxic response. 55 In recent years, there has been growing interest in targeting HIF-1α as an innovative approach for cancer therapy. Multiple mechanisms underlie KD's modulation of the hypoxic response. Primarily, KD induces significant decreases in insulin and IGF-1 concentrations, as noted previously, potentially impeding HIF-1α upregulation via inhibition of the phosphatidylinositol 3-kinase (PI3K)/protein kinase B (AKT)/mammalian target of rapamycin (mTOR) pathway. 56 Clearly, our study demonstrated KD's pronounced efficacy in lowering IGF-1 levels, offering partial elucidation for its impact on HIF-1α. Additionally, KD induces a metabolic shift, notably through glucose deprivation, resulting in an anti-glycolytic, anti-Warburg effect. 10 This attenuation of the glycolytic pathway could perturb HIF-1α activation, given the critical role of glycolysis in sustaining HIF-1α activity. 57 Our findings indicate reduced glucose availability and decreased activity of key glycolytic enzymes, PK and GAPDH, following KD, which may contribute to its observed influence on HIF-1α. Furthermore, BHB, the main ketone body found in the circulation after KD, is an endogenous histone deacetylase (HDAC) inhibitor. 58 Prior research has demonstrated HDAC inhibitors’ capacity to suppress HIF function by diminishing HIF-1α levels or repressing its transactivation activity. 59 However, our study did not observe elevated histone H3 acetylation following KD, suggesting that HDAC inhibition may not be the primary mechanism underlying KD's beneficial effect on the hypoxic response.
Our investigation uncovered notable anti-angiogenic effect associated with the KD, as evidenced by a significant reduction in VEGF levels subsequent to KD administration. Angiogenesis plays a critical role in tumor progression, and therapies targeting this process often focus on inhibiting VEGF, given its central role in promoting angiogenesis. 60 Tumor cells can upregulate VEGF expression through various stimuli, including cytokines, growth factors, hypoxia, as well as through oncogene activation and tumor suppressor mutation. 61
Our findings suggest that the observed anti-angiogenic effect of the KD may be attributed to its ability to suppress the hypoxic response, considering that hypoxia serves as the primary regulator of VEGF expression. 62 Furthermore, our study demonstrated a significant reduction in IGF-1 levels following KD administration. IGF-1 is a key growth factor known to stimulate the release of VEGF. 63 In contrast, our study did not observe similar anti-angiogenic activity with DOX monotherapy. This discrepancy underscores the potential complementary benefits of combining KD with DOX therapy to enhance angiogenesis control in cancer treatment.
Numerous studies have explored the impact of dietary factors on the regulation of inhibitory immune checkpoints such as programmed cell death protein-1 (PD-1) and its ligand, PD-L1.49,64,65 PD-L1 acts as an inhibitory ligand by binding to PD-1 on T cells, thereby suppressing their activation to maintain self-tolerance under normal physiological conditions. 66 In pathological contexts, such as cancer, PD-L1 is upregulated in tumor cells. 66 Its interaction with PD-1 on T cells inhibits their activation, fostering immune evasion and tumor progression. 66
In our study, treatment with DOX elicited a significant reduction in PD-L1 levels in EMT-6 tumor cells, indicating a potential role for DOX in bolstering anti-cancer immune responses by mitigating PD-L1 expression. However, the effect of conventional chemotherapy agents like DOX on PD-L1 expression exhibits variability, with divergent findings reported, likely influenced by factors such as cancer subtype and treatment dosage.67,68 Remarkably, our findings indicate that the KD potentiated DOX's suppressive effect on PD-L1 expression. Previous research has linked KD with enhanced anti-tumor immunity, partially attributable to the suppression of inhibitory immune checkpoints such as PD-1/PD-L1 and cytotoxic T-lymphocyte antigen-4 (CTLA-4).49,64,65 This suggests that the combination of KD with DOX therapy may offer a promising approach for enhancing the effectiveness of cancer treatment by targeting immune checkpoint pathways.
Of significant interest, our research findings suggest that cellular death in tumors predominantly occurred through a necrotic, caspase-3-independent mechanism rather than apoptosis. This form of necrotic cell death is primarily induced by the administered treatments rather than being a pathological change typically observed in treatment-naïve, high-grade, rapidly growing tumors, which often carry a poor prognosis. Several observations support our conclusion. Firstly, tumors in the control group, devoid of any treatment, displayed minimal necrosis despite a significant increase in volume. Secondly, tumors subjected to various treatments did not exhibit notable hypoxia, as reflected by HIF-1α levels. In fact, post-chemotherapy responses, as assessed by tumor necrosis, have been shown to define good responders in cancer patients and may predict improved event-free and overall survival.69,70
Recent studies have proposed that DNA damage induced by DOX may trigger apoptosis or other cellular events resulting in cell death or growth arrest. 71 Additionally, aside from apoptosis, there is evidence suggesting that autophagy, early or accelerated senescence, and necrosis may also play significant roles in DOX activity. 71 This becomes particularly crucial in cases of apoptosis resistance, where cancer cells evade this mode of death by inhibiting apoptotic signal activation. 71 Consequently, DOX can remain effective even independently of the apoptotic pathway. Our findings suggest the presence of apoptosis resistance, in which the tested treatments failed to overcome. However, the evaluated nutritional adjuvants may have enhanced DOX sensitivity to necrosis, as evidenced by histopathological analysis revealing increased necrosis occurrence in groups receiving nutritional adjuvants compared to DOX monotherapy. This enhanced sensitivity to necrosis could be attributed to the ability of both the KD and high-dose VitC to deplete ATP in tumor cells, leading to a bioenergetic catastrophe,24,72 thereby facilitating a more potent induction of necrosis compared to DOX monotherapy. It is important to note that considering the various mechanisms of action demonstrated by DOX, the drug's targets may vary among different tumors. Consequently, the significance of each target and mechanism may differ in individual cases. 73 Moreover, the concentration of DOX also appears to influence which of the drug's actions primarily contribute to cell death or growth arrest. 73
Several limitations should be considered when interpreting our findings. First, the long-term effects on tumor growth, recurrence, and metastasis remain unknown. Second, only a single EMT-6 breast cancer model was used, which may limit generalizability to other tumor types or to clinical settings. Third, the study was not blinded during treatment administration or data analysis, which could introduce observer bias, although standardized protocols were employed to minimize this. Additionally, not all hallmarks of cancer were studied, and not all relevant molecular markers or signaling pathways were evaluated. Finally, although valuable for controlled experimentation, the preclinical mouse model does not fully replicate the complexities of human cancer, including patient heterogeneity, variable tumor microenvironments, and the presence of comorbidities. These limitations highlight the need for further studies to validate and extend our findings in additional models and under varied experimental conditions.
Clinical relevance and future directions
The findings of this study carry several clinical implications despite the absence of a synergistic effect of the combined KD and high-dose VitC regimen on overall tumor volume. The KD demonstrated consistent modulation of clinically important biomarkers, including reductions in IGF-1, HIF-1α, VEGF, and PD-L1, as well as promotion of necrotic tumor cell death. These changes point to potential adjuvant benefits of the KD in attenuating proliferative, angiogenic, and immune-evasion pathways directly relevant to cancer progression and therapeutic resistance. Clinically, such effects may translate into improved responses to standard treatments, delayed tumor progression, or enhanced sensitivity to immunotherapy and chemotherapy.
High-dose VitC, although less impactful, showed clinically meaningful effects by reducing TERT expression and increasing collagen deposition within tumors. Reduced TERT may limit the proliferative capacity of residual cancer cells, while increased collagen deposition may impede local invasion and metastatic spread, collectively suggesting a potential to decrease the likelihood of tumor recurrence. These findings demonstrate that nutritional strategies may provide clinical benefit by influencing tumor biology and microenvironment.
From a translational perspective, these results highlight that nutritional interventions can influence clinically relevant pathways such as tumor metabolism, angiogenesis, immune evasion, and metastatic potential, potentially improving long-term outcomes. Future research should therefore focus on (i) optimizing treatment protocols, particularly dosing schedules for VitC (ii) identifying patient subgroups or tumor subtypes most likely to benefit, and (iii) evaluating combined nutritional and pharmacologic interventions in clinically relevant models. Ultimately, these steps are needed to advance from experimental findings toward safe and effective integration of nutritional adjuvants in clinical oncology practice.
Conclusion
Our study found that the integrated nutritional strategy did not significantly enhance the tumor-inhibitory effects of DOX compared to the individual nutritional adjuvants. However, the KD demonstrated positive outcomes across several cancer-associated markers, whereas high-dose VitC showed limited efficacy. It is imperative to explore different administration regimens for the combined use of the KD and high-dose VitC to determine the optimal approach for their joint utilization. Furthermore, considering the potential variability in efficacy based on cancer type or subtype and the specific drug used, extensive research is warranted to elucidate the mechanisms underlying these effects. Additionally, identifying potential responders to these nutritional adjuvants and elucidating relevant biomarkers for their efficacy represent crucial avenues for future investigation. Overall, our findings underscore the need for further research to better understand and optimize the potential of the proposed combinatory approach in cancer therapy.
Footnotes
Acknowledgment
None.
ORCID iDs
Ethical considerations
The animal experiment was conducted in accordance with the Public Health Service Policy on Humane Care and Use of Laboratory Animals (NIH Publication No. 15-8013, revised 2015). Ethical approval was obtained from the Research Ethical Committee at the University of Jordan (protocol number: 609/2021/19; approval date: 23 May 2021).
Authors’ contributions
DA Conceptualization, Methodology, Writing−original draft, Visualization, Investigation. HT Resources, Writing−review & editing, Funding acquisition. WT Resources, Writing−review & editing. MH Investigation. DA Investigation. NA Investigation. MA Resources, Writing−review & editing. All authors read and approved the final manuscript.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was funded by The Deanship of Scientific Research, The University of Jordan, Amman, Jordan, and The National Center for Research and Development, The Higher Council for Science and Technology, Amman, Jordan. Funding sources had no role in the design, analysis, and interpretation of the data; and preparation, review, or approval of the manuscript.
Declaration of conflicting interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Data availability
All relevant data are within the manuscript.