
Abstract
Chronic variable stress (CVS) is a well-established translational model that captures the unpredictability, persistence, and cumulative burden of long-term stress exposure. Through repeated application of heterogeneous mild-to-moderate stressors in a non-habituating manner, CVS induces sustained activation of the hypothalamic–pituitary–adrenal (HPA) axis, chronic glucocorticoid excess, oxidative stress, and neuroinflammatory signaling. These processes collectively drive coordinated dysfunction across multiple organ systems. Accumulating evidence now indicates that CVS pathology extends beyond the central nervous system (CNS), engaging the gut–liver–brain axis as a key integrative network linking stress to systemic metabolic and inflammatory outcomes. Stress-induced impairment of intestinal barrier integrity, dysregulation of nutrient transporters, hepatic lipid accumulation, iron dyshomeostasis, and cardiometabolic disturbances synergistically amplify both peripheral and neural vulnerability. Magnesium-L-theanine (MgT), a novel complex combining magnesium with the green tea-derived amino acid L-theanine, has emerged as a promising multi-target intervention against CVS-induced multisystem injury. Preclinical studies demonstrate that MgT preserves epithelial barrier architecture, restores tight junction and mucus-associated proteins, normalizes intestinal nutrient transporter expression, and attenuates hepatic steatosis, oxidative stress, and iron-driven metabolic damage under CVS conditions. These protective effects converge on the reactivation of key metabolic regulators, including nicotinamide adenine dinucleotide (NAD+)/sirtuin 1 (SIRT1) and PPARγ signaling pathways, positioning MgT as a modulator of metabolic flexibility and redox resilience. Co-administration with the NAD+ precursor nicotinamide riboside further potentiates these responses by reinforcing NAD+ availability and downstream metabolic control. In parallel, human studies of magnesium and L-theanine supplementation report anxiolytic, antidepressant, sleep-promoting, and anti-inflammatory benefits, supporting the translational relevance of this combined approach. This review integrates current evidence on CVS-induced multisystem pathology, with a focus on the gut–liver–brain axis, and proposes MgT as a trace-element-based, systems-oriented strategy to mitigate chronic stress-related gastrointestinal, metabolic, and neuropsychiatric complications, while highlighting key mechanistic gaps and translational priorities for clinical advancement.
Plain language summary
Chronic stress is not limited to psychological symptoms but affects multiple organs involved in metabolic and hormonal regulation. Long-term exposure to stress can disrupt stress hormone balance, increase inflammation, and impair communication between the gut, liver, and brain. This interconnected system plays a critical role in maintaining metabolic health, immune balance, and brain function. When it is disturbed, intestinal barrier function weakens, nutrient handling is altered, fat accumulates in the liver, iron balance becomes dysregulated, and the risk of metabolic and mood-related disorders increases. Magnesium-L-theanine is a nutritional compound that combines magnesium with L-theanine, a natural amino acid found in green tea. Experimental studies show that this combination helps protect the intestinal barrier, improves nutrient transport, reduces liver fat accumulation, and limits oxidative and inflammatory damage during chronic stress. These effects are linked to improved cellular energy balance and metabolic regulation. When combined with a supplement that supports cellular energy production, these protective actions appear to be further enhanced. Clinical studies of magnesium and L-theanine supplementation also report improvements in anxiety, sleep quality, and inflammatory markers. Together, these findings suggest that Magnesium-L-theanine may represent a promising supportive strategy for reducing the metabolic and endocrine consequences of chronic stress by targeting the gut–liver–brain axis.
Introduction
Stress responses are evolutionarily conserved adaptive programs that maintain physiological homeostasis under environmental challenge. When stress exposure becomes prolonged, unpredictable, or cumulative, however, these mechanisms transition from adaptive to maladaptive states, characterized by sustained neuroendocrine activation, metabolic inflexibility, and multisystem dysfunction.1 –3 Among experimental paradigms, chronic variable stress (CVS), also termed chronic mild or unpredictable stress, has emerged as one of the most informative models for capturing the integrated biology of human chronic stress, spanning behavioral, neuroendocrine, metabolic, and inflammatory domains.4,5
CVS employs rotating, unpredictable stressors over extended periods, thereby preventing habituation and driving persistent activation of the hypothalamic–pituitary–adrenal (HPA) axis, glucocorticoid excess, and oxidative burden.6,7 Behaviorally, this sustained neuroendocrine pressure manifests as anhedonia, reduced exploratory behavior, and increased behavioral despair, phenotypes that parallel key features of depressive disorders. 8 At the cellular level, CVS induces marked impairments in mitochondrial bioenergetics, particularly reduced activities of complexes II and IV, in the prefrontal cortex and hippocampus, identifying mitochondrial energy failure as a core signature of chronic stress pathology.6,9
A major conceptual advance in the field is the recognition that CVS-induced pathology extends beyond the CNS to engage the gut–liver–brain axis as an integrated stress–response network. Chronic stress disrupts intestinal epithelial architecture, downregulates tight junction proteins (including claudins, occludin, and ZO-1), and reduces mucin (MUC2) expression, resulting in increased intestinal permeability and the translocation of luminal antigens and lipopolysaccharide (LPS).10,11 Portal influx of microbial products subsequently promotes hepatic inflammation, oxidative injury, and fibrogenic signaling.12,13 In parallel, CVS induces a distinctive hepatic iron dyshomeostasis phenotype, characterized by increased Fe(II) accumulation, upregulation of transferrin receptor-1 (TFR1) and Zrt- and Irt-like protein 14 (solute carrier family 39 member 14; ZIP14), and suppression of ferritin subunits and miR-181, linking chronic stress to ferroptosis-prone metabolic vulnerability.12,14 These peripheral disturbances feed back onto central stress circuits, reinforcing neuroinflammation, autonomic imbalance, and cardiovascular dysfunction, including reduced heart rate variability and endothelial impairment.15,16
Although conventional pharmacotherapies can alleviate selected neuropsychiatric manifestations of chronic stress, their limited efficacy across organ systems and concerns regarding tolerance and long-term safety highlight the need for multi-target, biologically coherent strategies. Within this context, magnesium, a trace element essential for ATP-dependent metabolism, N-methyl-D-aspartate receptor (NMDA receptor) regulation, and HPA-axis buffering, and L-theanine, a green-tea-derived amino acid with GABAergic, monoaminergic, and nuclear factor erythroid 2-related factor 2 (Nrf2)-activating properties, have attracted increasing interest as modulators of stress physiology.17 –19
The magnesium–L-theanine (MgT) complex integrates these properties into a single molecular formulation. Experimental and early clinical studies suggest that MgT enhances central magnesium bioavailability, normalizes stress-sensitive neurotransmitter systems, and suppresses glucocorticoid hypersecretion more effectively than either component alone.20 –22 Importantly, emerging preclinical evidence indicates that MgT also restores intestinal barrier integrity, reactivates nutrient transport systems, attenuates hepatic steatosis and oxidative stress, and engages nicotinamide adenine dinucleotide (NAD+)-dependent metabolic regulators such as sirtuin 1 (SIRT1) and PPARγ, positioning MgT as a system-level modulator of stress biology.13,23 Synergistic effects with nicotinamide riboside (NR) further highlight NAD+ metabolism as a convergent node in stress-induced metabolic dysfunction.24,25
In this review, we integrate experimental, mechanistic, and translational evidence to propose a unified framework in which MgT counteracts chronic stress pathology through coordinated actions on neuromodulation, epithelial integrity, iron and lipid metabolism, and NAD+-centered bioenergetics. By moving beyond reductionist, single-pathway paradigms, this system-oriented perspective positions MgT and its potentiation by NAD+-boosting strategies as a promising next-generation approach for stress-associated gastrointestinal, metabolic, and neuropsychiatric dysfunctions. Collectively, these interconnected processes position CVS as a multisystem condition mediated through coordinated disruption of the gut–liver–brain axis (Figure 1). Given the growing translational interest in MgT, particular attention must also be paid to its dose–response characteristics, safety profile, and metabolic behavior, which remain critical determinants of its therapeutic development.

Conceptual overview of CVS and the gut–liver–brain axis. CVS models real-life chronic stress by exposing animals to unpredictable, mild-to-moderate stressors over several weeks. Persistent HPA axis activation, glucocorticoid excess and oxidative stress lead to multisystem dysregulation. In the brain, CVS impairs mitochondrial energy metabolism and monoamine balance, contributing to depressive- and anxiety-like behavior. In the gut, CVS disrupts tight junctions and the mucus layer, increasing intestinal permeability and facilitating the translocation of luminal antigens into the portal circulation.
Materials and methods
This review was developed through a comprehensive synthesis of experimental and translational evidence pertaining to CVS and the mechanistic actions of the MgT across the gut–liver–brain axis. Literature searches were conducted in PubMed, Web of Science, Scopus, and Google Scholar covering the period 1990–2025, using combinations of the following terms: “chronic variable stress,” “chronic mild stress,” “chronic unpredictable stress,” “magnesium,” “L-theanine,” “magnesium-L-theanine,” “intestinal permeability,” “gut–liver axis,” and “mitochondrial dysfunction.” Additional studies were identified through backward citation tracking of relevant articles. Inclusion criteria focused on experimental CVS/CMS/CUS studies reporting behavioral, neuroendocrine, mitochondrial, intestinal, hepatic, or cardiometabolic outcomes, as well as interventional studies evaluating magnesium, L-theanine, MgT, or NR. Studies without mechanistic data, that employed only acute stress paradigms, or that failed to report intervention composition or dosage were excluded. Eligible studies were examined for stress protocol characteristics, intervention type and dose, behavioral assays, biochemical and molecular endpoints, histopathology, mitochondrial function, NAD+ metabolite quantification, iron-handling markers, nutrient transporter expression, and indicators of barrier integrity.
Data extracted from individual studies were organized into mechanistic domains encompassing neuroendocrine regulation of the HPA axis; neurotransmission and synaptic plasticity; mitochondrial bioenergetics and redox control; gut epithelial structure and nutrient absorption; hepatic lipid metabolism and inflammatory signaling; and NAD+/SIRT1–PPARγ activity. Mechanistic diagrams presented in this review were constructed on the basis of convergent findings replicated across at least three independent studies or strongly supported by molecular evidence.
Study quality was evaluated heuristically against ARRIVE-aligned criteria, including clarity of stressor descriptions, randomization, blinding, sample-size justification, and methodological rigor in biochemical and histological analyses. Studies with substantial methodological ambiguity were incorporated only for contextual interpretation. No new animal or human experiments were conducted; all referenced studies reported adherence to institutional and national ethical guidelines. This methodological framework enabled the integration of multi-organ findings into a unified systems-level model of CVS pathology and MgT action, supporting subsequent analyses of MgT + NR synergy and outlining a mechanistic roadmap for future experimental and clinical investigations.
CVS: Models and multisystem effects
Conceptual basis and construct validity
The CVS model was developed to more faithfully recapitulate real-life stress exposure, which is rarely constant, uniform, or predictable.3,5 Unlike single-stressor paradigms, CVS employs a rotating schedule of mild-to-moderate stressors delivered in an unpredictable sequence and at variable times, thereby preserving novelty and preventing habituation.1,6,8,26 This design sustains activation of the HPA axis and produces stable behavioral and physiological alterations. CVS exhibits strong construct validity by capturing key dimensions of human chronic stress, including unpredictability, uncontrollability, and cumulative burden. The emergence of anxiety supports face validity- and depression-like behaviors, whereas predictive validity is demonstrated by the partial reversal of these phenotypes following chronic antidepressant treatment.2,27 Collectively, these features establish CVS as a leading experimental framework for investigating the multisystem consequences of chronic stress.
Behavioral and neurobiological outcomes
CVS elicits a robust and highly reproducible behavioral phenotype that closely mirrors core features of human stress-related psychopathology. A hallmark outcome is anhedonia, most commonly assessed as reduced sucrose preference, reflecting impaired reward processing. 27 CVS also induces depression-like behavior, evidenced by increased immobility in forced swim and tail suspension paradigms, indicative of behavioral despair and compromised stress coping.6,28 Anxiety-like behavior frequently co-emerges, characterized by reduced open-arm exploration in the elevated plus maze and decreased center exploration in the open-field test, accompanied by diminished locomotor and exploratory activity, consistent with motivational deficits and psychomotor slowing.27,29 These integrated alterations define the core neurobehavioral phenotype of chronic stress (Figure 2).

Integrated effects of CVS and MgT on gut–liver metabolic integrity and stress resilience. CVS induces sustained glucocorticoid exposure and reactive oxygen species generation, leading to coordinated barrier and metabolic injury across the gut–liver axis. CVS disrupts intestinal epithelial integrity by downregulating tight junction proteins (claudin-1, claudin-4, occludin, ZO-1) and mucin-2, leading to increased epithelial permeability and lipopolysaccharide translocation into the portal circulation. In parallel, stress impairs intestinal nutrient transport by suppressing glucose (SGLT1, GLUT2), peptide (PepT1, PepT2), and amino acid (LAT2, EAAT3) transporters. At the hepatic level, CVS promotes maladaptive iron handling characterized by increased redox-active Fe(II), dysregulation of the ZIP14/miR-181 axis, ferritin imbalance, and activation of lipogenic programs (SREBF1, FASN), culminating in triglyceride accumulation and metabolic dysfunction. MgT exerts multi-level protective actions by restoring tight junction architecture and mucus barrier integrity, normalizing nutrient transporter expression, attenuating ZIP14-mediated iron overload, suppressing lipogenesis, and replenishing cellular NAD+ pools.
At the neurobiological level, CVS disrupts multiple systems governing emotional and cognitive regulation. Repeated stress exposure reduces monoaminergic signaling, including serotonin, dopamine, and noradrenaline, within stress-sensitive regions such as the hippocampus and prefrontal cortex, paralleling neurotransmitter alterations observed in mood and anxiety disorders.3,8,30 In parallel, CVS compromises mitochondrial bioenergetics, markedly reducing the activity of respiratory chain complexes II and IV and limiting ATP production in corticolimbic circuits critical for affective regulation.6,31
A pronounced oxidative and inflammatory signature accompanies these alterations. CVS increases lipid peroxidation while impairing endogenous antioxidant defenses, including superoxide dismutase, catalase, and glutathione peroxidase, thereby heightening neuronal vulnerability.30,32 Sustained stress also promotes microglial activation and neuroinflammation, further destabilizing synaptic integrity and neuronal resilience.7,33 In the hippocampus, reductions in brain-derived neurotrophic factor and suppression of adult neurogenesis represent convergent mechanisms linking chronic stress to cognitive impairment and emotional dysregulation.27,30
Immune, endocrine, and metabolic consequences
Persistent exposure to CVS induces sustained dysregulation of the neuroendocrine system, driven primarily by prolonged HPA axis activation. This state is characterized by chronically elevated basal glucocorticoid levels, disruption of circadian cortisol/corticosterone rhythmicity, and impaired negative feedback sensitivity, collectively imposing a substantial allostatic load on the organism.34 –36 Such neuroendocrine maladaptation propagates systemic consequences that extend beyond the CNS, reshaping peripheral metabolic and immune homeostasis. 37 Concomitantly, CVS promotes a shift toward low-grade, chronic inflammation, characterized by elevated circulating levels of pro-inflammatory cytokines, including interleukin-1beta (IL-1β), IL-6, and tumor necrosis factor-alpha (TNF-α).7,38 These immune signals engage bidirectionally with HPA axis activity and microglial function, reinforcing neuroinflammatory cascades that contribute to stress-induced behavioral and cognitive alterations.
Metabolic homeostasis is similarly compromised under CVS. Sustained glucocorticoid exposure drives insulin resistance, hyperglycemia, and impaired glucose tolerance, while dyslipidemia, particularly elevated triglycerides and altered cholesterol profiles, increases cardiometabolic vulnerability.39,40 Changes in body weight and adipose tissue distribution further reflect altered energy partitioning and metabolic inflexibility, with outcomes dependent on sex, diet, and stress duration (Figure 2). Overlaying these endocrine, immune, and metabolic disturbances is a pervasive disruption of redox balance. CVS enhances lipid peroxidation and oxidative damage to proteins and DNA while suppressing endogenous antioxidant defenses, amplifying tissue vulnerability across multiple organ systems.10,32 Collectively, these interconnected perturbations provide a mechanistic basis for the emergence of hepatic, cardiovascular, and gastrointestinal comorbidities frequently observed in chronic stress states, underscoring the multisystem nature of CVS pathology. 11
Gut–liver axis and iron/lipid metabolism
As highlighted in the “Introduction” section, CVS disrupts gut barrier integrity and hepatic homeostasis, positioning the gut–liver axis as a central conduit of stress-induced pathology (Figures 1 and 4(a)). CVS reduces the expression of key tight junction proteins, induces mucosal injury, and promotes dysbiosis, collectively increasing intestinal permeability. The resulting translocation of LPS and microbial products into the portal circulation activates Kupffer cells and hepatic stellate cells, driving hepatic inflammation, oxidative stress, and pro-fibrotic signaling.12,23,41,42
Disruption of iron homeostasis has emerged as a pivotal mechanistic hub within the integrated stress–response landscape. CVS is characterized by a paradoxical hepatic iron phenotype, marked by a reduction in total iron stores alongside selective enrichment of redox-active ferrous iron. This shift coincides with the coordinated induction of iron uptake pathways, notably TFR-1 and ZIP14, as well as the repression of ferritin subunits and miR-181 expression.12,43 Such remodeling reflects a failure of adaptive iron buffering, creating a cellular milieu permissive to ferroptosis-related susceptibility and intensified lipid peroxidation-driven oxidative damage. Concomitantly, CVS provokes hepatic triglyceride deposition and stimulates lipogenic transcriptional programs, including the activation of sterol regulatory element-binding factor-1 and fatty acid synthase, thereby recapitulating key molecular hallmarks of stress-associated non-alcoholic fatty liver disease-like remodeling.44,45 These findings highlight the integrated nature of stress-induced epithelial, metabolic, and inflammatory injury along the gut–liver axis (Figure 2).
Beyond its metabolic and inflammatory consequences, stress-related liver pathology is increasingly recognized as an active driver of depression-like phenotypes through disruption of the gut–liver–brain axis. In preclinical models, hepatic ischemia/reperfusion injury and bile duct ligation induce a coordinated syndrome encompassing systemic inflammation, gut microbiota dysbiosis, and profound behavioral alterations, including anhedonia, behavioral despair, and synaptic deficits in the prefrontal cortex.46 –48 These findings indicate that hepatic injury propagates centrally relevant signals that can reshape neural circuits involved in mood regulation. Notably, subdiaphragmatic vagotomy markedly attenuates these phenotypes, implicating vagal pathways as a key conduit through which peripheral liver dysfunction is translated into affective disturbances.47,48 Convergent evidence from chronic stress paradigms further supports a bidirectional coupling between hepatic dysfunction and behavioral pathology. Chronic unpredictable mild stress induces hepatic metabolic disruption, oxidative stress, and inflammatory activation in parallel with anxiety- and depression-like behaviors.49 –52 Mechanistically, stress-induced gut dysbiosis and barrier failure promote portal translocation of microbial products, amplifying hepatic inflammation and systemic cytokine release, including IL-6 and TNF-α, which, in turn, drive neuroinflammation and synaptic remodeling within corticolimbic circuits.53 –55 Consistent with these experimental insights, clinical studies in chronic liver disease populations report a high prevalence of fatigue, social withdrawal, and depression, underscoring the translational significance of this axis.53,55
Vagal signaling in the gut–liver–brain axis
Vagal signaling constitutes a central, yet frequently underemphasized, axis of integration within the gut–liver–brain network. As a rapid and bidirectional neural conduit, the vagus nerve links peripheral metabolic and immune states to central autonomic and neuroendocrine control. Afferent vagal fibers relay signals derived from gut microbiota, intestinal metabolites, and hepatic metabolic activity to the nucleus tractus solitarius, enabling real-time integration of peripheral cues into brainstem regulatory circuits.56,57 In parallel, efferent vagal outputs modulate intestinal motility, epithelial barrier integrity, mucosal immunity, and hepatic metabolism, thereby establishing a coordinated vago–vagal reflex that sustains gut–liver homeostasis.58,59
Recent work has delineated a liver–brain–gut neural arc as a fundamental regulatory circuit governing intestinal immune equilibrium. This pathway is required for the maintenance of peripheral regulatory T cell (pTreg) niches in the gut, and its disruption leads to reduced pTreg abundance and heightened susceptibility to intestinal inflammation, positioning vagal signaling as a key determinant of mucosal immune tone.56,59 Beyond immune regulation, vagal afferent pathways transmit hepatic-derived inflammatory and metabolic signals to the brain, thereby contributing to early neurocognitive and affective alterations observed in liver disease.57,60 Importantly, experimental models provide direct evidence linking vagal signaling to behavioral outcomes. Depression-like phenotypes induced by hepatic ischemia/reperfusion injury or bile duct ligation are markedly attenuated by subdiaphragmatic vagotomy, indicating that vagal pathways mediate the translation of peripheral liver pathology into central affective dysfunction.47,48 Within CVS paradigms, impaired vagal tone, reflected by reduced heart rate variability and autonomic imbalance, may further amplify this axis by promoting intestinal barrier disruption, hepatic inflammation, and neuroinflammatory signaling. As a result, these findings position the vagus nerve as a systems-level integrator of metabolic, immune, and neural processes within the gut–liver–brain axis, and highlight vagal signaling as a tractable target for restoring resilience to chronic stress.
Cardiovascular and autonomic outcomes
Chronic stress disrupts autonomic cardiovascular regulation through a shift toward sympathetic predominance and impaired vagal/baroreflex control. In humans exposed to chronic psychosocial stress, this pattern is characterized by higher blood pressure, altered HRV spectral indices consistent with sympathetic dominance, and reduced baroreflex sensitivity.16,61,62 These autonomic alterations are accompanied by endothelial dysfunction, fostering a pro-inflammatory and pro-atherogenic vascular environment.63,64 Importantly, this cardiovascular phenotype closely parallels the mechanistic evidence linking chronic psychosocial stress to prothrombotic states, endothelial dysfunction, and increased vascular risk in both healthy individuals and cardiovascular disease patients 63 (Figures 2 and 6(a)). These findings underscore chronic stress as a system-level cardiovascular modifier and provide a strong rationale for integrative interventions, such as MgT, that simultaneously target central stress circuits and peripheral autonomic–metabolic pathways.
Strengths and limitations of the CVS model
The CVS paradigm has become a cornerstone preclinical model for interrogating the biological consequences of sustained stress exposure and for evaluating interventions targeting stress-driven multisystem dysfunction. Its major translational strength lies in its capacity to recapitulate defining features of human chronic stress, unpredictability, uncontrollability, and cumulative allostatic load, which collectively drive persistent neuroendocrine, metabolic, and inflammatory dysregulation. By employing a rotating schedule of heterogeneous stressors, CVS minimizes habituation and elicits durable alterations across both central and peripheral physiological systems, closely mirroring stress-associated phenotypes observed in clinical populations.
A key advantage of CVS is its ability to generate an integrated, system-level readout of stress biology. In contrast to single-stressor paradigms, CVS enables concurrent assessment of behavioral phenotypes, neural circuit adaptations, gut barrier integrity, hepatic metabolic remodeling, and autonomic-cardiovascular function within a unified experimental framework.2,65 This multidimensional resolution is particularly well suited for studying the gut–liver–brain axis, where bidirectional signaling between peripheral metabolic organs and central stress circuits critically shapes vulnerability to neuropsychiatric and metabolic disorders. Consistent with this translational breadth, CVS is responsive to multiple antidepressant interventions and remains a valuable preclinical screening model. 5 Nevertheless, several limitations temper cross-study comparability. Substantial heterogeneity in stressor composition, duration, and sequencing across laboratories introduces variability in behavioral and molecular outcomes, complicating synthesis and replication. 66 Moreover, biological modifiers such as sex, genetic background, and age exert strong influences on stress responsivity and adaptive capacity, underscoring the need for stratified designs and transparent reporting. 36 As with all rodent models, CVS also cannot fully capture higher-order dimensions of human stress, including cognitive appraisal and sociocultural context. 2
Despite these constraints, CVS remains an indispensable framework for probing integrated stress pathophysiology when applied with methodological rigor. Its relevance is particularly evident for evaluating pleiotropic interventions such as MgT, whose coordinated actions span neuroendocrine regulation, synaptic signaling, intestinal barrier function, and hepatic metabolism. The alignment between CVS-induced multisystem disruption and the network-level protective mechanisms engaged by MgT positions this model as a powerful platform for testing strategies aimed at restoring systemic homeostasis rather than targeting isolated molecular pathways (Figures 3–6).
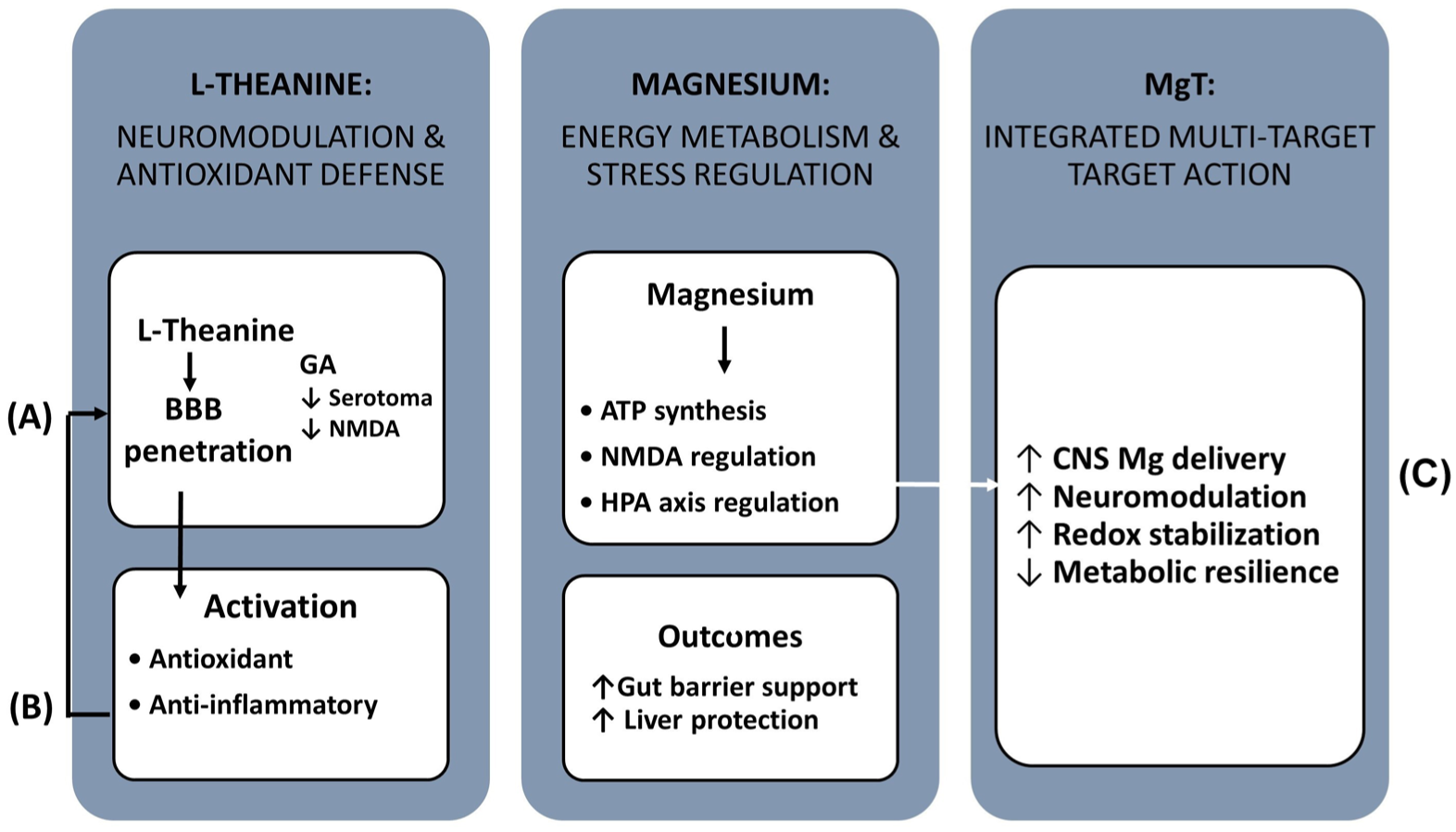
Pharmacological properties of L-theanine, magnesium, and the MgT complex. (a) L-theanine, a non-protein amino acid from green tea, crosses the blood–brain barrier and exerts anxiolytic, antioxidant, and neuromodulatory effects. It enhances GABAergic, serotonergic, and dopaminergic signaling, attenuates glutamatergic overactivation, and activates Nrf2-, AMPK–mTOR-, and JAK2/STAT3-related antioxidant and cytoprotective pathways. (b) Magnesium is an essential cofactor for ATP synthesis, nucleic acid metabolism, and signal transduction. It modulates NMDA receptor activity, supports mitochondrial function, regulates the HPA axis, and reduces stress-induced endocrine and inflammatory responses. (c) The MgT complex integrates the complementary actions of L-theanine and magnesium, potentially improving magnesium delivery to the central nervous system and providing multi-target effects on neurotransmission, oxidative stress, energy metabolism and stress hormone regulation.

Magnesium-L-theanine protects the gut–liver axis against CVS. (a) CVS disrupts the intestinal barrier by shortening villi, altering crypt architecture and decreasing the expression of tight junction proteins (Claudin-1, Claudin-4, Occludin, ZO-1) and the mucus component MUC-2. Increased intestinal permeability facilitates translocation of luminal antigens (e.g., LPS) into the portal circulation, promoting hepatic inflammation, oxidative stress, and fibrosis. In the liver, CVS induces Fe(II) accumulation via upregulation of TFR1 and ZIP14 and downregulation of ferritin and miR-181, while also increasing lipid peroxidation (MDA), decreasing GSH-Px activity and activating lipogenic factors (SREBF1, FASN), leading to triglyceride accumulation. (b) MgT supplementation dose-dependently preserves jejunal morphology, restores tight junction and mucus-related proteins, and reactivates key nutrient transporters for glucose (SGLT1, GLUT2), peptides (PepT1, PepT2), amino acids (LAT2, EAAT3), and lipids (FABP2, FATP4). (c) In the liver, MgT reduces triglyceride accumulation, improves redox balance, increases NAD+ and NADPH, and upregulates SIRT1 and PPARγ while suppressing SREBF1 and FASN expression. Collectively, these changes demonstrate that MgT mitigates CVS-induced structural and functional injury along the gut–liver axis.

Hypothesized effects of MgT on brain energy metabolism and synaptic function under CVS. (a) CVS impairs mitochondrial complex II and IV activities in the hippocampus and prefrontal cortex, decreases ATP production, increases ROS, reduces BDNF and neurogenesis, and promotes neuroinflammation and microglial activation. These changes contribute to depressive- and anxiety-like behaviors and cognitive deficits. (b) MgT is proposed to restore brain energy metabolism and synaptic function by improving mitochondrial ATP production (magnesium-dependent processes), enhancing antioxidant defenses via L-theanine-driven Nrf2 activation, normalizing glutamate–GABA balance (NMDA receptor inhibition and GABA support), modulating monoamine levels (serotonin, dopamine) and potentially enhancing NAD+-dependent SIRT1 activity and mitochondrial biogenesis.

Comparative and combined strategies for mitigating CVS. (a) Conceptual radar plot comparing the mechanistic coverage of selected interventions. Classical antioxidant vitamins (C/E) mainly provide direct ROS scavenging, whereas exercise exerts broad systemic effects on endocrine, neurotransmitter, and metabolic regulation. MgT integrates antioxidant regulation, neuromodulation, gut barrier protection, and metabolic control. The addition of NR is expected to further extend MgT effects toward NAD+-dependent pathways (e.g., SIRT1, PPARγ). (b) Mechanistic overview of the MgT and NR combination in CVS models. MgT preserves intestinal barrier integrity and improves hepatic redox and lipid homeostasis, while NR increases NAD+/NADPH availability and activates SIRT1- and PPARγ-dependent metabolic pathways. Together, MgT and NR synergistically support gut–liver axis integrity and metabolic resilience under chronic stress.
Pharmacological properties of MgT
L-theanine: Absorption, distribution, and mechanisms of action
L-theanine (γ-glutamylethylamide) is a non-proteinogenic amino acid abundant in green tea (Camellia sinensis) with well-documented neuroactive and stress-modulatory properties across species. Following oral administration, L-theanine is efficiently absorbed in the small intestine via sodium-dependent amino acid transporters and rapidly distributed systemically, reaching peak serum concentrations within approximately 50 min and peak brain concentrations within approximately 5 h.67 –69 Evidence from multiple animal species, including poultry and pigs, suggests that L-theanine may influence stress-related physiology, immune responses, and intestinal microbial balance. 70 Mechanistically, L-theanine exerts convergent effects on central neurotransmission and neuroendocrine regulation. It enhances GABAergic tone and increases dopamine and serotonin availability in corticolimbic regions, while modulating glutamatergic signaling through functional interactions with α-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid receptor (AMPA receptor) and NMDA receptor, thereby constraining stress-induced excitatory overdrive and excitotoxic vulnerability.68,71,72 These actions are accompanied by attenuation of HPA-axis activity, reflected by reduced ACTH, corticosterone, and noradrenergic output, alongside normalization of monoaminergic balance.68,73 –76
Beyond the CNS, L-theanine engages antioxidant and immunomodulatory pathways that are highly relevant to CVS. Experimental studies demonstrate activation of Nrf2-dependent transcriptional programs, upregulation of endogenous antioxidant enzymes (SOD, CAT, GSH-Px), and suppression of pro-inflammatory mediators, including inducible nitric oxide synthase (iNOS), IL-6, and TNF-α.19,69 In poultry and other animal models, dietary L-theanine improves mucosal immunity, enhances secretory IgA responses, and favorably shapes cytokine profiles, indicating increased resilience to environmental and nutritional stressors.70,77,78 In parallel, L-theanine modulates gut microbiota composition, reducing potentially pathogenic taxa while supporting beneficial genera such as Lactobacillus, thereby linking neural, immune, and microbial components of the stress response. 79 These data position L-theanine as a pleiotropic neuromodulator and gut–immune regulator that integrates neurotransmitter balance, redox control, mucosal immunity, and neuroendocrine restraint. This systems-level profile provides a strong biological rationale for its inclusion in the MgT complex as a strategy to counteract CVS-induced dysfunction across the gut–liver–brain axis. The complementary and overlapping mechanisms of L-theanine and magnesium provide a coherent pharmacological basis for the MgT complex (Figure 3).
Magnesium and stress physiology
Magnesium is a fundamental divalent cation that functions as a cofactor for several hundred enzymatic reactions and is therefore indispensable for metabolic, endocrine, and neurophysiological homeostasis. Its central role in cellular energetics stems from the fact that Mg–ATP constitutes the biologically active form of ATP, serving as the obligatory substrate for kinases, molecular motors, and energy-dependent ion pumps, including Na+/K+-ATPase and Ca²+-ATPases. Beyond energy metabolism, magnesium stabilizes DNA and RNA structures, modulates chromatin architecture, and thereby influences transcriptional programs engaged during adaptive responses to prolonged stress exposure.80,81
At the neurophysiological level, magnesium exerts a critical inhibitory function by acting as a voltage-dependent blocker of NMDA receptors, limiting Ca²+ influx and constraining glutamate-driven excitotoxicity. Dysregulation of this mechanism has been repeatedly implicated in stress-induced neuronal injury, impaired synaptic plasticity, and affective disorders, positioning magnesium as a key determinant of the excitatory–inhibitory balance within stress-sensitive circuits of the hippocampus, prefrontal cortex, and amygdala.82,83 Through these actions, magnesium directly contributes to neural resilience under sustained allostatic challenge. Magnesium also plays a pivotal role in regulating endocrine stress pathways. Experimental and clinical findings support a role for magnesium in the regulation of HPA axis activity, with evidence for altered CRH expression, ACTH release, and downstream glucocorticoid regulation under conditions of magnesium deficiency or stress.17,22 Acute psychological or physical stress is associated with a transient redistribution of magnesium from intracellular compartments to the extracellular space, followed by enhanced renal excretion. While initially adaptive, this response becomes maladaptive when stressors are chronic or recurrent, leading to progressive depletion of intracellular and skeletal magnesium stores and the emergence of chronic latent magnesium deficiency.
The concept of a “magnesium–stress vicious circle,” first articulated in the early 1990s, has since been reinforced by a substantial body of experimental and clinical evidence demonstrating a bidirectional interaction between stress exposure and magnesium homeostasis.17,84 Within this framework, sustained stress accelerates magnesium loss, while declining magnesium availability weakens inhibitory control over NMDA receptor activity, catecholaminergic signaling and HPA-axis feedback. As a consequence, stress responsiveness becomes exaggerated, autonomic balance shifts toward sympathetic dominance and vulnerability to anxiety- and depression-like phenotypes increases.
Animals exposed to thermal, restraint, noise, or social stress exhibit reductions in serum and erythrocyte magnesium concentrations, accompanied by elevated catecholamine and glucocorticoid levels. Magnesium-deficient rodents exhibit increased CRH expression and elevated ACTH levels, exaggerated catecholaminergic stress responses, greater susceptibility to stress-induced gastric ulceration, and more pronounced anxiety- and depression-like behaviors in paradigms such as the light–dark, open-field, and forced-swim tests compared with magnesium-replete controls.22,85 –87
Importantly, evidence from avian heat-stress models further demonstrates that magnesium supplementation does not merely correct deficiency but actively buffers stress-induced metabolic and oxidative dysregulation. In Japanese quail exposed to high ambient temperature, dietary magnesium supplementation dose-dependently improved feed intake, growth performance, nutrient digestibility, and carcass yield, while markedly reducing serum and tissue lipid peroxidation and circulating triglyceride and cholesterol concentrations. Notably, magnesium proteinate conferred significantly greater protection than magnesium oxide, indicating superior bioavailability and tissue efficacy under stress conditions. 88 These findings provide early experimental evidence that targeted, organic magnesium formulations are more effective than inorganic salts in preserving metabolic resilience, redox balance, and lipid homeostasis during sustained allostatic challenge, and are mechanistically consistent with the magnesium–stress vicious-circle paradigm.
Clinical observations converge closely with these experimental findings. Population-based surveys indicate that although overt hypomagnesemia affects a minority of individuals, up to 40%–45% may exhibit chronic latent magnesium deficiency when higher functional thresholds are applied.89,90 Chronic psychological or physiological stress has been associated with altered magnesium homeostasis, including lower serum or erythrocyte magnesium and increased urinary magnesium loss in selected human studies; related literature also links such stress exposures to cortisol dysregulation and inflammatory burden.17,91,92 Epidemiological studies further link low dietary magnesium intake to higher perceived stress, greater severity of anxiety and depressive symptoms, and elevated systemic inflammation. Importantly, randomized controlled data indicate that magnesium supplementation may improve subjective insomnia measures and related biochemical correlates, and these effects may be more evident in individuals with low baseline magnesium status.17,93
Within this neurobiological framework, magnesium can be viewed as an important modulator of stress-related neural homeostasis. Magnesium deficiency weakens inhibitory control over excitatory signaling, particularly through impaired GABAergic support and excessive NMDA receptor activation, while stress conditions can promote magnesium loss and reduce its protective buffering capacity.88,94 This “vicious circle” directly motivates the development of formulations such as the MgT, which are explicitly designed to restore and stabilize magnesium status within stress-responsive tissues and to combine this effect with neuromodulatory, antioxidant, and NAD+-supporting actions. By breaking the positive feedback loop between magnesium loss and stress reactivity, MgT may exert disproportionately large benefits compared with non-targeted supplementation, particularly in CVS paradigms that recapitulate human allostatic overload.17,23,24
Rationale for the MgT complex
The integration of magnesium and L-theanine into a single molecular complex represents a biologically coherent and experimentally substantiated strategy for targeting the multidimensional nature of chronic stress pathology. Chronic stress is not driven by isolated molecular defects but by convergent disturbances in excitatory–inhibitory neurotransmission, neuroendocrine regulation, mitochondrial bioenergetics, and redox homeostasis. By combining two agents that act on complementary yet intersecting pathways, MgT is uniquely positioned to exert broader, more durable stress-buffering effects than either agent alone.17,20,21,23
A central limitation of conventional magnesium supplementation is its modest capacity to elevate brain magnesium concentrations, largely due to limited permeability across the blood–brain barrier and tight homeostatic control of Mg²+ transport. L-theanine, by contrast, is efficiently absorbed and readily transported into the brain via amino acid transport systems. Complexation with L-theanine may enhance central magnesium availability and amplify neurochemical effects, including modulation of inhibitory neurotransmission and related signaling pathways. 20 This feature is particularly relevant in chronic stress states, where NMDA receptor overactivation and calcium overload contribute to neuronal hyperexcitability and synaptic instability.80,82
At the level of neurotransmission, magnesium and L-theanine exert synergistic and mechanistically complementary effects. Magnesium acts as a voltage-dependent NMDA receptor antagonist, limiting excessive Ca²+ influx and glutamate-driven excitotoxicity L-theanine may influence inhibitory and monoaminergic neurobiological processes, including dopaminergic and serotonergic signaling, in ways that support stress reduction and emotional well-being. 68 When combined within the MgT complex, these actions converge to restore excitatory–inhibitory balance more effectively than either compound alone, a property that aligns with the core neurobiological deficits observed in anxiety, depression, and stress-related sleep disturbances.95,96
Beyond neuromodulation, MgT provides integrated metabolic and redox protection. L-theanine activates antioxidant and anti-inflammatory programs, including Nrf2-driven transcriptional responses and the suppression of iNOS and pro-inflammatory cytokines, while magnesium is indispensable for ATP synthesis, mitochondrial enzyme activity, and redox cycling.17,19 This dual-action profile supports mitochondrial efficiency, stabilizes cellular energy metabolism, and mitigates oxidative damage, processes that are consistently disrupted in CVS models and stress-associated metabolic disorders. Notably, both constituents of MgT exhibit favorable safety and tolerability profiles in humans. Magnesium supplementation within recommended ranges is well tolerated, and L-theanine has demonstrated excellent safety across acute and chronic dosing paradigms, including in stressed and clinically vulnerable populations.17,68 Their combination into a single complex is therefore unlikely to introduce additive toxicity, a critical consideration for long-term stress-modulatory interventions.
Preclinical evidence further supports the mechanistic validity of MgT as an integrative modulator of stress physiology. Compared with conventional magnesium salts, MgT increases cerebrospinal fluid magnesium levels more effectively, indicating superior central bioavailability. In parallel, MgT has been shown to increase brain serotonin, dopamine, and melatonin levels and to improve sleep-related outcomes in experimental models of caffeine-induced sleep disturbance. 20 These effects are accompanied by suppression of stress-induced corticosterone elevation and enhancement of endogenous antioxidant defenses, including superoxide dismutase, catalase and glutathione peroxidase, reflecting coordinated attenuation of HPA hyperactivity and oxidative burden. Collectively, these findings position MgT as a biologically integrated, multi-target intervention that directly addresses the neurochemical, metabolic, and redox dimensions of chronic stress. This mechanistic coherence provides a strong rationale for subsequent investigations into MgT-mediated regulation of the gut–liver–brain axis, mitochondrial bioenergetics, and NAD+-dependent signaling networks, and supports its evaluation as a next-generation strategy for mitigating chronic stress-related multisystem dysfunction (Figures 3–5).
Dose, efficacy, safety, and metabolic considerations of MgT
MgT is emerging as a system-level modulator of stress biology rather than a conventional nutritional adjunct. Although direct evidence remains limited, preclinical data consistently support dose-dependent efficacy across central and peripheral domains. In preclinical CVS models, MgT has been administered at doses ranging from 100 to 400 mg/kg, with higher doses consistently producing greater improvements in metabolic, endocrine, and epithelial outcomes, although direct translation to human-equivalent dosing remains to be established. 23 MgT complexes enhance slow-wave activity, modulate GABAergic and monoaminergic signaling, and improve sleep architecture, 20 while recent CVS studies demonstrate coordinated protection of gut–liver integrity, including restoration of tight junctions and nutrient transporters, suppression of hepatic lipogenesis, and activation of PPARγ/SIRT1 pathways. 23 These findings position MgT as a regulator of integrated epithelial–metabolic networks. Mechanistically, this profile is supported by the complementary pharmacology of its components: L-theanine is rapidly absorbed, crosses the blood–brain barrier, and exerts neuromodulatory and antioxidant effects, whereas magnesium provides ATP-dependent metabolic support and NMDA receptor regulation, with bioavailability varying by formulation.97,98 Emerging evidence further suggests that MgT, particularly in combination with NR, restores NAD+ metabolism and reinforces metabolic resilience under stress. 24 Available data indicate a favorable but incompletely characterized safety profile. Both L-theanine and magnesium are generally well tolerated, and early human studies of combined formulations report reduced stress and improved sleep without serious adverse effects. 99 However, key translational parameters, including pharmacokinetics of the intact complex, dose equivalence, and long-term safety, remain unresolved, and current human data do not yet allow precise attribution of dose-specific effects to MgT alone. Thus, MgT should be considered a promising mechanistically integrated candidate requiring rigorous dose-ranging and clinical validation.
MgT and the gut–liver axis under CVS
The impact of MgT in the CVS model is most pronounced along the gut–liver axis, where epithelial integrity, nutrient handling, and hepatic metabolic control converge (Figure 4). In a dose-ranging CVS study, Sahin et al. 23 demonstrated that MgT counteracts the hallmark systemic consequences of chronic stress, including weight loss, hypomagnesemia, hypoinsulinemia, hypercorticosteronemia, and dyslipidemia, while restoring intestinal barrier function and attenuating hepatic injury in a dose-dependent manner. At the intestinal level, CVS induced marked architectural disruption, characterized by villus atrophy, crypt alteration, and inflammatory infiltration. MgT largely preserved villus–crypt morphology and reduced mucosal inflammation, effects that were paralleled by molecular restoration of epithelial sealing mechanisms. Stress-induced suppression of tight-junction and mucus-associated proteins (CLDN1, CLDN4, OCLN, TJP1, and MUC2) was reversed by MgT, indicating reinforcement of barrier integrity and permeability control. 23 These epithelial effects are mechanistically relevant, as barrier disruption is a recognized upstream driver of hepatic pathology under stress. Independent work by Jiang et al. 12 showed that CVS promotes hepatic injury through iron dyshomeostasis, characterized by Fe(II) accumulation, oxidative stress, upregulation of TFR1 and ZIP14, and suppression of ferritin and miR-181. When integrated with the reduced lipid accumulation and improved histology observed under MgT, these findings suggest that MgT modulates not only hepatic lipid flux but also redox and metal-handling pathways that amplify stress-related liver injury. Consistent with this interpretation, MgT suppressed CVS-induced activation of lipogenic programs (SREBF1, FASN) while restoring expression of PPARγ and SIRT1, central regulators of fatty acid oxidation, insulin sensitivity, and antioxidant defense. 23 CVS-associated disruption of hepatic NAD+ metabolism, reflected by reduced nicotinamide (NAM), nicotinic acid (NA), and nicotinamide adenine dinucleotide phosphate (NADPH), was partially corrected by MgT, supporting re-engagement of NAD+/SIRT1-dependent metabolic control.
A defining feature of MgT action is its normalization of intestinal nutrient transporter networks. CVS downregulated transporters governing carbohydrate, amino acid, and lipid uptake (SGLT1, GLUT2, PepT1/2, LAT2, EAAT3, FABP2, FATP4), consistent with functional malabsorption. MgT restored their expression, indicating recovery of absorptive capacity alongside repair of the structural barrier. 23 Synergistic effects become evident when MgT is combined with NR. In follow-up studies, MgT + NR robustly replenished hepatic NAD+ and NADPH pools, normalized SIRT1/PPARγ signaling, suppressed lipogenesis, and reduced triglyceride accumulation under CVS. 24 Parallel restoration of intestinal transporter expression further supports a cooperative model in which MgT stabilizes epithelial architecture and transport function, while NR amplifies NAD+-dependent metabolic resilience. Collectively, these data support a two-tier protective framework for MgT in CVS: first, reinforcement of intestinal barrier and absorptive function, limiting antigen translocation and energy imbalance; second, hepatic metabolic reprogramming that shifts stress-driven lipogenesis toward SIRT1/PPARγ-mediated resilience. Within this context, MgT emerges as a trace-element-based, multi-target intervention that acts at the level of epithelial metabolic coupling rather than on isolated downstream pathways. Future studies linking these robust peripheral effects to central neurobiological and behavioral outcomes will be critical for defining MgT’s integrative role across the gut–liver–brain axis. Together, these data demonstrate that MgT restores both structural and functional integrity across the gut–liver axis in a coordinated manner (Figure 4).
MgT and brain/energy metabolism: Potential mechanisms
Although current evidence on MgT in CVS models primarily centers on the gut–liver axis, converging mechanistic pathways strongly suggest that MgT may also influence brain bioenergetics, synaptic function, and stress-related neuroplasticity. These central effects may arise from both direct actions on neuronal substrates and indirect contributions mediated through systemic metabolic improvements. These convergent mechanisms suggest that MgT may support neuronal bioenergetics and synaptic stability under chronic stress conditions (Figure 5).
Mitochondrial bioenergetics and redox homeostasis
Mitochondrial dysfunction represents a central hallmark of CVS-induced neurobiology, reflecting the convergence of persistent allostatic load, impaired cellular energy metabolism, and heightened vulnerability to affective pathology. CVS has been associated with impaired brain energy metabolism, including reduced respiratory chain activity in stress-sensitive regions. In Tagliari et al., 6 CVS inhibited complex II and complex IV activity in the prefrontal cortex, whereas in the hippocampus, only complex IV activity was reduced, accompanied by increased immobility in the forced swim test. Oxidative damage to mitochondrial proteins, membrane lipids, and mtDNA may further compromise mitochondrial integrity and function, thereby amplifying vulnerability to cellular stress. 100 Within this energetic and redox-vulnerable landscape, the MgT exerts convergent protective actions. Magnesium is indispensable for ATP generation, since Mg–ATP constitutes the active substrate for oxidative phosphorylation and kinase-driven metabolic networks; restoring magnesium availability under stress is therefore predicted to stabilize mitochondrial ATP production and support electron transport chain efficiency. L-theanine may complement this response by activating Nrf2-dependent antioxidant signaling, increasing SOD, CAT, and GSH-Px activity, and reducing ROS accumulation, thereby limiting oxidative damage at the cellular level. 19 Notably, MgT and particularly MgT + NR combinations restore hepatic NAD+ and NADPH pools under CVS. 24 Given NAD’s essential role as a cofactor for mitochondrial dehydrogenases and complexes I and III, similar restoration in neural tissue would be expected to enhance electron transfer efficiency, improve redox cycling, and bolster mitochondrial metabolic flexibility. Although direct CNS measurements are not yet available, the integrated effects of magnesium-dependent ATP synthesis, L-theanine-mediated redox defense, and NAD+ replenishment provide a mechanistically coherent rationale for enhanced neuronal bioenergetics under chronic stress.
Neurotransmission, synaptic plasticity, and behavioral regulation
Chronic stress profoundly disrupts CNS homeostasis by reducing neurotrophic support and impairing structural and functional plasticity in limbic regions such as the hippocampus and prefrontal cortex. These alterations contribute to behavioral phenotypes linked to mood and cognition, including anxiety-like behavior, anhedonia-related features, and cognitive impairment. 101 Within this maladaptive neurobiological landscape, the MgT exerts multimodal regulatory effects.
Magnesium acts as a voltage-dependent physiological antagonist of the NMDA receptor, thereby limiting excessive Ca²+ entry and protecting neurons against glutamate-mediated excitotoxicity. 102 L-theanine complements these actions by enhancing GABA levels, modulating AMPA and NMDA receptor dynamics, and elevating serotonin and dopamine levels in limbic and prefrontal circuits. These combined effects contribute to synaptic stabilization and counteract the monoaminergic imbalances induced by CVS.20,68
Evidence from multiple preclinical models further supports L-theanine’s neuromodulatory role. Across chronic stress paradigms, stress exposure is consistently associated with cognitive impairment and anxiety-like behavior. In this context, L-theanine has been reported to attenuate corticosterone elevation, improve cognitive performance, and reverse stress-related catecholaminergic alterations in chronic restraint stress models.74,103 Moreover, prophylactic intake of 0.3% L-theanine in drinking water significantly attenuated corticosterone responses to acute water-immersion stress in young rats, suggesting a stress-buffering effect. 104 Consistently, in a chronic restraint stress model, L-theanine reduced serum corticosterone levels, improved cognitive performance, and reversed stress-related reductions in noradrenaline and dopamine in the serum, hippocampus, and cerebral cortex. 74 L-theanine also demonstrates antioxidant, and neuroprotective properties in models of ischemia–reperfusion injury and toxin-induced neuronal damage.70,105 In the context of MgT, these mechanistic and behavioral findings indicate that L-theanine not only augments the NMDA-blocking and ATP-stabilizing effects of magnesium but also adds a critical layer of neuroendocrine and neurotransmitter regulation. Through this synergistic interplay, MgT may help restore synaptic plasticity within the hippocampus and prefrontal cortex, regions particularly vulnerable to stress-induced reductions in dendritic architecture, neurogenesis, and adaptive behavioral flexibility.
NAD+/SIRT1, PPARγ, and neuroprotective signaling
NAD+ functions as a metabolic integrator linking redox state, mitochondrial function, and transcriptional stress adaptations. CVS diminishes NAD+ and NADPH availability, suppressing the activity of NAD+-dependent deacetylases such as SIRT1, which orchestrate mitochondrial biogenesis, antioxidant defense, and synaptic remodeling.106,107 MgT + NR co-treatment restores NAD+, NADPH, NAM, and NA levels and robustly upregulates hepatic SIRT1 and PPARγ expression under CVS. 23 Given the abundant expression of SIRT1 in hippocampal and cortical neurons, these peripheral findings strongly suggest analogous CNS effects. Increased SIRT1 activity would be expected to promote mitochondrial turnover, enhance resistance to oxidative stress, and suppress NF-κB-mediated neuroinflammation. PPARγ activation further complements these actions by modulating lipid signaling, anti-inflammatory gene expression, and metabolic homeostasis. Thus, MgT, either alone or combined with NR, may activate a neuroprotective signaling triad that shifts neural circuits from a stress-vulnerable to a stress-resilient state. Definitive studies quantifying brain NAD+ flux, SIRT1 activity, and mitochondrial biogenesis following MgT supplementation represent an important frontier for future work.
Integration with the gut–liver axis and systemic metabolism
The central effects of MgT are likely reinforced by improvements along the gut–liver axis, which transmits powerful bottom-up signals shaping neural energy metabolism and stress responsivity. CVS induces hepatic iron dysregulation and oxidative injury, characterized by Fe(II) accumulation, upregulation of TFR1 and ZIP14, and suppression of ferritin-related iron buffering. 12 MgT restores tight-junction integrity, replenishes nutrient transporters, reduces hepatic lipid accumulation, and reverses NAD+/SIRT1 metabolic collapse.23,24 These improvements reduce the peripheral inflammatory and metabolic stress burden that feeds forward into the brain, thereby attenuating systemic drivers of mitochondrial dysfunction, synaptic deficits, and behavioral pathology. Viewed as an integrated whole, MgT supports neural function not only through direct neuromodulation and redox control but also by recalibrating the metabolic and inflammatory environment originating in the gut and liver.
MgT, NR, and other interventions: Comparative and combined approaches
MgT versus classical antioxidant strategies
Conventional antioxidant vitamins, most notably vitamins C and E, attenuate selected features of CVS pathology by directly scavenging reactive oxygen species and limiting lipid peroxidation. In preclinical stress models, these agents reduce oxidative damage and partially normalize redox biomarkers; however, their biological reach remains largely confined to downstream redox buffering.6,8 Importantly, chronic stress pathology is governed not only by downstream oxidative imbalance but also by upstream regulatory systems, including HPA axis dynamics, reward-related monoaminergic circuitry, and NAD+-dependent metabolic control, which together shape allostatic load.106 –109 By contrast, the MgT engages multiple hierarchical layers of stress regulation. Magnesium supports ATP-dependent metabolism, constrains NMDA receptor-mediated excitotoxicity, and dampens endocrine stress signaling, whereas L-theanine modulates GABAergic, glutamatergic, and monoaminergic tone while activating antioxidant and cytoprotective pathways.19,68 Through this convergence, MgT attenuates HPA axis hyperactivation, normalizes neurotransmission, and restores redox and metabolic homeostasis across intestinal and hepatic tissues. Notably, MgT also reactivates NAD+/SIRT1–PPARγ signaling, positioning it upstream of mitochondrial function, lipid handling, and inflammatory control.23,24 Thus, MgT transcends the narrow antioxidant paradigm by integrating endocrine, neural, metabolic, and redox axes into a coordinated anti-stress response.
Synergy between MgT and NR
The rationale for combining MgT with NR arises from their complementary engagement of distinct yet convergent layers of stress biology. MgT primarily stabilizes upstream stress-sensitive systems, including intestinal barrier integrity, nutrient transporter networks, neurotransmission, and redox balance, as evidenced by its ability to restore tight junction proteins, mucosal components, and metabolic regulators such as PPARγ and SIRT1 in CVS models. 23 In contrast, NR acts predominantly at the level of cellular bioenergetics by replenishing NAD+ pools and restoring NAD+-dependent metabolic pathways that are disrupted under chronic stress conditions. 24 This mechanistic complementarity provides a basis for functional synergy. In CVS models, combined MgT and NR supplementation restores hepatic NAD+, NADPH, NAM, and NA levels, enhances SIRT1 and PPARγ signaling, suppresses lipogenic transcription factors (SREBF1, FASN), and improves intestinal barrier integrity and nutrient transporter expression. 24 These findings indicate that MgT and NR operate across different hierarchical levels: Mg-T attenuates upstream drivers of stress pathology, including epithelial disruption, oxidative stress, and neuroendocrine imbalance, whereas NR amplifies downstream metabolic resilience through NAD+-dependent reprogramming. Notably, MgT’s ability to partially restore NAD+-linked pathways on its own further supports convergence at this metabolic axis. 23 Within this framework, MgT can be conceptualized as a systems-level stabilizer of gut–liver–brain homeostasis, while NR functions as a metabolic amplifier that extends and deepens this protective response. Accordingly, their combination may offer broader therapeutic coverage than either intervention alone, particularly in chronic stress states characterized by concurrent epithelial dysfunction, hepatic metabolic injury, and impaired redox adaptation. This systems-level positioning distinguishes MgT from single-target interventions and supports its integration with NAD+-directed strategies (Figure 6).
Comparison with exercise and lifestyle interventions
Physical exercise exerts broad non-pharmacological benefits by improving autonomic balance, vascular function, and inflammatory regulation, with additional evidence suggesting benefits for neuroplasticity-related processes. 16 Exercise primarily exerts top-down effects on central neuroplasticity and autonomic regulation, whereas MgT predominantly acts through bottom-up stabilization of micronutrient status, neurotransmission, gut–liver integrity, and NAD+-dependent metabolic pathways. An integrated intervention combining exercise with MgT therefore holds strong potential for additive or synergistic benefit, simultaneously targeting central stress circuitry and peripheral metabolic resilience. Comparative studies evaluating exercise alone, MgT alone, and combined regimens using behavioral, autonomic, and multi-omics endpoints represent a critical translational priority for defining optimal anti-stress strategies.
Integration with phytochemicals and microbiota-targeted therapies
A growing body of evidence indicates that phytochemicals such as quercetin, curcumin, and resveratrol activate Nrf2–ARE signaling, suppress NF-κB-mediated inflammation, and enhance mitochondrial biogenesis, thereby intersecting with core pathways disrupted by chronic stress.110,111 Many dietary polyphenols can remodel gut microbial communities, favor short-chain fatty acid-related microbial functions, strengthen epithelial barrier integrity, and may reduce gut-derived inflammatory burden.112,113
Microbiota-targeted approaches, including probiotics, prebiotics, and synbiotics, further reinforce tight-junction networks, attenuate LPS-driven hepatic inflammation, and modulate neuronal and autonomic signaling via the gut–brain axis. 114 Given that MgT already improves barrier integrity, nutrient transporter expression, and hepatic redox metabolism, its combination with polyphenols or microbiota-directed therapies is mechanistically well-justified. Expected points of convergence include Nrf2 activation, NF-κB inhibition, improved mitochondrial dynamics, and normalization of microbial ecology.
Green tea provides a paradigmatic example of this convergence. Its bioactive matrix, comprising catechins such as epigallocatechin-3-gallate, caffeine, and L-theanine, exerts antioxidant, anti-inflammatory, hypolipidemic, and neuromodulatory effects across species. 70 In livestock systems, green tea and L-theanine supplementation improve growth performance, antioxidant capacity, and immune competence, while reducing lipid peroxidation and microbial spoilage in animal products, supporting the concept of functional feeds that modulate host stress biology.115 –117 At the gut level, L-theanine and green tea polyphenols reshape microbial communities and enhance mucosal barrier function, although microbiota responses are dose- and matrix-dependent, underscoring the need for optimized formulations.69,79
Clinical and translational implications
Accumulating data from preclinical stress models, supported by controlled human supplementation studies, indicate that the MgT has substantial translational relevance for stress-related physiological and neuropsychiatric disorders.21,68,109 Unlike single-target interventions, MgT engages multiple stress-responsive systems in parallel, including epithelial barrier integrity, redox and metabolic reprogramming, neuroendocrine regulation, and central neurotransmission. This multimodal profile closely aligns with disease states in which chronic stress operates as a causal or amplifying factor rather than a secondary epiphenomenon. 118 Stress-sensitive gastrointestinal disorders emerge as particularly compelling clinical targets. Experimental and translational evidence indicates that both magnesium and L-theanine enhance tight-junction architecture, modulate mucosal immune tone, and reshape microbial composition, thereby limiting stress-induced barrier disruption and endotoxemia along the gut–liver–brain axis. 114 These mechanisms support the potential utility of MgT in conditions such as irritable bowel syndrome, functional dyspepsia, and syndromes characterized by increased intestinal permeability.
Beyond the gut, MgT is mechanistically positioned to address metabolic pathologies driven by chronic glucocorticoid exposure, mitochondrial dysfunction, and oxidative imbalance. Restoration of NAD+-dependent metabolic pathways, improvement of redox homeostasis, and attenuation of hepatic lipid accumulation link MgT to key molecular features of metabolic syndrome and non-alcoholic fatty liver disease.100,119,120 In parallel, coordinated modulation of GABAergic, glutamatergic, and monoaminergic signaling provides a neurobiological basis for MgT’s anxiolytic, antidepressant, and sleep-promoting effects in stress-related affective disorders and hyperarousal-associated insomnia. 20 Despite this strong mechanistic rationale, important translational questions remain unresolved. Human dose–response relationships for MgT have not been systematically defined, and its pharmacokinetics may differ from those of conventional magnesium salts, which often exhibit limited bioavailability and poor CNS penetration. 121 Moreover, long-term safety and efficacy data for combined MgT and NAD+-boosting strategies, such as NR, are still limited despite robust preclinical justification.25,106
Future clinical translation will require integrated biomarker strategies capable of capturing MgT’s system-level activity. Endpoints should encompass endocrine stress markers, inflammatory and oxidative indices, NAD+-related metabolites, and gut barrier–microbiota readouts, complemented by neuroimaging, psychometric assessments, and autonomic measures such as heart rate variability. 122 Well-designed randomized, placebo-controlled trials in stress-associated gastrointestinal, metabolic, and occupational stress populations will be essential to establish therapeutic efficacy and inform regulatory positioning. Within emerging precision-medicine frameworks, MgT may ultimately be deployed as a modular intervention tailored to individual stress biology, metabolic resilience, and microbiome configuration. In this context, MgT represents not simply a micronutrient combination but a prototype of next-generation, systems-oriented strategies aimed at recalibrating stress physiology across interconnected organ networks. These findings support a translational framework in which MgT can be advanced from preclinical validation to targeted clinical application (Figure 7).

Translational pathway from preclinical findings to clinical application of MgT. Preclinical CVS models demonstrate that MgT, alone or in combination with NR, protects the gut–liver axis, normalizes metabolic and hormonal parameters, and improves stress-related outcomes. Mechanistic insights highlight NAD+/SIRT1 and PPARγ pathways, modulation of iron handling (ZIP14/miR-181), preservation of tight junctions and restoration of nutrient transporters. These findings support MgT as a candidate intervention for stress-related gastrointestinal disorders, NAFLD and metabolic syndrome, anxiety and depressive disorders with somatic symptoms, and stress-related sleep disturbances. Future randomized controlled trials should incorporate mechanistic biomarkers and clinically meaningful endpoints to establish optimal dosing, safety, and efficacy.
Conclusion and future directions
Accumulating preclinical evidence indicates that MgT is a mechanistically integrated, multi-target intervention capable of counteracting core features of CVS. Across experimental models, MgT preserves intestinal barrier integrity, restores nutrient transporter expression, and attenuates hepatic steatosis and oxidative stress, thereby stabilizing key components of the gut–liver axis, which are highly vulnerable to chronic stress.118,120 These tissue-level effects converge on reactivation of central metabolic regulators, including the NAD+/SIRT1 axis and PPARγ, which coordinate mitochondrial efficiency, lipid oxidation, and anti-inflammatory signaling. 106 Notably, co-administration of MgT with NR further amplifies these responses by expanding the NAD+ and NADPH pools, suppressing lipogenic transcriptional programs, and enhancing intestinal absorptive capacity, highlighting a synergistic strategy to reinforce metabolic resilience under stress.23,25 Through its combined actions on neuromodulation, redox homeostasis, and metabolic control, MgT occupies a unique position at the interface of trace-element biology and systems-level stress regulation.68,109 Future research should integrate mechanistic, clinical, and systems-level approaches to define the therapeutic scope of MgT across stress-related disorders (Figure 8). Priorities include brain-specific mechanistic interrogation, rigorous dose–response optimization, and long-term safety profiling, alongside well-powered clinical trials in stress-associated gastrointestinal, metabolic, and neuropsychiatric conditions. Within this framework, MgT emerges not merely as a nutraceutical combination, but as a prototype for next-generation, integrative interventions targeting the gut–liver–brain axis in chronic stress biology.

Proposed research roadmap for MgT in stress-related disorders. Future work should progress along four major levels. (1) Basic mechanistic studies are needed to examine MgT effects on NAD+/SIRT1 signaling in the brain, ferroptosis and iron homeostasis in the liver and multi-omics alterations (transcriptome, metabolome, microbiome) along the gut–liver–brain axis. (2) Refined animal experiments should define dose–response relationships, sex- and age-specific effects, long-term safety, and synergistic combinations with exercise, polyphenols, and microbiota-directed interventions. (3) Early-phase clinical trials should evaluate MgT and MgT + NR in well-defined patient populations (e.g., IBS with stress, NAFLD with metabolic syndrome, stress-related anxiety, and depression), using mechanistic biomarkers and clinically relevant outcomes. (4) Ultimately, implementation and personalization strategies may stratify patients by baseline NAD+ status, microbiota profiles, and stress load, integrating MgT into multimodal therapeutic programs alongside lifestyle and standard pharmacotherapies.