
Abstract
Objective
During and after menopause, the symptoms of vaginal atrophy cause great discomfort and necessitate effective treatment options. Currently, vaginally applied oxytocin is being investigated as a treatment for the symptoms of vaginal atrophy in postmenopausal women. To clarify the mechanisms behind oxytocins effects on vaginal atrophy, the present study investigated the effects of oxytocin on cell proliferation in the cells of the Vk2E6E7 line, a non-tumour vaginal cell line. The study also compared the effects of oxytocin with those of estradiol (E2).
Study design
The effects of both oxytocin and E2 on the proliferation of Vk2E6E7 cells were investigated using Cell Proliferation ELISA BrdU Colorimetric Assay. The expression of both oxytocin and oxytocin receptor was studied in Vk2E6E7 cells using quantitative real-time polymerase chain reaction and immunofluorescent staining.
Main outcome measures
Cell proliferation and gene expression.
Results
Oxytocin increased cell proliferation both time dependently and dose dependently. This differed from the effect pattern observed in cells treated with E2. In addition, in oxytocin-treated cells, the oxytocin receptor was found to be co-localized with caveolin-1, indicating pro-proliferative signalling within the cell.
Conclusions
Oxytocin stimulates cell proliferation and the co-localization of oxytocin receptor with caveolin-1 in oxytocin-treated cells, supporting the role of oxytocin signalling in cell proliferation. In addition, these findings suggest that increased cell proliferation is one mechanism by which local vaginal oxytocin treatment increases vaginal thickness and relieves vaginal symptoms in postmenopausal women with vaginal atrophy.
Introduction
Symptoms of vaginal dryness, itching, irritation and dyspareunia are very common during and after menopause. 1 The vaginal mucosa consists of a non-keratinized, stratified, squamous epithelium that undergoes constant renewal through cell proliferation, differentiation and maturation. 2 All these effects are regulated by estrogens 3 and therefore low levels of circulating estrogen lead to insufficient cell renewal and maturation.4–6 The vaginal mucosa becomes atrophic and is characterized by being thin, pale, dry and occasionally inflamed. Objectively, the mucosa has a higher vaginal pH and a lower maturation index, indicating few mature, superficial and intermediate cells and more immature, para-basal cells in the mucosa’s epithelial layer. 4 With the exception of estrogenic compounds, effective treatment for severe symptoms is still lacking, 4 in spite of continued hesitation about using estrogenic treatments on postmenopausal women. 1
Clinical trials using vaginal oxytocin (OXT) have shown promising results for relieving vaginal symptoms caused by menopause.7–9 OXT has been shown to decrease vaginal pH, to increase both the number of superficial epithelial cells7,8 and vaginal thickness and to improve the histological scores of the vaginal mucosa.7–9
OXT is a nona-peptide that is known to be very important during birth and lactation and to influence reproductive behavior.10,11 OXT is expressed in many peripheral tissues, 12 including skin. 13 In keratinocytes from skin biopsies, OXT affects cell proliferation in a dose-dependent manner, 14 and it has been shown to stimulate wound healing. 15
OXT mediates its effect through the OXT receptor (OXTR), a G-protein-coupled receptor in the cell membrane. OXT can either stimulate or inhibit cell proliferation, depending on whether the OXTR is located within or outside of a caveolin-enriched micro-domain on the cell membrane.16,17 Caveolins are scaffolding proteins that interact with G-protein sub-units to regulate cell signaling.18,19 In addition to the cellular localization of the OXTR, cell proliferation is affected by OXT concentration and the cellular distribution of various G-proteins. 10 OXTR is present in most reproductive organs, 12 and its expression is sometimes regulated by estrogens. 20 OXTR has also been shown by the Human Protein Atlas to be expressed in the vaginal mucosa. 21
The aim of the present study was to investigate the effect of OXT on the proliferation of cells in the Vk2E6E7 line of non-tumour cells established from normal vaginal mucosa. 22 The study also compared OXT-treated cells with those treated with estradiol (E2).
Materials and methods
Cell culture
Vk2E6E7 cells (catalogue no. ATCC CRL-2616; LGC Standards GmbH) were cultured according to manufacturer’s instructions. Keratinocyte serum-free medium (KSFM) supplemented with 0.1 ng/ml of recombinant epidermal growth factor (EGF), 0.05 mg of bovine pituitary extract (catalogue no. 17005-075; Thermo Fisher Scientific) and 44 mg/L of calcium chloride (catalogue no. 1.02382.0500; Merck Millipore) was used, and the cells were cultured at 37℃ in 5% CO2.
Vk2E6E7 cells were exposed to either OXT (Grindex, Latvia) or E2 (catalogue no. E2758; Sigma-Aldrich AB, Sweden) for 24, 48 or 72 h. OXT was dissolved in H2O, and stock solutions were prepared in H2O and diluted in supplemented KSFM to final concentrations of 0.01 nM, 0.1 nM, 1 nM, 10 nM, 100 nM, 1 μM and 10 µM of OXT in KSFM. E2 stock solutions were prepared in ethanol and diluted in KSFM to final concentrations of 0.1, 1, 10, 100 and 1000 nM of E2 in KSFM. Water or ethanol was used as a vehicle/control, and the final H2O or ethanol concentration in all treatment groups was 0.1% (v/v) in KSFM.
Quantitative real-time polymerase chain reaction
The expression of OXTR and OXT in Vk2E6E7 cells grown in supplemented KSFM was investigated using quantitative real-time polymerase chain reaction (qPCR). Total RNA was extracted from Vk2E6E7 cells using the RNeasy Mini Kit (Qiagen; Germany), and cDNA was synthesized using the SuperScript VILO Master Mix (Thermo Fisher Scientific). A total of 10 ng cDNA/wells were analysed in triplicate. TaqMan Gene Expression Assays for OXTR (catalogue no. Hs00168573_ml; Thermo Fisher Scientific) and OXT (catalogue no. Hs00792417_g1; Thermo Fisher Scientific) and the endogenous control actin B (catalogue no. 4326315E; Thermo Fisher Scientific) were used together with TaqMan Fast Universal Master Mix (catalogue no. 4352042; Thermo Fisher Scientific) as previously described. 6
Vk2E6E7 cell proliferation
The proliferation of Vk2E6E7 cells was assessed using Cell Proliferation ELISA BrdU Colorimetric Assay (catalogue no. 11647229001; Sigma-Aldrich) according to the manufacturer’s instructions. At sub-confluency, the cells were sub-cultivated to sterile 96-well plates. Cell concentration was 50,000 cells/ml, and each well was seeded with 100 µl of cells in KSFM. The absorbance was measured at 450 nm and was directly correlated with the number of proliferative cells.
Immunofluorescent staining of OXTR and caveolin-1 in Vk2E6E7 cells
To further understand the effect of OXT on the proliferation of Vk2E6E7 cells, immunofluorescent staining of OXTR and caveolin-1 (CAV1) was performed. Cells were cultured on four-well chamber slides in supplemented KSFM or treated with vehicle or OXT 10 nM. Then, the cells were rinsed with phosphate buffered saline (PBS) before being fixated by immersion in 4% paraformaldehyde for 15 min followed by repeated washings in PBS. Then, the cells were permeabilized with cooled methanol for 5 min followed by repeated washings with PBS. A blocking solution consisting of 1% bovine serum albumin in PBS with 0.1% Tween was used to block unspecific binding, to incubate the antibodies and to repeatedly wash between incubations. Primary antibodies for OXTR (catalogue no. ab87312; Abcam, Cambridge, UK) and CAV1 (catalogue no. ab2910; Abcam) were diluted 1:200 and 1:750, respectively, and the secondary antibodies were donkey anti-Goat Alexa Fluor 594 conjugate (catalogue no. A11058; Thermo Fisher Scientific) and donkey anti-Rabbit Alexa Fluor 488 conjugate (catalogue no. A21206; Thermo Fisher Scientific), both diluted 1:100. The slides were mounted using VECTASHIELD Antifade Mounting Medium with DAPI (catalogue no. H-1200; Vector Laboratories) and observed using a fluorescent microscope (Axio Observer Z1; Carl Zeiss AG, Germany).
Statistical analysis
The overall effect of treatment on cell proliferation was analysed using a non-parametric Kruskal–Wallis test, and treatment concentrations were compared with the vehicle/control using a non-parametric Mann–Whitney U-test. Non-parametric correlations were tested according to Spearman’s rank correlation. A p value < 0.05 was considered significant. Statistical analysis was performed using the IBM SPSS Statistics 20 software program (IBM, Chicago, IL, USA).
Results
OXTR and OXT gene expression in Vk2E6E7 cells
Expression of OXTR mRNA was detected, with the median Ct-value being 30.5 (30.2–30.8), and in relation to the endogenous control, actin B, the median Ct-value was 24.0 (23.2–24.8). In contrast, the expression of OXT was low or undetectable, with the median Ct-value being 37.9 (37.2–undetectable).
Proliferation of OXT-treated cells
Cells treated with 1 nM to 10 µM of OXT showed increased proliferation after a 24-h incubation period. After 72 h of incubation, cells treated with 0.01 nM to 1 µM of OXT also showed increased proliferation compared with the vehicle. However, in cells treated for 48 h, only 0.01 nM of OXT increased the proliferation, whilst 10 µM of OXT inhibited proliferation (Figure 1(a) to (c)). The increased proliferation of cells treated with OXT for 24 h was dose dependent, with proliferation correlating positively with the increased concentrations of OXT (r = 0.459, p < 0.001). However, there was a negative correlation between OXT concentration and cell proliferation in the cells incubated for 48 h (r = −0.438, p < 0.01). There was no correlation between OXT concentration and cell proliferation demonstrated in the cells treated for 72 h.
Cell proliferation in Vk2E6E7 cells measured using Cell Proliferation ELISA BrdU colorimetric assay, which directly correlates cell proliferation to the absorbance measured. Cells were incubated in either OXT (a, b, and c) or E2 (d, e, and f) for 24, 48, or 72 h, respectively using six wells per treatment. Significant differences are indicated as follows: *p < 0.05, **p < 0.01, and ***p < 0.001. Data were analysed using non-parametric Kruskal–Wallis and Mann–Whitney U-tests.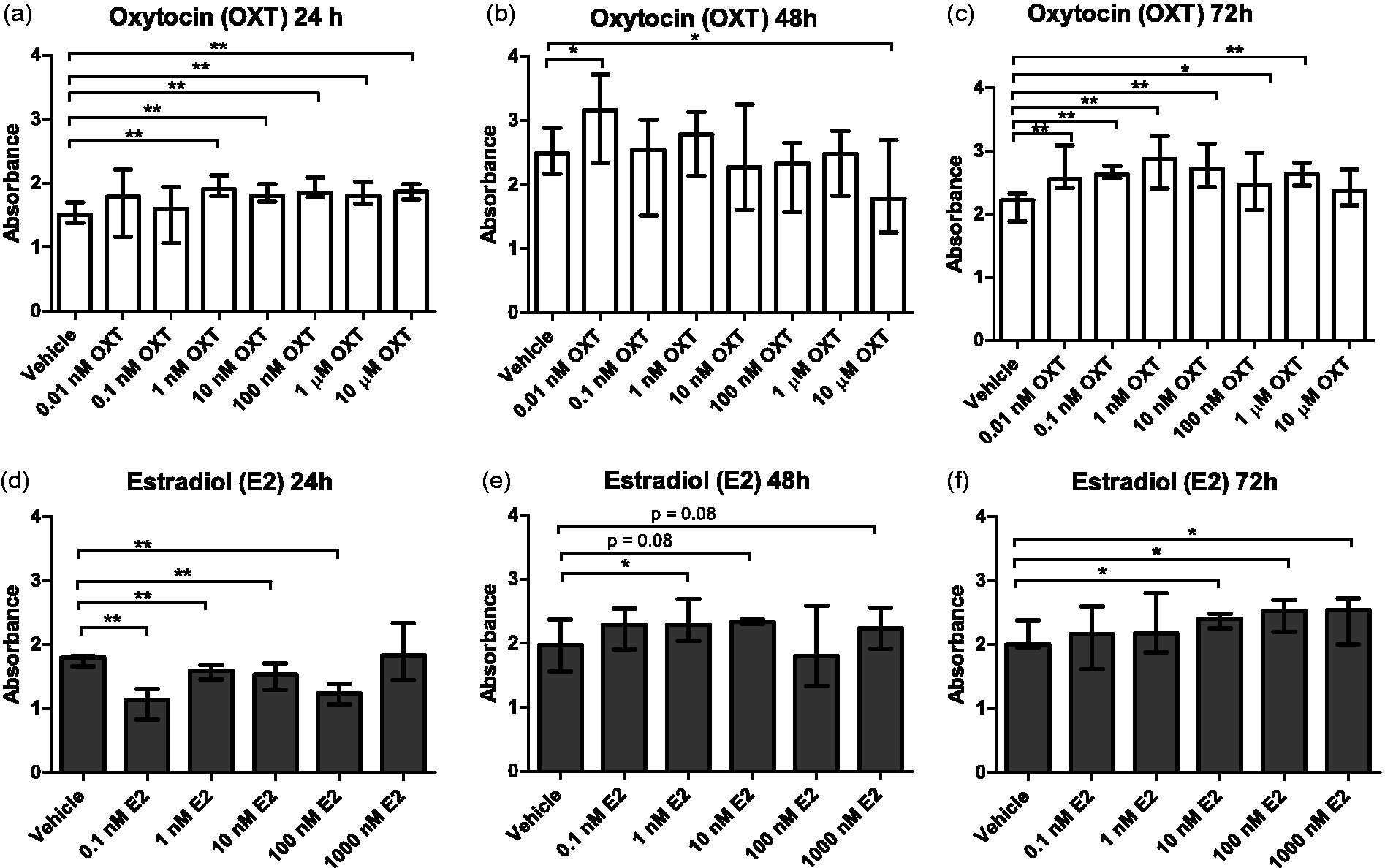
Proliferation of E2-treated cells
Cells treated with E2 showed a decreased rate of proliferation compared with vehicle-treated cells after 24 h of incubation. However, this effect was reversed after 48 and 72 h of incubation, with E2-treated cells showing increased cell proliferation compared with vehicle-treated cells (Figure 1(d) to (e)). However, a dose-dependent increase in proliferation was observed only in the cells incubated for 72 h, in which increased proliferation could be correlated with E2 concentration (r = 0.498, p < 0.01).
Immunofluorescent staining of OXTR and CAV1 in Vk2E6E7 cells
In Vk2E6E7 cells, the OXTR was localized mainly to the cell nucleus and the plasma membrane in the vicinity of the nucleus (Figure 2). To investigate the co-localization between OXTR and CAV1 in caveolin-enriched micro-domains, the cells were simultaneously double-stained for CAV1. In control experiments in which Vk2E6E7 cells were cultured in either supplemented KSFM or vehicle-treated supplemented KSFM, CAV1 was found to cover the entire cytoplasm and was not co-localized with OXTR. In contrast, in cells treated with 10 nM of OXT for 15 min, CAV1 was localized in the plasma membrane in the vicinity of the nucleus and was co-localized with OXTR. This co-localization occurred in a large proportion of cells, but not all of them. Localization of CAV1 close to the nucleus did not persist over time, and cells treated for 24 h showed localization of CAV1 similar to that found in controls and no co-localization with OXTR.
Immunofluorescent staining of Vk2E6E7 cells presented first as an overlay and then with specific antibodies for OXTR (red), CAV1 (green), and nuclear-specific DAPI (blue). Cells were treated with either vehicle or 10 nM of OXT for 15 min or 24 h.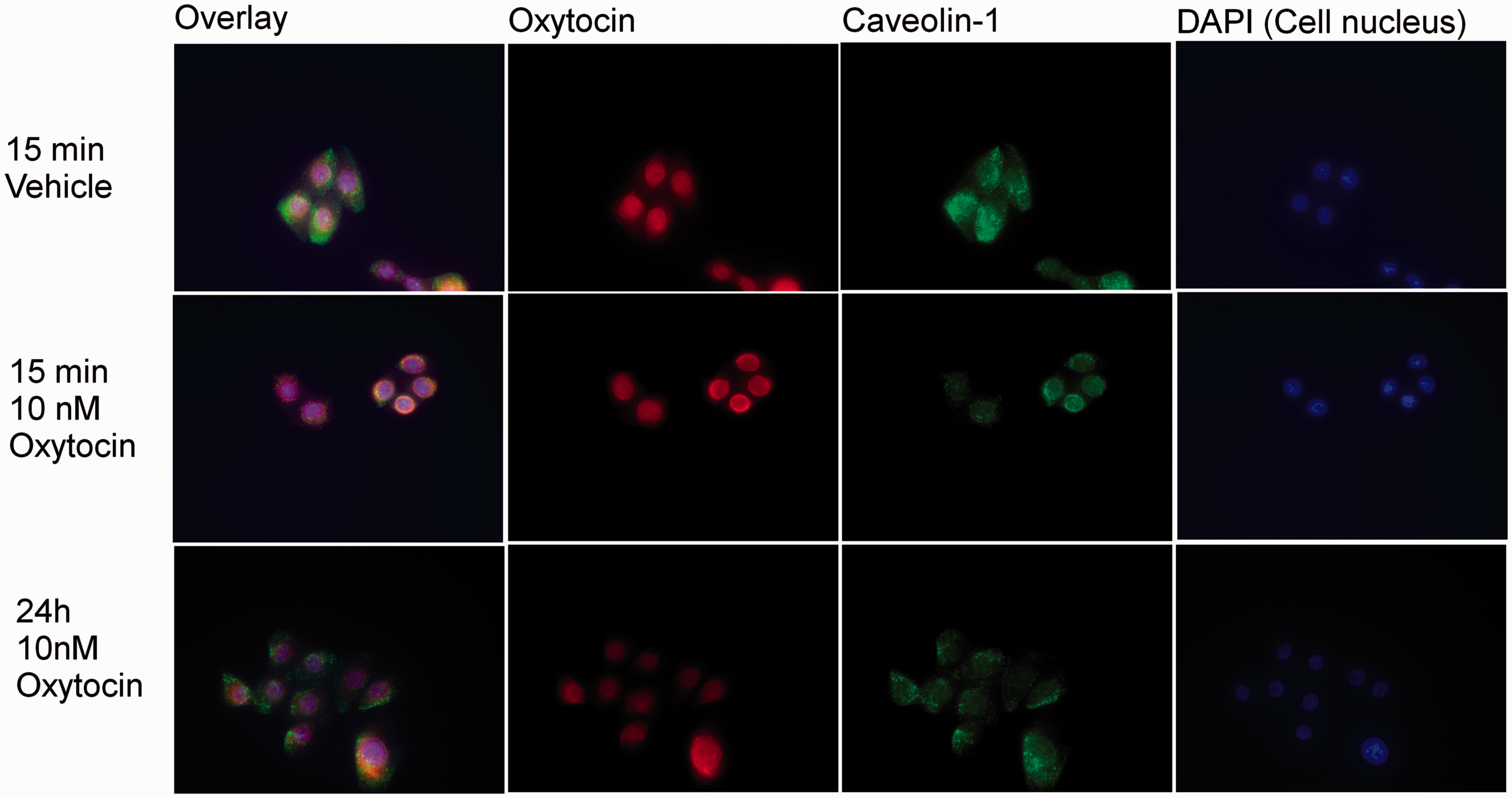
Discussion
The aim of this study was to investigate the effects of OXT on Vk2E6E7 cell proliferation. Treatment with OXT increased Vk2E6E7 cell proliferation both time dependently and dose dependently. The presence of OXTR in the Vk2E6E7 cells was verified by both qPCR and immunofluorescent staining. The co-localization of OXTR with CAV1 in OXT-treated cells, which indicated pro-proliferation signalling by the OXTR, strengthened the results obtained by proliferation analysis. In addition, the present study compared the proliferative effects of OXT and E2, both of which influenced Vk2E6E7 cell proliferation. However, the pattern of the effect differed between the two test substances, both in dose dependence and time dependence.
The thin, pale and dry vaginal mucosa observed clinically in women suffering from symptoms of vaginal atrophy is associated with a decreased number of epithelial-cell layers. 4 In addition, a decreased rate of vaginal-cell proliferation has been shown to correlate with clinical assessment of vaginal atrophy and increased vaginal pH. 5 Therefore, stimulating vaginal-cell proliferation with new treatment options for vaginal atrophy is of great clinical interest for postmenopausal women. OXT treatment has been shown to increase mucosal thickness,7–9 to increase the percentage of superficial cells and to decrease the percentage of para-basal cells,7,8 indicating a positive effect of OXT on vaginal-cell proliferation.7–9
In the present study, proliferation analysis showed a dose-dependent increase in proliferation in Vk2E6E7 cells treated with OXT and incubated for 24 h. However, in cells incubated for 48 h, only those treated with 0.01 nM of OXT showed increased proliferation. In addition, after 15 min of incubation with 10 nM of OXT, OXTR and CAV1 were co-localized, which is consistent with the increase in cell proliferation observed after 24 h of incubation with OXT and which may signify pro-proliferative signalling. This signalling may occur through activation of genes important for cell proliferation by translocation of CAV1 via the activation of OXTR and its transport to the vicinity of the nucleus. 19 The lack of co-localization between OXTR and CAV1 after 24 h of incubation and the lack of effect on cell proliferation after 48 h of incubation might be signs of desensitization of the OXTR 12 or possibly of a transition to an inhibitory effect of OXTR.16,17 After 48 h of incubation, only cells incubated with 0.01 nM of OXT had higher proliferation than the vehicle-treated cells, which may indicate either a dose-dependent desensitization or a transition to an inhibitory effect. The increased cell proliferation observed after 72 h of incubation might be due to a resensitization of the OXTR, but the mechanism for this type of effect is not known 12 and was not analysed further in the present study.
OXT can activate, inhibit or have no effect on cell proliferation. 10 In addition, OXT’s effect on cell proliferation differs among target cells and tissues, depending on the localization of the OXTR, 10 which in caveolin-enriched micro-domains is linked to activation of a proliferative response. Hence, co-localization of OXTR and CAV1 are of great interest when studying the effects of OXT on cell proliferation.
Immunofluorescent staining of the OXTR in Vk2E6E7 cells showed the presence of OXTR in the nucleus or on the plasma membrane in the vicinity of the nucleus. Nuclear localization of OXTR has been observed in other cell types and has been suggested as being involved in regulating gene-specific expression. 23 Immunostaining of CAV1 showed OXT-dependent localization. In untreated cells, the CAV1 was spread out over the entire plasma membrane, whilst in OXT-treated cells, the CAV1 was localized in the vicinity of the nucleus and occasionally co-localized with the OXTR. The localization of CAV1 in the vicinity of the nucleus has been described previously 19 and might be important for OXTR regulation of gene expression and cell proliferation. 19 In addition, the co-localization of OXTR with CAV1 in the vicinity of the nucleus might be a marker for the intracellular components needed for signalling pathways to induce cell proliferation. However, co-localization was not found in all cells, which may explain the moderate but still significant increase in proliferation by OXT treatment. The explanation for the inconsistent co-localization between OXTR and CAV1 can only be speculated upon but may depend on varying OXT concentrations affecting the interaction between OXTR and CAV1 and also by the production of CAV1. 24 Incubation time may also affect co-localization, with a longer incubation period may cause desensitization of the OXTR and uncoupling from the CAV1. 12
To the best of our knowledge, there are no published studies investigating the effect of OXT on vaginal-cell proliferation. However, previous work (Uvnäs-Moberg and Sjögren, manuscript to be submitted) has shown increased cell proliferation and a distinct dose–response relationship in primary vaginal epithelial cells treated with 1 nM to 1 µM of OXT, with a clear maximal effect at 1 µM of OXT and an inhibitory effect at a dose of 10 µM of OXT. These data might reflect the effects of OXT in vivo. In other cell types, the effect of OXT on cell proliferation differs10,14,24–26 because it depends on the localization of the OXTR on the plasma membrane,16,17 on the OXT concentration and on the type of G-proteins within the cell. 10 For instance, although the present study found that OXT increased proliferation in vaginal cells, OXT inhibits proliferation in the epidermis. This difference is unexpected, since the epidermis and vaginal mucosa are very similar in many aspects. 2 The differences in results between the present study and the prior one cited might reflect the diversity of OXTR localization in differing cell types. In addition, differences in the incubation times and dosages used 14 might have caused the differing results. Furthermore, the differences may reflect different experimental conditions and age-dependent changes on cellular properties. OXT may not have the same positive effect on proliferation and growth in cell lines derived from young women as in those derived from menopausal women, in whom the spontaneous growth rate of vaginal epithelial cells is reduced due to lack of estrogen, which is associated with low expression of genes important for cell proliferation in vaginal mucosa. 6
Vaginal-cell proliferation is estrogen dependent,3,5,27 but the exact mechanism by which estrogen exerts its proliferative effects is not known, although it most likely involves several signalling pathways. 2 The present study found differences between OXT and E2 that involved both incubation time and the size of the effect on cell proliferation. This indicates a difference in signalling pathways for OXT, with its membrane-bound, G-protein-coupled receptor, 12 and E2, which acts via the nuclear estrogen receptor. 28
As an in vitro model for proliferation of vaginal epithelial cells, the Vk2E6E7 cell line has some limitations that should be considered. First, the properties of these cells may differ from those of primary cells, and the role of stromal cells cannot be studied in this in vitro model. The present study observed the co-localization of OXTR and CAV1 in Vk2E6E7 cells treated with OXT, which supported the finding of increased proliferation in Vk2E6E7 cells. However, it should be noted that caveolins, which are scaffolding proteins important for cellular signalling, 18 and their role in OXT signalling depend on interactions with cavins, also known as PTRF, 18 which the present study did not investigate. The increase in proliferation observed in Vk2E6E7 cells was not as prominent and the dose–response curve was not as distinct as those observed with primary vaginal cells (Uvnäs-Moberg and Sjögren, manuscript to be submitted), which may reflect different properties in primary vaginal cells from those in a cell line of vaginal epithelial cells. In future studies, it might be of interest to investigate the effect of OXT on cell proliferation in primary cells from both young and postmenopausal women, as their responsiveness to OXT might differ, thus affecting the results.
OXT signalling has been shown to transactivate the EGF receptor as part of the chain of events leading to cell proliferation.17,29 Therefore, adding EGF to the growth medium might affect the results of OXT treatment, perhaps by inducing a slight background effect or by facilitating the effect at lower doses of OXT. However, in the present study, all cells, irrespective of whether they were treated with the vehicle, OXT or E2, were incubated with supplemented KSFM that included EGF. Therefore, the effect of EGF should be negligible.
Overall, Vk2E6E7 cells offer an interesting in vitro model of vaginal epithelial cells. 22 However, the mechanism for vaginal-cell proliferation is complex, and much data remain to be obtained. In addition, transferring information obtained from in vitro studies to the in vivo situation should be done with caution.
Conclusions
OXT increased the proliferation of Vk2E6E7 cells in a time-dependent and dose-dependent manner. The expression of OXTR in Vk2E6E7 cells was shown using both qPCR and immunofluorescent staining. The co-localization of OXTR with CAV1 in OXT-treated cells both supports the assumption that OXT-induced stimulation of cell proliferation is induced via activation of OXT signalling and supports the results obtained from the proliferation experiments. In addition, these findings support that OXT-induced proliferation of the vaginal epithelium could be a mechanism by which vaginal OXT treatment increases thickness of the vaginal epithelium, thereby relieving symptoms of vaginal atrophy in postmenopausal women.7,8 The restorative effects of OXT on vaginal mucosa should be further explored.
Footnotes
Author contributorship
TKK and KUM designed the study and lab work was performed by TKK. Results were analysed by the two authors who also contributed equally to the finalized manuscript.
Declaration of conflicting interests
The author(s) declared the following potential conflicts of interest with respect to the research, authorship, and/or publication of this article: This study was enabled through commissioned research whereby Peptonic Medicals financed the salary of TKK and the lab costs. KUM is a board member and consultant and owns stock in Peptonic Medical.
Ethical approval
This study only included cell lines and there was no need for ethical approval.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This study was enabled through commissioned research where Peptonic Medicals financed the salary of TKK and the lab costs. No additional funding was received.