
Abstract
Background:
Tinnitus is an auditory phantom perception in the absence of any corresponding acoustic stimulus whose pathophysiology remains poorly understood. This study aimed to investigate alterations in the functional organization of the brain in individuals with tinnitus using resting-state functional magnetic resonance imaging (rs-fMRI) and graph theory analysis.
Methods:
We conducted a study including 44 individuals with tinnitus and 32 healthy controls. Using rs-fMRI and graph theory measures, we characterized whole-brain topological properties, including network segregation, integration, small-worldness, and global efficiency. In addition, regional segregation and integration were assessed using clustering coefficient and participation coefficient analyses to identify alterations in brain hub regions.
Results:
Our findings revealed altered topological properties in the tinnitus brain, particularly in the balance between cerebral segregation and integration, leading to deviations from optimal small-world architecture. We also observed alterations in the topology of specific auditory and nonauditory brain regions associated with phantom sound perception. Notably, patients with tinnitus exhibited a decreased nodal participation coefficient in the thalamus, suggesting reduced connectivity between this region and different functional modules as well as long-range connections.
Conclusions:
These results suggest that tinnitus is associated with alterations in the functional organization of the brain, leading to disrupted information processing and sensory integration.
Impact Statement
This article investigates functional modifications in patients with chronic tinnitus using resting-state functional magnetic resonance imaging and graph theory analysis. The results show that individuals with tinnitus, compared with healthy volunteers, exhibit distinct topological patterns in their resting-state brain activity. We demonstrate decreased segregation in several brain regions in patients with tinnitus, as well as decreased integration in the right thalamus. These results suggest that tinnitus may be associated with alterations in the functional organization of the brain, leading to disrupted information processing and sensory integration.
Introduction and Aim
Tinnitus is a phantom perception of sound in the absence of any corresponding auditory input from the environment (Moller, 2011). It is a common condition that affects ∼15% of the population, with 1–3% of individuals experiencing severe distress due to chronic tinnitus (when tinnitus is experienced more than 3 months or 6 months, depending on the definition) (Cima et al., 2019; Eggermont and Roberts, 2004). Despite its high prevalence, the precise underlying functional neuroanatomy of tinnitus remains poorly understood. Several theories have been proposed to explain the central compensatory mechanisms involved in tinnitus. These include increased spontaneous firing rates and neuronal synchrony, often linked to cochlear damage (Roberts et al., 2010), abnormal thalamocortical dysrhythmia, characterized by disrupted thalamic and cortical oscillatory activity (Moazami-Goudarzi et al., 2010), and the central gain theory, which posits that reduced peripheral input leads to amplification of auditory signals in central pathways (Sedley, 2019). However, these plastic changes in central auditory structures alone cannot fully account for the diverse clinical characteristics of tinnitus or explain why some individuals recover while others develop chronic, disabling tinnitus. To explain this variability, it has been suggested that dysfunctional interactions between auditory and nonauditory regions—such as the limbic system, frontostriatal circuits, prefrontal cortex, and parietal regions—may sustain the conscious awareness of tinnitus and hinder habituation (De Ridder et al., 2011; Maudoux and Lefebvre, 2014; Rauschecker et al., 2010; Schlee et al., 2009; Vanneste and De Ridder, 2012). These interactions are believed to amplify the emotional distress and attentional focus associated with tinnitus, creating a self-perpetuating feedback loop. Even if those models differ, they share the common concept that tinnitus involves a network-level deficit rather than in a single structured abnormality.
Traditional functional connectivity approaches, such as region-of-interest (ROI) correlations and independent component analysis (ICA), have significantly advanced our understanding of brain network alterations in tinnitus. However, these methods rely on predefined regional selection or statistical decomposition, which may limit reproducibility and overlook complex organizational principles of the connectome. In contrast, graph theory analysis enables a data-driven, whole-brain characterization of network topology, capturing both segregation (local specialization) and integration (global communication) (Deco et al., 2015; Rubinov and Sporns, 2010). Graph-theoretical metrics (including clustering coefficient, participation coefficient, efficiency, and small-worldness) have proven effective not only for delineating normal brain organization (Bassett and Bullmore, 2006; Mohan et al., 2016), but also for detecting clinically relevant alterations in a wide range of neurological and psychiatric disorders, from Alzheimer’s disease and schizophrenia to depression (Chang et al., 2020; Stam, 2014).
From a theoretical perspective, cognitive models emphasize that failures of sensory gating and aberrant salience attribution can cause intrinsic auditory signals to propagate abnormally into higher-order attentional and emotional systems. In line with this, the Global Neuronal Workspace theory postulates that conscious access arises when information becomes globally integrated and broadcast across distributed subcortical–cortical networks, including the thalamus, frontal, temporal, and parietal regions (Dehaene et al., 2011; Panda et al., 2022). Tinnitus, as a persistent and consciously accessible percept, may thus reflect maladaptive recruitment of this workspace, in which aberrant auditory activity disrupts the normal balance between local segregation and global integration. Accordingly, in order to test this hypothesis, we focused on graph-theoretical measures that directly capture these properties: clustering coefficient to quantify local segregation, and participation coefficient to index cross-network integration, together with complementary efficiency-based metrics to assess information transfer at local and global scales. Notably, the thalamus has emerged as a potential hub in this process. Functional imaging studies report reduced thalamo-limbic and thalamo-frontal connectivity, abnormal coupling with visual cortices, and decreased intrinsic activity within the thalamus itself (Chen et al., 2014; Kok et al., 2024; Zhang et al., 2015). This suggests that thalamic dysfunction could play a central role in the persistence of tinnitus, although its contribution within large-scale network organization remains unclear.
The present study applies high-resolution, whole-brain graph theory analysis to a cohort of 44 tinnitus patients and 32 healthy controls. By examining both global topology and regional hub properties, we aimed to capture tinnitus-related reorganization across multiple scales. We specifically hypothesized that tinnitus would be associated with disruption of the normal balance between segregation and integration at the global level, together with altered regional properties, thereby supporting the view that tinnitus reflects large-scale functional reorganization rather than dysfunction confined to auditory pathways.
Materials and Methods
Subjects and magnetic resonance imaging acquisition
Forty-four participants with tinnitus (18 females, age range 19–78 years) and 32 healthy controls (13 females, age range 19–72 years) underwent resting-state functional magnetic resonance imaging (rsfMRI). Exclusion criteria were contraindication for magnetic resonance imaging (MRI) (e.g., the presence of ferromagnetic aneurysm clips and pacemakers), hyperacusis, or phonophobia. All patients were free of major neurological, neurosurgical, or psychiatric history.
Resting-state blood oxygen level-dependent (BOLD) data were acquired on a 3 T MRI scanner (Siemens, Trio Tim, Germany) with a gradient echo-planar sequence using axial slice orientation (32 slices; voxel size = 3.0 × 3.0 × 3.75 mm3; matrix size = 64 × 64 × 32; repetition time = 2000 ms; echo time = 30 ms; flip angle = 78º; and field of view = 192 mm). A protocol of 300 scans lasting 600 sec was performed. A T1-weighted MPRAGE sequence was also acquired for registration with functional data on each subject.
For patients, audiological and clinical examinations were performed. Hearing levels were assessed using standard audiological testing. Pure tones ranging from 250 Hz to 8 kHz were presented to each ear until the detection threshold was reached. Patients were tested to identify the frequency that best matched their perceived tinnitus pitch. They were also asked to identify the best match to the perceived intensity of their tinnitus; this intensity was corrected for the hearing threshold measured at the corresponding frequency.
For each patient, we calculated the four-frequency pure-tone average (4fPTA) by obtaining the means of the thresholds at 500, 1000, 2000, and 4000 Hz for each ear, and then selected the better-ear 4fPTA for each participant. Ear-specific and better-ear 4fPTAs were then used to assign a World Health Organization-proposed hearing impairment (HL) grade as follows: (1) normal, ≤19.50 dB HL; (2) slight/mild, 19.51–34.5 dB HL; (3) moderate, 34.51–49.5 dB HL; (4) moderately severe, 49.51–64.5 dB HL; (5) severe, 64.51–80.5 dB HL; and (6) profound, ≥ 80.51 dB HL).
All included patients had experienced tinnitus for at least 3 months, with a mean duration of 3.2 ± 6.5 years.
Tinnitus intensity during MRI was also reported on a scale between 0 (not heard) and 10 (maximum). Self-reported severity of tinnitus impact was measured using the Tinnitus Handicap Inventory (THI), which measures the impact of tinnitus on daily living (Newman et al., 1996). THI is composed of 25 questions, with higher scores indicating more tinnitus-related disability (from 0 to 100). Results are classified into five categories: slightly disability (from 0 to 16), mild disability (from 18 to 36), moderate disability (from 38 to 56), severe disability (from 58 to 76), and catastrophic (from 78 to 100).
Written informed consent was obtained from all patients and healthy volunteers. The study was approved by the Ethics Committee of the Faculty of Medicine of the University of Liège, Belgium (reference number: B707 2009-6160), and was conducted in accordance with the General Data Protection Regulation and with the 1964 Helsinki declaration and its later amendments. A subset of participants in the current tinnitus group (13 out of the 44 included) were previously included in previous work (Maudoux et al., 2012a, 2012b), which used ICA and seed-based functional connectivity approaches to investigate rsfMRI data.
Data preprocessing and analysis
The structural (T1-weighted MPRAGE) and rsfMRI data were preprocessed using SPM12 (http://www.fil.ion.ucl.ac.uk/spm/software/spm12/) with the following steps: discarding the first five functional volumes, slice-time correction, realignment, coegistration, transforming the functional images of all the subjects to the Montreal Neurological Institute standard space, and normalization. The structural data were segmented into gray matter, white matter, and cerebrospinal fluid (CSF) to regress out signals related to physiological artifacts.
First, we extracted the head movement from six direction motions (three directions translational and three rotational directions), and physiological signals were extracted from white matter and CSF signals in the BOLD fMRI images. Following this, physiological signals were regressed out by removing the first five principal component analysis (PCA) components of the white matter and CSF signals, and the head motion was regressed using the Friston 24-parameters model regression (Friston et al., 1996), as recommended by the aCompCor denoising approach (Behzadi et al., 2007). Finally, the rsfMRI data were filtered using a 0.01–0.09 Hz band-pass filter (Holla et al., 2017). We additionally applied motion scrubbing, excluding frames with framewise displacement >0.5 mm to minimize residual motion-related artifacts.
Brain regions time series extraction and connectivity matrix construction
After preprocessing, the rsfMRI data were parcellated into 268 brain ROIs covering cortical, subcortical, and cerebellar areas using the Shen 268-node functional atlas (Shen et al., 2013) through the MarsBaR toolbox® (http://marsbar.sourceforge.net). We selected this atlas because it offers a fine-grained, high-resolution, data-driven parcellation derived from rsfMRI, ensuring that each region is functionally homogeneous. This provides greater sensitivity and functional specificity than coarser anatomical atlases. Moreover, the Shen atlas has been widely used to assess brain network alterations in various neurological conditions, including in our previous work (López-González et al., 2021; Panda et al., 2023). Regional time series were extracted as the mean BOLD signal across all voxels within each ROI. Although alternative methods, such as PCA (extracting the first eigenvector), can be employed, averaging was retained here due to its robustness, interpretability, and widespread adoption (Power et al., 2011; Rubinov and Sporns, 2010), as well as its use in tinnitus functional connectivity studies, allowing comparability with previous work.
Pearson correlation coefficients were computed between all ROI pairs to construct subject-level connectivity matrices. The correlation coefficients were subsequently transformed into Fisher’s Z values to stabilize variance and normalize their distribution prior to group-level analysis. Connectivity matrices were then represented as weighted, undirected graphs. To improve interpretability and align with common rsfMRI practice, negative correlations were set to 0 prior to thresholding so that only positive edges were retained for graph construction (Rubinov and Sporns, 2010). Because global signal regression can induce artifactual anticorrelations, excluding negative edges reduces sign ambiguity and improves metric stability (Murphy et al., 2009). Pearson correlation coefficients were first transformed into Fisher’s Z values. Negative correlations were then set to 0 prior to sparsity thresholding, such that only positive edges were retained for graph construction. Sparsity-based thresholding was then performed on the positive weighted graphs, and metrics were computed on the resulting positive weighted graphs, and metrics were computed on the resulting positive weighted networks. For path-based measures, edge lengths were defined as follows:
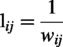
Graph theory analysis
The topological properties of the functional brain networks were defined based on 268 × 268 weighted undirected networks, G (N, E), where N represents the set of nodes (ROIs), and each edge Eij represents the strength of functional connectivity between node i and node j.
To characterize the functional brain networks, graph theory connectivity measures, including clustering coefficient (γ), characteristic path length (λ), participation coefficient (PC), small-worldness (σ), global efficiency (Eglob), and local efficiency (Eloc), were computed using the Brain Connectivity Toolbox (http://www.brain-connectivity-toolbox.net) (Rubinov and Sporns, 2010). Graph measures were computed on thresholded networks across a range of sparsity values, as detailed in the Thresholding section below. Sparsity (S) was defined as the proportion of possible edges retained in the thresholded network,
Clustering coefficient (γ)
Brain network segregation was quantified using the clustering coefficient, which represents how well brain regions are connected through short-range connections to form segregated modules within the whole network. For an undirected graph, the local clustering coefficient of node i (called as absolute clustering coefficient, [Ci]), is defined as the ratio between the number of existing connections (Ei) among the neighbors of node i and the number of all feasible connections among these neighbors. The global clustering coefficient C is then calculated as the average of the local clustering coefficients across all nodes (Watts and Strogatz, 1998):

Followed by, we calculated the normalized clustering coefficient (γ), taking the ratio of the absolute clustering coefficient of the network (C) and the random clustering coefficient (CRand) (γ = C/CRand). CRand is the mean clustering coefficient of randomized networks with the same degree distribution as the real network. Following standard practice (Humphries and Gurney, 2008; Rubinov and Sporns, 2010; Watts and Strogatz, 1998), we generated 1000 random networks for each sparsity threshold using an edge-rewiring algorithm that preserves the degree distribution of the original network.
For each brain region, a nodal clustering coefficient was obtained by calculating the AUC of its normalized clustering coefficient values across the range of sparsity thresholds, yielding a threshold-independent metric. Finally, we performed between-group comparisons of these nodal clustering coefficients to identify regions with significantly altered clustering in the patient group as compared to healthy control. Brain regions with high clustering coefficients, often referred to as “provincial hubs,” are characterized by having densely interconnected neighboring regions, reflecting predominantly short-range, within-module connections.
Normalized path length (λ)
The normalized path length (λ) is a metric used to quantify network integration and describes how effectively the nodes in a network are interconnected. For weighted graphs, edge lengths were defined as the inverse of the correlation weights,
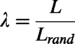
Small-worldness (σ)
Small-worldness (σ) describes the extent to which a network simultaneously exhibits high local clustering (segregation) and short path lengths (integration), reflecting an optimal balance between specialized and distributed processing. In brain networks, small-world topology supports efficient information transfer with minimal wiring cost. The concept of small-world networks was originally introduced by Watts and Strogatz (1998). Small-worldness (σ) was quantified as the ratio of the normalized clustering coefficient (γ = C/Crand) to the normalized characteristic path length (λ = L/Lrand), where C and L are the clustering coefficient and path length of the empirical network, respectively, and Crand and Lrand are the corresponding averages from degree-preserving randomized networks. Mathematically, a real network would be considered as small-world if it meets the two conditions: (1) γ > 1 and (2) λ ≈ 1. A network follows a small-world organization (toward regular properties) when σ = γ/λ > 1 (Humphries and Gurney, 2008). Following standard practice (Humphries and Gurney, 2008; Rubinov and Sporns, 2010), we generated 1000 random networks for each sparsity threshold using an edge-rewiring algorithm that preserved the degree distribution of the original network. Results were then summarized across the full sparsity range (0.06–0.45) using the AUC approach to reduce sensitivity to single thresholds.
Participation coefficient
The participation coefficient is a measure of brain integration that captures the diversity of intermodular connections of individual nodes (Guimerà and Nunes Amaral, 2005). Nodes with a high participation coefficient have strong connections across segregated modules and long-range connections, a property that has proven useful in recent applications to brain networks (Acharya et al., 2022; Deco et al., 2015). To compute the participation coefficient, modular partitions were obtained using the Louvain algorithm (Blondel et al., 2008) with a resolution parameter of γres = 1.0 (renamed here to avoid confusion with the normalized clustering coefficient γ defined above). To ensure stability, the algorithm was repeated 100 times at each sparsity threshold, and consensus partitions were derived. The participation coefficient of a node (PCi) is defined as (Guimerà and Nunes Amaral, 2005) follows:
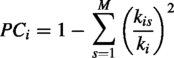
Network efficiency
The topological efficiency of a network can be measured by two metrics: (1) global efficiency (Eglob) and (2) local efficiency (Eloc), which reflect how well information is transmitted across the network (Iturria-Medina et al., 2008; Latora and Marchiori, 2001).
Eglob quantifies the network’s ability to efficiently exchange information at the global level and is defined as the average inverse shortest path length across the network:
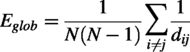
Conversely, Eloc reflects the efficiency of information transfer within local neighborhoods, measuring the resilience of subnetworks to node removal. It is defined as follows:
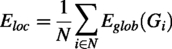
All path-based measures were computed on positively weighted graphs, with edge lengths defined as
Thresholding
To ensure comparability of network measures across individuals, networks were thresholded using a sparsity range 0.06 ≤ S ≤ 0.45 (step 0.01), as recommended by Holla et al. (2017). Graph measures were integrated across thresholds by computing the AUC, a widely accepted approach that increases robustness to threshold selection.
Statistical analysis
In this study, global graph measures were compared between healthy controls and patients suffering from tinnitus using a two-sample t test (two-tailed). Brain regional clustering coefficients and participation coefficients were compared between the two groups across 264 brain regions, and multiple comparisons were corrected using the false discovery rate correction, with statistical significance set at p < 0.05. Subsequently, for brain regions that showed significant differences in clustering and/or participation coefficients between patients and healthy controls, these regions clustering and/or participation, along with whole-brain graph measures, were correlated with clinical variables using linear Pearson linear correlation analysis. Before computing correlations, the assumptions of linearity and normality were verified, and outliers (>3 standard deviation [SD] from the mean) were excluded. Given the variability of the data, these analyses were considered exploratory, and the results should be interpreted with caution. Correlation coefficients (r values) were considered statistically significant at p < 0.05.
Results
Population
Thirty-two healthy controls were recruited (13 females; age range, 19–72 years). Forty-four patients suffering from tinnitus for at least 3 months (18 females; age range, 19–78 years) were recruited for the study. Six patients were discarded because of technical issues. Therefore, analyses were conducted on 38 patients. Details of the characteristics of these 38 patients are provided in Table S1 of the Supplementary Files.
Patients had tinnitus for an average duration of 3.2 ± 6.5 years (mean ± SD; range, 0.25–33 years). Tinnitus-matched frequencies ranged from 250 Hz to 12.5 kHz (5000 ± 3270 Hz). Corrected tinnitus-matched intensities (objective loudness) ranged from 0 to 5 dB HL (19 ± 18 dB HL). Tinnitus Handicap Inventory (THI) (Newman et al., 1996) scores ranged from 2 to 94 (44.4 ± 25.0; N = 37, with one patient missing).
The numeric rating of the tinnitus subjective intensity during scan on scale of 0–10 had an average value of 5 ± 3. For each patient, the 4fPTA of the better ear was calculated. Twenty-five patients had normal auditory threshold, 12 subjects had a slight/mild hearing loss, and 1 subject had moderate hearing loss. No patient had moderately severe, severe, or profound hearing loss.
Difference in global network parameters
The topological properties of the whole-brain functional networks were compared between patients with tinnitus and healthy controls, revealing significant differences in normalized clustering coefficient (γ), small-worldness (σ), and normalized local efficiency (Fig. 1 and Supplementary Fig. S1). Specifically, patients showed a significantly reduced normalized clustering coefficient (γ) of 1.71 ± 0.59 compared to controls with 2.30 ± 0.53 (p = 0.0002), small-worldness (σ) of 1.51 ± 0.42 compared to controls with 1.76 ± 0.47 (p = 0.017), and normalized local efficiency of 1.40 ± 0.42 compared to controls with 1.63 ± 0.44 (p = 0.032). Although the global participation coefficient tended to be lower in patients with tinnitus than in controls, the difference did not reach statistical significance.

Modifications in brain topology in patients with tinnitus, characterized by reduced brain network segregation in tinnitus. Asterisks (*) in the graphs
Differences in regional nodal parameters
Our analysis of nodal topological properties revealed significant differences between patients with tinnitus and healthy controls. Specifically, we observed decreases in both the clustering coefficient (representing short-range connections/local connectivity) and the participation coefficient (representing long-range connections/integration across modules) in specific nodes in patients compared with controls. The decrease in nodal clustering coefficient was observed across a widespread distribution of temporal, frontal, parietal, and occipital regions, indicating a breakdown in local connectivity within these regions (Fig. 2 and Supplementary Table S2). Moreover, we found a reduction in the nodal participation coefficient in the right thalamus, particularly in the pulvinar and lateral posterior thalamus, suggesting a disconnection from different functional modules and long-range connections (Fig. 3). These findings suggest that the thalamus may have lost its connector hub properties in patients with tinnitus.
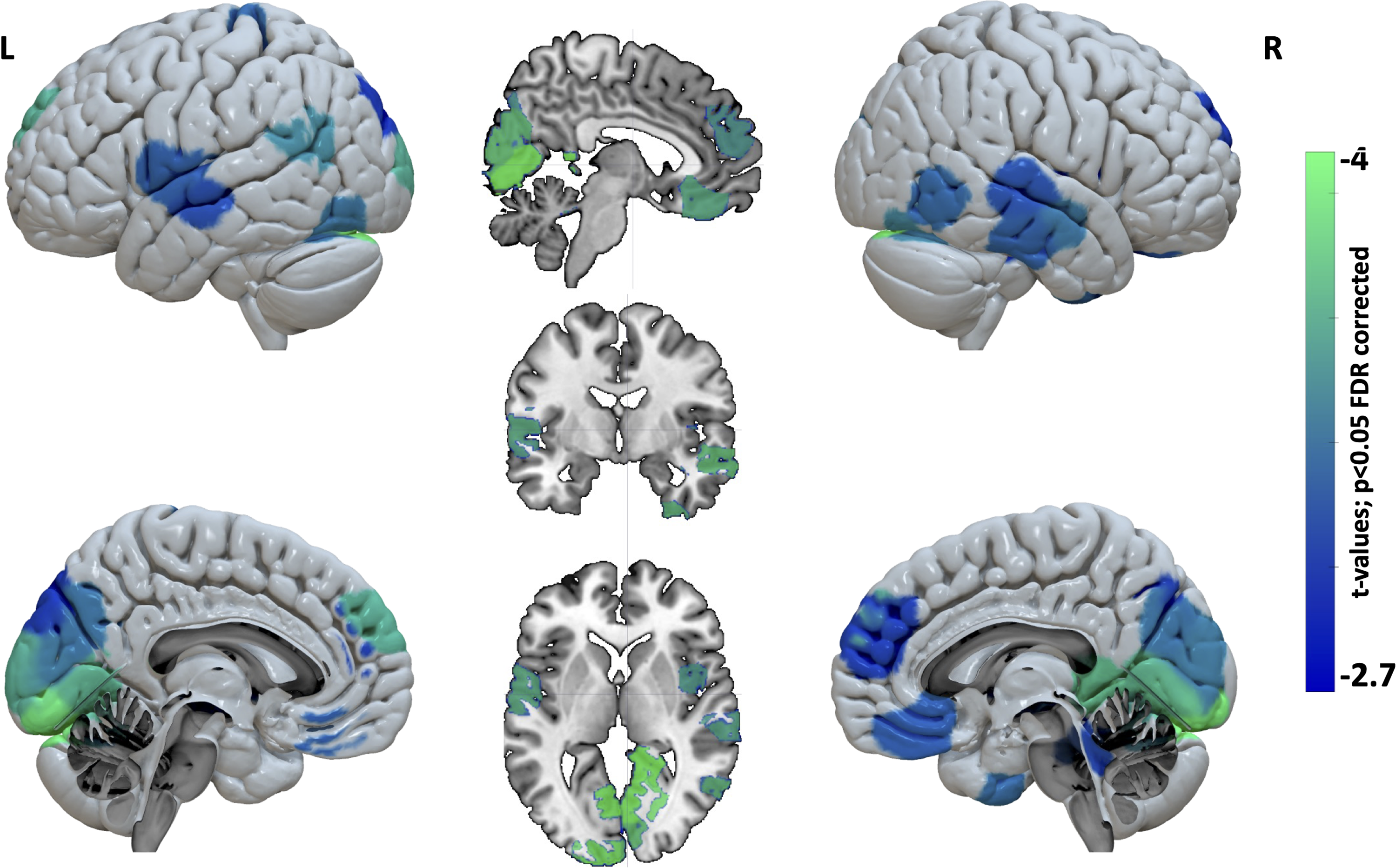
Decreased network segregation in patients with tinnitus compared with healthy controls. Brain regions showing decreased segregation in patients included temporal regions, the dorsolateral prefrontal cortex (DLPFC), orbitofrontal/ventromedial prefrontal cortex, dorsomedial prefrontal cortex, occipital cortex, sensorimotor cortex, salience network, default mode network (DMN) and posterior fossa. L, left; R, right.
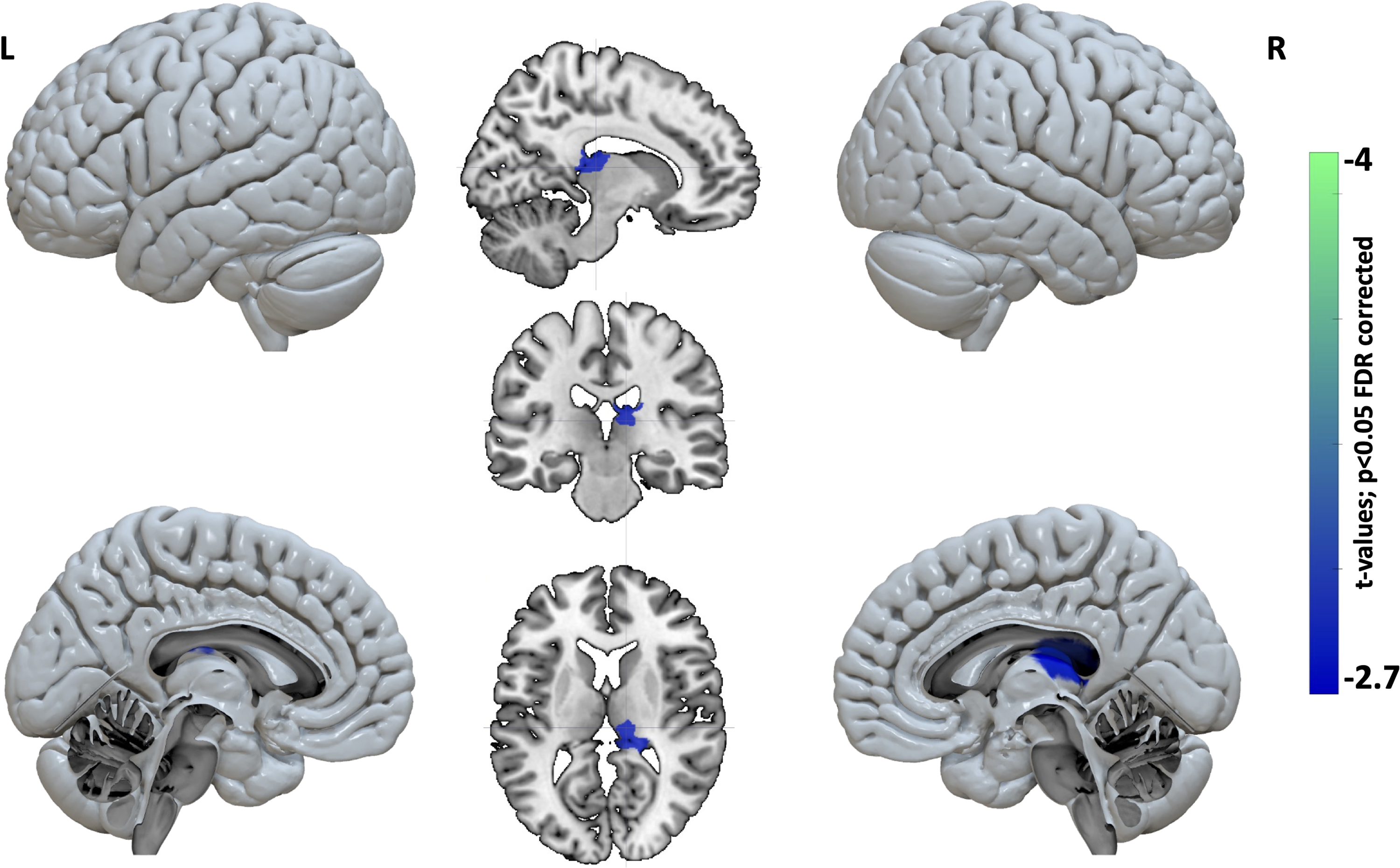
Decreased network integration in patients with tinnitus compared with healthy controls, particularly in the right thalamus, including the pulvinar and lateral posterior thalamus. L, left; R, right.
Further analyses were performed to investigate the associations between modifications in global and local topological properties and various clinical variables related to tinnitus and population characteristics. To address whether the observed modifications in brain connectivity were specifically associated with tinnitus perception rather than other confounding factors, we performed correlation analyses between tinnitus-related behavioral scores and network topological properties. These analyses focused on both global metrics and regional properties within brain areas identified as significantly altered in the primary group comparisons. Specifically, we observed negative correlations between the whole-brain clustering coefficient and tinnitus intensity experienced during the scanning session, tinnitus objective loudness, and global subjective tinnitus intensity evaluated with the Visual Analog Scale (Fig. 4a–c). Moreover, we found that the observed decrease in thalamic network integration (nodal participation coefficient) was negatively correlated with perceived tinnitus intensity during the scan (Fig. 4d). In addition, we observed negative correlations between tinnitus intensity experienced during the scan/tinnitus objective loudness and left Rolandic opercular clustering coefficient (Fig. 4e,f) and also between tinnitus subjective intensity and right inferior temporal/left lingual/right cerebellum clustering coefficient (Fig. 4g–i). Importantly, we found no significant correlations between global or local network properties and demographic or audiological factors such as age, sex, or hearing level. This result provides reassurance that the observed brain network alterations in tinnitus patients are not attributable to these characteristics but are instead closely related to tinnitus perception and distress.
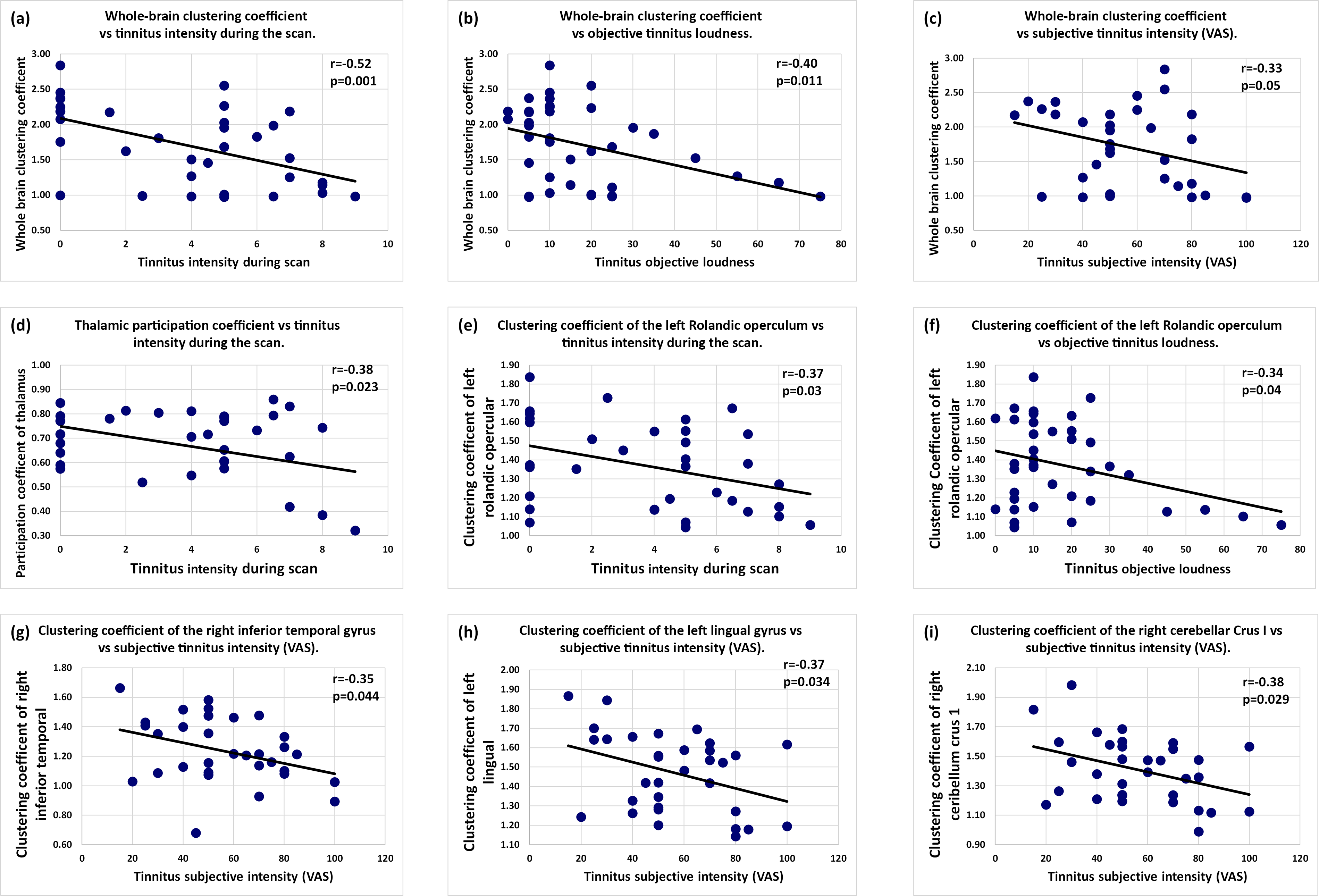
Correlations between brain network measures and clinical measures. (
Discussion
In this study, we investigated distinct topological patterns of resting-state brain activity in patients with tinnitus compared with healthy controls by employing fMRI and graph theory analysis. Our primary objective was to understand how tinnitus affects the functional organization of large-scale cortical networks, which may reveal the underlying mechanisms associated with phantom auditory perception and its persistence.
The brain’s optimal organization has been characterized as a “small-world” topology, a network configuration that balances high local clustering with efficient global connectivity. Small-world networks facilitate both localized and specialized processing within clusters (segregation) and effective communication across distant brain regions (integration) (Bassett and Bullmore, 2006). This configuration supports the brain’s capacity for both rapid information processing and modular specialization, enabling it to respond flexibly and adaptively to a range of cognitive and sensory tasks (Shine, 2019). Our findings suggest a deviation from this optimal organization in tinnitus patients, as evidenced by modification of global whole-brain connectivity (Supplementary Fig. S1) and alterations in key topological properties. Specifically, we observed a reduction in the clustering coefficient and local efficiency, indicating less effective communication within local networks (Fig. 1).
To gain insight into the impact of specific brain regions on global functional organization, we examined topological properties at the node level. Our analysis revealed that tinnitus patients exhibit significant reductions in nodal clustering coefficients across both auditory and nonauditory regions, implying a disruption in local connectivity that may affect the brain’s ability to process sensory information efficiently (Supplementary Table S2, Fig. 3). Indeed, regions with reduced nodal clustering coefficients are regions that are less connected with their immediate neighbors, potentially impairing local processing dynamics by losing their “provincial hubs” characteristics. This reduced clustering coefficient was particularly pronounced in the temporal cortex, dorsolateral prefrontal cortex, orbitofrontal cortex, dorsomedial prefrontal cortex, occipital cortex, sensorimotor cortex, as well as regions involved in the salience network and default mode network (DMN). These regions are known to support a range of cognitive, sensory, and emotional processes (Jones and Graff-Radford, 2021; Kawashima, 2018; Rushworth et al., 2004; Xiong et al., 2020). The implication of these regions is consistent with observations previously described in tinnitus using fMRI (Araneda et al., 2018; Lv et al., 2020). By losing their hub-like connectivity, these regions may contribute to the conscious perception of tinnitus, as well as its associated distress and persistence. For instance, the salience network, which includes the anterior insula and dorsal anterior cingulate cortex, plays a crucial role in detecting and filtering salient sensory information, enabling the brain to prioritize certain stimuli over others (Seeley et al., 2007). Alterations in this network could lead to an exaggerated focus on the tinnitus sound, making it difficult for patients to ignore or habituate to the perception. Similarly, the DMN, which is typically active during rest and internally focused thought, may become dysregulated in tinnitus, leading to increased awareness of the phantom sound even in the absence of external stimuli. Notably, the involvement of extra-auditory regions, including key nodes of the salience network and the DMN, is highly consistent with influential theoretical models of tinnitus proposed by Jastreboff, Schlee, and De Ridder. These models posit that the persistence and conscious awareness of tinnitus are not solely driven by aberrant activity in the auditory pathways, but also rely on dysfunctional interactions with broader networks responsible for attention, salience attribution, and emotional regulation (De Ridder et al., 2011; Jastreboff, 2011; Schlee et al., 2011).
In line with the Global Workspace Theory proposed by Baars and further developed by Dehaene and Naccache, conscious experiences emerge when information is globally broadcast across widespread brain networks (Baars, 1988; Dehaene and Naccache, 2001). According to this theory, only one consistent content can dominate consciousness at any given moment, implying that the conscious perception of tinnitus may result from the brain’s inability to filter the tinnitus signal from this global workspace. Alterations in connectivity, particularly within networks associated with attention and sensory processing, may prevent the brain from downregulating tinnitus perception. The decreased nodal clustering observed in our study suggests that tinnitus patients may experience difficulty in segregating and processing sensory information locally, allowing the phantom sound to occupy the global workspace more persistently.
One of the most intriguing findings in our study is the significant decrease in the participation coefficient of the posterior thalamus, specifically within the pulvinar nucleus (Fig. 3). Traditionally, the thalamus has been viewed as a passive relay station for sensory information. However, recent research has revealed that the thalamus plays a critical role in integrating and coordinating sensory information across cortical networks, acting as a connector hub in facilitating cortico-cortical communication (Hwang et al., 2017). The thalamus is organized into first-order and higher-order nuclei, with the latter, including the pulvinar, involved in more complex functions such as sensory integration and multimodal processing. A decrease in the participation coefficient within the thalamus indicates reduced connectivity between the thalamus and cortical networks, potentially limiting its ability to modulate and regulate sensory input effectively. This may explain the reduced modular organization and increased between-network connectivity observed in our study. Several previous studies have highlighted the involvement of the thalamus and its connectivity in chronic tinnitus, providing essential context for our findings. Seed-based functional connectivity analyses, such as those by Zhang et al. (2015), demonstrated disrupted thalamocortical connectivity in tinnitus patients, with reduced coupling between the thalamus and cortical regions (including the temporal, frontal, and occipital cortices), correlating with both tinnitus distress and duration. Amplitude-based approaches, like Amplitude of Low-Frequency Fluctuations (ALFF) analyses (Chen et al., 2014), further revealed decreased spontaneous activity within the bilateral thalamus in tinnitus patients. Additionally, Kok et al. (2024) recently reported increased resting-state connectivity between the thalamus and the right visual association cortex, underscoring the thalamus as a central node affected in tinnitus pathophysiology. While these prior studies have illuminated region-specific alterations, they largely relied on seed-based or amplitude measures, which, although informative, provide a more limited view of the thalamus’s role at the whole-brain network level. In contrast, our graph-theoretical approach enables us to assess the thalamus’s role as a connector hub across large-scale networks. The observed reduction in participation coefficient within the pulvinar nucleus suggests a weakened integrative capacity of the thalamus, reflecting not only disrupted local connectivity but also a global breakdown in network-level communication. This reduced hubness may limit the thalamus’s ability to coordinate information flow between distributed brain regions, particularly between sensory and higher-order modules, thus contributing to the altered sensory experience characteristic of tinnitus. The thalamocortical dysrhythmia (TCD) hypothesis, initially described by Llinás and later applied to tinnitus by De Ridder, proposes that tinnitus arises from disrupted thalamocortical activity resulting in a self-sustaining loop of abnormal rhythmic firing between the thalamus and cortex (De Ridder et al., 2011; Llinás et al., 1999). This model suggests that damage to the auditory pathway or reduced sensory input leads to thalamic hyperpolarization, initiating abnormal synchronous firing that cascades through cortical regions. Our findings support this hypothesis, as the observed reduction in thalamic integration (reflected by lower participation coefficient) may indicate a loss of regulatory control over cortical networks, contributing to the heightened and persistent perception of tinnitus. The pulvinar’s role in mediating attention and integrating sensory information across cortical regions makes it a crucial node for maintaining sensory coherence, and its reduced participation in tinnitus patients may disrupt the normal sensory gating mechanisms that filter out irrelevant stimuli. Since the introduction of the TCD concept, numerous studies have implicated the thalamus in both the generation and maintenance of tinnitus, suggesting that thalamocortical interactions play a critical role in the development and persistence of this condition (De Ridder et al., 2011; Koops and Eggermont, 2021; Vanneste et al., 2018). These studies propose that tinnitus may result from an imbalance between excitatory and inhibitory activity within the thalamus, leading to abnormal thalamocortical oscillations and the perception of phantom sounds. This shift disrupts the typical rhythm of thalamic signaling, creating a state of dysrhythmia that affects sensory processing in the auditory cortex. Research by Berlot et al. (2020) demonstrated reduced thalamocortical and cortico-cortical connectivity in tinnitus patients, reinforcing the idea that altered thalamic integration may contribute to tinnitus persistence. These findings underscore the importance of the thalamus as a central hub in the pathological network dynamics underlying tinnitus and suggest that its disrupted connectivity with other brain regions may be a core feature of tinnitus pathogenesis.
Although graph theory has become an established tool in the study of various neurological and psychiatric disorders, its application to tinnitus remains relatively novel, with only a limited number of publications to date (Han et al., 2025; Lan et al., 2022; Lin et al., 2020). Lan et al. (2022) demonstrated, using graph theoretical analysis of rsfMRI data, that patients with tinnitus, both acute and chronic, exhibit marked reorganization of brain network topology, particularly affecting the distribution of rich-club nodes that play a central role in global brain communication. Furthermore, they observed centrality changes that varied with tinnitus chronicity. Our results complement and extend these findings. Like Lan et al., we observed deviations from optimal brain network organization in tinnitus, as evidenced by significant alterations in key topological properties (Fig. 1). We also found that these network alterations were not confined to auditory regions but extended to associative and limbic areas, including the prefrontal cortex, hippocampus, and amygdala (Figs. 2 and 3). These regions are implicated in the conscious perception, sensory filtering, and emotional regulation of tinnitus. Our findings can also be meaningfully compared to the results reported by Han et al. (2025), who examined the topological properties of brain functional networks in tinnitus patients with different symptom durations. In their study, recent-onset tinnitus patients exhibited compromised network segregation compared with both persistent tinnitus patients and healthy controls. Interestingly, Han et al. observed that many of these topological measures tended to normalize in the persistent tinnitus group, suggesting a partial restoration of network organization over time, although some alterations in intra- and inter-modular connectivity persisted. In our study, which included primarily patients with chronic tinnitus, we observed enduring reductions clustering coefficient and local efficiency, both globally and regionally. The persistence of diminished segregation and hub disruption in our cohort may reflect a subgroup of chronic tinnitus patients with sustained network dysfunction, underscoring the heterogeneity of the condition and highlighting the need for longitudinal studies to better define the trajectories of large-scale network reorganization in tinnitus. Nevertheless, both studies converge on the notion that tinnitus, particularly at onset, involves a breakdown in local network specialization and that these alterations may be only partially reversible with chronicity. In contrast to the work of Lin et al. (2020), who observed increased global and local efficiency, as well as clustering coefficient in anatomical networks of tinnitus patients, our results instead point to a reduction in clustering coefficient and local efficiency, indicating less effective communication within local subnetworks (Fig. 1). This apparent discrepancy may be explained by differences across studies in the modality (structural vs. functional networks), included clinical populations, and the chronicity of tinnitus. While Lin et al. highlighted increased anatomical efficiency that may reflect compensatory mechanisms or plastic reorganization, our functional network findings instead suggest marked deficits in local processing and network specialization. Importantly, Lin et al. also identified a shift in network hubs from classical auditory and thalamic regions in controls to limbic areas (amygdala and parahippocampus) in tinnitus, reinforcing the notion that tinnitus is associated with a redistribution of network centrality from sensory to emotion-related areas, an idea our results also support through the identification of altered nodal properties in the orbitofrontal and limbic cortices.
Despite heterogeneity in data types and patient characteristics, a consistent finding has emerged from those studies, including our: tinnitus is associated with significant modifications in global brain network organization. Alterations in measures such as clustering coefficient, efficiency, and hub distribution indicate that tinnitus involves a distributed reorganization of both segregation and integration processes at the network level. These network-level disturbances commonly extend beyond the primary auditory cortices, involving associative, limbic, and prefrontal regions, and may underlie the complex sensory, cognitive, and emotional dimensions of tinnitus.
One limitation of our study is the reliance on a control population that did not undergo detailed auditory and psychological assessments. Instead, control participants were included based on self-reported absence of hearing or psychological issues. This approach leaves open the possibility that undetected conditions among controls could, in principle, influence group differences and thus impact the robustness of our findings. To mitigate this risk and validate the clinical relevance of the observed brain network alterations, we conducted correlation analyses between tinnitus-related behavioral measures and both whole-brain and regional network topological properties. We deliberately focused these analyses on brain regions identified as significantly different in our group comparison, a decision guided by both statistical power considerations and the exploratory nature of the study. While this approach might miss associations present in other brain regions and is less comprehensive than whole-brain correlational mapping with multiple comparison correction, it allowed us to provide targeted evidence for a relationship between connectomic changes and the perceptual or emotional impact of tinnitus. Our findings revealed significant negative correlations between whole-brain clustering coefficient, regional nodal parameters, and clinical variables related to tinnitus, particularly perceived tinnitus intensity and related distress (Fig. 4 and Supplementary Table S3). These correlations indicate that the functional reorganization observed in tinnitus patients is closely linked to the perception of the phantom sound. Specifically, we found that the perceived tinnitus intensity during the scanning session was negatively correlated with the thalamic participation coefficient (Fig. 4d and Supplementary Table S3). This suggests that as the thalamus becomes less integrated across functional modules, tinnitus perception intensifies. These findings reinforce the critical role of thalamocortical dysfunction in the conscious perception and maintenance of tinnitus, supporting the hypothesis that disruptions in thalamic connectivity contribute to the persistent awareness of the phantom sound. Nevertheless, given the variability in individual data and the cross-sectional design of our study, these correlations should be interpreted with caution, as robust associations rather than evidence of direct causality. For example, as illustrated in Figure 4c, some relationships appeared weak and scattered, emphasizing the exploratory nature of these findings and the potential limitations of Pearson correlation in this context. Importantly, we found no significant correlations between global or local network properties and demographic or audiological variables such as age, sex, or hearing level. This absence of association with these potential confounders increases our confidence that the observed brain network changes are specifically related to tinnitus processes, even though the influence of other unmeasured factors cannot be fully excluded. Several methodological considerations should also be taken into account regarding our analytical approach. First, we acknowledge that the use of parametric two-sample t tests may not fully capture the network-level dependencies and spatial autocorrelation inherent in graph-theoretical data. Although our results were consistent across metrics, nonparametric permutation-based inference would offer a more robust statistical framework and should be considered in future studies. Another important consideration is the absence of explicit assessments of network stability or reproducibility, such as test–retest, split-half, or bootstrap procedures. Given our relatively modest sample size, conducting such analyses would have resulted in very small subsamples, thereby limiting statistical power and reducing the interpretability of stability indices. As a result, we cannot fully determine the extent to which the observed topological alterations reflect stable neurobiological features versus sample-specific variability. We therefore recommend that future studies with larger cohorts incorporate these validation techniques to confirm the robustness and generalizability of network-level findings in tinnitus.
Another limitation of our study is the inherent challenge of translating resting-state connectivity findings into clinical applications. Although resting-state analysis can reveal altered connectivity patterns associated with conditions such as tinnitus, depression, and schizophrenia, resting-state connectivity can vary significantly between individuals due to factors such as age, attention, fatigue, and recent experiences. This individual variability complicates the generalization of results across populations and may contribute to the high degree of variability seen in existing tinnitus literature. Despite these challenges, our findings align with previous models of tinnitus generation that share a common perspective: tinnitus reflects a deficit at the network level rather than being confined to a single brain structure. This network-centered view underscores the complexity of tinnitus and highlights the importance of examining large-scale brain connectivity to gain a deeper understanding of its persistence and clinical variability.
Conclusion
This study highlights significant disruptions in brain network organization in tinnitus patients, manifesting as deviations from the optimal small-world architecture typically observed in healthy brains. We observed reductions in clustering coefficient and local efficiency, indicating a compromised ability for efficient local processing, together with a decreased thalamic participation coefficient, reflecting impaired long-range communication across the brain. Specifically, the reduced participation coefficient within the thalamus suggests thalamic dysfunction that not only affects local thalamic connectivity but also appears to disrupt broader network dynamics across cortical and subcortical regions. Indeed, decreased nodal clustering coefficients in both auditory and nonauditory brain areas suggest a breakdown in localized connectivity, impacting regions involved in auditory processing as well as cognitive and emotional regulation. Our findings both confirm and extend the current understanding of the role of thalamocortical and cortico-cortical dysconnectivity in the perception and persistence of tinnitus by demonstrating, via whole-brain network topology, that chronic tinnitus is linked to a disconnection of the thalamus from broader functional modules, highlighting the value of graph-theoretical analyses in unraveling complex network reorganization in this condition. Finally, correlation analyses revealed that stronger tinnitus perception is associated with reduced clustering coefficient, supporting the idea that tinnitus fundamentally reshapes brain connectivity.
Understanding the role of specific brain regions and thalamic modifications could provide new insights and potential treatments for tinnitus and other phantom perception. However, we must remain critical, as recent studies tend to show that the profile of tinnitus (i.e., with hearing loss or without hearing loss, bothersome tinnitus) and its associated comorbidities (i.e., distress) are important factors to take into account in the brain changes connectivity (Husain and Khan, 2023).
Data and Code Availability
The MRI and patient behavioral data supporting the findings of this study are available upon reasonable request from M.D. at the University of Liege, Belgium (
Authors’ Contributions
Marie Detroz and Audrey Maudoux conceived the study. Marie Detroz performed the analyses, contributed to data acquisition and preprocessing, and conducted clinical correlations. Audrey Maudoux supervised the study and critically revised the manuscript. Following completion of the preprocessing pipeline, Rajanikant Panda conducted the functional MRI analyses, including graph theoretical network analyses and related neuroimaging investigations. All authors reviewed and approved the final manuscript.
Footnotes
Acknowledgments
The authors thank the technicians of the Department of Radiology for their active participation in the MRI studies of patients with tinnitus.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This research was funded by the Belgian National Funds for Scientific Research and the University and University Hospital of Liège.
Supplemental Material
Supplemental Material
Supplemental Material
Supplemental Material
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.