
Abstract
Introduction:
Brain networks and meditation have recently gained attention, with studies suggesting that more efficiently organized meditation brain networks are linked to better cognitive performance. This efficiency is exemplified in small-world brain networks, which combine local segregation with global integration, facilitating optimal information processing. This study examines the small-world propensity (SWP), a marker of neural efficiency, among three functional brain networks: the default mode network (DMN), fronto-parietal network (FPN), and attention network (AN) in three groups: advanced meditators (AM), beginner meditators (BM), and control meditators (CM).
Methods:
Using magnetoencephalography (MEG), we recorded 10-min meditation sessions from AM and BM groups practicing Surat-Shabda-Yoga meditation at different stages. The CM (baseline group), with no formal training in meditation, had introductory exposure to “four chakra meditation” and practiced the same. SWP was computed using coherence-based connectivity measures across frequencies ranging from 4 to 45 Hz during stable meditative states.
Results:
Significant differences were observed between meditators and non-meditators, with AM and BM groups compared with the CM group. Specifically, the AN in the AM group compared with CM exhibited higher SWP at the beta frequency range (19 Hz), while the FPN in the BM group compared with the CM showed increased SWP at the theta frequency range (8 Hz).
Discussion:
These findings highlight how meditation engages the brain’s intrinsic network architecture in a frequency- and stage-specific manner, supporting efficient information processing and offering a scientific basis for its cognitive and regulatory benefits.
Impact Statement
This study reveals how meditation naturally resonates with the brain’s intrinsic architecture, promoting efficient network organization through small-world topology. Using MEG and coherence-based connectivity, we demonstrate frequency-specific adaptations in key functional networks such as DMN, FPN, and -AN across meditation stages. These changes highlight how meditation leverages neural connectivity for optimized information processing by linking contemplative practice to intellectual benefits. It contributes to the field of brain connectivity by positioning meditation as a modulator of functional network dynamics with potential cognitive applications.
Keywords
Introduction
The growing interest in exploring the core aspects of meditation stems from its demonstrated benefits on various cognitive and psychological functions, highlighting its role in promoting mental and physical well-being (Guidotti et al., 2023; Jones, 2019). These benefits are closely associated with changes in brain’s connectivity (Guidotti et al., 2023; Lardone et al., 2022). The brain, as a complex system, functions through the coordination of its regions, and connectivity plays a vital role in facilitating these interactions (Sporns, 2022). Recent advancements in network science provide a robust framework for investigating brain organization by modeling it as a network of interconnected nodes (representing brain regions) and edges (representing connections). This approach allowed the researchers to explore large-scale brain interactions and their role in higher-level functions like attention, memory, and emotional regulation (Friston, 2011; Vecchio et al., 2020). This method facilitates the examination of widespread changes in communication between brain regions through the lens of graph theory (Bullmore and Sporns, 2009).
Interestingly, studies have shown that the connectivity of meditators’ brains dynamically reorganizes during both meditation and rest (Cotier et al., 2017; Mooneyham et al., 2016; Zhang et al., 2021). Brain connectivity has likely evolved to maintain cost efficiency and optimized processing capacity (Bullmore and Sporns, 2009), reflecting principles of neural efficiency in large-scale brain organization (Bassett and Bullmore, 2017). It seems possible to define a theoretically “optimal” network for brain functioning, characterized by both local segregation and global integration. This optimum may be the “small world” network, which has indeed been confirmed in several studies with human subjects (Bassett and Bullmore, 2017; Douw et al., 2011; Liao et al., 2017). Some studies have reported that meditators and yoga practitioners have a relatively small world like organizational structures (Gard et al., 2014).
Beyond functional magnetic resonance imaging (fMRI), neurophysiological techniques like electroencephalography (EEG) and magnetoencephalography (MEG) offer valuable insights into brain network activity by capturing electrical and magnetic signals arising from neuronal postsynaptic potentials. MEG, with its precise temporal and spatial resolution, has proven effective for investigating neural oscillations and their interactions across brain regions (Velmurugan et al., 2014). Studies utilizing EEG (Cahn and Polich, 2006; Yordanova et al., 2020), fMRI (De Filippi et al., 2022; Manna et al., 2010) and MEG (Calvetti et al., 2021; Lardone et al., 2022) have demonstrated that meditation modulates activity and connectivity in networks responsible for attention, self-awareness, and self-monitoring.
Interestingly, brain connectivity and network topology seem to be predictive of functioning even during rest. Several studies have confirmed the presence of a robust “default mode network (DMN)” which is consistently active during rest (Damoiseaux et al., 2006; Power et al., 2011). Recent research has shown that several other brain networks known to become active during certain tasks can also be identified during rest. Among them are the fronto-parietal network (FPN) as well as the dorsal and ventral attention networks (AN). These networks seem to be stable networks, as they are connected even when no task is given (Dosenbach et al., 2007; Power et al., 2011). The topology of these resting-state networks has been associated with higher-order functions, including working memory and attention, but their specific roles in meditation remain underexplored (Douw et al., 2011).
Meditation practices are believed to influence large-scale brain networks, yet the underlying mechanisms remain poorly understood (Gibson, 2024; Zhang et al., 2021). To address these gaps, the present study investigates the brain network topology of two groups of meditators, advanced meditators (AM) and beginner meditators (BM) practicing “Surat-Shabda-Yoga (SSY)” meditation at different stages. Their topological features were compared with those of individuals with no meditation experience but practicing introductory “four chakra meditation.” For each participant, three functional networks (DMN, FPN, and AN) were constructed using coherence as a measure of synchronization/connectivity (Bastos and Schoffelen, 2016). Furthermore, to avoid potential biases induced by varying densities and sensitivity to edge weights, we used small world propensity (SWP) as a measure of small worldness, enabling robust topological evaluations (Muldoon et al., 2016).
Materials and Methods
Participants
We acquired the MEG signals of 12 advanced meditators (AM, 5 F) with a mean age of 48.7 years, ±SD 6.42, seven beginner meditators (BM, 4 F) with a mean age of 29 years, ±SD 6.1, and 10 control meditators (CM, 6 F) with a mean age of 21.2 years, ±SD 1.5. The demographic characteristics of participants, including age and gender distribution, are summarized in Table 1.
Demographic Characteristics of Participants
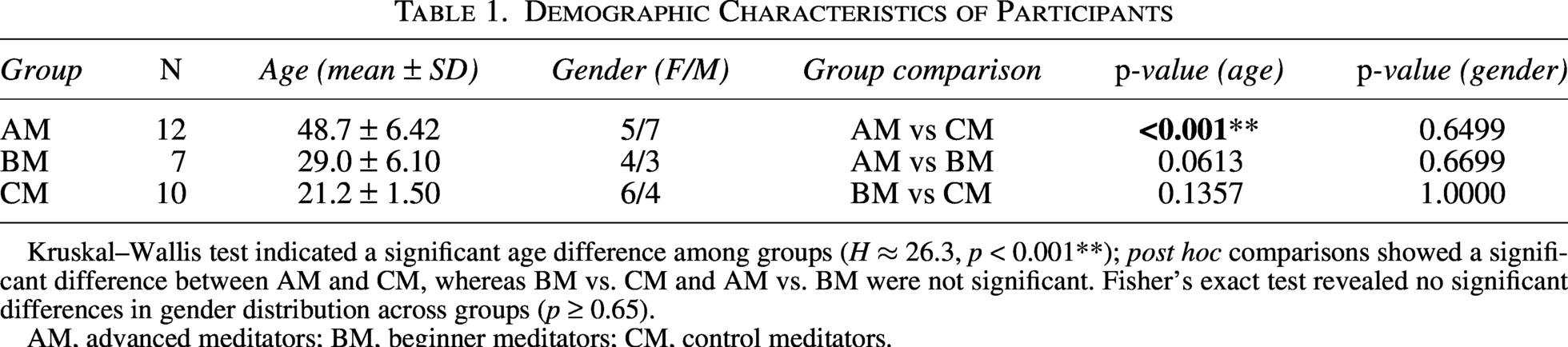
Kruskal–Wallis test indicated a significant age difference among groups (H ≈ 26.3, p < 0.001**); post hoc comparisons showed a significant difference between AM and CM, whereas BM vs. CM and AM vs. BM were not significant. Fisher’s exact test revealed no significant differences in gender distribution across groups (p ≥ 0.65).
AM, advanced meditators; BM, beginner meditators; CM, control meditators.
The CM participants were students at Dayalbagh Educational Institute (DEI) and had been introduced to basic meditation as part of their coursework in the comparative study of religion. This practice, known as “four chakra meditation,” involves focusing attention sequentially on the navel, heart, throat, and eye regions while repeating a mystical word (Sahab, 2004).
In the present study, meditators were classified into BM and AM groups according to the structured framework of SSY, as practiced in the Radhasoami tradition (Sahab, 2004; Satsangi et al., 2016). SSY meditation is rooted in the evolution of Hindu philosophy, which describes six chakras along the spinal cord. Unlike practices that primarily emphasize these lower centers, SSY directs attention toward higher-order reference points in the brain that function as focal centers for meditation. These reference points can be conceptually aligned with the attention network (AN), an ascending arousal system that modulates states of consciousness and regulates the activity of other neural systems (Tang et al., 2015). Furthermore, SSY practitioners report inner experiences that correspond to distinct frequency bands and reference centers, suggesting possible neurophysiological correlates (Satsangi et al., 2016). Thus, this practice provides phenomenological descriptions that closely parallel current neuroscientific models, making it a compelling system to investigate in contemporary brain research.
BMs in this study are practitioners in their initial stage, “sumiran dhyan”. This practice involves the focused repetition of a specific word or directing recurring attention towards it. AMs are practitioners who, after completing at least five years of BM practice, underwent a formal evaluation process. This process includes a structured questionnaire and interview, where their meditation-related experiences are reviewed by mentors. Advancement to the next stage requires evidence of sustained meditative depth and consistency. At this level, practitioners continue the basic practice of attentional practice while incorporating an inward focus on reference centers, known as “bhajan abhyas”, which also involves attending to inner auditory experiences. This resonance with frequency characteristics of the perceived internal sounds is pivotal, as it enables the practitioners to access and progress through successive meditative states.
We excluded one AM participant from the study due to noisy recordings (Gross et al., 2013). All participants provided written informed consent, and the study was approved by the research degree committee of DEI.
MEG system
The MEG data for this study were recorded using the Elekta Neuromag TRIUX system with 306 channels, located at National Brain Research Center (NBRC), Manesar, India. The system is housed in a magnetically shielded room, ensuring minimal interference from external electromagnetic fields during data acquisition. To monitor head movement during the recordings, five head-positioning indicator coils were attached to the participant’s scalp. These coils were activated and localized at the beginning of each recording session. For precise co-registration of MEG data with MRI images, a 3D digitization process was conducted. This process involved the use of a wooden chair, goggles with a transmitter, and a stylus with a receiver to capture the head surface geometry and the positions of fiducial points (nasion, left preauricular, and right preauricular). Electrooculogram and electrocardiogram (EKG) were co-recorded to aid artifact removal (Velmurugan et al., 2014). MEG data were acquired with a sampling frequency of 1,000 Hz, providing high temporal resolution. The recorded data were subsequently band-pass filtered between 0.5 and 45 Hz to isolate the frequency ranges relevant to neural oscillations and remove slow drifts and high-frequency noise.
Experimental procedures
MEG recordings were conducted during a 10-min meditation phase. CM participants practiced the basic “four chakra meditation” during the recordings, while AM and BM participants engaged in their respective meditation practices. The CM group served as the baseline group, as they had no formal meditation training beyond the introductory exposure provided in their coursework. During the recordings, no specific instructions were given to any group, allowing all participants to meditate in their usual manner.
Preprocessing
Preprocessing and data analysis were performed using Brain Electrical Source Analysis software (BESA; MEGIS Software GmbH, Munich, Germany). Initially, raw signals underwent spatial filtering, a method that defines spatial topographies describing artifacts such as EKG and allows the program to automatically scan and correct the data. The topographies were defined manually by marking a time range containing the artifact, typically around an EKG spike, or by creating a buffer with an averaged artifact. Specifically, EKG artifacts were averaged prior to defining the spatial topography for improved precision. Subsequently, a manual inspection was conducted to identify and remove noisy channels and segments of the recording with pervasive noise across all channels (Gross et al., 2013). This procedure yielded clean recordings for each participant. For subsequent functional connectivity analysis, we focused on the last 3- min of the 10-min meditation session to capture stabilized meditative states, minimizing the influence of transitional effects or initial adaptation to the experimental setup (Cahn and Polich, 2006; Lutz et al., 2004). The 3-min data were divided into three 1-min epochs, each further segmented into 5-sec sub-epochs. This segmentation was chosen to ensure reliable connectivity measures within each epoch while balancing temporal resolution and mitigating variability from noise or artifacts (Rojas et al., 2018).
Functional brain networks
We analyzed three widely recognized brain networks: DMN, FPN, and AN. The DMN is a well-documented network associated with self-referential thinking, introspection, and resting-state activity. It consists of 12 nodes representing key brain regions such as the medial prefrontal cortex and posterior cingulate cortex (Power et al., 2011). The FPN plays a crucial role in goal-directed attention and adaptive cognitive control and comprises 13 nodes, including the dorsolateral prefrontal and inferior parietal cortices (Dosenbach et al., 2007). The AN, responsible for sustained and selective attention, has 24 nodes distributed across regions like the frontal eye fields and superior parietal lobule (Power et al., 2011). While these networks are well-characterized in cognitive neuroscience, their roles in meditation practices remain under exploration. Furthermore, to measure connectivity among these networks, we used standard source montages within BESA software. Raw data segments were processed in the source analysis module, where source solutions for the DMN, FPN, and AN were loaded and saved as user-defined source montages. These montages ensured consistency across all participants during subsequent analyses.
Connectivity and network analysis
We estimated functional connectivity among the DMN, FPN, and AN using a connectivity measure (coherence). Connectivity measures in M/EEG research can be broadly categorized into linear and nonlinear approaches. Linear metrics such as coherence quantifies the frequency-specific linear relationship between two signals, capturing both amplitude and phase consistency within defined frequency bins. In contrast, nonlinear measures such as phase-locking value (PLV), mutual information, and related metrics estimate phase synchrony or general statistical dependencies beyond linear relationships (Bastos and Schoffelen, 2016; Sporns, 2022). While nonlinear methods are valuable for probing cross-frequency coupling or nonlinear interactions, our study was specifically designed to investigate frequency-specific group differences in predefined networks (DMN, FPN, AN). For this purpose, coherence provides a direct and interpretable characterization of functional connectivity that aligns with our experimental goals. Importantly, coherence has been successfully used in meditation neuroimaging studies under comparable designs, including recent MEG work demonstrating frequency-specific functional reorganization during eyes-open meditation (Pradeep Kumar et al., 2023).
For this study, connectivity (adjacency matrix) was measured at eight key frequencies: 4 Hz, 7.9 Hz, 12.83 Hz, 18.93 Hz, 25.33 Hz, 30.76 Hz, 37.36 Hz, and 45.37 Hz. These frequencies were selected from the coherence spectrum based on prespecified analysis parameters in BESA, which included a 1 Hz lower cutoff, a 50 Hz upper cutoff, and frequency and time resolutions of 0.25 Hz and 200 msec, respectively. For simplicity and consistency in presentation, these frequencies are interchangeably referred to as their rounded values: 4 Hz, 8 Hz, 13 Hz, 19 Hz, 25 Hz, 31 Hz, 37 Hz, and 45 Hz throughout this article. Frequencies from 4 to 8 Hz are associated with the theta (θ) band, 8 to 13 Hz with alpha (α), 13 to 30 Hz with beta (β), and above 30 Hz with gamma (γ), and these frequency bands are linked to various cognitive functions (Pavlov and Kotchoubey, 2022). The connectivity matrices generated through this process provided a comprehensive view of functional interactions among the networks, serving as the foundation for subsequent network analyses conducted in MATLAB (Mathworks®, version R2022b).
A graph is a topological representation of a network, constructed by nodes (“vertices”) and links (“edges”) between them (Douw et al., 2011). Graphs can be unweighted (binary) or weighted; in this study, we used undirected weighted graphs. Each node in the graphs corresponds to a specific brain region from the DMN, or FPN, or AN, and the edge weight reflects the coherence connectivity between the connected nodes. These network graphs enabled us to compute a wide range of network measures, with a focus on the global properties of the brain’s topological organization (Bullmore and Sporns, 2009).
A key measure used in this study was SWP, which quantifies the extent to which a network exhibits small-world topology (Muldoon et al., 2016). Small-world networks are characterized by high local clustering and short path lengths, which are commonly observed in human brain networks and are thought to underlie cognitive processes, including cognitive insights (Bassett and Bullmore, 2017; Liao et al., 2017; Schilling, 2005). SWP, Φ, was calculated by comparing the observed network’s clustering, 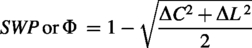
Where
SWP offers several advantages over traditional methods such as the small-world index (SWI). Unlike SWI, SWP allows for unbiased comparisons across networks with varying densities and is sensitive to edge weights, making it particularly suited for coherence-based functional network analysis (Muldoon et al., 2016). This sensitivity ensures that subtle differences in connectivity are captured, enabling a more nuanced understanding of brain networks. In this study, SWP, a measure of neural efficiency, was used to compare the meditator and control groups.
Statistical analysis
All statistical analyses were performed in MATLAB. To compare the meditators and control groups across each frequency and network, we employed Wilcoxon rank sum test, nonparametric alternative to independent t-test (Hollander et al., 2014). This test was chosen due to its suitability for the requirements of the present study. Specifically, we calculated the SWP for the last 3 epochs of each individual at a given frequency and network. For each comparison, we had 33 SWP values (11 subjects × 3 epochs) for AM, 21 SWP values (7 subjects × 3 epochs) for BM, and 30 SWP values (10 subjects × 3 epochs) for CM. Since the BM group sample size was less than 30, we did not test for normality and directly opted for the nonparametric/rank-sum test. This decision aligns with the test’s strength in handling non-normally distributed data, uneven group sizes, and sensitivity to outliers, ensuring robust and reliable comparisons (Hollander et al., 2014). To address the issue of multiple comparisons, we applied the false discovery rate (FDR) correction, a widely accepted method that controls the expected proportion of false positives (Benjamini and Hochberg, 1995). This approach is particularly effective in neuroimaging studies, where multiple comparisons are performed across frequencies and networks. The significance level was set at p ≤ 0.05 after FDR correction, ensuring that the reported results are both statistically rigorous and reliable.
Results
Table 1 presents the demographic comparison across groups. Kruskal–Wallis test revealed a significant difference in age among the three groups (H ≈ 26.3, p < 0.001**). Post hoc comparisons showed that the AM group was significantly older than the CM group, while no significant age differences were found between AM vs. BM or BM vs. CM. Fisher’s exact test showed no significant group differences in gender distribution (p ≥ 0.65).
To analyze the possible effects of meditation on brain network organization, we compared the brain topology of meditator groups (AM and BM) with that of the control group (CM) during meditation. Small-world network topology, measured using SWP, was evaluated across three functional networks: the DMN, the FPN, and the AN (see Fig. 1). BrainNet Viewer was employed to visualize the spatial distribution and organization of the three functional networks on the brain surface, providing a detailed representation of the DMN, FPN, and AN across various perspectives (Qin et al., 2021; Xia et al., 2013).
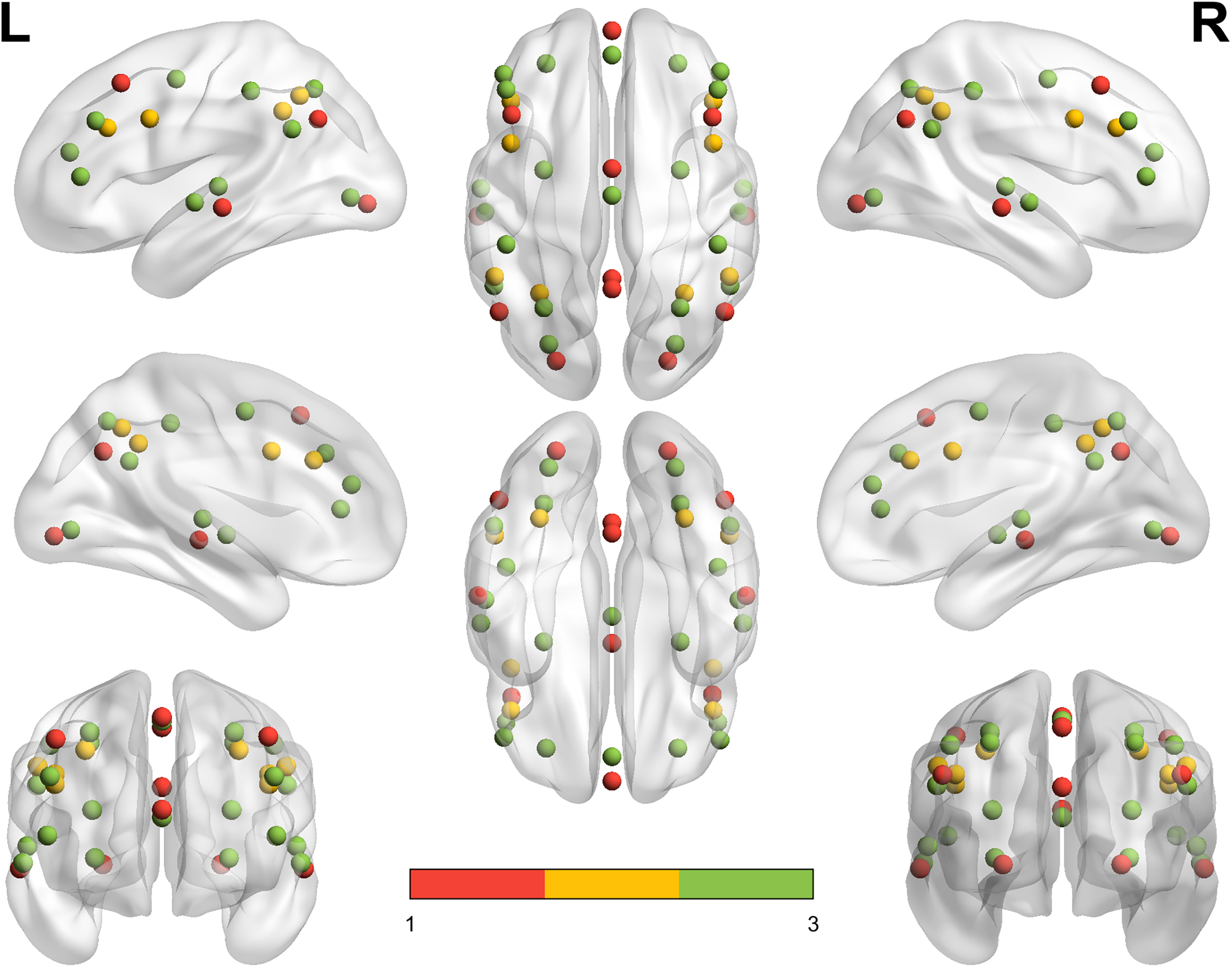
Visualization of functional networks (DMN, FPN, and AN) on the brain surface using brainNet viewer. The figure provides a comprehensive view of the brain, showcasing lateral, medial, dorsal, ventral, anterior, and posterior sides of both hemispheres. The visualization was created using BrainNet Viewer with the ICBM152 smoothed surface template, designed to support the mapping of MNI coordinates for precise node placement. The three functional networks are represented with color-coded nodes: default mode network (DMN) in red, fronto-parietal network (FPN) in yellow, and attention network (AN) in green. This full-brain view highlights the spatial distribution and organization of these networks.
Connectivity, assessed using coherence, was evaluated at eight frequencies (4 Hz, 8 Hz, 13 Hz, 19 Hz, 25 Hz, 31 Hz, 37 Hz, and 45 Hz). For each network-frequency pair, SWP values from the AM group (33 data points) were statistically compared with those from the CM group (30 data points). This resulted in 24 pairwise comparisons (3 networks × 8 frequencies). Among these comparisons, a significant difference was observed between the AM and CM groups in the AN at 19 Hz (pFDR = 0.0069), which falls within the β frequency range (see Fig. 2). This result suggests that meditation practice enhances neural efficiency (increased SWP) in the AN at β frequency. No other comparisons between AM and CM were statistically significant.
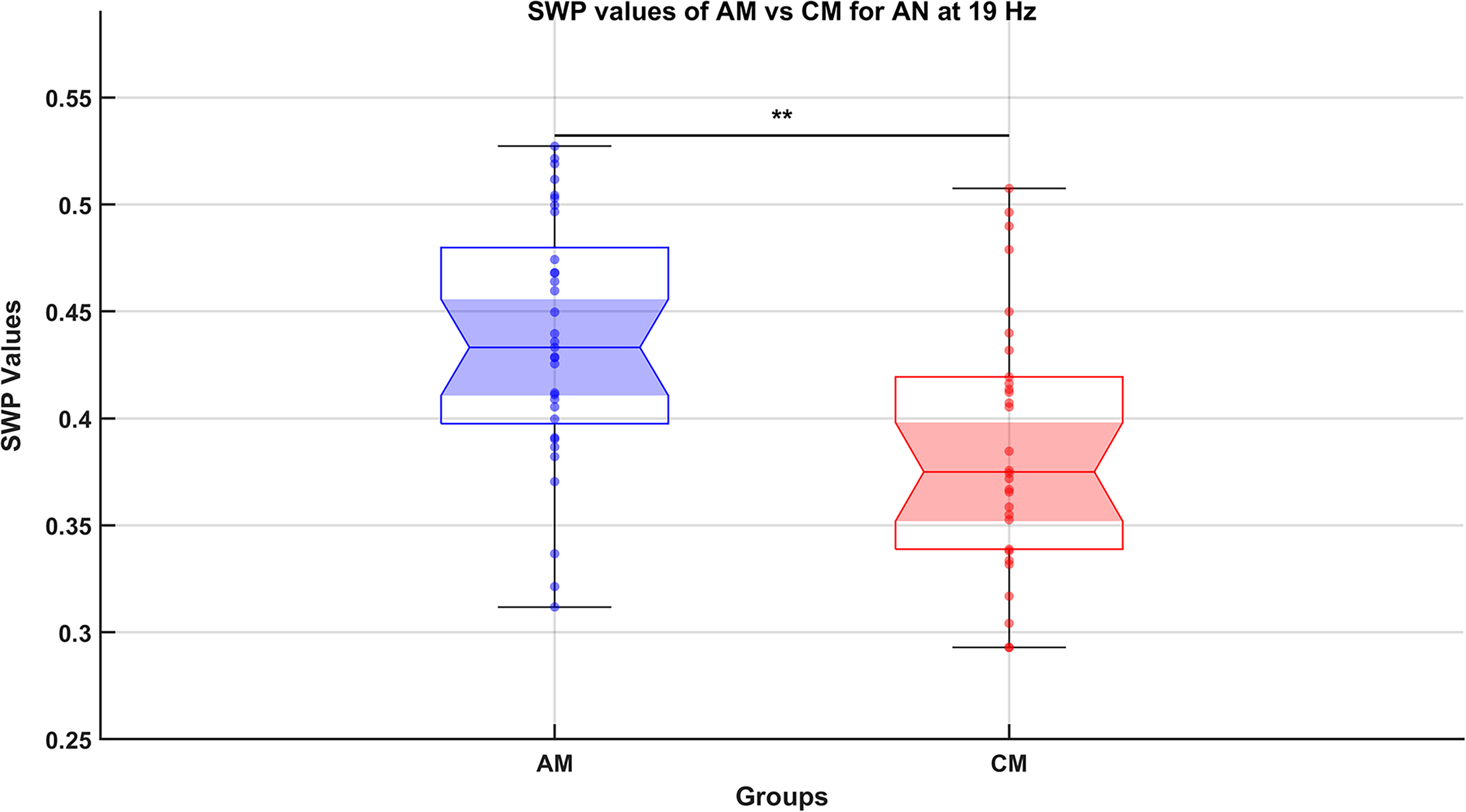
Comparison of small-world propensity (SWP) values between advanced meditators (AM) and control meditators (CM) groups for the attention network (AN) at 19 Hz (falls within β frequency range). This notched box plot represents the SWP values, a measure of neural efficiency in the functional network AN at 19 Hz, comparing the AM and the CM groups. The functional network was derived using coherence-based connectivity measures across multiple frequencies (4 Hz, 8 Hz, 13 Hz, 19 Hz, 25 Hz, 31 Hz, 38 Hz, and 45 Hz), with significant differences observed only at 19 Hz for the AN. Data points are based on three 1-min epochs from the final 3 min of a 10-min meditation session, yielding 33 data points (11 subjects × 3 epochs) for AM and 30 data points (10 subjects × 3 epochs) for CM. The notched design allows visualization of the median and its confidence interval, alongside individual data points and potential outliers, providing insights into group variability. The nonparametric rank-sum test, applied to compare groups, was followed by false discovery rate (FDR) correction to account for multiple comparisons. Statistical significance (FDR-corrected p < 0.01) is indicated by double asterisks (**). The results show higher SWP values in the AM group compared with the CM group, suggesting enhanced neural efficiency within the AN at 19 Hz of AM.
Similarly, comparisons between the BM (21 data points) and CM groups (30 data points) revealed one significant difference in the FPN at 8 Hz (pFDR = 0.0375), within the θ frequency range (see Fig. 3). The higher SWP values observed in BM suggest enhanced neural efficiency in the FPN during meditation at θ frequency. No significant differences were found in the remaining 23 comparisons among the networks and frequencies.
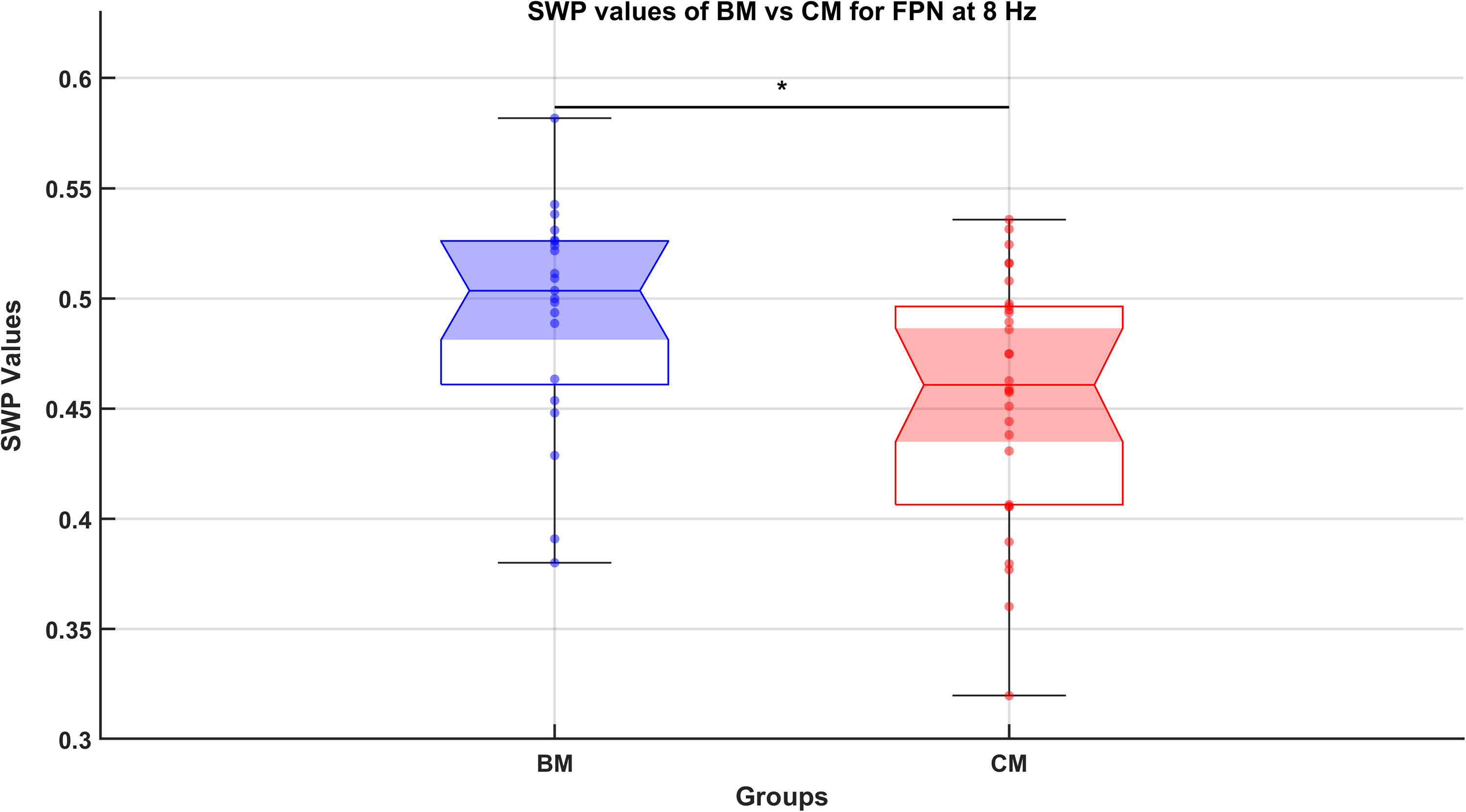
Comparison of small-world propensity (SWP) values between beginner meditators (BM) and control meditators (CM) groups for the fronto-parietal network (FPN) at 8 Hz (falls within θ frequency range). The figure presents a notched box plot comparing SWP values, which measures neural efficiency of functional brain networks, between BM and CM for the FPN at 8 Hz. Connectivity analysis was performed using coherence measures across frequencies (4 Hz, 8 Hz, 13 Hz, 19 Hz, 25 Hz, 31 Hz, 38 Hz, and 45 Hz), with a significant group difference observed at 8 Hz. BM group data include 21 points, derived from 7 subjects over three epochs (1 min each) from the final 3 min of a 10-min meditation session. CM group data consist of 30 points from 10 subjects with a similar epoch division. The notched box plot showcases the median, interquartile ranges, confidence intervals for the medians, and potential outliers, along with all data points plotted for clarity. A rank-sum test was used for statistical comparison, with corrections for multiple comparisons applied via the false discovery rate (FDR) method. A significant difference at p < 0.05 (FDR-corrected) is indicated by *. The higher SWP values in BM compared with CM suggest enhanced neural efficiency within the FPN at 8 Hz.
Furthermore, to enhance network-level interpretability, we computed average adjacency matrices for the most significant contrast (AM vs. CM) in the AN at 19 Hz (β-band). The AN node coordinates were defined as the node file, and the edge weights were derived from the averaged adjacency matrices of each group. Visualization was performed using BrainNet Viewer, generating lateral, medial, and dorsolateral views of AN nodes and their connectivity for AM and CM groups (Fig. 4a,b) (Xia et al., 2013). In addition, a thresholder graph plot (circular layout, edge threshold = 0.34) was constructed to depict retained functional connections, enabling a direct visual comparison of network topology between AM and CM (Fig. 4c). The AM group demonstrated denser local clustering and shorter path lengths relative to the CM group, consistent with its higher SWP (0.4846 vs. 0.2929). Together, these visualizations complement the statistical results and provide an intuitive representation of group-level differences. Importantly, the observed AN topology at β-band frequencies aligns with its established role in attention and cognitive flexibility, supporting a functional interpretation of the SWP differences (Markett et al., 2022; Power et al., 2011).
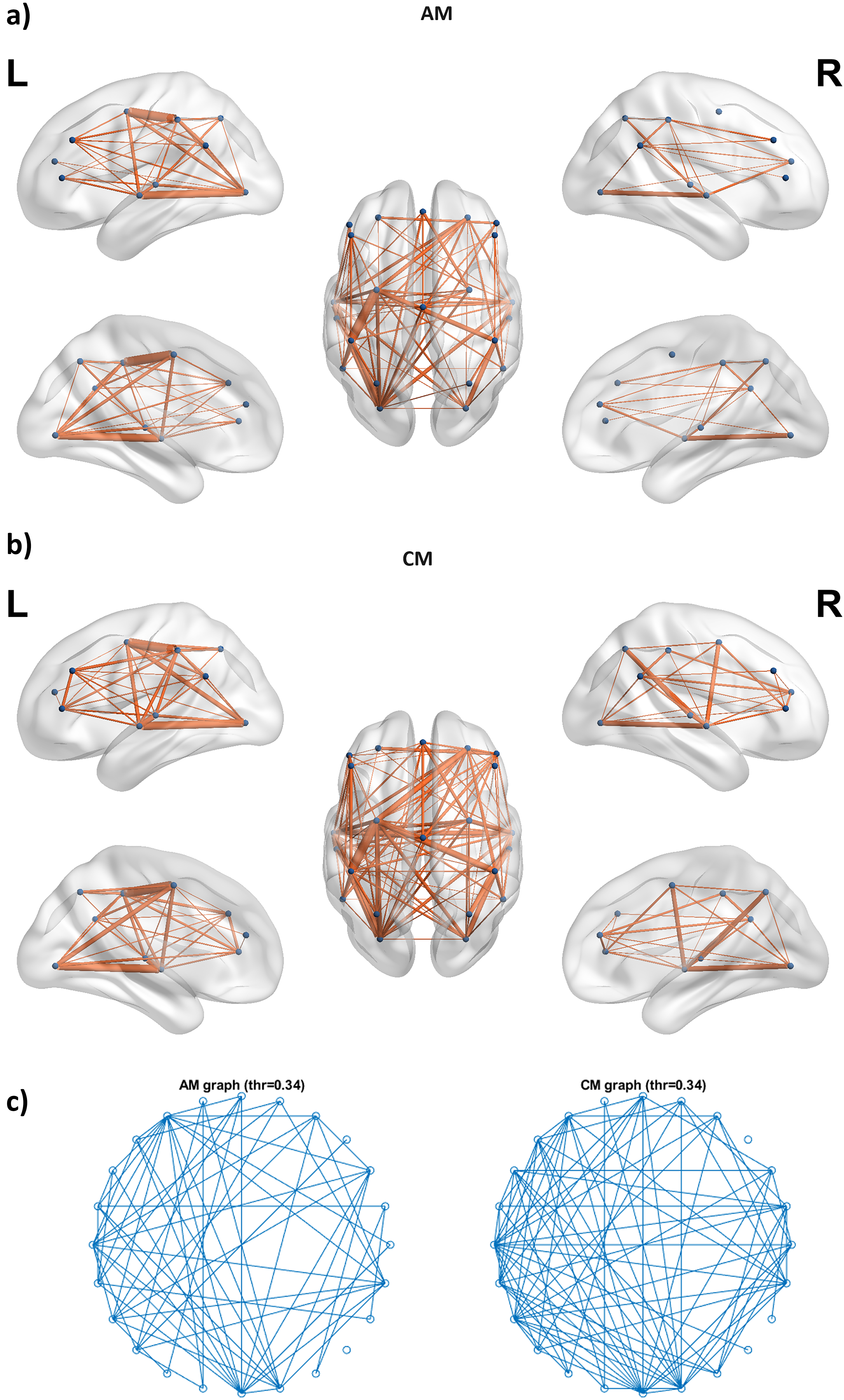
Lateral, medial, and dorsolateral views of the attentional network (AN) nodes with average connectivity (edge weights from adjacency matrices) for the last three epochs across:
Overall, these findings reveal that meditation engages the brain’s intrinsic network architecture in a frequency- and stage-specific manner. Notably, increased SWP or enhanced neural efficiency was observed in the AN at 19 Hz in AM and in the FPN at 8 Hz in BM, each relative to CM.
Discussion
The present study aims to investigate the neural efficiency by measuring SWP during meditation phase in two groups of meditators, AM and BM, practicing distinct stages of SSY and to compare them with individuals in control group CM with no prior meditation experience.
As expected, the AM group was significantly older than the CM group. This difference arises naturally because eligibility for advanced practices in SSY requires prior completion of at least five years of BM practice and progression to advanced stages based on spiritual competence (Satsangi et al., 2016). In contrast, the CM group primarily comprised student participants with only introductory exposure to meditation as part of coursework. Prior research indicates that small-world properties of brain networks tend to decline with aging and are associated with age-related reductions in cognitive performance (Hong et al., 2016). Therefore, age-related effects alone would predict lower SWP in the AM group relative to CM. However, our results demonstrate the opposite pattern: AM participants exhibited significantly higher SWP in the AN compared with CM. This finding suggests that the observed differences cannot be explained by aging and instead point toward meditation practice as the main factor enhancing neural efficiency.
Our findings indicate that meditator groups (AMs and BMs), compared with control group (CM), exhibit distinct brain network topology as measured by SWP, a graph-theoretical parameter. Beyond its description as a graph-theoretical property, SWP can also be interpreted as a marker of neural efficiency. Small-world organization reflects an optimal balance between functional segregation, supported by high local clustering, and functional integration, enabled by short characteristic path lengths (Bassett and Bullmore, 2017; Muldoon et al., 2016). Networks exhibiting this balance are thought to support efficient information processing by enabling specialized local computations while maintaining rapid global communication.
Within this framework, the increased SWP observed in the AN of AM at the β-band frequency (19 Hz), and in the FPN of BM at the θ-band frequency (8 Hz), relative to CM, suggests stage- and frequency-specific enhancements in neural efficiency (Figs. 2–4).
The AN play a critical role in cognitive functions such as attention management and cognitive flexibility (Markett et al., 2022; Power et al., 2011). Its reorganization in AMs may reflect enhanced capacity for sustained and flexible attentional control. Moreover, β-band oscillations are closely tied to active engagement, top-down regulation, and decision-making processes (Engel and Fries, 2010; Spitzer and Haegens, 2017). Taken together, the β-band small-world increase in AN plausibly represents a neural mechanism that enables AMs to maintain stable focus while flexibly shifting across meditative states, processes closely related to attentional regulation and cognitive flexibility. This interpretation aligns with the advanced stages of SSY practice (bhajan abhyas), where practitioners engage in inward auditory focus and heightened attentional regulation (Satsangi et al., 2016). These results converge with prior work showing meditation-related reorganization of large-scale networks, including attention-related systems, together with frequency-specific topological changes across frequency bands (Lardone et al., 2022).
The FPN play a critical role in cognitive functions such as cognitive control, task set maintenance, and adaptive responses (Corbetta and Shulman, 2002; Dosenbach et al., 2008; Vincent et al., 2008). The observed small-world topology within the FPN at θ (8 Hz) frequency in BMs compared with CMs suggests more efficient information processing and connectivity within this network (Bassett and Bullmore, 2017; Muldoon et al., 2016). Moreover, θ frequencies are associated with introspection, memory processing, and cognitive control (Cavanagh and Frank, 2014; Herweg et al., 2020). This enhanced topology may allow BMs to direct their attention effectively to maintain focus along with managing the distractions, functions that overlap with cognitive control and task-set maintenance. In line with this evidence, these capabilities are crucial for the meditative practice of SSY, where the initial stage (sumiran dhyan) emphasizes withdrawing attention from external distractions and focusing inwardly while mentally repeating a mystical word (Satsangi et al., 2016).
Beyond their relevance to specific stages of SSY, the frequency-specific effects observed in the θ and β bands reflect neural mechanisms that are consistently implicated across contemplative traditions emphasizing attentional regulation and cognitive control. Focused attention (FA) practices, such as mantra- or breath-based meditation, are commonly associated with increased θ-band activity and enhanced fronto-parietal coordination, reflecting sustained attentional engagement and top-down control processes (Barrows et al., 2024; Cavanagh and Frank, 2014; Tang et al., 2015). In line with this literature, the increased neural efficiency observed within the FPN of BM at θ frequency suggests that early-stage SSY engages attentional control mechanisms that are broadly shared with FA traditions, while extending prior findings by demonstrating their expression at the level of large-scale network topology rather than solely oscillatory power or synchrony.
In contrast, open monitoring practices are more frequently associated with α-band modulation, reduced DMN coherence, and increased flexibility in switching between intrinsic and task-positive networks (Barrows et al., 2024; Devaney et al., 2021; Lardone et al., 2022). The absence of significant α- or DMN-related small-world effects in the present study is therefore consistent with the attentional demands and practice structure of SSY, which emphasize sustained, goal-directed attentional engagement rather than nonreactive monitoring of experience. This distinction highlights style-specific differences across contemplative traditions while reinforcing the interpretation that the observed θ- and β-band network reorganization in SSY reflects mechanisms aligned with focused attentional control.
Overall, these results reveal a progressive adaptation in network topology across meditation stages. Interpreting SWP as an index of neural efficiency places the present findings within a broader network neuroscience framework, in which large-scale brain systems such as the AN and FPN support adaptive cognitive functions including sustained attention and cognitive control. In this view, meditation-related increases in small-world organization reflect more efficient coordination within these systems (see Supplementary Table S1 for a comparative overview of recent EEG/MEG studies).
Conclusion, Limitations, and Future Directions
Our findings must be considered in the light of several limitations. One limitation of this study is the relatively small number of participants in the BM group (n = 7), which restricts broad generalizability. To address this, we segmented each 10-min meditation session into 1-min epochs and focused on the final 3 min, representing stable meditative states (Cahn and Polich, 2006; Lutz et al., 2004). This procedure yielded 21 epochs for the BM group, thereby increasing the number of data points available for analysis. However, these epochs are repeated measures within individuals and are not equivalent to additional independent subjects. Thus, the limitation of a small participant sample remains. However, to mitigate the risk of Type I error due to within-subject dependence, we employed nonparametric rank-based tests and applied FDR correction for multiple comparisons. Importantly, topological measures such as SWP are designed to capture subtle but reliable group differences even with limited samples (Muldoon et al., 2016). Furthermore, the structured progression of SSY practice provides clear and internally consistent group categorization, which adds confidence to within-group comparisons. Comparable sample sizes have also been reported in other meditation neuroimaging studies (Tang et al., 2015). Thus, while caution is warranted in extending these findings to larger populations, the results remain robust within the current framework, supported by both methodological design and statistical evidence.
While coherence provides a well-established and interpretable measure of frequency-specific functional connectivity, it primarily captures linear relationships between neural signals. Future studies could extend the present framework by incorporating nonlinear and cross-frequency connectivity metrics, such as phase–amplitude coupling, PLV, or mutual information, to probe interaction dynamics that are not accessible through coherence alone (Bastos and Schoffelen, 2016; Sporns, 2022). Such approaches may be particularly relevant for examining resonance-like interactions across frequency bands, which have been hypothesized to support integrative states during meditative practice and may complement the frequency-specific topology effects observed here.
In addition, the present analysis focused on measuring neural efficiency using SWP, without explicitly examining node-level graph metrics. Future work could incorporate measures such as degree centrality or nodal clustering within attention- and control-related networks to identify hub regions that contribute most strongly to observed group differences (Bullmore and Sporns, 2009; van den Heuvel and Sporns, 2011). Identifying such hubs would provide anatomically specific regions of interest for targeted analyses and enable investigation of how highly connected nodes participate in nonlinear or cross-frequency coupling during meditation (Bastos and Schoffelen, 2016; Canolty and Knight, 2010). Together, combining node-level characterization with advanced connectivity metrics may offer deeper insight into how meditation reorganizes brain networks to support efficient, coordinated, and potentially resonance-based neural communication.
Beyond these limitations, it is worth emphasizing that SSY is a contemplative practice that appears to resonate with the brain’s intrinsic architecture. By engaging large-scale brain networks in a stage- and frequency-specific manner, it reflects a natural process for cultivating cognitive insights and enhancing intellectual capacity (Schilling, 2005). While our findings focus on measuring neural efficiency, the underlying meditative philosophy of SSY aims toward attaining “param-anand”, a state of ultimate bliss and transcendence (Satsangi et al., 2016). In this view, mental clarity and psychological well-being are not isolated outcomes but emerge naturally as byproducts of the deeper spiritual progression. Thus, this study highlights not only the cognitive and regulatory benefits of meditation but also its potential role in facilitating holistic human development. Future research may further investigate how these philosophical dimensions are mirrored in neural function.
Authors’ Contributions
C.M.M.: Supervision, writing—review, and data curation; G.P.C.D.: Conceptualization, analysis, software validation, visualization, and writing—original draft, review, and editing; M.T.: Data curation and writing—review; S.A.: Investigation, data curation, and writing—review.
Data Availability
Due to Institutional Review Board (IRB) restrictions, raw participant data cannot be publicly shared. Derived connectivity data and analysis scripts are openly available at OSF (https://osf.io/8yu5k/). Additional data are available from the corresponding author upon reasonable request.
Footnotes
Acknowledgments
The authors are deeply grateful to Prof. P.S. Satsangi, Emeritus Chairman, Advisory Committee on Education, DEI, for visionary talks and novel experimentation that has inspired this research. We wish to thank all the participants who volunteered for this study. We acknowledge the support received from the Director of DEI for undertaking this study. We are obliged to Dr. Manjari Tripathi for access to MEG facility at NBRC. We are also thankful to Dev Saran Sujan, an undergrad, for his contributions in scripting. Damisetti Geeta Prem Chandoo thanks Rajya Anand Sansthan (RAS) for providing a doctoral fellowship (RAS/2/ADF/I/11/2022).
Author Disclosure Statement
Authors declare that they have no conflict of interest.
Funding Information
This research did not receive any specific grant from funding agencies in the public, commercial, or not-for-profit sectors.
Supplemental Material
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.