
Abstract
The developmental hazard screening of oligonucleotide therapeutics (ONTs) presents challenges due to their frequent lack of pharmacology in nonclinical species and embryo–fetal exposure is presumed to be limited in vivo. This study demonstrates that direct culture in media containing mipomersen, a 2′-O-methoxyethyl phosphorothioated antisense oligonucleotide (ASO), results in dose-responsive morphological changes in rat and rabbit whole-embryo culture (WEC). Automated miRNAscope in situ hybridization was used to confirm dose-dependent embryonic exposure and visualize the distribution pattern of mipomersen in the embryo and extraembryonic membranes, suggesting that ONTs may enter the umbilical vasculature and pass into embryo circulation. Neither microinjections nor assisted transfections were required to achieve embryonic exposure. These findings support the utility of WEC as a new approach method (NAM) for developmental hazard identification of ONTs. WEC could complement or partially replace in vivo studies, reducing animal use and required test material amounts, while enabling robust developmental hazard identification for ONTs. This work informs future safety assessment strategies and regulatory guidance for ONTs.
Introduction
The emergence of oligonucleotide therapeutics (ONTs) has challenged the pharmaceutical industry to develop new strategies and standard practices for non-clinical safety assessment of this modality, which has the characteristics of both small molecules and biologics. Biodistribution of ONTs in developmental and reproductive toxicity (DART) assessments is an important consideration, as nearly all marketed ONTs have been detected in placentas of non-clinical species but generally have not been detectable in the fetal liver, kidney, or homogenized whole fetus. 1 However, two marketed GalNAc siRNAs, LEQVIO (inclisiran sodium) and RIVFLOZA (nedosiran sodium), have been detected in fetal plasma. 1 Depending on ONT-specific dosing strategies and in vivo DART study design considerations, it may not be feasible to achieve meaningful ONT exposure throughout the entire period of organogenesis.
Whole-embryo culture (WEC) is an ex vivo hazard identification screening assay and is considered a new approach method (NAM). WEC allows rodent or rabbit embryos to be directly exposed to the test item during an early critical period of major organogenesis, offering the opportunity to perform developmental hazard assessment for ONTs that may not achieve meaningful fetal exposure during in vivo DART studies or when surrogate molecule use is necessary. It was previously thought that microinjection into the amniotic cavity, the embryo itself, or the largest yolk sac blood vessels was necessary to ensure exposure to the embryo due to the larger size of ONTs. 2 Previous studies have successfully evaluated ONTs in mouse WEC using a microinjection approach.3–6 In contrast, WEC studies evaluating small molecules are regularly performed, as they are presumed to be small enough to diffuse into the yolk sac cavity and be absorbed into the embryo. While it is unlikely that ONTs could diffuse through the yolk sac, endocytosis could potentially allow for their uptake resulting in exposure to embryos cultured in media containing ONTs, 7 given gymnotic exposure was successful. This approach is much simpler in terms of technical execution compared with the microinjection approach and prevents experimental artifacts that may interfere with interpretation. Demonstration that microinjection is not necessary for ONTs would greatly increase the utility of WEC studies for developmental toxicity hazard assessments.
KYNAMRO (mipomersen), a 20-nucleotide long, 2′-O-methoxyethyl phosphorothioated ASO (2′-MOE ASO), was used to investigate the utility of rat and rabbit WEC to assess ONTs using unassisted transfection. Mipomersen was approved by the FDA to treat homozygous familial hypercholesterolemia and is an inhibitor of apolipoprotein B-100 synthesis, a protein essential for the production of low-density lipoprotein, although it was later withdrawn from the market due to unacceptable risk of hepatotoxicity. 8 Mipomersen is not pharmacologically active in rats or rabbits. 9
For the first time, we show concentration-dependent embryonic exposure to an ONT following culture in media containing 2′-MOE ASO (mipomersen) alone, with no microinjection, for both rat and rabbits on gestation day (GD) 11. Exposure was confirmed via off-target dose-responsive effects on yolk sac and/or embryo parameters and automated miRNAscope in situ hybridization showing distribution of mipomersen in embryos and yolk sac umbilical vasculature at the point of connection with the embryo.
Materials and Methods
Rat husbandry and WEC
Rat embryos were cultured according to established techniques. 10 Untreated time-mated female Sprague Dawley rats (∼10–12 weeks of age, supplied by Charles River, Laboratories, Inc., Raleigh, NC) were used for this study. Pregnant dams were approximately GD 5 upon arrival (day of mating is designated as GD 0). Mated female rats were housed in Allentown NexGen reusable individually ventilated caging system with clear plastic cages containing Alpha-dri™ bedding (Shepherd Specialty Papers, Inc., Kalamazoo, MI) in a controlled environment (68°F–79°F; 30% to 70% relative humidity) with an approximate 12-h light/12-h dark cycle. Food (Certified Rodent Diet #5001, PMI Nutrition International Brentwood, MO) was provided ad libitum. Filtered tap water (supplied and periodically tested by American Water) was available ad libitum from an automatic watering system. Rat enrichment (plastic or cardboard hut or tube, wood gnawing block) was provided and documented. Available information indicated that no substance was present in the diet, drinking water, enrichment, or bedding at a concentration likely to influence the outcome of this study.
Dams were anesthetized to a deep plane of anesthesia with isoflurane and euthanized via exsanguination in the afternoon on GD 9. Death was ensured by thoracotomy as a secondary form of euthanasia. Embryos were dissected out of the uterus for subsequent WEC. Using a dissecting microscope, the Reichert’s membrane was removed and embryos at the appropriate stage of development (i.e., late headfold stage and 1 or 2 somite pairs) were chosen for the experiment. 11 Selected embryos were placed into prewarmed, sterile bottles containing approximately 2.0 mL culture media (70% heat-inactivated rat WEC serum, 30% Tyrode’s solution, 35 μg/mL streptomycin) as well as either vehicle control (0.04% phosphate-buffered saline [PBS]) or 1, 10, 50, or 100 µM mipomersen (WuXi). The culture bottles were maintained at 37°C ± 0.5°C in an incubator (Cullum Starr Precision Engineering Limited, Cambridge, England) for approximately 40 h. The bottles rotated at a rate of approximately 30 rotations per minute and were continuously gassed using an intermediate low flow regulator set at a final flow rate of approximately 30 cc/min with scheduled, increasing oxygen concentrations from 5% to 10% to 20% O2 (with a constant 5% of CO2 and the balance of nitrogen between 75% and 90%). Ninety-four embryos were individually cultured, 30 vehicle control and 16 treated per group. Embryos were distributed such that each litter was represented in each group and each group was distributed between two or three incubators. At the end of the culture period on GD 11, embryos were examined for viability, morphology, size (crown-rump length), and somite number using a modified version of a morphological scoring system. 12 In addition, visceral yolk sacs (VYS) were evaluated for morphology and size (diameter) before embryo evaluation.
Rabbit husbandry and WEC
Untreated time-mated female New Zealand White rabbits (approximately 6 to 9 months of age, supplied by Envigo Global Services, Inc., Denver, PA) were used for this study. Pregnant does were approximately GD 3 to 5 upon arrival (day of mating designated as GD 0). Mated female rabbits were housed individually in stainless steel Allentown Rabbit Euro Cages with plastic fenestrated flooring in a controlled environment (61°F–72°F; 30%–70% relative humidity) with an approximate 12-h light/12-h dark cycle. Food [Rabbit Chow (HF) #5322, PMI Nutritional International, Brentwood, MO] was provided (approximately 125 g daily). Filtered tap water (supplied and periodically tested by American Water) was available ad libitum from an automatic watering system. Animals received appropriate music for a period during the light hours. Rabbit enrichment (plastic resting board with hide, wood gnawing block, stainless steel bell, nylon dumbbells) was provided and documented. Available information indicates that no substance is present in the diet, drinking water, or enrichment at a concentration likely to influence the outcome of this study.
Does were euthanized in the morning on GD 9 via intravenous overdose of Fatal Plus® (pentobarbital sodium) via the marginal ear vein. Death was ensured via exsanguination as a secondary form of euthanasia. Embryos were dissected out of the uterus and the bed of chorionic tissue was removed, such that only the embryo, amnion, and attached VYS were included for subsequent WEC. Criteria for placement into culture were as follows: clearly developed eyes, embryos positioned all on the same side of the yolk sac, intact vessel encircling yolk sac, visible blood flow, and lack of abnormalities. Selected embryos were placed into prewarmed, sterile bottles containing approximately 2.5 mL culture media [75% heat-activated rabbit WEC serum, 25% PBS, 10 µg/mL streptomycin, 10 µL/mL penicillin, 2 mg/mL glucose] as well as either vehicle control (0.04% PBS) or 1, 10, 50, or 100 µM mipomersen (WuXi). The culture bottles were maintained at 37°C ± 0.5°C in an incubator (Cullum Starr Precision Engineering Limited, Cambridge, England) for approximately 45 h. The bottles rotated at a rate of approximately 30 rotations per minute and were continuously gassed using an intermediate low flow regulator set at a final flow rate of approximately 30 cc/min with scheduled, increasing oxygen concentrations from 20% to 95% O2, (with a constant 5% of CO2 and the balance of nitrogen between 0% and 75%). Ninety-four embryos were individually cultured, 30 vehicle control and 16 treated per group. Embryos were distributed such that each litter was represented in each group, and each group was distributed between two or three incubators. At the end of the culture period on GD 11, embryos were examined for morphology, size (crown-rump length), and somite number using a modified version of a morphological scoring system.13,14 In addition, VYS were evaluated for morphology and size (diameter) before embryo evaluation.
Embryo evaluation of morphology and fixation
On GD 11, rat and rabbit embryos were dissected from the yolk sac and amnion, which were retained, and evaluated for viability, morphology, size, and somite number. VYS were evaluated for morphology and size. Dead and/or grossly malformed embryos were only evaluated for viability. Embryos found outside the yolk sac following the culture period were not evaluated as these were considered experimental artifacts.
The following regions were evaluated for both rat and rabbit: yolk sac, embryo rotation, somite, caudal, forelimb, hind limb, neural tube, heart, pharyngeal arches, and craniofacial. Supplementary Tables S2 and S4 list all the detailed potential observations for each region. In the heart region, observations of dextrocardia were recorded in the raw data, but not summarized in Table 1, because this finding is considered an artifact of culture and not treatment related.
Observational Macroscopic Scoring of WEC Samples
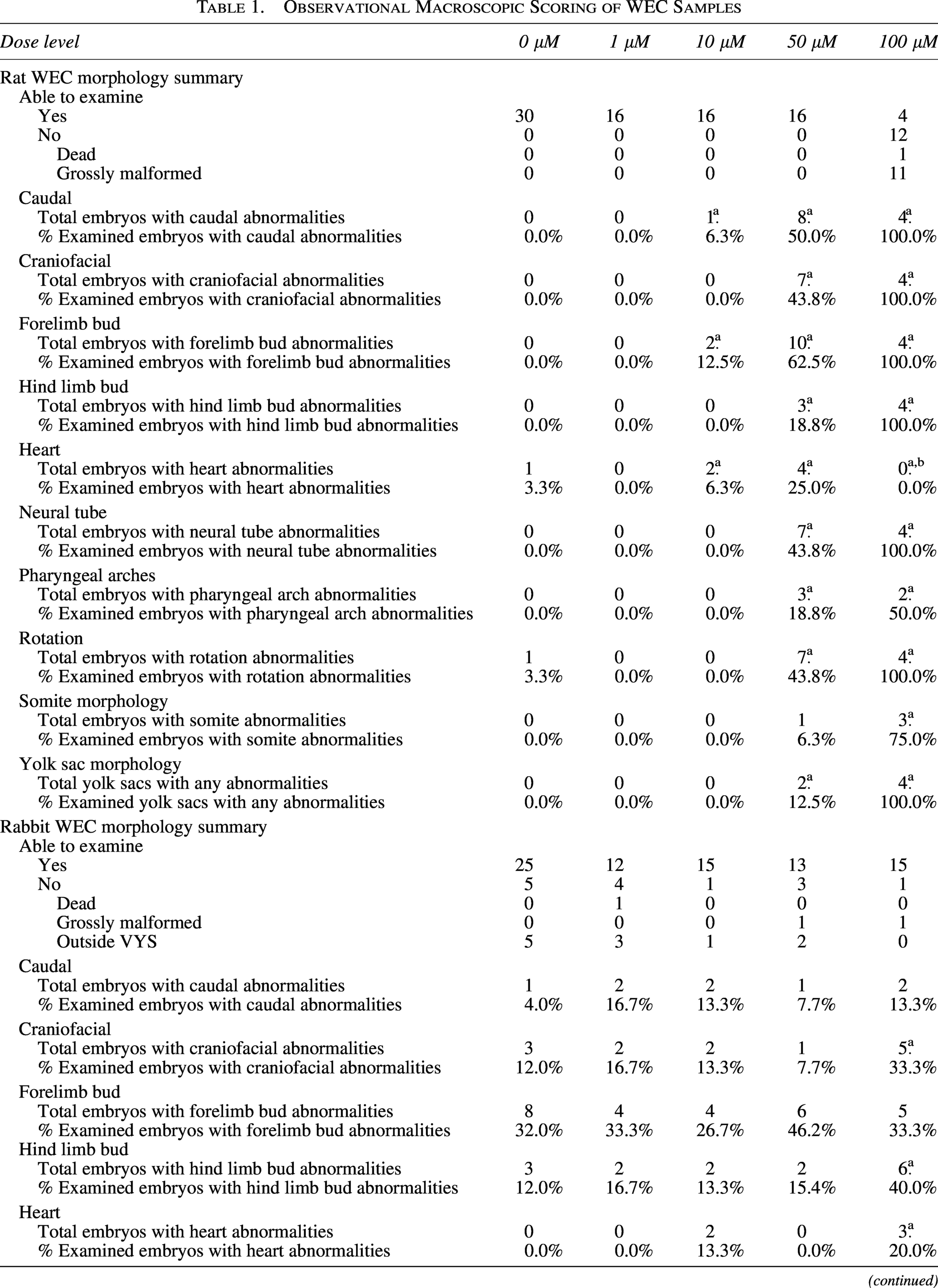
Mipomersen-related observation.
Although no heart observations were recorded at 100 µM in the examined embryos, this region is considered impacted by mipomersen due to the dose response seen at lower doses and the high percentage (68.8%) of grossly malformed embryos at this dose. Heart abnormalities likely occurred in grossly malformed embryos that were not captured in the data because grossly malformed embryos were not examined.
VYS, visceral yolk sacs; WEC, whole-embryo culture.
Following this evaluation, all embryos and their extraembryonic membranes were fixed for 24 h in 10% neutral buffered formalin, initially orientated in HistoGel (Catalog # HG-4000-012, Epredia), processed, and embedded as formalin-fixed paraffin-embedded (FFPE) samples for potential miRNAscope analysis.
Animal work was performed at GSK, Collegeville, Pennsylvania, USA. 15 All studies were conducted according to GSK’s Policy on the Care, Welfare, and Treatment of Laboratory Animals and reviewed by the Institutional Animal Care and Use Committee at GSK or by the ethical review process at the institution where the work was performed. GSK is committed to the replacement, reduction, and refinement of animal studies (3Rs). Non-animal models and alternative technologies are part of our strategy and used where possible. When animals are required, application of robust study design principles and peer review minimizes animal use, reduces harm, and improves benefit in studies.
Statistical analysis for WEC data
Power calculations showed that using 16 rat or rabbit embryos will give a confidence rate of 86% for a true incidence rate of 20%. While there is no industry standard for group size in WEC studies, an N = 12 has been used in a streamlined rat WEC assay to classify teratogenic potential of pharmaceutical compounds with high predictivity. 16 For quantitative data, statistical significance was determined using the Kruskal–Wallis test (2-tailed) for somite number and Dunnett’s test for yolk sac size and embryo size. Statistical analysis was not performed for qualitative morphological observations.
miRNAscope
Detection of the unlabeled mipomersen oligonucleotide was carried out using an automated miRNAscope in situ hybridization assay, as previously described. 17 FFPE tissue sections cut at 5 µm, onto positively charged slides, were stained according to the manufacturer’s recommendations on an automated staining platform (Leica BOND RX, Leica) using a mild heated target retrieval (88°C for 15 min). Tissues were checked for global mRNA integrity using both universal positive control (RNU6) (Catalog # 727878-S1) and negative control (Scramble) (Catalog # 727888-S1) probes and then with a target-specific probe that detects the mipomersen nucleotide sequence (Catalog # 1763658-S1). Whole-slide images were scanned at 40× (Nanozoomer S360, Hamamatsu).
Results
Rat WEC
Rat embryos were treated from GD 9 to 11 with vehicle (n = 30) or 1, 10, 50, or 100 µM mipomersen (n = 16). All control embryos and embryos treated with ≤50 µM could be examined. Related to treatment at 100 µM, there were 11 (68.8%) grossly malformed and 1 (6.3%) dead embryos. Therefore, only four embryos at 100 µM mipomersen could be examined (Fig. 1A, Table 1).

Representative macroscopic photographs of rat
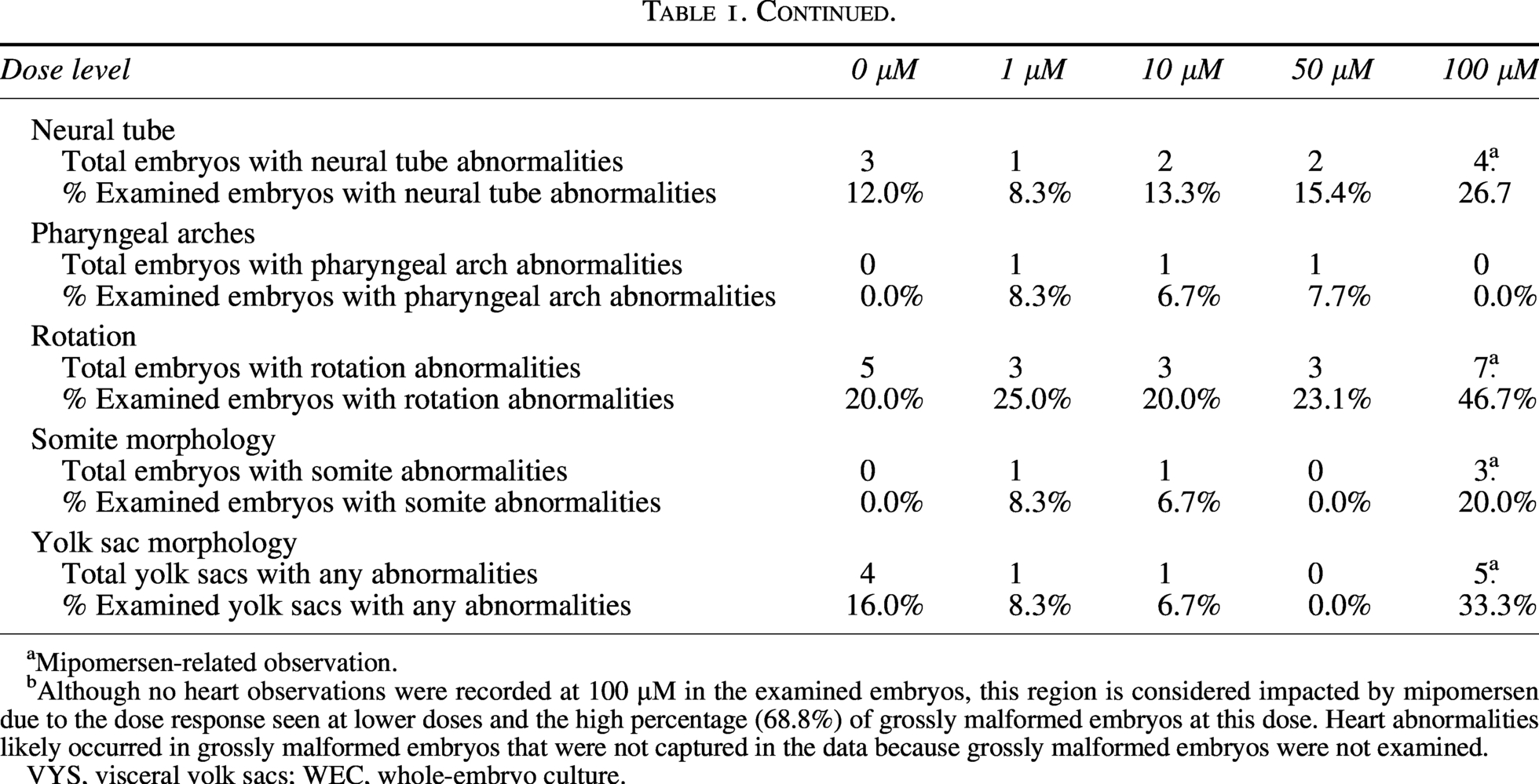
Only 1 of 4 embryos examined at 100 µM mipomersen had somites that could be visualized and counted. This embryo had a count of 23 somites, which is below the concurrent control mean of 27 and historical control mean of 26.5 (Supplementary Table S1). Statistical analysis was not performed on somite number due to low sample size. Mipomersen-related effects on somites also occurred at 50 µM; the mean somite number was 25 (0.92× of control). There was mipomersen-related statistically significant, dose-dependent decrease in mean VYS size at 50 and 100 µM (0.93× and 0.89× of control, P < 0.05). There was a mipomersen-related dose-dependent decrease in embryo size at 50 and 100 μM (0.93× and 0.81× of control, respectively), but statistical significance was only achieved at 100 μM (P < 0.0005) (Table 2). There were no statistically significant, mipomersen-related differences in mean VYS size, mean embryo size, or somite number ≤10 μM.
Only one of four embryos examined at 100 µM mipomersen had somites that could be visualized and counted; therefore, statistical analysis was not performed.
Mipomersen-related observation.
Statistically significant difference (P < 0.05).
WEC, whole-embryo culture.
Mipomersen-related dose-responsive morphological abnormalities occurred at ≥10 µM.
At 100 µM, morphological abnormalities occurred in all four embryos that were examined in the following regions: VYS (abnormal vasculature, blood islands, and/or paleness), caudal region (narrow or moderately short), craniofacial (cleft between the eye and first pharyngeal arch, loss of cells, absent mesencephalic flexure, short nasal prominence, optic vesicle, and/or otic placode abnormalities), neural tube defects (fourth ventricle collapsed, hemorrhage, spinal cord narrow, exencephaly, short, truncated, and/or narrow brain segments), forelimb and hind limb buds (remnant or not evident), and embryo rotation (squirrel or squirrel with tail fused to the body). Abnormalities were evident in pharyngeal arches (narrow/short) for two (50.0%) embryos examined. Although no heart observations were recorded at 100 µM in the examined embryos, this region is considered impacted by mipomersen due to the dose response seen at lower doses and the high percentage (68.8%) of grossly malformed embryos at this dose. Heart abnormalities likely occurred in grossly malformed embryos that were not captured in the data because grossly malformed embryos were not examined.
At 50 μM, abnormalities occurred in VYS (paleness) in 2 (12.5%) embryos, caudal (mild or moderately short, remnant, and dysgenesis) in 8 (50.0%) embryos, craniofacial (cleft, loss of cells, mesencephalic flexure misshapen, nasal prominence short or not evident, and optic vesicle abnormalities) in 7 (43.8%) embryos, neural tube defects (fourth ventricle collapsed or swollen, hemorrhage, exencephaly, short, truncated, and/or narrow brain segments) in 7 (43.8%) embryos, forelimb buds (remnant or small) in 10 (62.5%) embryos, hind limb buds (not evident) in 3 (18.8%) embryos, heart (outflow tract swollen or abnormal looping, chambers not well defined) in 4 (25.0%) embryos, embryo rotation (bend or squirrel) in 7 (43.8%) embryos, and pharyngeal arches (narrow/short, remnant, or not evident) in 3 (18.8%) embryos. At 10 μM, abnormalities were observed in the caudal region (moderately short) in one (6.3%) embryo, forelimb buds (small) in two (12.5%) embryos, and heart (outflow tract abnormal looping, atrium swollen) in one (6.3%) embryo (Table 1, Supplementary Table S2).
There were no mipomersen-related effects in rat embryos at 1 µM.
Rabbit WEC
Rabbit embryos were treated from GD 9 to 11 with vehicle (n = 30) or 1, 10, 50, or 100 µM mipomersen (n = 16). There were five (16.7%) control embryos and three (18.8%), one (6.3%), and two (12.5%) embryos at 1, 10, and 50 μM, respectively, found outside the VYS, which were not examined (considered to be an artifact of culture). There was one (6.3%) grossly malformed embryo in each group at 50 and 100 μM, which was considered unrelated to treatment due to being within historical control incidence (Supplementary Table S3). There was one dead embryo at 1 μM, which was not considered treatment related due to lack of dose response. These embryos were not examined; therefore, the total number of embryos examined was 25 (vehicle) and 12, 15, 13, and 15 at 1, 10, 50, and 100 μM, respectively (Fig. 1B, Table 1).
There was a test article-related statistically significant decrease in mean somite number at 100 μM (0.91× of control, 31 somites at 100 μM vs. 34 somites controls, P < 0.01), but no effects on VYS size or embryo size (Table 2).
Mipomersen-related morphological abnormalities occurred only at the high dose of 100 μM. Effects were observed in the VYS (collapsed, paleness, abnormal vasculature, blood island[s], blebbing/blister) in five (33.3%) embryos at 100 μM compared with four (16.0%) control embryos. Embryo rotation defects (bend, S-shape turning, or squirrel) occurred in 7 (46.7%) embryos at 100 μM compared with 5 (20.0%) control embryos. Hind limb bud abnormalities (small or remnant) occurred in six (40.0%) embryos at 100 μM compared with three (12.0%) control embryos. Craniofacial defects occurred in five (33.3%) embryos (short nasal prominence, optic vesicle observations) compared with three (12.0%) controls. Mipomersen-related abnormalities at 100 μM also occurred in somite morphology in three (20.0%) embryos (not well defined, not able to visualize, blister) and in the heart in three (20.0%) embryos (abnormal outflow tract looping, swollen pericardial sac) compared with the vehicle control group in which none of these abnormalities was observed (Table 1, Supplementary Table S4).
Due to lack of dose response and/or incidence within concurrent or historical control, there were no mipomersen-related effects in rabbit embryos at ≤50 μM (Supplementary Table S3).
miRNAscope
To confirm exposure, automated miRNAscope in situ hybridization was used to visualize the distribution of mipomersen within the embryos and on the VYS and amnion retained following measurements and examination of embryo morphology. Although morphological examinations were not performed on grossly malformed samples, miRNAscope was performed on sections through 12 rat embryos at 100 μM mipomersen dose, 16 at 50 μM, 16 at 10 μM, 15 at 1 μM, and 12 controls. In addition, 8 rabbit embryos at 100 μM mipomersen dose, 16 at 50 μM, 16 at 10 μM, 14 at 1 μM, and 15 control rabbit embryos were assessed. For all samples, an initial hematoxylin and eosin stain was generated and serial sections stained with the universal positive and negative control probes to quality control for overall morphology and global RNA quality (data not shown).
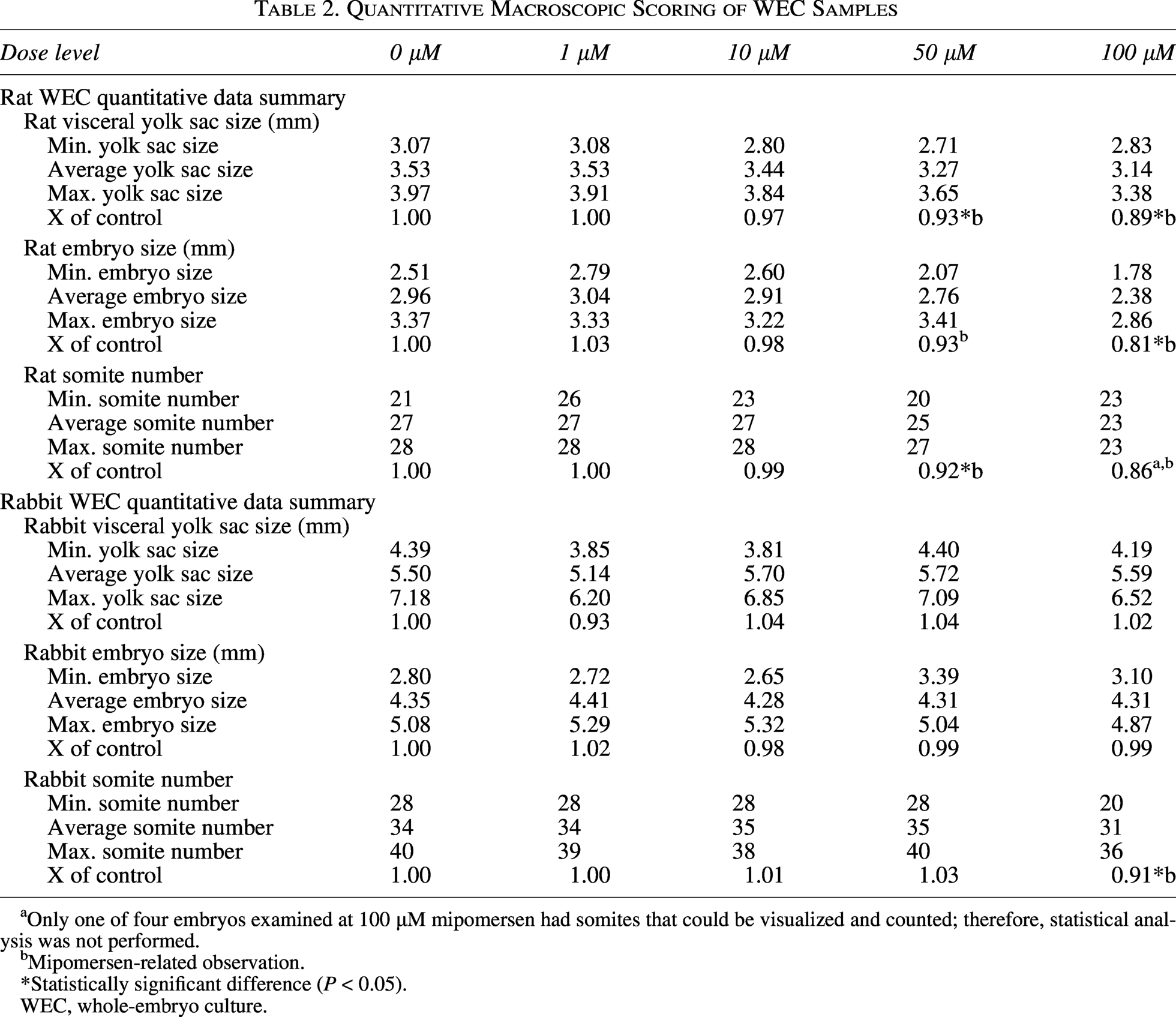
The embryo and the extraembryonic membranes of rat and rabbit control embryos were all completely negative for the ONT (Figs. 2A and 3A). At the 1 µM dose, all rat and rabbit embryos demonstrated strong staining on the VYS endoderm, the outermost layer of the yolk sac, but staining was noticeably absent from the embryonic mesoderm-derived endothelial, mesothelial, and hematopoietic cells, and the amnion. The embryo itself was negative for the oligo signal with the notable exception of the connection between the vitelline blood vessels and the embryonic gut in both rats and rabbits (Figs. 2B and 3B).

Representative midline sagittal sections through the rat embryos and extraembryonic membranes were taken through all samples and stained by miRNAscope with a probe complementary to mipomersen. Whole-slide scans were taken and the staining patterns documented. No (red chromogen) signal was detected in either the embryo or the yolk sac (ys) in the controls
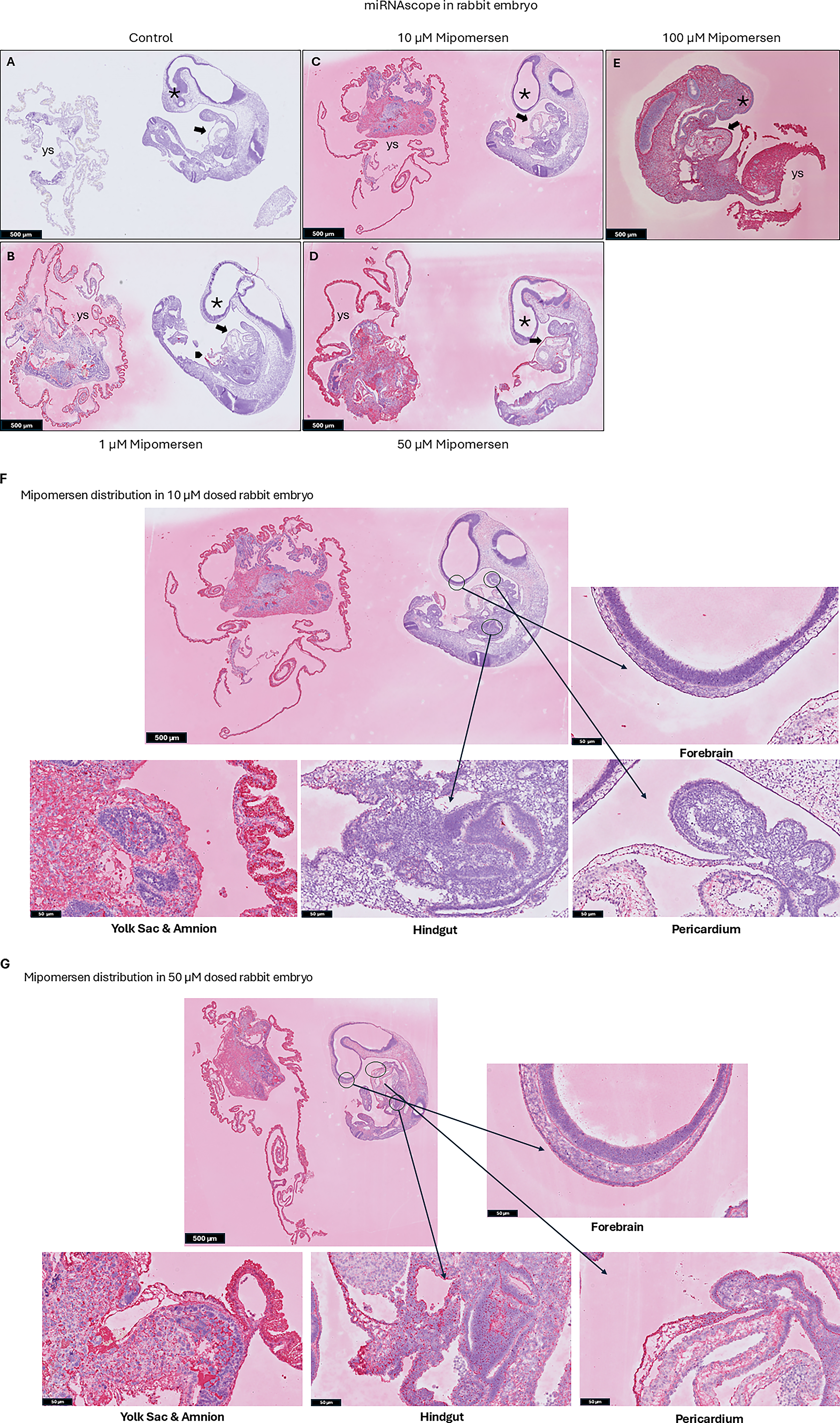
Representative midline sagittal sections through the rabbit embryos and extraembryonic membranes were taken through all samples and stained by miRNAscope with a probe complementary to mipomersen. Whole-slide scans were taken and the staining patterns documented. No (red chromogen) signal was detected in either the embryo or the yolk sac (ys) in the controls
At 10 µM, the rat yolk sac staining was largely the same as at 1 µM, although a weak signal was seen in the endothelial cells and in the amnion itself. However, the rabbit yolk sac staining at 10 µM showed higher mipomersen expression compared with the 1 µM treated embryos (Figs. 2C and 3C). ONT was detected on the pericardium wall of the developing heart, the surface ectoderm of the craniofacial region, and at low levels in the forebrain neuroepithelium in both rat and rabbit embryos (Figs. 2F and 3F). Most notable at this stage was the appearance of the ONT in the hindgut endoderm (Figs. 2F and 3F).
At both the 50 µM and the 100 µM doses for rat and rabbits, all embryos and extraembryonic membranes were heavily stained in all the epithelial and mesenchymal cell layers throughout the samples (Figs. 2D, E, G and 3D, E, G).
Discussion
After culture in media containing 0, 1, 10, 50, or 100 µM mipomersen, both rat and rabbit embryos showed dose-responsive changes in yolk sac and/or embryo parameters suggestive of exposure to the ONT. At the high dose of 100 µM, rats were more severely impacted compared with rabbits, with most rat embryos being grossly malformed and those that were examinable were characterized by decreased yolk sac and embryo size, decreased somite number, and changes to yolk sac and embryo morphology. Most rabbit embryos at 100 µM were examinable and were less severely impacted, only showing a decrease in somite number and changes to embryo morphology. In rats, we observed a clear dose–response relationship. No effects were noted at the lowest dose of 1 µM even with demonstrated exposure in the vitelline blood vessels and embryonic gut, with progressively greater effects seen at each higher dose. In rabbits, a dose-response was evident only at the high dose, where test item effects were observed despite demonstrated exposure in the embryo at all dose levels.
The differences observed between species may be partially due to differences in off-target profiles. Since mipomersen is not pharmacologically active in rat or rabbit, 9 all observed effects are presumably due to off-target toxicity, which may be hybridization dependent or independent, and/or chemical/physical property-related toxicity. Mipomersen is a full phosphorothioate backbone single-stranded ASO, which is associated with non-specific binding and proinflammatory responses. Since the innate immune responses by primitive macrophages may also begin during the culture period of GDs 9–11, inflammatory cytokines and chemokines could have been stimulated having an impact on signaling relevant for embryonic development.18–20
Rats and rabbits also have structural and functional differences in their extraembryonic membranes. 21 Rat embryos are completely enclosed by the yolk sac during the culture period and have a Reichert’s membrane, which is rodent-specific, whereas rabbit embryos are not completely enveloped by the yolk sac until after the culture period on approximately GDs 12–13.13,14,21,22 Rats and rabbits have an inverted yolk sac meaning that the endodermal surface, which is specialized for nutrient absorption, directly interfaces with the maternal environment in vivo or with the cell culture media in WEC.23,24 The yolk sac endoderm is highly endocytotically active and supplies nutrients via histiotrophic nutrition. Disruption of these processes at higher doses in rat WEC could have contributed to the higher incidence of abnormalities, compared with rabbits, which were not completely enveloped in yolk sac during the culture period, as interference with histiotrophic trafficking has been shown to cause malformations and resorptions in vivo. 23 This potential disruption in histiotrophic nutrition may also be contributing to the discordant outcomes between the mipomersen in vivo studies, which did not detect any effect, and these WEC studies. ASOs have been shown to bind to transporters and act as competitive inhibitors in specific contexts, for example, binding to renal transporters, such as megalin and cubilin, to interfere with protein uptake in proximal tubule epithelial cells. 25 Therefore, mipomersen may be competing with nutrients for transfer into the embryo, especially in rats as megalin and cubilin are highly expressed in the rat yolk sac and have been shown to mediate endocytosis of multiple nutrients in VYS.26–29 Evidence for a similar role in the rabbit yolk sac is not available, but megalin and cubilin are expressed in human yolk sac with functions in nutrient transfer. 30 Histiotrophic nutrition has human relevance as maternal circulation to the placenta is not fully established until 10–12 weeks of gestation and histiotrophic nutrition from the uterine glands may be occurring throughout the first trimester. 31 Therefore, disruptions to histiotrophic nutrition could potentially lead to adverse pregnancy events in humans.
There was a mipomersen-related dose-dependent effect on rat embryo morphology in every region evaluated. Similar generalized grossly malformed phenotypes, as well as embryonic lethality across entire dose levels, have been previously observed when testing small molecules in WEC. Positive controls have not yet been established for ONTs in DART-specific NAMs. Future work should evaluate ONTs with pharmacological activity against targets with a known role in embryonic development between GDs 9 and 11.
Automated miRNAscope in situ hybridization was used to confirm direct mipomersen exposure inside the embryos at all dose levels, including dose levels at which no embryo or VYS changes occurred. This suggests that mipomersen, and possibly other ASOs with a similar chemical composition, can get into the embryo without inherently causing toxicity. At lower doses, the allantois and heart were the first structures to show positive signal, suggesting that ONTs may enter the umbilical vasculature and pass into embryonic circulation in a dose-dependent manner until saturating at higher doses where a nearly ubiquitous mipomersen distribution was seen throughout all major tissues and structures in all dosed rat and rabbit embryos.
Endocytosis may allow some ONTs to be taken up by the endodermal surface of the yolk sac and pass into the umbilical vasculature, as ONTs are often modified to enhance endocytosis and improve resistance to nuclease degradation, leading to longer half-lives and increased stability within the target cell. 8
ONTs have properties of both small molecules and biologics. In the absence of internationally harmonized regulatory guidance specific to ONTs, guidelines for both small molecules (i.e., ICH M3) and biologics (i.e., ICH S6) as well as DART-specific principles [i.e., ICH S5(R3)] are typically considered during the development of ONTs as potential medicines.32–34 In addition, the Guideline for Preclinical Safety Assessment of Oligonucleotide Therapeutics issued by the Pharmaceuticals and Medical Devices Agency in Japan and the draft Nonclinical Safety Assessment of Oligonucleotide-Based Therapeutics Guidance for Industry issued by the FDA should be considered when appropriate.35,36
When a candidate ONT is not pharmacologically active in preclinical species, the current consensus is that in vivo DART study designs should include several dose levels of the clinical candidate in two species (for embryo–fetal development studies) in addition to one group in at least one species that utilizes an appropriate species-specific surrogate molecule for hazard identification. 1 This approach was implemented for the mipomersen DART evaluation in the mouse combined fertility and early embryonic development study using a mouse surrogate and the rabbit embryo–fetal development study using a rabbit surrogate. 9 There was no evidence of impaired fertility or embryo–fetal development in mice or rabbits at doses up to 2× and 5× clinical exposure, respectively. This lack of effects compared with our WEC results could be because mipomersen was unable to access embryonic tissue in vivo as it was not detectable in the fetal liver or kidney but was detectable in the mouse and rabbit placenta; alternatively, the method of detection may not have been sensitive enough and mipomersen may have been present at a low concentrations in the embryo–fetal compartment without any effects in vivo. When using surrogate molecules, only hazard identification is possible and not risk assessment. As the current data demonstrate the ability of ONT to access embryonic tissues via WEC, it suggests that WEC assays could be used as a hazard identification assay for early embryonic development. Since both WEC evaluation and the surrogate group of an in vivo study should only be used for hazard identification, a WEC evaluation of a surrogate molecule could potentially replace the hazard identification surrogate arm of an in vivo study. This would reduce animal use and limit the amount of surrogate molecule quantities needed as WEC would require less test material compared with an in vivo study. WEC studies may provide more robust hazard identification, compared with embryo–fetal development studies including a surrogate group, as embryonic exposure in WEC may be confirmed while maternal components may limit ONT transfer to the embryo–fetal compartment in vivo. Maternal toxicity may also limit dosing to lower levels, thus limiting embryo–fetal exposure. The data suggest that current embryo–fetal development studies in two species for ONT development could be replaced with a hazard identification WEC study with the appropriate surrogate molecule and an in vivo second species using the clinical candidate to identify non-specific toxicity, thereby reducing animal use.
Mipomersen exposure in placental tissue in the DART studies was 23 and 15 µg/g in mice and rabbits at high doses of 25 and 15 mg/kg/dose, respectively. The dose of 100 µM mipomersen in WEC media represents approximately 33× or 50× the placental exposure in mice and rabbits, respectively, before accounting for protein binding. Mipomersen is expected to be >85% protein-bound. 37 Since the culture media are abundant in proteins, a relatively high concentration of 100 µM mipomersen was selected to ensure that adequate levels of unbound mipomersen would be available to enter embryos. Lower doses of 1, 10, and 50 µM mipomersen were selected to evaluate a dose response and the low doses are approximately at parity with the placental exposures observed during the in vivo DART studies.
Because ONT properties vary, future WEC experiments should assess distribution within embryos for different antisense chemistries, as well as different tool ONT classes (e.g., siRNAs, microRNA mimics, transfer RNA, decoys, and aptamers). ONTs with varied chemical structures will have different protein binding properties, which can impact the profile of non-specific effects, as well as the ability of certain ONTs to cross the yolk sac and be taken up into the embryo. Future studies should also utilize ONTs that are pharmacologically active in the test species, enabling assessment of productive uptake, which was not possible in the current study due to lack of target engagement. Knockdown levels of the target protein should be measured to confirm the functional effect of ONT treatment within the 40- to 45-h culture period. It will be important to establish the time course of the knockdown effect to determine if changes in embryo measurements and morphology might be on-target effects, which is plausible given that the WEC period occurs during organogenesis, a developmental window characterized by rapid cell proliferation, growth, and protein synthesis. Consideration should be given to selecting ONTs that target mRNAs encoding proteins with known roles in embryo–fetal development and protein expression during the culture period. These studies will further validate the utility of WEC models for hazard identification in this context.
In addition, in vivo studies administering ONTs should assess placental and embryo–fetal distribution using an in situ hybridization-type approach or immunohistochemistry to provide single-cell resolution and spatial context. 38 Most previous studies have used tissue lysate-based quantification methods such as LC-MS/MS, hybridization ELISA, or q-PCR, which measure concentrations in a diluted homogenate. These methods may or may not offer the level of sensitivity required to reliably detect ONTs in fetal samples. ONTs can enter the umbilical vasculature and embryos in WEC and placental exposure has been confirmed in vivo. Therefore, it is plausible that ONTs are entering the embryo–fetal compartment in vivo, unless the placenta is acting as a barrier to prevent exposure. A thorough investigation of ONT distribution in placenta is warranted to determine if evidence of a barrier can be visualized. At early developmental stages before the placenta develops, the human yolk sac provides initial nutrient transfer and gives rise to the first blood cells of the embryo. 39 ASGR1 is expressed in the human yolk sac endoderm at ∼8 weeks postconception. 40 ASGR1 is a receptor that is highly expressed in hepatocytes and specifically binds the GalNAc molecule to achieve targeted exposure of GalNAc siRNAs in the liver; however, ASGR1 expression in the human yolk sac could lead to early embryonic exposure. This further highlights the need for additional investigations using a variety of ONT classes.
ASGR1 expression in rats is generally absent in the fetal liver and its expression is rapidly induced around birth. 41 However, ASGR1 expression has not been evaluated in rat or rabbit embryos specifically during the developmental period relevant for WEC from GDs 9 to 11 and extraembryonic membranes have not been evaluated. ASGR1 expression patterns should be determined for key developmental stages within the embryo and in extraembryonic membranes, as this information would have implications for the interpretation of in vivo DART study’s dosing GalNAc-conjugated ONTs.
Conclusions
We have demonstrated that WEC experiments involving mipomersen, a 2′-O-methoxyethyl phosphorothioated ASO, do not require microinjection or assisted transfection, enabling developmental hazard identification of off-target toxicity by directly exposing rat and rabbit embryos to this ONT in culture media. Hazard identification is especially important if embryo–fetal exposure may be limited in vivo, as it is not understood if transfer of ONTs between the maternal and embryo–fetal compartments in non-clinical species is predictive of that in humans. While WEC studies only identify hazard potential and are not risk assessment studies due to the absence of maternal factors, identification of potential hazards may help inform prescribers and patients. The demonstration that mipomersen can access embryonic tissues in WEC supports the relevance of this system for identifying intrinsic embryotoxic potential, including off-target, chemistry-related, or exposure-driven effects that may not be manifest in vivo due to limited embryo–fetal exposure or dose-limiting maternal toxicity. However, many questions remain regarding whether and how these data can be incorporated into a risk assessment weight of evidence. These questions are not unique to the WEC model but are applicable to all NAMs. As NAMs are more widely adopted, a consensus needs to be reached on the advantages and appropriate use case for each model. The utility of WEC as an NAM for assessing developmental hazards of ONTs, potentially complementing or partially replacing in vivo studies thereby reducing animal use and the required test material amounts, should be considered as the new International Council for Harmonization of Technical Requirements for Pharmaceuticals for Human Use (ICH) S13 guidelines are drafted for the non-clinical safety evaluation of ONTs. 42
Footnotes
Acknowledgments
The authors thank Heather Laurie-Williams, Joseph Hosford, and Doug Fuerst (GSK Drug Substance Supply) for the synthesis and supply of mipomersen.
Author Disclosure Statement
All authors are employed by a company, GSK, that develops oligonucleotide therapies. In addition, J.P. is the European Federation of Pharmaceutical Industries and Associations (EFPIA) Lead on ICH S13 Expert Working Group.
Funding Information
This work was funded by GSK.
Supplemental Material
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.