
Abstract
Significance:
Recognized for its role in energy storage and thermal insulation, white adipose tissue (WAT) has garnered increasing attention given its functions beyond metabolism. In particular, dermal and subcutaneous WATs have been shown to play critical roles in wound healing and scarring. This review proposes that dermal and subcutaneous adipose tissues are not passive bystanders but active regulators of wound healing and fibrosis.
Recent Advances:
Significant advances in wound healing biology have identified critical roles and signaling pathways that immune cells and fibroblasts are involved with in wound healing; however, fewer investigations have studied adipocytes in this context. This review focuses on the roles of WAT and specifically white adipocytes at different stages of wound healing: inflammation, proliferation, and remodeling. We also discuss applications of WAT and its derivatives as therapeutic strategies for improved wound healing.
Critical Issues and Future Directions:
With multiple fibrotic conditions associated with decreased adipose tissue, understanding the different ways by which adipocyte subpopulations and WAT participate in repair and fibrotic processes will help shed light on how we can modulate cellular response for future therapeutic applications.
Derrick C. Wan, MD

SCOPE AND SIGNIFICANCE
Extensive work has previously focused on the roles of immune cells and scar-depositing fibroblasts during wound healing; yet, fewer investigations have been conducted evaluating the roles of adipocytes in this process. Emerging evidence suggests that dermal white adipose tissue (DWAT) and subcutaneous white adipose tissue (SWAT) are not passive structural elements but active regulators of wound healing and fibrosis. This review will focus on our current understanding of the dynamic roles DWAT and SWAT play in different phases of this process. We will provide a summary of the current translational research investigating adipose tissue and its derivatives as potential therapeutic strategies to overcome fibrotic processes and chronic wound conditions.
TRANSLATIONAL RELEVANCE
Current research focuses on understanding the underlying mechanisms by which dermal and subcutaneous adipose tissues participate in the different stages of wound healing, ranging from immunomodulatory to scar-depositing roles. Given that adipose tissue appears to play roles in both fibrotic outcomes and regenerative healing, by understanding the fundamental biology underlying dermal pathological conditions involving adipose tissue, many groups hope to develop adipose tissue and adipose-derived therapies to facilitate wound healing or improve fibrotic outcomes in the future.
CLINICAL RELEVANCE
Wound healing and scarring is a condition affecting the majority of the human population, with serious dermal injuries often leading to pain, decreased function, and undesirable esthetic outcomes. While adipocytes have been shown to participate in scar formation, many fibrotic conditions such as radiation-induced fibrosis and systemic scleroderma are often associated with decreased fat distribution, suggesting some protective effects of the tissue. However, it is also possible that fat loss in these contexts reflects secondary effects of the disease process (e.g., local ischemia, inflammation, or catabolic signaling) rather than a primary driver of fibrosis.1–4 Thus, investigations into molecular therapies derived from increased understanding of the cell types and mechanisms in adipose tissue functions show promise as potential future directions in the management and treatment of wound healing and scarring.
BACKGROUND/OVERVIEW
White adipose tissue (WAT) is found in distinct locations throughout vertebrate species. Comprised of white adipocytes, these depots serve as key players in energy storage, metabolism, thermal insulation, and mechanical cushioning. 5 WAT distribution and function are distinct from brown adipose tissue, which plays a critical role in thermoregulation and heat production 6 (Fig. 1). Two large WAT depots are typically described: subcutaneous WAT (SWAT), found below the skin, as well as visceral WAT, surrounding internal organ systems. However, WAT depots exist in other anatomical locations. Notably, although adipose tissue associated with the skin has historically been considered a singular depot known as SWAT, over the past decade, the field has recognized that a distinct layer of adipose tissue lies within the reticular dermis, known as dermal WAT (DWAT). 7 In mice and rats, SWAT is physically separated from DWAT by the panniculus carnosus. Albeit that humans lack this layer of skeletal muscle, these two depots of WAT remain distinguishable.8,9 This anatomical difference is important to consider when interpreting preclinical studies of wound healing, as differences in tissue organization and components (e.g., panniculus carnosus in small animals) may influence adipose tissue behavior and wound healing dynamics in preclinical models. 10

Distribution of adipose tissue in humans and mice. White adipose tissue is primarily distributed within subcutaneous tissue and surrounding viscera. Brown adipose tissue in humans is situated primarily paravertebrally, within axilla, and supraclavicularly, while principally found intrascapularly in mice. Biorender.com Adapted from “Adipose Tissue Depots” and “Mouse Adipose Tissue Distribution and Types.” Retrieved from https://app.biorender.com/biorender-templates. Created in BioRender. Parker, J. (2025) https://BioRender.com/ehh3oz7.
Dermal adipose tissue has garnered increasing attention given its roles outside of metabolism, such as in inflammatory regulation, thermogenesis, and hair follicle cycling.11–13 For instance, adipocytes within DWAT expand during hair follicle growth and retract during hair follicle regression. Importantly, there is growing interest in the role that adipose tissue plays in wound healing and scarring. Although different groups have demonstrated that DWAT plays critical roles in this process,14–17 it has been demonstrated that SWAT participates in wound healing as well, suggesting that both WAT layers within the skin assist in this process. 18
Notably, fibrotic conditions are often associated with decreased fat distribution. For example, although radiotherapy is one of the primary components of cancer treatment, it is associated with collateral tissue injury often leading to radiation-induced fibrosis (RIF). 19 RIF is characterized by increased dermal thickness and loss of vascularity and tissue elasticity (Fig. 2A). RIF is a prominent sequela of radiation therapy for multiple cancers, with the incidence of grade-2-or-higher neck fibrosis following surgical dissection sometimes higher than 50%. 22 Another example of fibrotic disease is systemic sclerosis (SSc), a complex autoimmune disease associated with fibrosis in multiple organs for which the etiology remains incompletely understood. 23 Similar to RIF, it is also characterized by decreased DWAT, with excisional biopsies from patients demonstrating replacement of subcutaneous tissue by fibrotic connective tissue 24 (Fig. 2B). For both conditions, few effective treatment modalities are available, and adipose tissue and derived cells therefore offer promise as potential therapeutic strategies for these conditions.
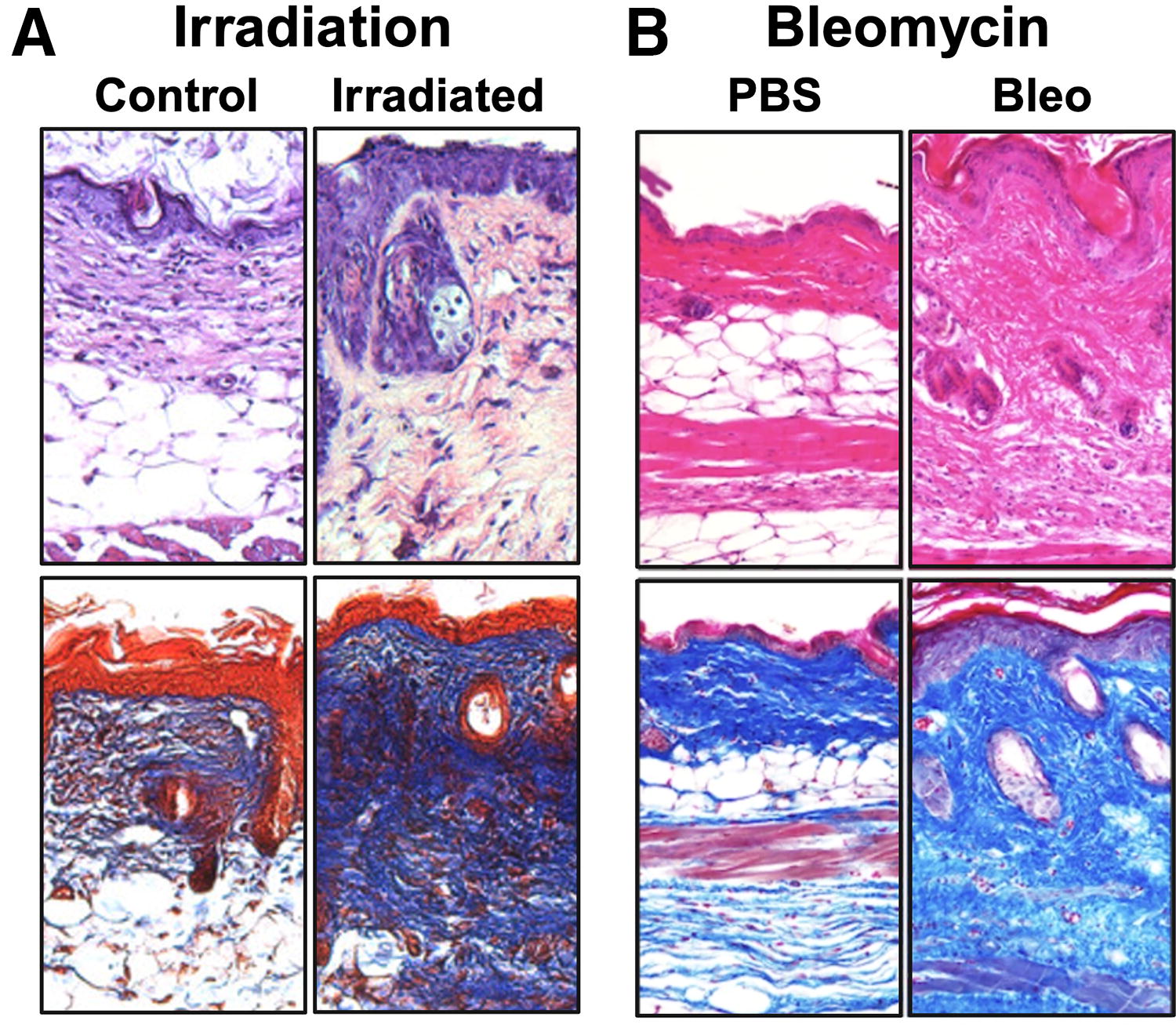
Loss of white adipose tissue in fibrotic conditions.
In this review, we begin our discussion with wound healing and the roles that WAT plays during the different stages of repair. We then touch on molecular signaling pathways and mechanisms influencing adipocyte biology within WAT. Finally, we discuss the applications of WAT as a therapeutic strategy for improved wound healing in a clinical context.
Translational insight
Fat loss is a hallmark of RIF and SSc. Exploring mechanisms of DWAT depletion in these settings may yield adipose-targeted therapies to reduce fibrotic burden.
DISCUSSION
Wound healing overview
As the primary barrier to the external environment, the skin is a common site for tissue damage. Following injury, the skin undergoes an orchestrated and complex process to repair the damaged site, involving multiple cell types and signaling pathways. Wound healing is typically described as a series of four overlapping stages: (1) hemostasis; (2) inflammation; (3) proliferation; and (4) remodeling. 25 Shortly after injury, bleeding from damaged blood vessels is controlled via the coagulation cascade. Inflammatory mediators secreted within the wound bed attract immune cells such as neutrophils and monocytes, which assist in sterilizing the wound and debris removal within the first few days of the wound healing process.26,27 Five to seven days following injury, inflammatory and platelet cell activity leads to migration of fibroblasts to the wound, the main cell type responsible for extracellular matrix (ECM) synthesis and deposition.28,29 A subset of fibroblasts activated via both mechanical forces and certain cytokines including transforming growth factor β (TGF-β) differentiate into myofibroblasts, which participate in wound contracture.30,31 During remodeling, the final stage of wound healing, fibroblasts slowly replace the provisional ECM with one that is stiffer and stronger than the original granulation tissue, ultimately establishing a mature scar.25,32 WAT participates in different roles throughout these stages, as described in further detail in the following sections. A summary of the key events involved in wound healing and the role of WAT is presented in Table 1.
Summary of wound healing stages and the role of WAT in each
DWAT, dermal white adipose tissue; ECM, extracellular matrix; WAT, white adipose tissue.
WAT participation during the inflammatory phase
Adipose tissue, in addition to its primary role in energy homeostasis as an endocrine organ, 33 also contains multiple types of immune cells and participates in the immune response. 34 In particular, adipocytes within WAT secrete pro- and anti-inflammatory factors known as adipokines, which directly influence the composition of immune cells within adipose tissue.35–37 In the skin, for example, subcutaneous adipocytes express adipochemokines when exposed to ultraviolet radiation, including C-C motif chemokine ligand (CCL) 3, CCL5, C-X-C motif chemokine ligand (CXCL) 1, and CXCL5. 38 SWAT and DWAT are known to also secrete adipokines that play key roles in energy homeostasis and metabolism, such as leptin and adiponectin. Interestingly, these adipokines participate directly in the inflammatory response, 39 with leptin evoking a proinflammatory response in macrophages and neutrophils 40 and adiponectin enhancing polarization of macrophages toward an anti-inflammatory state17,41,42 (Fig. 3, left). Mechanistically, leptin promotes inflammatory signaling in macrophages through the leptin receptor and Janus kinase/signal transducer and activation of transcription pathway, promoting proinflammatory cytokine production (e.g., interleukin [IL]-6, tumor necrosis factor ɑ [TNF-ɑ]). 40 Leptin receptor-deficient or leptin-deficient mice have shown impaired cytokine production and phagocytosis, highlighting leptin’s essential role in macrophage activation and function.43,44 In contrast, adiponectin exerts anti-inflammatory effects via AMP-activated protein kinase and IL-10-mediated pathways.17,41,42
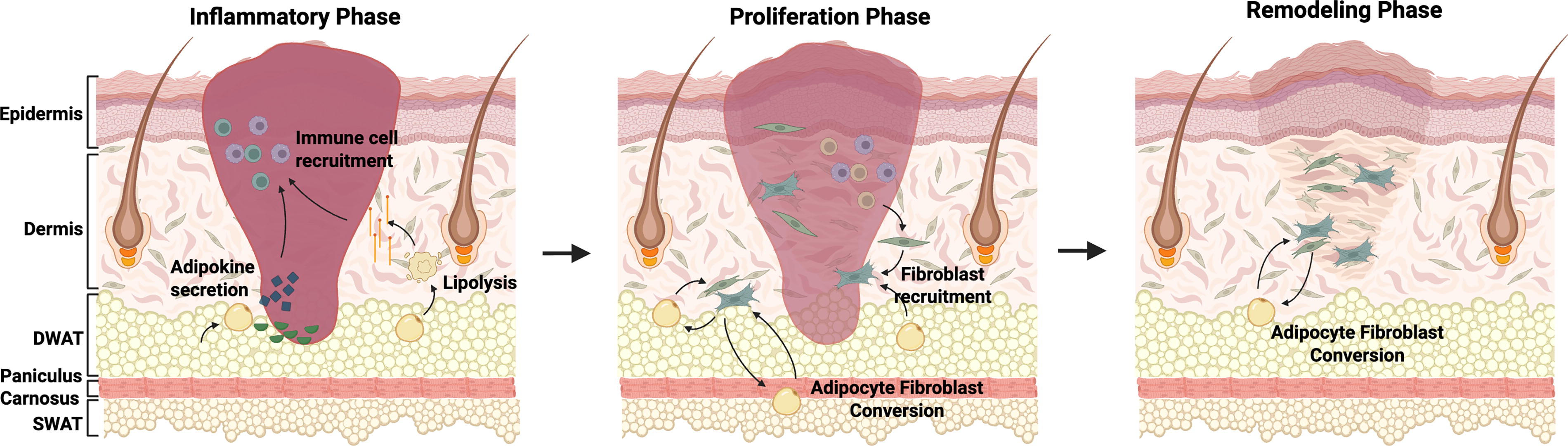
Adipose tissue involvement at different stages of wound healing. Left, inflammatory phase: adipocytes secrete adipokines that recruit immune cells such as monocytes and neutrophils to the injury site. Some adipocytes within dermal white adipose tissue (DWAT) also undergo lipolysis, leading to additional macrophage recruitment. Middle, proliferation phase: adipocytes near the wound site recruit fibroblasts. Some adipocytes also undergo adipocyte to fibroblast transition, participating directly in matrix deposition. Right, remodeling phase: temporary granulation tissue is replaced with mature scar. Fibrosed skin is characterized by continued absence of DWAT and secondary skin structures (e.g., hair follicles). Created in BioRender. Parker, J. (2025) https://BioRender.com/ehh3oz7.
Within WAT, adipocytes also express receptors that can detect and respond to inflammatory signaling, such as toll-like receptors (TLRs). TLRs respond to foreign materials such as cell surface components of microbes and are one of the initial contributors to activation of the immune system. 45 Of note, TLRs are one of three primary activators of proinflammatory nuclear factor kappa B (NF-κB) signaling. 46 In conjunction with bacterial stimuli, NF-κB signaling can lead to expression of multiple proinflammatory cytokines, including CCL4, CXCL10, intercellular adhesion molecule 1, IL-6, and TNF-ɑ. 47 Adipocytes are known to also express nucleotide-binding oligomerization (NOD)-like receptors and RIG-I-like receptors (RLRs). 48 Certain NOD-like receptors contribute to adipocyte differentiation, NF-κB signaling, and the inflammasome,49–51 whereas types of RLRs sense double-stranded RNA and viruses. 52 How these two types of receptors function in the context of adipocytes’ inflammatory response, specifically, remains unclear.
In the context of wound healing, adipocytes within the dermis have been shown to be necessary for macrophage-based inflammation following injury. 16 Shook and colleagues established that following wounding and prior to immune cell infiltration, adipocytes at the wound edge undergo lipolysis and release fatty acids. Prevention of lipolysis within wounds led to decreased inflammatory macrophage infiltration and delayed repair. That said, the specific mechanism by which adipocytes and metabolic products activated macrophages within the wound microenvironment remains unclear. Macrophages express fatty acid receptors and transporters. Thus, as inflammatory macrophages conduct glycolysis for energy, one theory is that these cells benefit from the increased release of lipids by adipocytes. 53 In support of this hypothesis, inhibition of adipocyte lipolysis decreased medium-chain fatty acid release, limited macrophage migration, and decreased G protein-coupled receptor 84 (GPR84) signaling. 47 In this signaling pathway, GPR84 interacts with Gi/o proteins, leading to phosphorylation of Akt, extracellular signal-regulated kinases, and NF-κB-p65 signaling. Increased activity of this cascade led to elevated expression of multiple inflammatory cytokines, including IL-6 and TNF-ɑ. Chemokines, including C-C motif ligand 2 and CXCL1, and proteins known to recruit inflammatory cells, such as neutrophils and macrophages, to the site of inflammation also increased.54,55
Further, adipocyte progenitors demonstrate a role in immune cell regulation within the dermis. DWAT contains a rich source of adipocyte-derived stem cells (ASCs). 56 Previous work from Zhang and colleagues demonstrated that dermal fat expanded following Staphylococcus aureus infection, and that impairment of adipogenesis increased infection. 57 Following injury, preadipocytes within the area underwent rapid proliferation and the dermal adipose layer expanded, secreting antimicrobial peptides such as cathelicidins known to promote re-epithelialization. These data suggested that adipocytes not only direct and modulate how immune cells respond to signaling but participate directly in host defense. As inflammation resolves, adipose tissue shifts from immune modulation to promoting tissue regeneration, marking the transition to the proliferative phase.
Translational insight
Enhancing adipocyte signaling and antimicrobial responses during early inflammation could help control infection and prevent chronic wound states.
Function of WAT during proliferation phase
Adipocyte–fibroblast interactions
Following the inflammatory phase, signaling from immune cells leads to migration and recruitment of fibroblasts to the wound site, which in turn contribute a major role in preliminary ECM deposition. 25 During the proliferation phase, DWAT also plays multiple functions. Notably, adipocytes within DWAT appear to participate in fibroblast recruitment. Adipocyte-derived factors such as platelet-derived growth factor (PDGF), connective tissue growth factor, and fibroblast growth factor 2 activate mitogen-activated protein kinase and phosphatidylinositol 3-kinase (PI3K)/protein kinase B (AKT) signaling in fibroblasts, promoting fibroblast migration and ECM production.58–61 Inhibition of adipogenesis leads to a reduction in fibroblast migration. Schmidt and Horsley demonstrated that “fatless” AZIP/F1 mice, which lack mature adipocytes, have decreased cellular organization following wound healing and decreased fibroblasts within the wound bed. 15 Interestingly, fibroblasts remained present around wound edges, suggesting that white adipocytes were necessary for fibroblast repopulation within wounds. In keeping with these results, El-Hattab et al. established that conditioned medium from adipocytes in a spheroid culture system induced human dermal fibroblast migration and differentiation into a myofibroblast-like phenotype. 62
Of note, not only do adipocytes recruit fibroblasts, but rather, some adipocytes also undergo adipocyte-to-fibroblast transition to participate directly in scar deposition as well. For instance, applying lineage tracing models, Shook and colleagues demonstrated that following wounding, adipocytes migrated into the wound bed, dedifferentiated, transitioned into myofibroblasts, and contributed to scar deposition. 16 Another study applying a bleomycin-induced skin fibrosis model observed substantial loss of DWAT following bleomycin injection, with fibrous tissue found in its stead. 63 Loss of DWAT also correlated with a loss of adipogenic and gain of fibrotic markers (Fig. 3, middle).
Adipocyte interactions with other cell types
Although fewer studies have been conducted, adipocytes within WAT are believed to also participate in regulation of angiogenesis and re-epithelialization. For example, ablation of dermal adipocytes delays both re-vascularization and re-epithelialization, as described by Shook and colleagues, wherein adipocyte ablation experiments suggested that adipocytes were required for efficient activation of the inflammatory response and downstream steps in wound repair. 16 Additional work performed with ASCs supported the role of adipocytes in re-epithelialization and re-vascularization. These studies indicated that application of ASCs to full-thickness wounds in vivo led to improved re-epithelialization via increased keratinocyte proliferation, along with increased blood vessel density.64,65
Neurovascular interactions
Adipose tissue is richly innervated and vascularized, and data suggest that neural input and vascular remodeling jointly influence adipocyte function during wound healing. Sympathetic nerve fibers contribute to wound healing by releasing neurotransmitters such as norepinephrine, neuropeptide Y (NPY), acetylcholine, and vasoactive intestinal peptide (VIP). 66 NPY promotes angiogenesis and wound contraction and regulates collagen synthesis, whereas VIP regulates cell proliferation and differentiation and exerts anti-inflammatory effects. 67 Vascular endothelial growth factor (VEGF) and PDGF are crucial for angiogenesis and are significantly influenced by neurovascular interactions.68,69 Denervation or altered neurovascular interactions, such as in the context of diabetic neuropathy, therefore impair the healing process. 70 As proliferation settles, the wound transitions into the remodeling phase, where matrix reorganization determines the quality of healing.
Clinical takeaway
Leveraging DWAT to support fibroblast recruitment and angiogenesis during the proliferation phase may enhance granulation tissue formation and improve healing outcomes.
Remodeling phase
During the final phase of wound healing, the remodeling phase, temporary granulation tissue is replaced with mature scar. Type III collagen is replaced with Type I collagen, and the scar gains elasticity and strength. Meanwhile, vasculature within the scar bed decreases, and cells initially involved in repair including endothelial cells, macrophages, and myofibroblasts either undergo apoptosis or leave the initial wound site. 32
Long-term scars are characterized by the continued absence of secondary skin structures, including hair follicles or glands. Fibrotic skin is also characterized by an absence of DWAT, with lineage tracing suggesting that adipocytes originally derived from DWAT become one of the contributors to long-term scar deposition once converted to adipocyte-derived fibroblasts. 63 Adipocyte-to-myofibroblast transition during remodeling is driven by profibrotic signaling pathways, particularly TGF-β and wingless/integrated (WNT)/β-catenin, as detailed in the section “Signaling pathways”.71,72 Further investigations into adipocyte-fibroblast dynamics within long-term scars are warranted in order to better understand how scars can be reverted toward a more regenerative state. As this is believed to be a stage of wound healing that lasts upwards of years following injury, much remains unknown at a molecular level about the signaling pathways and cell interactions involved in scar homeostasis (Fig. 3, right).
Although wound healing typically leads to scar formation and an absence of hair follicles, a unique model applying a large wound in an adult mouse can lead to hair follicle neogenesis.73,74 Using this model, Plikus and colleagues demonstrated that adipocytes regenerated within the scar and were derived from myofibroblasts. They demonstrated that reprogramming of myofibroblasts required the presence of neogenic hair follicles and expression of bone morphogenetic protein (BMP). 75 Culturing fibroblasts from human keloids, they also found that adipocytes formed in vitro following treatment with BMP or placement of these fibroblasts adjacent to human hair follicles. These data ultimately demonstrated the plasticity of the adipocyte-fibroblast lineage. Notably, as hair follicle neogenesis in this model occurs following re-epithelialization, and at the start of early wound remodeling, these data underscored the potential that this plasticity could be applied for the modulation of existing, long-term scars in a clinical setting.
Translational insight
Preserving adipocyte lineage plasticity during remodeling could offer therapeutic potential to reduce long-term scarring and promote regenerative healing, including follicle and gland restoration.
Signaling pathways
Canonical WNT signaling appears to play a critical role in driving fibrosis and altering adipocyte-lineage cell fate. Activation of β-catenin on its own, for example, leads to a fibrotic phenotype. 76 Notably, in studies overexpressing β-catenin in fibroblasts and fibroblast lineage cells, the layer of DWAT decreases, eventually getting replaced with dense, fibrotic tissue. Applying a reversible WNT activation model, Jussila and colleagues demonstrated that expression of canonical WNT alone was sufficient to cause fibrosis, ECM expansion, and shrinkage of adipocytes. 77
Peroxisome proliferator-activated receptor gamma (PPARγ) signaling, a primary regulator in adipocyte differentiation, is believed to increase the regenerative capacity of DWAT. 78 Interestingly, PPARγ is known to block TGF-β signaling, a prominent factor secreted by macrophages during the inflammatory phase of wound healing. Expression of TGF-β leads to fibroblast activation into myofibroblasts, whereas adipocyte to myofibroblast transition has also been shown to be driven by elevated TGF-β signaling. 79 Inhibition of profibrotic signaling pathways such as TGF-β is therefore promising option to decrease adipocyte-derived fibrosis. In addition to canonical signaling pathways, adipocyte browning has emerged as a mechanism that influences wound healing.
Translational insight
Modulating WNT, TGF-β, and PPARγ pathways in adipose tissue may offer new antifibrotic strategies, particularly in diseases characterized by excessive scarring.
Adipocyte browning
Although adipocytes have historically been categorized as either white or brown, the term “adipocyte browning” has emerged over the past decade and refers to the appearance of brown-like adipocytes within WAT, coined “beige adipocytes” 80 (Fig. 4). These beige adipocytes are transcriptionally distinct from white and brown adipocytes and are believed to appear in WAT depots in situations of increased thermogenic or energy demand. 81 It is posited that transcription factors stimulating the expression of UCP1, a gene deemed critical for the function of brown and beige adipocytes, drive adipocyte browning. Recently, application of lineage tracing of adipocytes from SWAT demonstrated that cells from this layer of adipose could migrate into wound beds and transdifferentiate toward a fibroblast-like fate. Notably, they observe that adipocytes from SWAT underwent browning following wounding, and that inhibition of browning in SWAT decreased macrophage polarization and myofibroblast activation. 18 Given the ability of adipocytes to influence immune and fibroblast activity, recent studies have investigated how adipose tissue and its derivatives can be harnessed therapeutically to enhance wound healing.

Comparison of adipocyte types. Adipocytes have historically been classified as white or brown. White adipocytes are UCP1 negative, contain one large lipid droplet and low mitochondrial density, and function primarily in lipid storage and metabolism. Brown adipocytes play critical roles in thermogenesis and heat production, containing a high mitochondrial density and multiple lipid droplets. Under certain conditions such as low temperature or injury, some white adipocytes undergo browning, reprogramming into beige adipocytes. These cells, similar to brown adipocytes, contain multiple lipid droplets, are UCP1 positive, and have increased mitochondrial density. Created in BioRender. Parker, J. (2025) https://BioRender.com/ehh3oz7.
Translational insight
Manipulating browning of SWAT adipocytes may influence immune and fibrotic activity, presenting a possible therapeutic target.
Adipose tissue and derived cells in wound care
To date, given the complex nature of wound healing, traditional interventions including surgical debridement and infection prevention remain limited in their effectiveness. Adipose-derived therapies, however, have garnered significant attention as novel therapeutic alternatives for wound repair. In the following section, we will summarize the primary ways in which adipose tissue and its derivatives have been applied to wound healing: (1) ASCs; (2) autologous fat grafting; and (3) cell-free adipose tissue extract. A summary of studies applying these different techniques in wound healing is presented in Table 2. Application of these techniques at different stages alongside their limitations is summarized in Table 3.
Summary of studies applying adipose-derived therapeutic strategies in wound healing and fibrosis
Summary of adipose-derived therapies in wound healing
ECM, extracellular matrix; EGF, epidermal growth factor; PDGF, platelet-derived growth factor; TGF-β, transforming growth factor β; VEGF, vascular endothelial growth factor.
Adipose-derived stem cells
ASCs are defined as plastic-adherent, multipotent cells derived from adipose tissue.116,117 These cells are isolated from lipoaspirate, debris and blood removed, and the cells filtered and plated on tissue culture plates (Fig. 5, left). Isolation and expansion of these cells often take in the order of days to weeks prior to use. These cells have been applied in the context of wound healing, and it is posited that they assist with regeneration, reconstruction, and scar reshaping and repair. Studies suggest that ASCs promote improved wound healing at multiple stages; during the inflammatory phase, ASCs are believed to secrete certain cytokines that revert macrophages toward an anti-inflammatory M2 phenotype, 98 whereas they secrete factors including VEGF and PDGF that promote fibroblast migration and proliferation along with neovascularization during the proliferative and remodeling phases.99,100
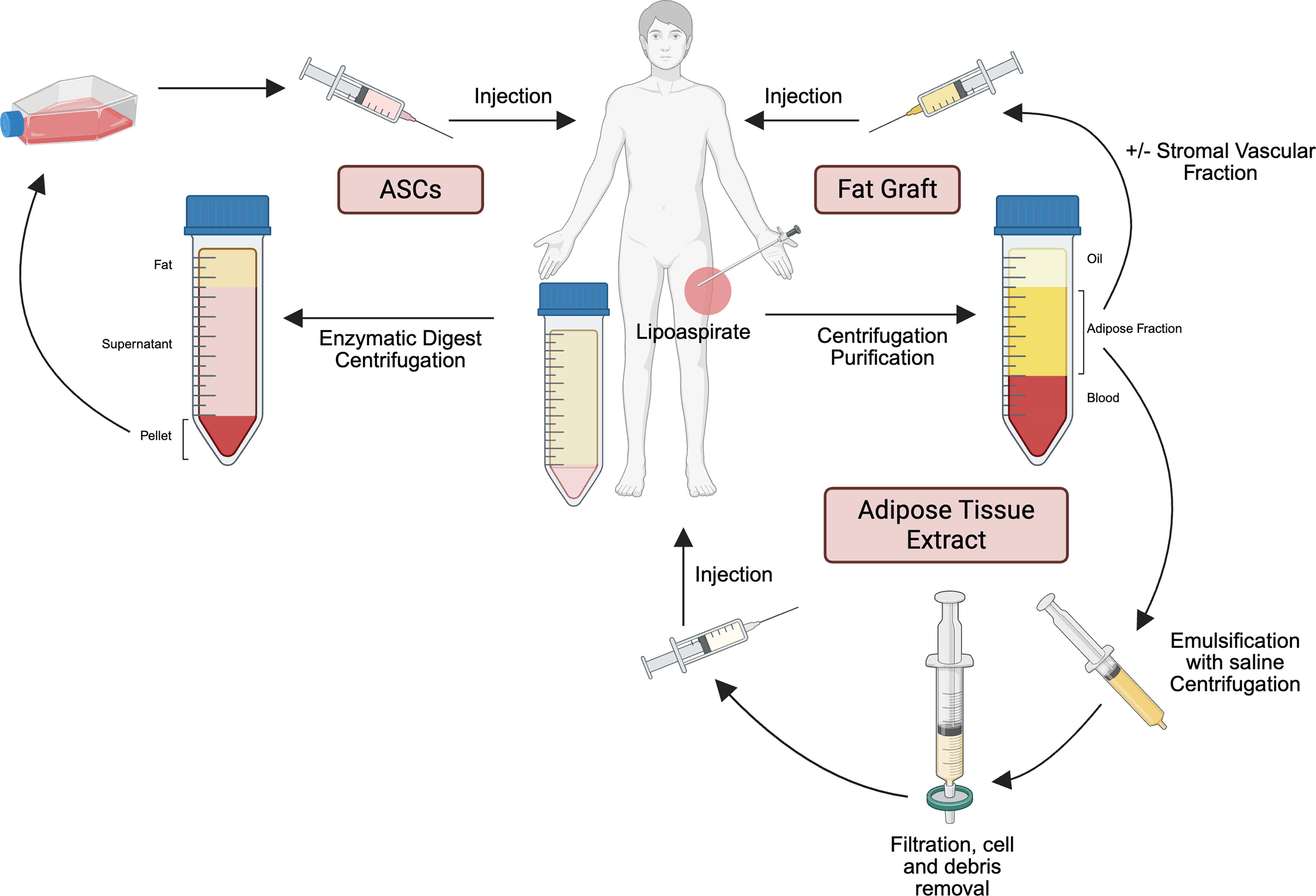
Adipose tissue-derived therapeutics. Left, retrieval and expansion of adipose-derived stem cells (ASCs). Right, retrieval and purification of adipose for autologous fat grafting with or without stromal vascular fraction. Bottom, retrieval and purification of adipose tissue extract. Created in BioRender. Parker, J. (2025) https://BioRender.com/ehh3oz7.
In one preclinical study, ASCs resuspended in an atelocollagen matrix with silicon membrane were applied to wounds in diabetic (db/db) mice. Histology demonstrated that treatment with the ASC-enriched matrix led to increased granulation tissue deposition, as well as improved revascularization and re-epithelialization. 86 Some prospective studies in humans have also been performed. Of note, one group injected ASCs intramuscularly into patients with critical limb ischemia with or without nonhealing ulcers and necrotic feet. 85 After a mean follow-up of 6 months, clinical improvement was observed in over 66% of patients, with decreased pain rating, increased vasculature, and improvements in claudication walking distance.
Autologous fat grafting
During autologous fat grafting, adipose tissue is taken from one site of a patient and placed in another. This is typically performed by retrieving fat from an adipose-rich donor site, such as the abdomen, periumbilical region, buttocks, or thigh using a blunt fat harvesting cannula. Following this, the collected fat is refined to remove components that could negatively impact fat uptake, including oil, dying or dead cells, and blood. Processed fat is then transferred via syringe to the desired recipient site of the patient. 118 In some cases, this technique is enriched using cellular stromal vascular fraction (SVF) 106 (Fig. 5, right). Here, collected fat is processed to separate the tissue into three layers: oil, SVF, and blood. The oil and blood are discarded, whereas the SVF is then used for injection. Enrichment of the graft using the SVF is grounded in the observation that the SVF contains the cellular components, including mature adipocytes, endothelial cells, ECM, ASCs, and vascular cells believed to have a regenerative effect on the target tissue.
In the context of wound healing, autologous fat grafting can be applied directly to the wound site. In one study, for example, lipoaspirate from healthy regions of diabetic foot ulcer patients was injected into the edges of their ulcers. These patients experienced a mean 50% reduction in ulcer size in 4 weeks. At a mean 68 days after the treatment, ulcers from 88% of patients completely resolved. 97 Another study collected fat from the posterior cervical regions of miniature pigs and placed the SVF back into the wounds. They observed that pig wounds treated with autologous fat grafting with SVF demonstrated increased re-epithelialization and decreased wound surface area at postwounding day 7 and 14 relative to control wounds. 89
RIF is another condition for which adipose-based therapy offers significant potential, with fat grafting improving skin architecture following treatment. In one study, Deleon and colleagues enriched fat grafts with CD34+CD146+ ASCs and injected this enriched lipoaspirate into the irradiated scalps of CD1 nude mice. 107 Enriched fat grafts improved graft vascularization and retention and led to improved architecture of the irradiated soft tissue. Further, human trials have also been performed applying autologous fat grafting with SVF for RIF. In one prospective cohort study, 20 patients with RIF underwent injections of autologous fat graft lipoaspirate with SVF derived from healthy donor sites. At 31 months follow-up, these patients experienced improved hydration and neovascularization in the affected areas. 82 Finally, in addition to applications in the context of RIF, groups have demonstrated the benefit of autologous fat grafting for other fibrotic dermal conditions, including chronic wounds and burns.92,119–121
Cell-free adipose tissue extract
A third technique demonstrating potential in wound healing is cell-free adipose tissue extract. To apply this technique, adipose tissue is purified, and the liquid fraction separated, removing cellular and lipid components (Fig. 5, bottom). First developed in the mid 2010s, adipose tissue extract contains growth factors, including PDGF, TGF-β, and epidermal growth factor. 111 In one study, Yin and colleagues loaded a hydrogel with cell-free adipose tissue extract and applied the combination to diabetic wounds. 113 By using a hydrogel formulation, components of the adipose tissue extract were able to release slowly into the wound bed. Seventeen days following treatment, the combined treatment group observed increased capillary density, a small reduction in wound area ratio, and increased cell proliferation. In another study, cell-free adipose tissue extract was topically applied to dermal wounds in mice. 112 Dermal wounds following treatment closed significantly faster relative to control wounds, and in vitro experiments demonstrated that cell-free adipose tissue extract led to enhanced dermal fibroblast proliferation and migration.
Downsides to adipose-derived treatments
Although adipose-derived treatments do show promise for applications in wound healing, these therapies have resulted in adverse events in some patients. For example, fat grafting can be associated with fat necrosis, cyst formation, and volume loss due to variable graft survival.108–110 Additionally, ASCs may exacerbate fibrosis in specific contexts. In SSc, for example, ASCs can exhibit a profibrotic and antiadipogenic phenotype.101,102 The ASCs from SSc patients have been found to abnormally express cell surface markers that may contribute to inadequate immunoregulatory functions. 122
Several challenges limit the widespread clinical application of adipose-derived treatments, including the need for standardization, optimal dosing, and robust comparisons of their efficacy in different clinical settings. The therapeutic consistency of adipose-derived treatments is hindered by a lack of standardized protocols for cell isolation, processing, and characterization, with donor variability and adipose depot source further complicating reproducibility.104,105 Emerging evidence suggests that the efficacy of ASCs is dose-dependent, with subtherapeutic and supratherapeutic doses potentially reducing effectiveness or triggering adverse effects. 103 Comparative studies indicate that ASCs may outperform other cell types, such as bone marrow-derived mesenchymal stem cells, in certain clinical contexts; however, differences in delivery methods, depot origin, and experimental design limit generalizability.123–125 Further, the growth factors in cell-free fat extract have been found to be unstable and quickly degraded in vivo, necessitating delivery strategies that prolong bioactivity and minimize the need for repeated dosing.114,115 These limitations and conflicting findings highlight key opportunities for future research and clinical translation.
Clinical takeaway
Adipose-derived therapies—including ASCs, fat grafting, and cell-free extracts—represent promising regenerative tools for enhancing wound repair. While early studies show encouraging outcomes, challenges remain around reproducibility, standardization, and context-specific effects that must be addressed before broad clinical adoption.
Influence of systemic factors on wound healing
It is important to note that factors such as age, sex, and metabolic state modulate adipose function and wound healing capacity. Increased age is associated with defective adipogenesis, diminished ASC function, and altered expression of collagen and matrix metalloproteinases, impairing wound healing.126,127 Although less explored, sex differences can influence collagen and metalloproteinase expression during healing. 126 Circulating estrogen has also been shown to support adipocyte function and fat graft viability. 128 Both diabetes and obesity contribute to a prolonged and dysregulated inflammatory phase in wound healing, increasing the risk of chronic wounds.129,130 In both animal models and older human subjects (approximately 60–70 years), diabetic adipose tissue displays chronic senescent cell accumulation and reduced secretion of key repair mediators. 131 Transplantation of diabetic adipose tissue into nondiabetic hosts has been shown to impair the wound healing process. 131 Similarly, obesity or a high-fat diet promotes vascular insufficiency and chronic inflammation, disrupting the normal reparative processes.126,132 Obesity creates a microenvironment within adipose tissue characterized by an imbalance of adipokines, with elevated proinflammatory mediators such as leptin and resistin alongside decreased anti-inflammatory adiponectin contributing to impaired healing. 130 In addition to systemic factors, local molecular signaling pathways also play a key role in regulating adipocyte behavior and wound healing outcomes.
Clinical takeaway
Accounting for factors such as age, sex, and metabolic state or chronic disease may improve our understanding of adipose tissue behavior and allow for more personalized wound care to enhance therapeutic efficacy.
The study of transcriptional and proteomic signatures of adipocyte subpopulations
Advancements in omic-based techniques enabling the identification of transcriptional or proteomic signatures at cellular or even subcellular resolution have opened the door for groups to explore the functions of distinct subpopulation of a cell type in a given context. 133 Our group, for example, has investigated fibroblast heterogeneity in detail in various models of fibrosis, identifying profibrotic and proregenerative populations.134–136 As adipocytes are technically difficult cells to capture on single cell RNA-seq, more recent developments in applying these transcriptomic techniques to nuclei are a promising alternative circumventing these challenges. 137 These techniques are relatively recent developments and, to our knowledge, have not yet been applied in the contexts of skin scarring or fibrosis; however, they have been applied in investigations on thermoregulation and high-fat diet.
For example, one study determined that a subpopulation of adipocytes collected from inguinal WAT had a distinctly thermogenic signature, and that subpopulations changed in their proportions following cold stimulation and eventual return to thermoneutrality, 138 illustrating not only that subpopulations of adipocytes exhibit disparate functional signatures but also that their profiles are dynamic in nature. In another study that explored mouse adipose tissue remodeling during obesity, So et al. identified diverging subpopulations of adipocytes, with hypertrophic adipocytes displaying signatures of senescence, inflammation, and stress. 139 Whether parallel subpopulations within adipocytes in SWAT or DWAT are present in normal and wounded skin will be critical to explore in order to further understand adipocytes’ roles in healing and fibrosis.
FUTURE DIRECTIONS AND CONCLUSIONS
In this review, we have summarized the field’s current understanding of the role of dermal and subcutaneous adipose tissue during wound healing. Although much remains to be understood regarding the exact mechanisms by which adipose tissue participates across the different stages of repair, work within this field has established that adipocytes within WAT are a dynamic cell type that plays roles in dermal homeostasis and fibrosis. Recent work also suggests WAT and adipose tissue-derived cells offer significant potential for therapeutic application in wound healing and other dermal fibroses.
Fibroblasts, one of the key drivers of the proliferative phase of wound healing, demonstrate significant heterogeneity within both normal and injured skin. 29 Multiomic approaches have allowed for molecular explorations into the broad transcriptional and proteomic landscape of this cell type within the skin. These investigations have provided insights into which subpopulations of fibroblasts and signaling pathways to target, prevent, or reduce scar formation. With growing understanding of the participation of adipose tissue in wound healing, a similar approach to assess adipocyte plasticity and heterogeneity at a molecular level may allow us to understand which subpopulations of adipocytes to target in order to improve wound repair. Learning how adipocytes change when perturbed, and how different subpopulations of adipocytes are influenced by external cues from their environment and adjacent cells, will help shed light on adipose tissue function and therapeutic applications in years to come.
TAKE HOME MESSAGES
WAT plays important roles in energy storage, metabolism, thermal insulation, and mechanical cushioning
WAT situated in the skin (DWAT and SWAT) has been shown to play critical roles in homeostasis and injury
While WAT participates in scar formation, decreased adipose tissue in fibrotic conditions such as scleroderma suggests that adipose tissue may have a protective effect as well.
Adipose tissue participates in the inflammatory phase of wound healing by assisting in the recruitment of immune cells to the wound site
WAT also participates in proliferation and remodeling phases of wound healing, wherein they help recruit scar-depositing cells (fibroblasts). Some also participate directly in scar deposition.
Translational research exploring adipose tissue and its derivatives to treat chronic wounds and fibrotic conditions bode promise as future therapeutic strategies.
SUMMARY
Wound healing and scarring remain a clinical burden with few treatments available. Although the field has gained significant insights into understanding the roles of cell types and signaling pathways involved in this process, details regarding the profibrotic and protective roles of adipose tissue remain incompletely understood. Researchers have demonstrated that adipocytes from WAT participate in the inflammatory phase through signal mediation and response, while also participating in the proliferation and remodeling phases by recruiting scar-depositing fibroblasts. Studies investigating adipocyte plasticity have also shown how these cells can transition into fibroblasts to participate directly in scar formation.
Meanwhile, clinical fibrotic conditions often present with decreased adipose tissue, suggesting that adipose tissue offers protective benefits as well. Adipose tissue and its derivatives such as autologous fat grafting, ASCs, and cell-free adipose tissue extract, show promise as potential therapies to improve wound healing and remodel fibrosis in clinical settings in the future.
ACKNOWLEDGEMENTS AND FUNDING SOURCES
Figures 1, 3, 4, and 5 were created with Biorender.com. This work was supported by the Hagey Laboratory for Pediatric Regenerative Medicine and the Wu-Tsai HPA Moonshot 2. J.B.P. was supported by the Bio-X Bowes Graduate Student Fellowship. M.T.L. and D.C.W. were supported by NIH Grant AR081343/DE 032677.
Footnotes
AUTHORS’ CONTRIBUTIONS
J.B.P. and M.F.G. wrote the article. D.A., N.E.L., C.V., A.G.M., M.K., M.T.L., and D.C.W. edited the article. Written consent was received from all authors to submit the article, and all authors accept complete responsibility for the contents of the articles.
AUTHOR DISCLOSURE AND GHOSTWRITING
No competing financial interests exist. No ghostwriters were used to write this article.