
Abstract
Significance:
Keloids represent a persistent clinical challenge, with recurrence rates approaching 100% after conventional surgical excision. Historically viewed as fibroblast-driven scars, emerging evidence positions immune dysregulation as a central driver of keloid pathogenesis, reshaping diagnostic and therapeutic paradigms.
Recent Advances:
Multi-omics and single-cell analyses reveal profound alterations in immune cell populations and cytokine networks within keloid tissue. M2 macrophages, mast cells, Th2 and Th17 subsets, and dendritic cells dominate the inflammatory microenvironment, sustaining fibroblast activation and excessive extracellular matrix deposition. Key signaling axes—including transforming growth factor-β (TGF-β)/Smad, IL-4/IL-13, IL-6/JAK-STAT-3, and PI3K/AKT/mTOR—intersect with mechanical stress pathways, creating a self-perpetuating fibrotic loop. Notably, soluble human leukocyte antigen-E emerges as a predictive biomarker for disease progression and recurrence, while tissue-resident memory T cells may underlie postoperative relapse.
Therapeutic Implications:
Beyond corticosteroids and silicone gel, immune-targeted strategies are gaining traction. Dupilumab (IL-4/IL-13 blockade) demonstrates clinical efficacy in reducing keloid burden and pruritus. Experimental approaches targeting TGF-β signaling (Fresolizumab, AVID200), NKG2A/CD94 checkpoints (Monalizumab), IL-6 (Tocilizumab), and TSLP (Tezepelumab) hold promise for precision immunotherapy. Localized delivery and combination regimens may optimize outcomes while minimizing systemic toxicity.
Critical Issues and Future Directions:
The absence of validated keloid models and large-scale trials limits translation. Future research must integrate immune profiling, biomarker validation, and mechanistic modeling to enable personalized interventions. Immune dysregulation is not merely an epiphenomenon—it is the Achilles’ heel of keloids, offering unprecedented opportunities for targeted therapy and recurrence prevention.
BACKGROUND
Scope and significance
Keloid is a fibroproliferative tumor resulting from abnormal wound healing in the skin. 1 It tends to grow progressively, often extending beyond the boundaries of the original lesion, and is challenging to resolve spontaneously.1,2 Keloids are typically associated with symptoms such as redness, swelling, and itching. Furthermore, keloids located on joints and facial areas may restrict limb movement and lead to disfigurement, imposing significant physiological and psychological burdens on patients.3,4 Despite the availability of various treatment options for keloids, including intralesional injections, surgical interventions, radiotherapy, and laser therapy,1,5 there remains a lack of systematic evidence supporting specific therapeutic approaches. The recurrence rate following simple surgical excision can reach up to 100%, 6 and surgery may exacerbate collagen deposition, leading to larger lesions postoperatively.7,8 In addition, although the diagnostic accuracy for keloids is approximately 81%, no specific diagnostic biomarkers have been identified. 3 Therefore, elucidating the underlying pathogenesis of keloids, identifying specific diagnostic markers, establishing safe and effective therapeutic targets, and preventing disease recurrence remain key challenges in keloid management.
Min Zhang, MD

Keloids are known to be influenced by genetic factors, wound tension, and gender. Accumulating evidence suggests that immune-mediated inflammation plays a significant role in keloid formation. 9 As early as the 1970s, researchers identified an association between keloids and immune inflammation.10,11 The development of advanced technologies and experimental methodologies has sparked considerable interest and extensive research into the immunological mechanisms underlying keloid pathogenesis over the past two decades. In 2022, Zhang Yixin et al. 3 discovered that T cells and their surface, the Natural Killer Group 2 A (NKG2A)-CD94–human leukocyte antigens (HLA-E) axis, may serve as potential biomarkers and therapeutic targets for keloids. In 2024, Rolin G et al. 12 first demonstrated that M1 and M2 macrophages exert distinct effects on in vitro keloid fibroblasts; moreover, restoring the M1/M2 balance could inhibit keloid fibrosis. Therefore, elucidating the roles of immune cells in keloids is expected to provide novel insights into both the diagnosis and treatment of the diseases.
Translational relevance
The absence of effective diagnostic markers and treatment methods for keloids is closely related to the fact that the molecular mechanism of the disease remains unclear. Inflammation plays a crucial role in the occurrence, development, and recurrence of keloids. However, at present, there is still a lack of systematic research on the mechanism of immunity in keloids. This makes translational research work aimed at developing effective prevention and treatment strategies for keloids even more difficult.
Clinical relevance
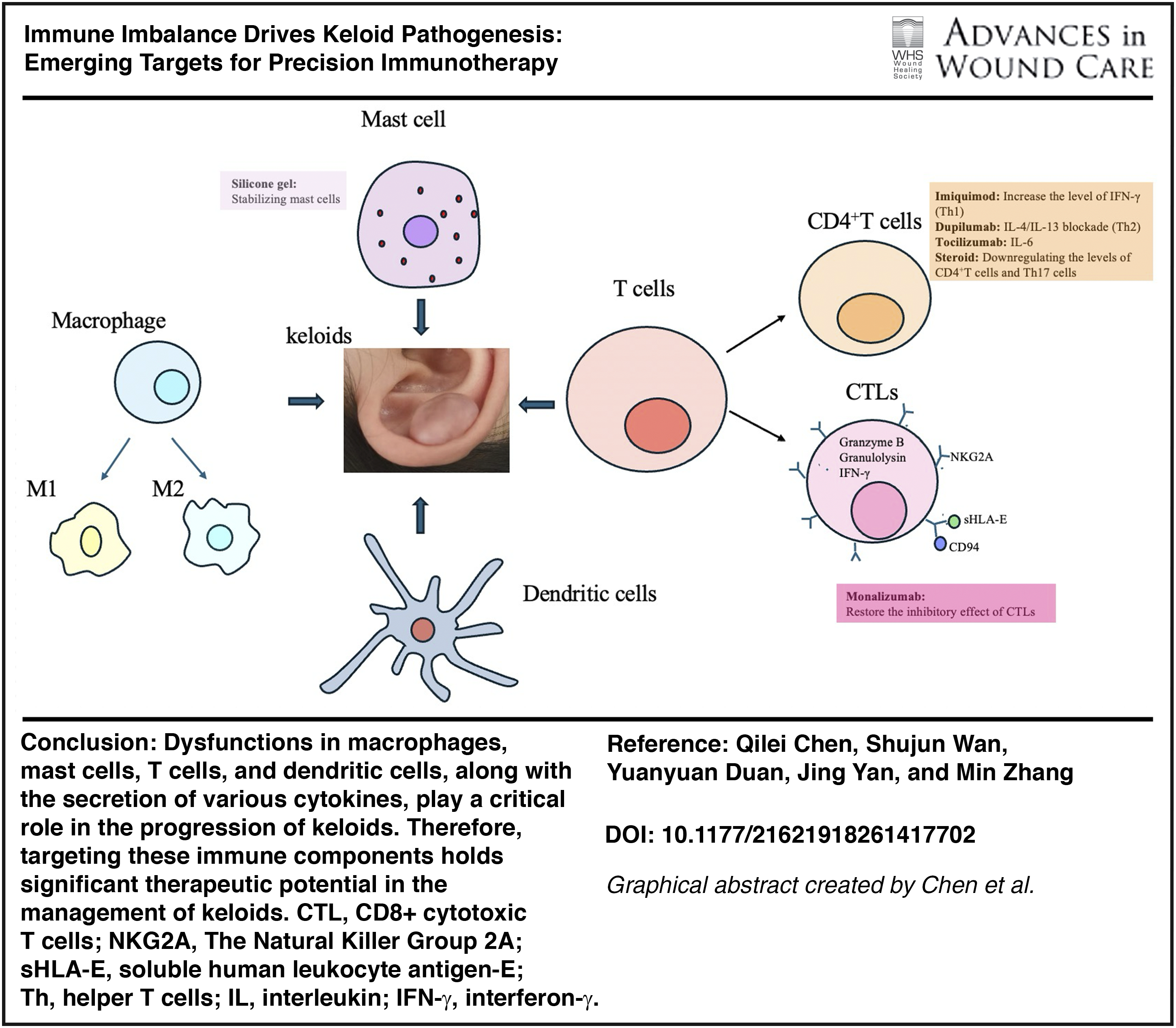
Through our analysis, we have identified that immune cells and the cytokines they secrete are crucial for the pathophysiology of keloids. Notably, M2 macrophages play a pivotal role in the dysregulated growth and invasion of keloids, while mast cells contribute to angiogenesis. In addition, T cells are implicated in collagen deposition, and the cytokine transforming growth factor-β (TGF-β), along with its associated signaling pathways, is involved throughout the entire pathogenesis of keloids by interacting with other signaling cascades. Investigating alterations in immune cell populations and their secreted cytokines, as well as their molecular mechanisms, is essential for improving diagnosis and treatment approaches while preventing recurrence of keloids.
OVERVIEW
Immune cells
Macrophages
Circulating blood mononuclear cells can infiltrate exposed wounds and differentiate into either M1 (classically activated, CD86+) or M2 (alternatively activated, CD163+) macrophages. 13 Normal wound healing involves a coordinated transition from an early inflammatory phase dominated by M1 macrophages to a reparative phase characterized by M2 macrophages. In contrast, dysregulation of this transition may lead to prolonged inflammation, delayed wound closure, and excessive scar formation. 14
M1 macrophages–fibroblast crosstalk
Monocytes differentiate into M1 macrophages under the influence of interferon-γ (IFN-γ), interleukin-12 (IL-12), tumor necrosis factor-alpha (TNF-α), or pathogen-associated molecular patterns such as lipopolysaccharides. 15 M1 macrophages play a crucial role in engulfing invading pathogenic microorganisms and contribute to the inflammatory phase of wound healing. Furthermore, matrix metalloproteinases secreted by M1 macrophages exhibit anti-fibrotic properties.15–17
M2 macrophages–fibroblast crosstalk in keloids
Monocytes differentiate into M2 macrophages in response to IL-4, IL-13, or apoptotic neutrophils. These M2 macrophages are capable of secreting various growth factors, including platelet-derived growth factor (PDGF), TGF-β, insulin-like growth factor-1 (IGF1), and vascular endothelial growth factor (VEGF), which promote the abnormal proliferation of wound fibroblasts. 16 Zhou et al. 18 demonstrated that increased infiltration of M2 macrophages significantly contributes to the uncontrolled growth and invasion of keloid fibroblasts. Furthermore, studies have indicated a bidirectional cell–cell communication or symbiotic relationship between fibroblasts and macrophages in keloids, suggesting that these cells can exchange essential growth factors through their interactions. For instance, fibroblasts sustain the expression of chemokine C–C motif ligand 2 (CCL2), colony-stimulating factor 1, and IL-6, which leads to the recruitment and activation of macrophages at the wound site. In turn, macrophages secrete TGF-β, IGF1, VEGF, and PDGF that further promote the abnormal proliferation of fibroblasts19,20 (Fig. 1A).
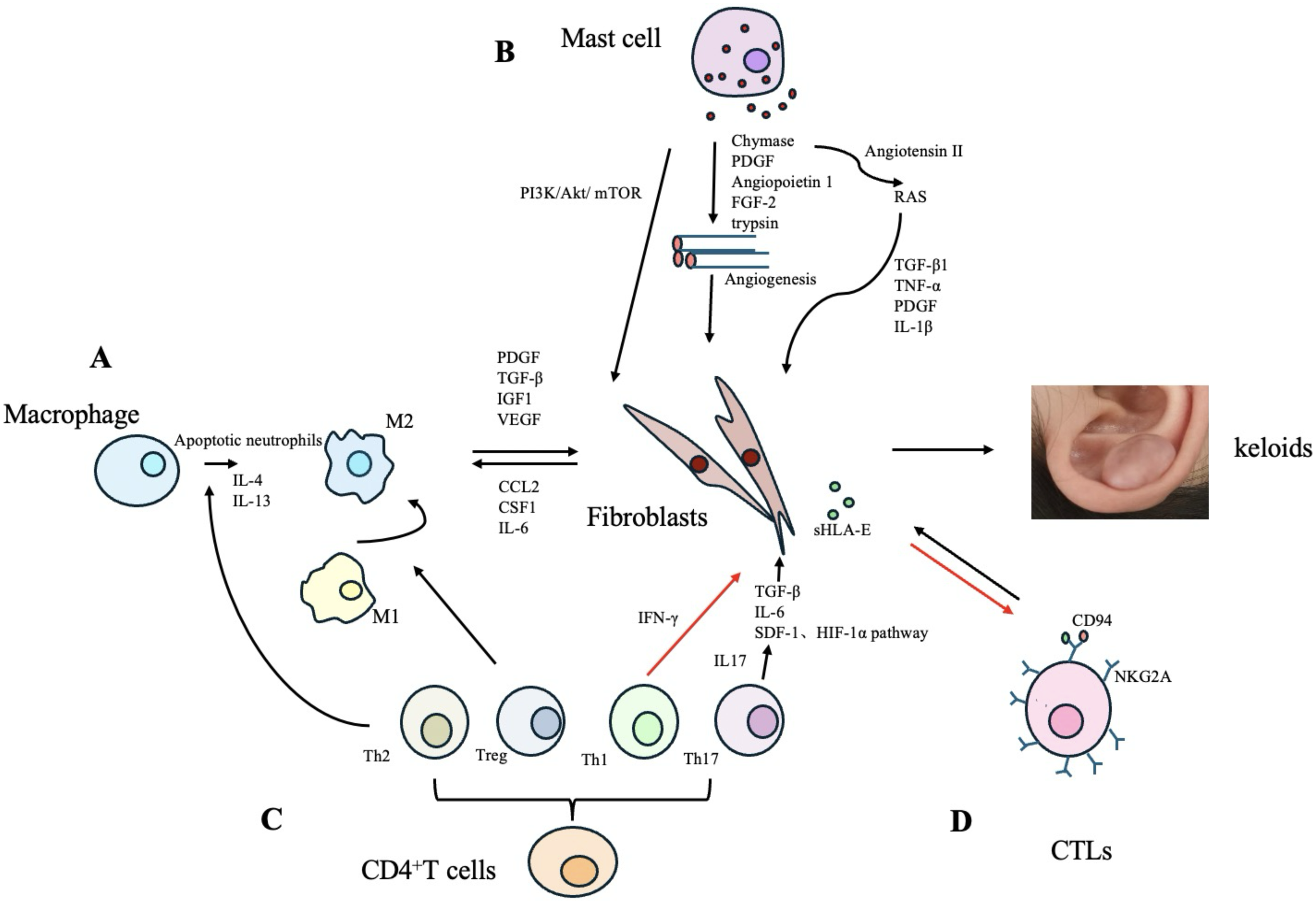
The role of immune cells and their secreted cytokines in the pathogenesis of keloids.
Mast cells
Compared to the mast cells in mouse embryos on day 18, those present on day 15 were fewer in number, exhibited lower maturity, and were unable to undergo degranulation following injury. The scarless wound healing process observed on day 15 can be disrupted by the immediate injection of mast cell lysate post-injury. Furthermore, when compared with Kit+/+ mice from the same litter, KitW/W-v mice with mast cell deficiency displayed significantly smaller scars during the wound healing process at day 18. 32 These results indicated that mature mast cells significantly contributed to scar formation.
Dysregulation of mast cell degranulation in keloids
RNA sequencing analysis has revealed a significant increase in the number of mast cells present in keloids. 33 Studies have indicated that mast cell degranulation frequently interacts with activated fibroblasts, suggesting a potential bidirectional communication between these two cell types. 34 Furthermore, green tea extract (EGCG), which contains polyphenols such as epigallocatechin-3-gallate, has been shown to inhibit this interaction by suppressing the phosphatidylinositol 3-kinase (PI3K)/Akt/mTOR signaling pathway, thereby reducing type I collagen production 21 (Fig. 1B).
Mast cells may promote angiogenesis by secreting PDGF, chymotrypsin, angiopoietin-1, fibroblast growth factor 2 (FGF-2), and trypsin at the wound site22,23 (Fig. 1B). Increased numbers and activity of trypsin and chymotrypsin in mast cells may stimulate fibroblast proliferation and collagen deposition through the TGF-β1/Smad signaling pathway in keloids.35,36 Furthermore, chymotrypsin enhances the expression of angiotensin II, thereby activating the local renin-angiotensin system, leading to elevated levels of TGF-β1, TNF-α, PDGF, and IL-1β, which further promote fibroblast activation and collagen production 24 (Fig. 1B).
Mast cell degranulation and itchy keloids
Hawash et al. 28 found that, compared with non-itchy keloids, the number of mast cells and their intracellular granules was significantly higher in itchy keloids. Mast cells were observed to undergo degranulation and secrete histamine and nerve growth factor (NGF). On one hand, NGF may induce mast cells to release additional histamine; on the other hand, histamine promotes keratinocytes to secrete NGF, thereby establishing a positive feedback loop between histamine and NGF, which leads to histamine-driven itching in keloids.28,37 The above findings indicate that mast cells are closely involved in the pathogenesis of keloids, and that targeting mast cells and their secreted mediators may represent a promising therapeutic strategy for keloids, particularly for itchy keloids.
T cells
CD8+ T cells
CTLs imbalance and fibrosis in keloids
Scientists have found that the levels of CD8+ cytotoxic T cells [also known as cytotoxic T Lymphocytes (CTLs)] are downregulated in both tissue and peripheral blood samples from patients with keloids, as revealed by single-cell RNA sequencing (scRNA-seq) analysis, particularly for CTLs expressing granzyme B, granulysin, and IFN-γ in peripheral blood.3,32 Studies have demonstrated that both direct and indirect cultures of CTLs and fibroblasts significantly inhibit the proliferation of fibroblasts, leading to a reduction in their growth vitality and an increase in apoptosis. 38 It has been hypothesized that the abnormal proliferation of fibroblasts and excessive deposition of extracellular matrix (ECM) observed in keloids may be associated with the reduced levels of CTLs. 38
The NKG2A protein is expressed on the surface of CTLs, where it forms a complex with CD94 and regulates the activation and proliferation of CTLs. 3 Keloid-derived fibroblasts can secrete soluble human leukocyte antigen-E (sHLA-E) into the surrounding tissue and circulating blood, where it interacts with the NKG2A/CD94 complex on the surface of CTLs, thereby inhibiting CTL activity and proliferation and promoting tolerance to fibroblast growth 3 (Fig. 1D). sHLA-E is closely linked to the development of keloids. A study has shown that sHLA-E holds promise as a diagnostic biomarker for keloids, with a sensitivity and specificity of 83.69% and 92.16%, respectively. 3 In addition, sHLA-E is closely associated with treatment response and recurrence in keloids. It demonstrates strong predictive value for the clinical efficacy of intratumoral injection in keloid patients. 32
Research has shown that the efficacy rate of intratumoral dexamethasone injection alone is 73%, but this increases to 92% when dexamethasone is combined with 5-fluorouracil in the treatment of keloids. This improvement may be attributed to the enhancement of CTLs activity by 5-fluorouracil, 39 suggesting that CTLs play a beneficial role in improving keloid outcomes.
The role of TRM in the recurrence of keloids
However, flow cytometry analysis has revealed an increased proportion of CD8+ tissue-resident memory T cells (TRM) within the T cell population in keloid tissues. 40 Although these cells may provide partial protection against secondary wound infection, they can also trigger a sustained inflammatory response, thereby promoting keloid progression.40,41 In addition, due to their long lifespan and tissue-restricted localization, TRMs are thought to contribute to postoperative keloid recurrence, making them a potential key factor in disease relapse. 42 Targeting TRMs may represent a promising therapeutic strategy for preventing keloid recurrence.
CD4+ T cells
Th-cell imbalance and fibrosis in keloids
Immature CD4+ T cells can differentiate into various subsets of helper T (Th) cells, including Th1, Th2, Th17, and regulatory T cells (Tregs).25,26 Th1 cells suppress fibroblast proliferation and the expression of type I and type III collagen genes by secreting IFN-γ. 27 Th2 cells secrete IL-4 and IL-13, thereby promoting collagen synthesis and the abnormal deposition of reticular fibrin. 28 Dupilumab, a monoclonal antibody targeting the Th2 axis, shows significant therapeutic efficacy in the treatment of keloids. 43 The infiltration of Th17 cells and the expression of IL-17 are elevated in keloids. On one hand, IL-17 induces neutrophils and macrophages to produce cytokines such as TGF-β, thereby promoting collagen deposition and fibrosis. 29 On the other hand, IL-17 contributes to fibrotic processes by enhancing IL-6 expression and activating the stromal cell-derived factor-1 and hypoxia-inducible factor 1α signaling pathways 30 (Fig. 1C). In addition, studies have shown that markers associated with T cell activation and migration (ICOS, CCR7), Th2 cells (IL4R, CCL11, TNFSF4/OX40L), Th1 cells (CXCL9, CXCL10, CXCL11), and Th17/Th22 cells (CCL20, S100As) are upregulated in keloids. Similar expression patterns have also been observed in the surrounding tissue of keloids. 33
Treg-cell imbalance and fibrosis in keloids
Compared with the T cell population without Tregs, the T cell population enriched with Tregs demonstrated a significantly enhanced capacity to inhibit collagen synthesis in keloid-derived fibroblasts. 44 However, in the absence of other CD4+ T cell subsets, Tregs promoted collagen synthesis in fibroblasts. 45 The above findings indicate that Tregs exert differential effects on fibroblast collagen synthesis depending on their interactions with other T cell subsets. Moreover, Tregs are capable of secreting interleukin-10 (IL-10) and prostaglandin E2 to inhibit tissue fibrosis. 46
Research has shown that the accumulation of Tregs in injured muscle tissue promotes the polarization of M1 macrophages toward the M2 phenotype. 47 Qi et al. 31 also reported that Tregs can induce the polarization of M1 macrophages toward the M2 phenotype and enhance the deposition of type III collagen in keloid tissue (Fig. 1C). The proportion of CD4+CD25+FOXP3+ Tregs is reduced in the peripheral blood mononuclear cells of patients with multiple keloids, which may be attributed to increased local infiltration of Tregs in the affected tissue.22,40 However, the specific mechanism by which the increased local accumulation of Tregs influence keloids tissue remains unclear.
CD4+/CD8+ T cell imbalance and keloids
Furthermore, Boyce et al. 48 found that, unlike the gradual decrease in the CD4+/CD8+ T cell ratio observed during normal wound healing, the CD4+/CD8+ T cell ratio remained elevated in keloids for an extended period, resulting in prolonged inf lammatory activity and heightened response characteristic of the early healing stage. Therefore, modulating the CD4+/CD8+ T cell ratio may be expected to be a new direction for the treatment of keloids.
Dendritic cells
Research has shown that the expression of dendritic cell (DC) surface markers CD80 and CD86 is elevated in keloids. In addition, DCs associated with atopic dermatitis (OX40L+, FCεR1+) also show an increasing trend in keloids. 33 DC precursor cells (possibly monocytes) are recruited by autoimmune antigens, migrate from blood vessels to the lesion site, and differentiate into inf lammatory DCs within keloids, thereby attracting memory T cells. 49 DCs and memory T cells interact, secrete excessive amounts of A Disintegrin and Metalloproteinase 10 (ADAM10) at the lesion site, induce fibroblast activation and proliferation, and thus promote keloid progression. 49 Moreover, keloids may recruit and activate inf lammatory DC precursor cells (monocytes), trigger inflammatory responses independent of autoimmune antigen presentation, and are associated with Th2, Th17, and Th22 differentiation.49,50
Far more than that, inflammatory DCs may also activate fibroblasts to secrete metalloproteinases such as ADAM10, ADAM17, and CD10, which interact with the novel profibrotic mediator soluble Ephrin-B2 in the skin, thereby promoting fibroblast activation and proliferation. 51 As is well known, most immune effector molecules, including cytokines and chemokines, are synthesized as inactive precursors, and their activation requires proteolytic processing. In this context, ADAM10 and ADAM17 serve as key proteolytic enzymes involved in cell differentiation and proliferation. ProTNF and Notch receptors are proteolytically processed by these enzymes and converted into active effectors, thereby promoting fibroblast proliferation and ECM deposition. 49 Therefore, DC-related metalloproteinases may represent promising therapeutic targets for keloids.
Comprehensive analysis of a systems biology approach to keloids
RNA and scRNA sequencing analyses have revealed the presence of immune-related differentially expressed genes between keloids and normal tissues. 52 These genes are predominantly enriched in key signaling pathways associated with keloid pathogenesis, including TNF, IL-17, TGF-β, mitogen-activated protein kinase (MAPK), and JAK-signal transducer and activator of transcription (STAT). 52 Notably, the genes encoding prostaglandin F(2α) receptor (PTGFR), Retinol-binding protein 5 (RBP5), and leukemic inhibitory factor (LIF) were found to be significantly upregulated, suggesting their potential as biomarkers for keloids. 52 Moreover, the expressions of these three genes vary across different fibroblast subtypes, potentially influencing the progression of keloid formation. 52 Additional scRNA sequencing data demonstrated a marked increase in the proportion of Th17 cells in keloids compared to normal scars. Functional studies suggest that Th17 cells may contribute to fibroblast proliferation, migration, and collagen deposition through the secretion of IL-17A, thereby exacerbating keloids development. 53
Tumor necrosis factor superfamily member 4 (TNFSF4) is found to be upregulated in keloids. 54 Enrichment analysis indicates that the elevated expression of TNFSF4 is associated with various immune processes, including neutrophil chemotaxis, dendritic cell chemotaxis, and the regulation of antigen processing and presentation. 54 Results from scRNA sequencing demonstrate that TNFSF4 expression is significantly increased in mesenchymal fibroblasts, which are key cells involved in skin fibrosis. The high levels of TNFSF4 enhance intercellular interactions within fibrosis-related pathways, particularly those involving fibronectin 1 and collagen. 54 In addition, mesenchymal fibroblasts exhibit significant expression of TNFSF4, which notably upregulated gene expression in extracellular matrix tissues and the wound healing process. 54 This study elucidated the impact of the immune molecule TNFSF4 on both protein and gene expression levels within keloids. 54 Moreover, transcriptomic profiling revealed that differentially expressed genes in keloids are closely associated with immune responses and ECM deposition. Consistent with these findings, subsequent proteomic analysis showed that differentially expressed proteins are predominantly involved in ECM-related pathways, leading to the hypothesis that local immune dysregulation may trigger excessive ECM accumulation. 55 Systems biological analysis suggests that differentially expressed immune genes significantly contribute to the progression of keloids by modulating the expression of cytokines and influencing the activity of fibroblasts.
Immune cells interaction in keloids
The dysregulated immune cells present in keloids engage in interactions with one another, collectively impacting the progression of the disease. Keloid-derived macrophages have the capacity to significantly enhance the expression of Foxp3 in CD4+ T cells, thereby facilitating their differentiation into Treg cells 56 (Fig. 2A). In addition, Tregs have the capacity to inhibit the activation of both CD4+ and CD8+ T cells while simultaneously promoting the polarization of M2 macrophages 57 (Fig. 2B). The upregulated levels of IL-4 and IL-13, secreted by Th2 cells, can facilitate the polarization of macrophages towards the M2 phenotype, thereby promoting the development of keloids 28 (Fig. 2C). In keloids, the level of IL-10 secreted by FOXP3+CD8-memory T cells is diminished, resulting in an overactive and dysregulated T cell response. 40 Inflammatory DCs within keloids may originate from blood monocytes and interact with memory T cells, thereby inducing the activation and proliferation of fibroblasts 49 (Fig. 2D). Furthermore, the inflammatory DCs in keloids are closely associated with the differentiation of Th cell subsets49,50 (Fig. 2D).
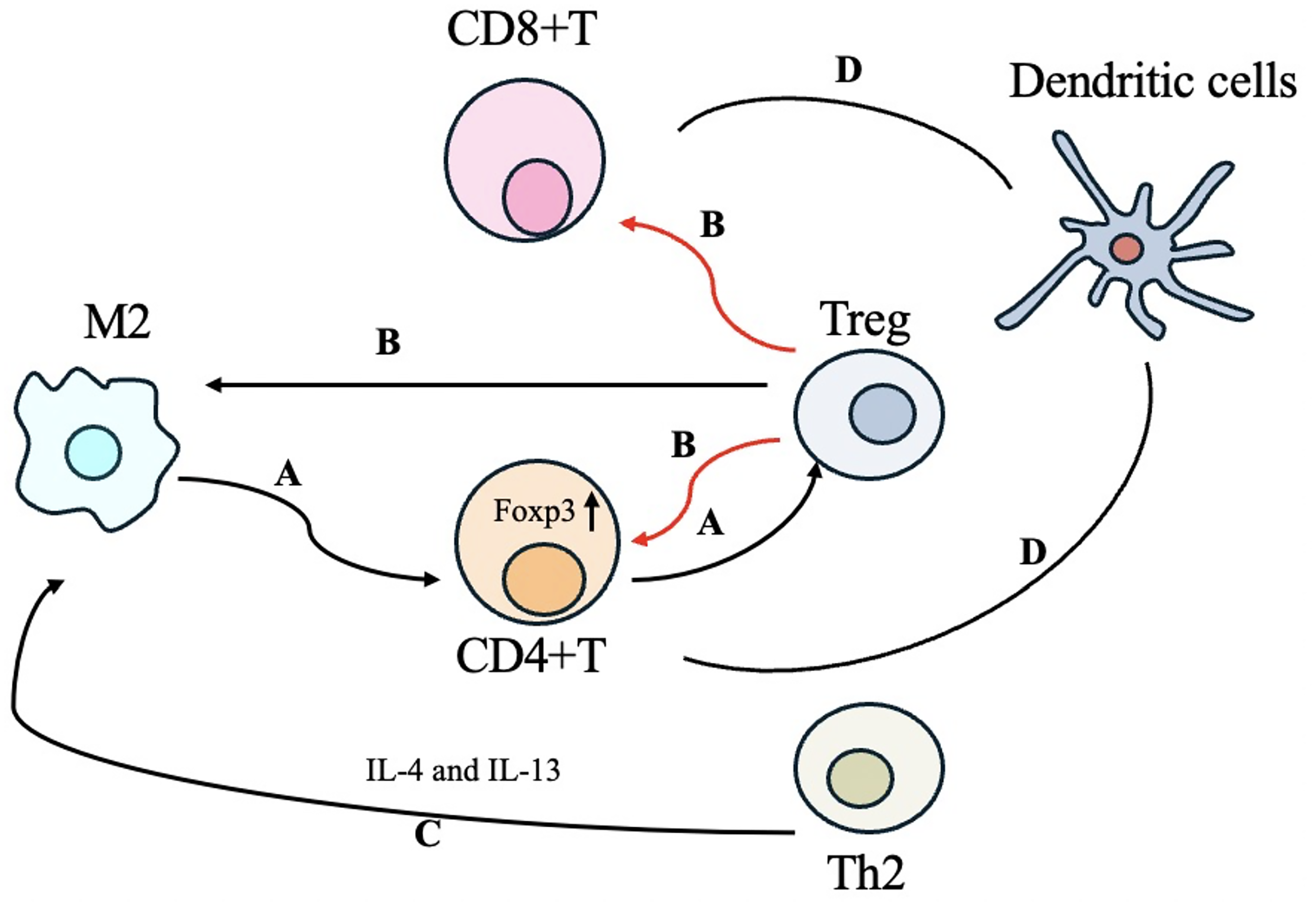
Immune cell interaction in keloids.
Genetic susceptibility, mechanical tension, and wound healing dynamics interact with immune dysregulation in keloids
Genetic susceptibility interacts with immune dysregulation in keloids
Among the general Black and Hispanic populations, the incidence of keloids ranges from 4.5–6.2% to as high as 16%. 58 In contrast, reports indicate that the incidence among Caucasian individuals is less than 1%. 58 It is underscoring the significant role of genetics in keloid formation. Studies have shown that the expression levels of CXCL8 and IL-17RA, both of which play a role in the IL-17 signaling pathway and fibroblast activation, are diminished in keloids. Moreover, CXCL3 is typically recognized as a neutrophil chemotactic agent during inflammation and is found to be overexpressed in keloid fibroblasts. 59
Mechanical tension interacts with immune dysregulation in keloids
Wong et al. 60 conducted a comparative analysis of gene expression in stretched versus non-stretched wounds using a mouse high-throughput screening model, employing microarray technology. The findings revealed that mechanical stimulation was significantly associated with the continuous signaling of T-cell-dependent Th2 cytokines (IL-4 and IL-13) and chemokines (MCP-1). 60 These results suggest that the fibrotic pathways regulated by T cells exhibit a pronounced degree of mechanical reactivity; the mechanical forces can induce a chronic inf lammatory state through immune-mediated activation of both local and systemic cell populations. 60
Immune changes in wound healing dynamics during the normal wound healing and keloids process
Normal wound healing commences with hemostasis (Fig. 3). 71 The primary cells involved in this process are platelets, while the crucial matrix component is fibrinogen. 72 Hemostasis is achieved through the contraction of the blood vessel wall, platelet aggregation, and the formation of a platelet thrombus, alongside the conversion of soluble fibrinogen into insoluble fibrin chains that establish a fibrin network. 72 Furthermore, platelets directly bind to, activate, or trigger the extravasation of monocytes, neutrophils, and even T lymphocytes to the injured site by releasing heterogeneous protein molecules such as TGF-β, p-selectin, CD40L, and RANTES. 73
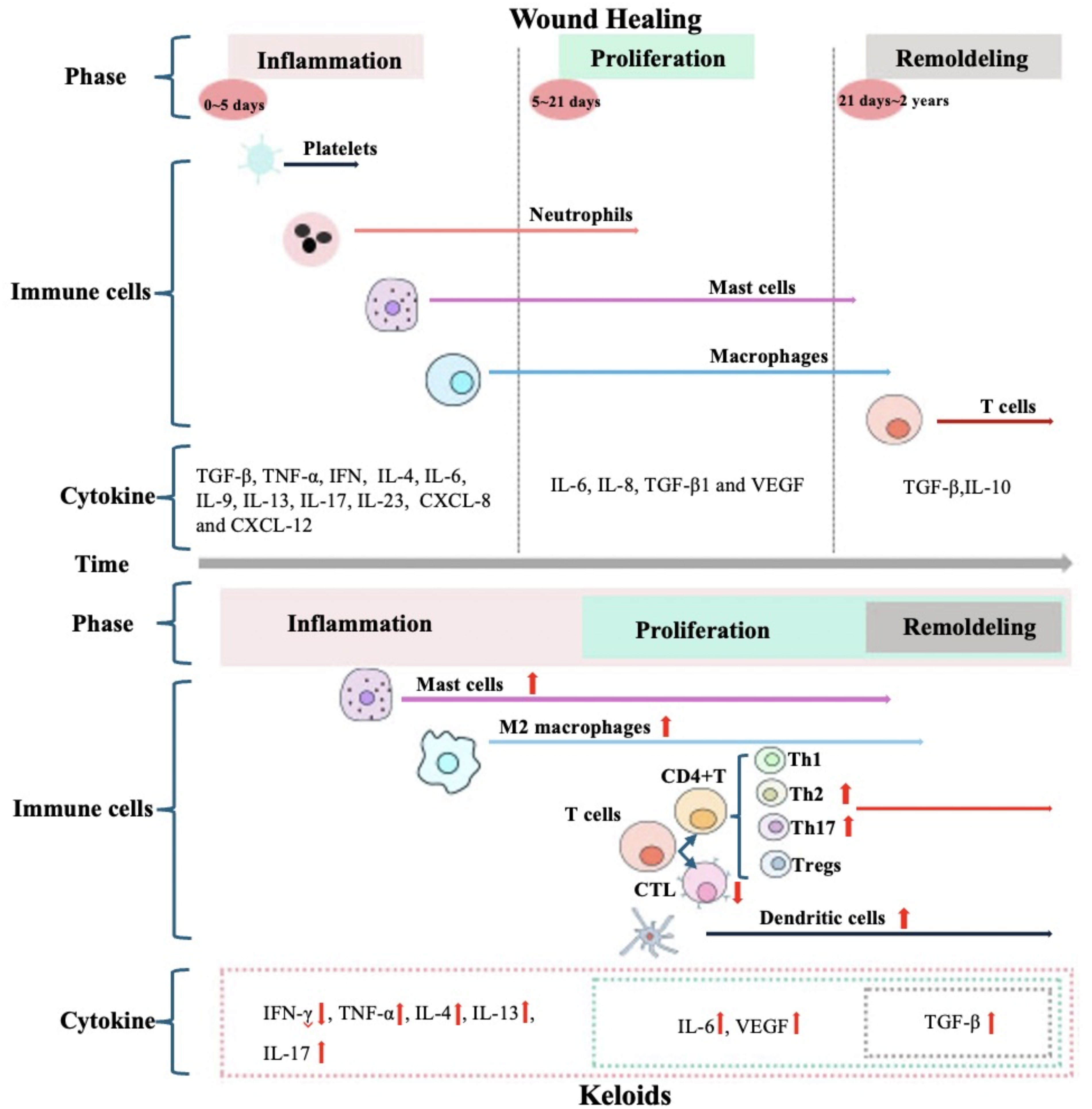
Immune changes in wound healing dynamics during the normal wound healing and keloids process. Wound Healing: Inflammatory phase spans from 0 to day 5 post-injury,22,61 granulocytes (neutrophils and eosinophils), macrophages and mast cells participate in this phase, as well as cytokines such as TGF-β, TNF-α, IFN-γ, interleukins (IL-4, IL-6, IL-9, IL-13, IL-17, IL-23), and chemokines (CXCL-8, CXCL-12), 62 M1 transforms into M2 macrophages and persists during the proliferation and remodeling stages, 63 Mast cells play a pivotal role in later proliferative and remodeling phases. 64 Proliferative phase occurs between 5 and 21 days post-injury,22,65 IL-6, IL-8, TGF-β1, and VEGF are crucial mediators in this phase;22,66,67 Remodeling phase typically occurs between 21 days and 2 years post-injury,22,68 T cells, IL-10, and TGF-β are involved in it.22,69 Keloids: Excessive inflammatory responses run through the phases of inflammation, proliferation and remodeling, 70 the levels of M2 macrophages, mast cells, Th2 and Th17 cells, dendritic cells, and the cytokines they secrete—such as TNF-α, IL-4, IL-13 and IL-17—are significantly upregulated,18,24,28,29,33 CTLs and IFN-γ are downregulated,3,32 the levels of IL-6, VEGF, and TGF-β that are involved in the normal proliferation and remodeling phases of wounds also elevated.19,20,30 CTLs, CD8+ cytotoxic T cells; CXCL, C–X–C motif chemokine ligand; IFN-γ, interferon-γ; IL, interleukin; TGF-β, transforming growth factor-β; TNF-α, tumor necrosis factor-alpha; VEGF, vascular endothelial growth factor.
Recruited immune cells subsequently trigger the onset of the inflammatory phase, which generally spans from day 0 to day 5 post-injury.22,61 During this phase, a diverse array of immune cells—including granulocytes (neutrophils and eosinophils), macrophages, and mast cells—participate in orchestrating the inflammatory response. M1 macrophages predominate during the early stages of inflammation and play a crucial role in coordinating initial cellular responses. In contrast, M2 macrophages subsequently become dominant at the wound site; They are critical for resolving inflammation as well as promoting fibroblast proliferation and tissue remodeling. 63 Mast cells also exert significant influence by facilitating angiogenesis, collagen deposition, and granulation tissue formation, playing a pivotal role in later proliferative and remodeling phases. 64 Concurrently, immune cells secrete a wide range of bioactive mediators including cytokines such as TGF-β, TNF-α, IFN-γ, interleukins (IL-4, IL-6, IL-9, IL-13, IL-17, IL-23), chemokines (CXCL8, CXCL12), along with multiple proteases—all collectively regulating inflammatory processes during wound healing. 62
A successful anti-inflammatory phase is signified by the onset of the proliferative phase, which typically occurs between 5 and 21 days post-injury.22,65 IL-6 and IL-8 not only facilitate inflammation but also contribute to tissue fibrosis. 22 During this phase, TGF-β1 emerges as a pivotal factor driving the differentiation of fibroblasts into myofibroblasts. 66 Concurrently, the expression of VEGF is upregulated, thereby promoting angiogenesis at the wound site to accommodate the demands for rapid proliferation. 67 However, excessive secretion of VEGF is closely associated with scar formation. 74
After the proliferative phase, wound healing enters a remodeling stage that typically occurs between 21 days and 2 years post-injury.22,68 This process necessitates the degradation of immature ECM components and the subsequent remodeling of mature substrates. 62 T cells recruit specialized (pro-lysis) macrophages and establish cell–cell interactions with skin cells during this stage. 22 Furthermore, research has demonstrated that IL-10, TGF-β signaling pathways, and TGF-β type II receptors play critical roles in the remodeling process.22,69
In normal wound healing, the inflammatory phase, proliferative phase, and remodeling phase interweave and transition in an orderly manner. During this natural progression, excessive intervention in treatment is generally unnecessary. Conversely, persistent and excessive inf lammatory responses are critical contributors to keloid formation. 70 Notably, elevated levels of M2 macrophages, 18 mast cells, 33 Th2 cells, 28 Th17 cells, 29 dendritic cells, 33 as well as key cytokines19,20,24,28–30 persist throughout the inflammatory and remodeling phases. Therefore, in the management of keloids, it is essential to closely monitor the variations in these immune factors and implement appropriate therapeutic measures.
Immunotherapy
Why immune dysregulation matters clinically
Currently, various treatment methods are available for keloids.1,5 However, there remains a lack of specific agents targeting this condition. Moreover, the existing therapeutic approaches exhibit limited efficacy in completely preventing recurrence. Even when surgical resection is combined with radiotherapy, the recurrence rate remains approximately 22%. 75 Consequently, there is an urgent need to develop more effective treatment strategies. In previous work, we summarized the pathogenic roles of various immune cells in keloid formation, suggesting that immunotherapy may represent a promising breakthrough in addressing the challenges associated with keloid management. In this context, we outline potential targets and pathways related to keloid immunotherapy (Table 1).
The immunotherapeutic strategies of keloids in clinical employ
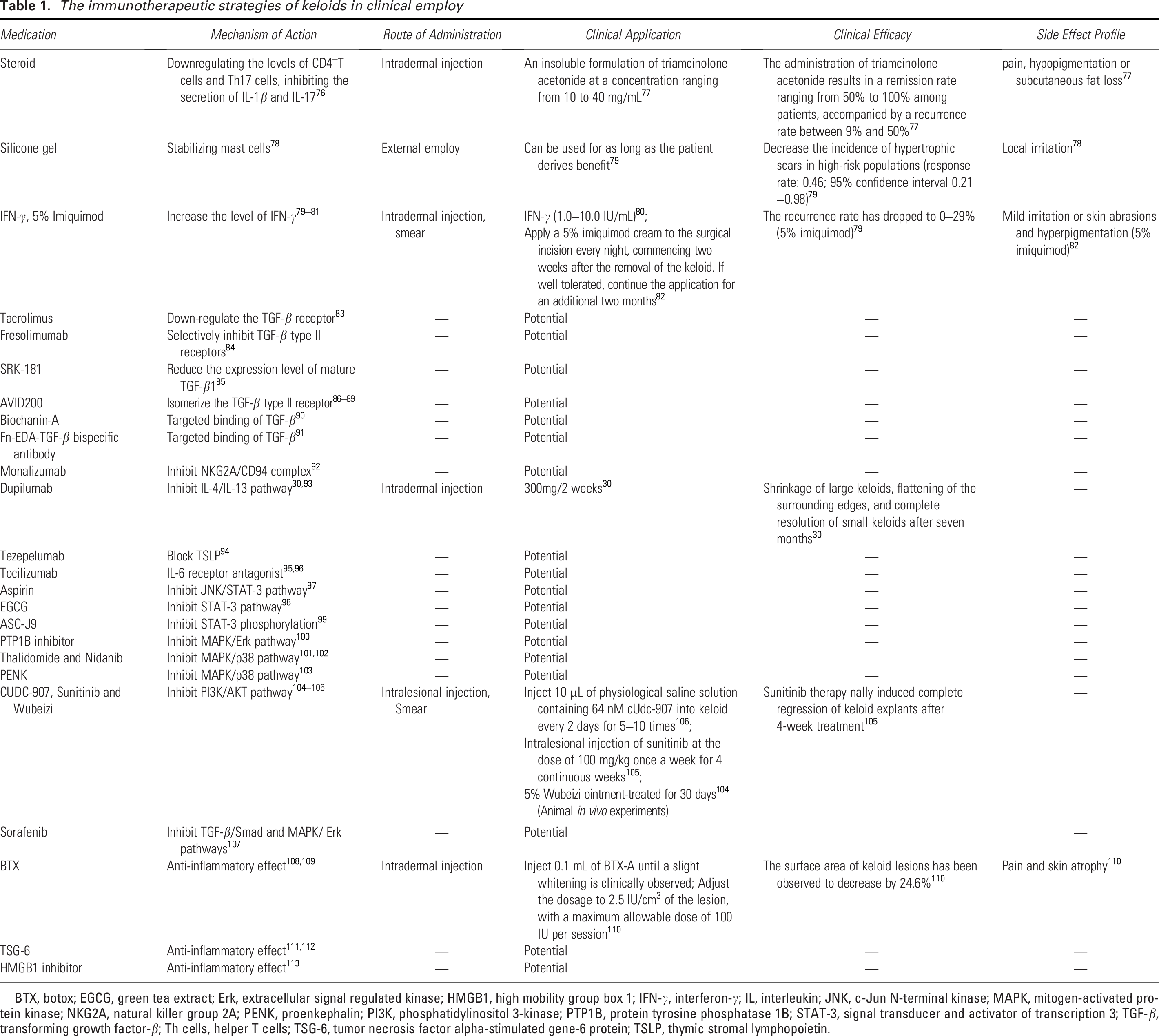
BTX, botox; EGCG, green tea extract; Erk, extracellular signal regulated kinase; HMGB1, high mobility group box 1; IFN-γ, interferon-γ; IL, interleukin; JNK, c-Jun N-terminal kinase; MAPK, mitogen-activated protein kinase; NKG2A, natural killer group 2A; PENK, proenkephalin; PI3K, phosphatidylinositol 3-kinase; PTP1B, protein tyrosine phosphatase 1B; STAT-3, signal transducer and activator of transcription 3; TGF-β, transforming growth factor-β; Th cells, helper T cells; TSG-6, tumor necrosis factor alpha-stimulated gene-6 protein; TSLP, thymic stromal lymphopoietin.
Immunotherapy targeted immune cell populations
Steroids
Steroids represent one of the commonly used therapeutic approaches for keloids. 114 Research has shown that steroids can downregulate CD4+ T cell and Th17 cell levels, suppress the secretion of IL-1β and IL-17, and contribute to the modulation of the inflammatory response. 77 Intradermal injection of triamcinolone acetonide is among the recommended treatment options for keloids.7,76
Silicone gel
When mature wounds exhibit itching and red streaks, daily application of silicone gel dressings has been shown to effectively reduce scar tissue formation. 82 Research has demonstrated that the hydration and occlusive environment created by silicone gel plays a crucial role in stabilizing mast cells, thereby effectively blocking the inflammatory cascade triggered by mast cell activation. 115
Immunotherapy targeted immune cytokines and signaling pathways
IFN-γ
As early as 2002, IFN-γ was incorporated into the international clinical guidelines for scar management and demonstrated significant therapeutic efficacy in the treatment of keloids. 78 IFN-γ can downregulate the expression of type I and type III collagen in keloids, antagonize the overexpression of IL-6 and TGF-β, and inhibit fibroblast proliferation, thereby effectively reducing keloid volume while alleviating associated symptoms. 116 When IFN-γ was used in combination with retinoic acid, 5-fluorouracil, and bleomycin, it showed favorable therapeutic effects on keloids.80,116 5% imiquimod is a topical immune response modulator that inhibits collagen synthesis by inducing IFN-γ production. 81 In addition, studies have shown that IFN-γ not only induces ferroptosis in fibroblasts but also inhibits the synthesis of solute carrier family 7 member 11 and solute carrier family 3 (activators of dibasic and neutral amino acid transport), member 2 (SLC3A2) by downregulating serine protease inhibitor 2, thereby enhancing fibroblast sensitivity to ferroptosis. 79
TGF-β
TGF-β functions as a vital mediator in keloids formation, promoting collagen synthesis by fibroblasts.20,45,117 Therefore, targeting TGF-β-related receptors or signaling pathways may represent a promising therapeutic strategy for the management of keloids.
Tacrolimus is an immunosuppressive agent capable of inhibiting T cell activation. In keloids, tacrolimus effectively suppresses the TGF-β/Smad signaling pathway in fibroblasts by downregulating TGF-β receptors, thereby reducing fibroblast proliferation, migration, and collagen production. 118 Fresolizumab selectively inhibits TGF-β type II receptors, leading to reduced expression of collagen type I alpha 1 chain in fibroblasts in a mouse fibrosis model, indicating its potential for treating fibrotic diseases. 83 In addition, studies have shown a significant reduction in fibroblast infiltration in deep dermal tissue following Fresolizumab treatment. 84 Therefore, Fresolizumab holds therapeutic promise for the management of keloids. SRK-181 is a tumor immunotherapy agent that targets and binds to the TGF-β1 precursor, thereby reducing the expression of mature TGF-β1, 119 which is speculated to have therapeutic potential for keloids. AVID200, as a highly selective inhibitor, has the potential to promote the binding of the TGF-βII receptor to TGF-β3, thereby inhibiting collagen synthesis and deposition.85–88 Phase I clinical trials have further confirmed that AVID200 exhibits favorable anti-fibrotic effects,85,86,89 supporting its potential role in mitigating abnormal fibrosis associated with keloids. In addition, studies have demonstrated that Biochanin-A suppresses bleomycin-induced pulmonary fibrosis by targeting TGF-β and blocking the TGF-β1/Smad signaling pathway, with minimal immunological or histological side effects, 120 thus indicating its potential as a high-quality therapeutic agent for keloids.
However, due to the involvement of TGF-β in multiple biological processes, prolonged inhibition of TGF-β may lead to a range of adverse effects. 90 Studies have shown that systemic administration of TGF-β inhibitors not only exerts anti-fibrotic effects but also disrupts anti-inflammatory responses, potentially leading to disease exacerbation. 121 Local injection of TGF-β antibodies or inhibitors is proposed as an alternative therapeutic strategy for keloids. Furthermore, fibronectin extra domain A (Fn EDA), as a key component of the ECM, is specifically expressed in fibrotic lesions. The Fn EDA–TGF-β bispecific antibody designed by McGaraughty can deliver TGF-β antibodies specifically to the site of renal fibrosis by targeting Fn EDA. 122 Given that keloids are enriched in ECM, bispecific molecular targeted therapy directed against both Fn EDA and TGF-β may offer greater efficacy with fewer side effects compared to applied TGF-β antibodies alone.
NKG2A/CD94
Monalizumab, a novel inhibitor of the NKG2A/CD94 complex, is widely used in tumor immunotherapy. 91 Studies have shown that Monalizumab enhances anti-tumor immunity by blocking NKG2A and promoting the functional recovery of CTLs. 91 Given that CTLs can inhibit fibroblast proliferation and activity in keloids, it is hypothesized that Monalizumab may reduce excessive ECM deposition by restoring the inhibitory effect of CTLs on fibroblasts.
IL-4/IL-13 pathway
Dupilumab could inhibit the Th2-type inflammatory response by targeting the IL-4/IL-13 pathway, and it served as an effective biological agent for the treatment of moderate to severe atopic dermatitis. 92 In 2020, Diaz et al. 43 reported a novel application of Dupilumab in the treatment of keloids: in patients with severe atopic dermatitis and concomitant keloids, subcutaneous administration of Dupilumab 300 mg every two weeks led to significant improvement in atopic dermatitis, a 50% reduction in fibrotic plaques, shrinkage of large keloids, flattening of the surrounding edges, and complete resolution of small keloids after seven months. Furthermore, in a phase 3 clinical trial involving patients with moderate to severe Alzheimer’s disease, Dupilumab was found to alleviate pruritic symptoms by blocking neuronal itch signaling, 93 indicating its potential for improving itching associated with keloids. Targeting the IL-4/IL-13 pathway with Dupilumab may represent a promising therapeutic strategy for patients with keloids, particularly those experiencing itching symptoms, and further large-scale clinical trials are warranted.
TSLP
Thymic stromal lymphopoietin (TSLP) may promote skin fibrosis by stimulating Th2 cells to produce cytokines, and that neutralization of TSLP or genetic deletion of TSLPR significantly suppresses fibroblast proliferation and skin fibrosis.123,124 Given the significant elevation of TSLP levels in keloid tissues and their surrounding areas, 33 it is hypothesized that TSLP may play a crucial role in keloid fibrosis. Tezepelumab is a human monoclonal antibody targeting TSLP. In clinical practice, it has been widely used for the treatment of severe and uncontrolled asthma in adults and adolescents. 125 Studies have shown that Tezepelumab may reduce immune cell levels in keloids. 125 Although no clinical trials have been conducted on the employ of Tezepelumab for keloid treatment, it is anticipated to be a potential therapeutic agent with effects comparable to those of Dupilumab.
IL-6
Research has shown that IL-6 levels are elevated in keloids. It may promote fibroblast proliferation, migration, inflammatory signaling, and ECM deposition, thereby playing a significant role in the progression of keloids. 88 Tocilizumab is an IL-6 receptor (IL-6R) antagonist that is currently approved for the treatment of rheumatoid arthritis and juvenile idiopathic arthritis. The 2018 ACR conference reported the results of a phase III randomized controlled trial of tocilizumab in systemic sclerosis, 94 which demonstrated that tocilizumab significantly reduced pulmonary fibrosis, with treated patients exhibiting improved forced vital capacity compared to the placebo group. 95 Therefore, it is hypothesized that tocilizumab may serve as a promising therapeutic candidate for skin fibrosis.
STAT-3 pathway
Studies have shown that the STAT-3 signaling pathway is activated in keloids,96,126 and that inhibition of STAT-3 expression or phosphorylation can significantly reduce the proliferation and migration of keloid fibroblasts, as well as collagen synthesis. 126 In vivo experiments have demonstrated that aspirin may inhibit inf lammatory responses and reduce scar formation following injury through modulation of the JNK/STAT-3 signaling pathway. 127 In addition, EGCG has been demonstrated to inhibit the STAT-3 signaling pathway, thereby contributing to improvements in keloid formation. 128 ASC-J9 functions as a phosphorylation inhibitor of STAT-3 and exhibits efficacy in suppressing the proliferation of keloid fibroblasts. 129
The elevated expression and phosphorylation levels of STAT-3 in keloids are associated with the activation of JAK2. 87 Inhibitors targeting the JAK2/STAT-3 pathway or STAT-3 siRNA have been shown to effectively inhibit the progression of keloids.87,128 Oral administration of small-molecular-weight JAK inhibitors has proven effective in treating systemic sclerosis affecting skin or lungs. 97
MAPK Pathway
The MAPK signaling pathway constitutes a complex network that includes extracellular signal-regulated kinases (Erk), c-Jun N-terminal kinases (JNK), and p38 kinases, all of which play a crucial role in the progression of keloids. 98 The down regulation of protein tyrosine phosphatase 1B can inhibit the activity of keloid fibroblasts through the MAPK/Erk pathway. 99 Thalidomide and nintedanib, which target p38 in the MAPK pathway, have demonstrated efficacy in restricting the growth of keloid fibroblasts.130,131 Silencing Osteomodulin may ameliorate fibroblast proliferation by inhibiting p38. 100 In addition, proenkephalin derived from placental mesenchymal stem cells has been shown to inhibit the proliferation and migration of keloid fibroblasts while promoting apoptosis through the suppression of p38. 132
PI3K/AKT Pathway
It has been reported that differentially expressed genes in keloids are significantly enriched within the PI3K/AKT pathway. 101 CUDC-907, sunitinib, and Rhus chinensis Mill. ointment have been demonstrated to effectively target and inhibit the PI3K/AKT signaling pathway. These agents exhibit significant therapeutic potential in preclinical models of keloids.102,103,133
Crosstalk Signaling Pathways
MAPK and TGF-β1/Smad Signaling Pathway
Research has demonstrated the existence of crosstalk between the MAPK and TGF-β1/Smad signaling pathways (Fig. 4). 141 In addition to its role in binding to the TGF-β receptor and modulating Smad expression, elevated levels of TGF-β1 observed in keloids can also activate the MAPK signaling pathway, which subsequently influences the phosphorylation of Smad proteins and promotes ECM deposition 138 (Fig. 4B). The multi-kinase inhibitor-sorafenib has been shown to simultaneously inhibit both intracellular TGF-β/Smad and MAPK/Erk signaling pathways, thereby inducing growth arrest in keloid fibroblasts. 142
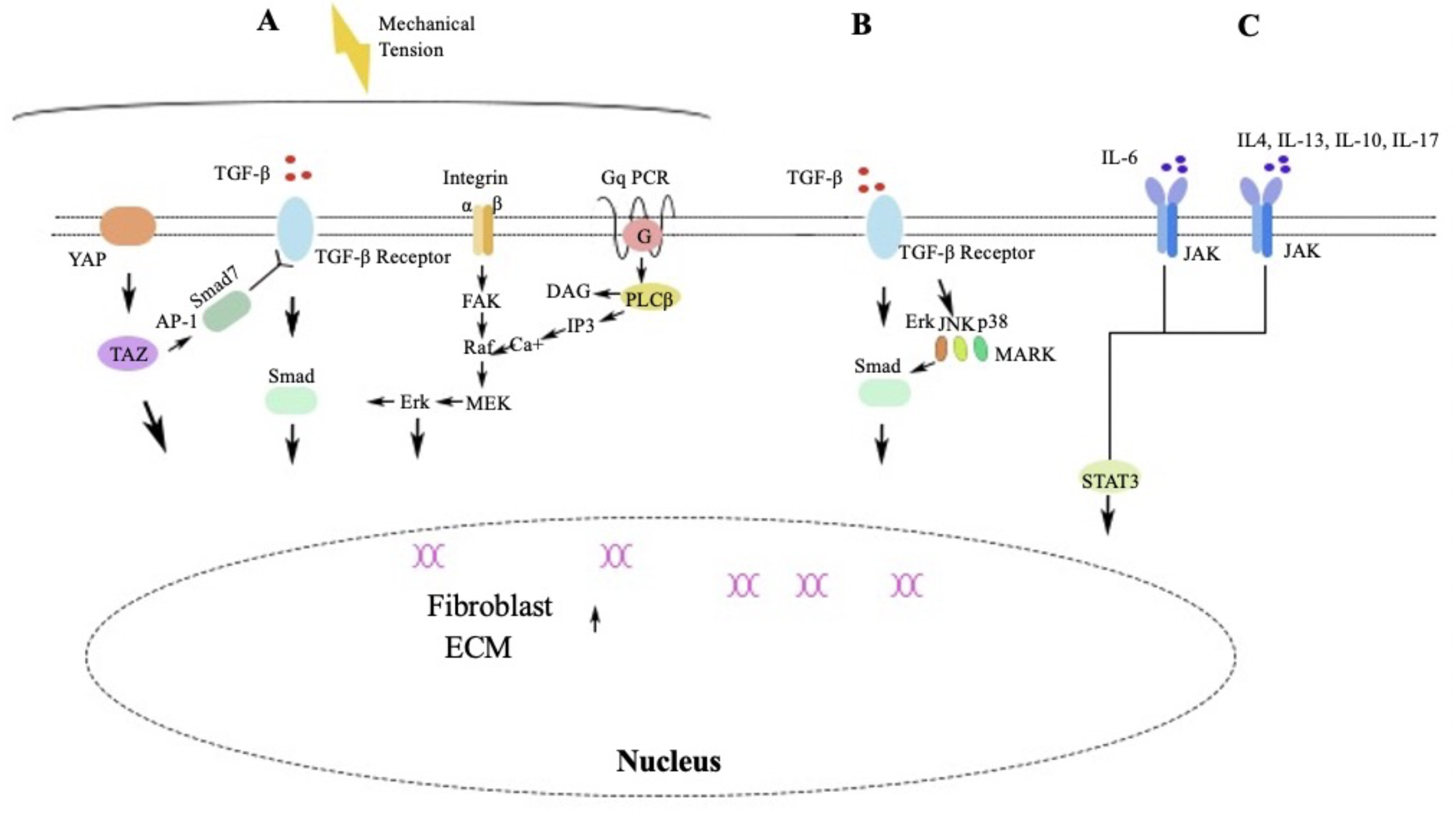
Mutually Interfering Pathways.
Mechanical Transduction Pathways
The interplay between mechanical transduction pathways and TGF-β is increasingly acknowledged in the literature. 135 Mechanical force stimulates integrins, thereby enhancing fibroblast proliferation, differentiation, and collagen synthesis through the FAK/Erk signaling pathway,104,135 which is also involved in the activation of the TGF-β/Smad pathway 105 (Fig. 4A). Yes-associated protein (YAP)/Transcriptional coactivator with PDZ-binding motif (TAZ) serves as a crucial mechanical sensor. 106 It can decrease Smad7 levels by inhibiting activator protein-1 (AP-1), thereby facilitating the activation of the TGF-β/Smad signaling pathway 137 (Fig. 4A). Stretching stimulation promotes the synthesis of inositol triphosphate (IP3). 136 It triggers the release of Ca2+ from intracellular calcium stores, subsequently enhancing fibroblast proliferation through the Raf/MEK/Erk signaling pathway, which interacts indirectly with the TGF-β/Smad signaling pathway 107 (Fig. 4A). When addressing mechanical force stimulation in relation to keloid growth, a combined inhibition of multiple pathways may yield more effective results.
IL-6 and the JAK/STAT Signaling Pathway
IL-6 specifically induces the overactivation of the STAT-3 and JAK/STAT pathways through Gremlin, a bone morphogenetic protein antagonist, and plays a crucial role in promoting fibrosis. 139 In addition, Th2 and Th17 cytokines, including IL-4, IL-10, IL-13, and IL-17, can also facilitate keloid progression through the JAK/STAT signaling pathway33,140 (Fig. 4C). Therefore, alongside inhibiting IL-6, the adjunctive administration of inhibitors targeting Th2 and Th17 cytokines may enhance therapeutic efficacy.
Others
Botox (BTX) is widely used in the field of aesthetics, where it can reduce wrinkles, inhibit masseter hypertrophy, and improve facial contours by inhibiting acetylcholine release at the neuromuscular junction. Recently, studies have demonstrated that botulinum toxin exhibits anti-inflammatory effects on wounds and scars. 143 It has also been found to be more effective in treating keloids compared to intradermal corticosteroid and placebo injections. 144
Tumor necrosis factor alpha-stimulated gene-6 protein (TSG-6) is a secreted glycoprotein containing multiple adjacent functional domains. As TSG-6 can bind to various proteoglycans and polysaccharides, it inhibits neutrophil migration, regulates protease networks, and reduces inflammatory responses by stabilizing or remodeling the ECM. 145 Studies have shown that the level of TSG-6 in keloids is significantly lower than that in normal skin, and this expression level is negatively correlated with keloid severity. 146 In addition, TSG-6 may reduce the ratio of TGF-β1 to TGF-β3, 147 as well as the levels of collagen, myeloperoxidase, and inflammatory cytokines including IL-1β, IL-6, and TNF-α, 148 indicating its significant clinical potential in the treatment of keloids.
High mobility group box 1 (HMGB1) is a driver of inflammatory responses. During skin injury, HMGB1 induces inflammatory cell infiltration and promotes collagen synthesis. Researchers have discovered that HMGB1 inhibitors can significantly enhance the treatment of keloids. 149 Furthermore, polydeoxyribonucleotides that inhibit the secretion of HMGB1 may mitigate the inflammatory response and reduce keloid formation in rat models. 149
Review of keloid models
The evidence supporting current keloid treatments primarily derives from low-quality studies, non-randomized trials, and case reports, limiting the ability to make evidence-based decisions regarding therapy. Ideally, effective therapies should be based on a thorough understanding of the underlying pathology and should be developed and assessed through preclinical models. However, to date, there remains no optimal keloid model available for research purposes. The existing keloid models can be broadly categorized into in vitro and in vivo models.
In vitro models encompass both indirect and direct co-culture systems. 150 Indirect co-culture primarily involves the cultivation of two distinct cell types in separate chambers, 150 while direct co-culture includes mixed monolayer cultures as well as full-thickness skin equivalent cultures composed of keloid-derived cells, which represents the in vitro culture model that most closely mimics the in vivo environment. 150
In vivo models of keloids encompass both human and animal subjects. In vivo models for human keloids are less frequently used, as They present a potential risk for exacerbating the progression of preexisting keloids.151,152 Animal models for keloid research are typically established by inducing keloids or implanting human keloid tissue. However, the implantation of human keloid cells or tissue fragments into animal models has yielded more successful results, 153 compared with inducing keloids which often product hypertrophic scar rather than true keloids. 108 The researchers further enhanced this method by implanting a hydrogel matrix containing keloid fibroblasts into nude mice, simultaneously exposing it to IL-6. This approach led to the development of nodular masses accompanied by the synthesis of new collagen. 109 To enhance the similarity between animal and human models, researchers have attempted to transplant human keloid tissue into immunodeficient mice 111 to create humanized mouse models. Nonetheless, any animal model for keloids has an important limitation: keloids only occur naturally in humans. 154
In contrast, in vitro culture models may be more closely aligned with our objective of studying human keloids. In recent years, researchers have dedicated themselves to advancing the development of tissue culture models. For instance, in 2024, Lee et al. 155 utilized keloid fibroblasts and endothelial cells to establish a novel three-dimensional (3D) keloid spheroid model. Furthermore, certain CRISPR/Cas9-based lineage tracing technologies have opened new avenues for constructing human skin organoid models. 112 Consequently, full-thickness organoid equivalents of keloid skin cultivated in vitro are significant models for future research on keloids. Once these equivalents are confirmed to exhibit biological characteristics akin to those of actual keloids, They will signify a substantial progression in understanding the molecular mechanisms underlying this disease and developing therapeutic strategies.
Future directions
A review of relevant studies indicates that immune imbalance plays a crucial role in the occurrence, development, and recurrence of keloids. Alterations in immune cell populations and the cytokines they secrete can significantly influence fibroblast proliferation and collagen deposition. However, existing research is neither comprehensive nor specific. For example, most studies have primarily focused on analyzing the levels and/or functions of immune cells and their secretory factors in keloids, while rarely exploring the underlying molecular mechanisms involved. Some proposed mechanisms are extrapolated from findings related to other diseases but remain unverified within the context of keloids. This gap is closely linked to the absence of an ideal keloid model. Moreover, although various immunotherapeutic approaches for treating keloids exist, significant deficiencies persist. For instance, the cellular-level anti-inflammatory mechanisms associated with conventional treatments such as steroids, silicone injections, and botulinum toxin remain poorly understood. Many therapeutic strategies targeting TGF-β or other immune factors and pathways are largely speculative; they are based on their roles in conditions like tumors, pulmonary fibrosis, or atopic dermatitis but lack validation through either in vivo or in vitro models specifically designed for keloids. Furthermore, there is a notable absence of supporting clinical research data. Consequently, developing tissue culture models and humanized mouse models that accurately reflect the biological characteristics of keloids is essential for advancing our comprehensive understanding of their immune molecular mechanisms as well as formulating effective immunotherapies.
In recent years, significant advancements have been made in immunotherapies for keloids, however, numerous practical challenges persist. Firstly, there is a notable absence of effective treatment options approved by the Food and Drug Administration for keloids, particularly those pertaining to immunotherapy. 45 Secondly, the cost of monoclonal antibodies tends to be prohibitively high, and their therapeutic efficacy is significantly affected by environmental factors such as temperature and pH levels. Furthermore, there remains a scarcity of large-scale clinical trial data at present, resulting in considerable gaps within the field of keloid immunotherapy. Finally, while various studies indicate that combination therapies for keloids yield better outcomes than monotherapies—such as when steroid injections are administered within the lesion alongside surgical resection—this approach has demonstrated a markedly lower recurrence rate compared to surgery alone. 113 However, the integration of skin immunotherapy with surgical intervention, radiotherapy, and the combined application of multiple skin immunotherapies remains devoid of comprehensive theoretical guidance. This represents an area where researchers and clinician must focus their efforts moving forward.
CONCLUSION
Although it was widely believed that abnormally proliferating fibroblasts played a dominant role in keloid formation, subsequent studies have demonstrated that immune imbalance also plays a significant role in the initiation and progression of keloids. The analysis of aberrantly activated immune cells and the cytokines They secrete is instrumental in the diagnosis and treatment of keloids.
Immune imbalance factors are anticipated to function as diagnostic biomarkers. sHLA-E has emerged as a predictive biomarker for keloids progression and recurrence, while TRM may play a significant role in postoperative relapse. The upregulation of Th2 subsets (IL-4 and IL-13) suggests a heightened likelihood of keloid formation and associated pruritus. Furthermore, the increased presence of CD4+CD25+FOXP3+ Tregs in tissues is closely associated with the progression of multiple keloids.
Immune-targeted strategies may inhibit the development of keloids at an upstream stage, representing a promising breakthrough for both treatment and prevention of recurrence. Dupilumab (IL-4/IL-13 blockade) demonstrates clinical efficacy in reducing keloid burden and pruritus. Experimental approaches targeting TGF-β signaling (Fresolizumab, AVID200), NKG2A/CD94 checkpoints (Monalizumab), IL-6 (Tocilizumab), and TSLP (Tezepelumab) hold promise for precision immunotherapy. Furthermore, localized treatments that are based on the characteristics of immune dysregulation within keloids, as opposed to systemic immunotherapy, may offer new insights into enhancing treatment outcomes, minimizing adverse reactions, and fostering personalized treatment strategies. The hypothesis that combined therapy—particularly combined immunotherapy—is more effective than monotherapy necessitates further research and validation. Effective keloid models and large-scale clinical trials could present significant opportunities for the clinical transformation of immune-targeted therapies. Research into immune imbalance provides unprecedented avenues for targeted treatment and recurrence prevention in keloids.
TAKE-HOME MESSAGES
Due to the absence of standardized and effective treatment interventions, keloids continue to pose a significant clinical challenge, with recurrence rates reaching as high as 100% when conventional treatment methods are employed. Effective diagnostic markers and treatment strategies are grounded in a comprehensive understanding of the molecular mechanisms underlying keloids. Immune dysregulation serves as a key mechanism that facilitates the development of keloids. Regulating imbalanced immune cells and their cytokines may inhibit the development of keloids in the early stages, representing a promising breakthrough in both treatment and recurrence prevention. In addition, imbalanced immune molecules may serve as potential biomarkers for keloids. Combined and localized immune-targeted therapy may offer a novel strategy for the future management of keloids. Establishing tissue culture models and humanized mouse models that accurately reflect the biological characteristics of keloids is essential for exploring immune mechanisms and developing effective immunotherapy strategies.
AUTHORS’ CONTRIBUTIONS
Q.C.: Wrote the article. S.W. and M.Z.: Contributed to the conception of the study. Y.D.: Data curation. J.Y.: Contributed significantly to the article preparation. Y.D.: Helped perform the analysis with constructive discussions. All the authors contributed to the article and approved the submitted version.
Footnotes
ACKNOWLEDGMENTS AND FUNDING SOURCES
This work was supported by the Key projects at the school level of Wannan Medical College (WK2023ZZD27), Youth Innovation Research Fund of Jiangning Hospital, Nanjing Medical University–Youth Project (JNYYZXKY202502), A Collaborative Open Project between the Jiangsu Provincial Key Laboratory of Traditional Chinese Medicine Efficacy and Safety Evaluation and Jiangsu Zhengyang Pharmaceutical Co., Ltd. (JKLPSE201901).
AUTHOR DISCLOSURE AND GHOSTWRITING
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.