
Abstract
Significance:
Wound healing is as much about opportunity (Gk. Karios) and sequence (Gk. Chronos). The sequential healing responses involve concerted responses of cells of various lineages along with both soluble (cytokines and growth factors) and insoluble (matrix composition and topology) that are precisely coordinated for ultimate healing. This motivated the current scoping review to examine the biological relevance and utility of current in vitro models of wound healing, focusing on specific cellular responses.
Recent Advances:
We examined PubMed, Web of Science, and EBSCO of Science for the years 2020–2025, using keywords in vitro wound healing and individual cell type, namely epithelial cells, keratinocytes, fibroblasts, macrophages, platelets, and endothelial cells. This identified 126 relevant studies were selected and categorized by healing stage, cell types, and prototypical biological responses such as proliferation, migration, and lineage-specific functions.
Critical Issues:
This analysis outlined the utility of a temporal framework to organize current in vitro models as acute—initial, early and late, and chronic—delayed. Existing two-dimensional and three-dimensional models focus on platelet function (hemostasis), neutrophil or macrophage chemotaxis and phagocytosis (inflammation), epithelium (closure), fibroblast (matrix synthesis, contraction), and endothelial (angiogenesis). While these models provide key insights into specific phases, cell functions, and molecular mechanisms, they do not fully replicate the integrated, multiorgan processes of wound healing observed in vivo.
Future Directions:
Sequential combinations of these models lend themselves to the advances in artificial intelligence, machine learning, and robotic automation that are integral to developing prognostic and therapeutic wound care agents.
Praveen Arany

SCOPE AND SIGNIFICANCE
Among the three prototypical biological processes, namely, embryogenesis, carcinogenesis, and wound healing, there is a striking dichotomy of phasic changes in healing tissues. This includes features of organized, profuse proliferation and migration in early wounds simulating tumors, compared with healing during later stages which are organized and undergo remodeling and reconstitution, simulating development. These disparate but coordinated responses offer significant insights into biological regulations that could be harnessed to improve therapeutic clinical outcomes.1–3 Wound healing has emerged as a key area of biomedical research due to its direct relevance across various chronic diseases that have a profound impact on clinical outcomes. With increasing global interest, many studies have explored discrete aspects of wound healing, focusing on individual phases and cell types, leading to a broad range of experimental models now available.4–6 These models represent isolated mechanistic aspects of wound healing, such as keratinocyte migration or fibroblast-mediated matrix contraction, rather than complete wound closure or systemic repair.
The temporal framework enables researchers to better address contributions specific to in vitro cell responses and ultimately to in vivo scenarios. For example, macrophage polarization defects in the late stage have been strongly linked to chronic wound development. 7 However, multiple macrophage transgenic knockout mice have demonstrated both delayed and no effects on wound healing.8,9 These observations indicate specific macrophage responses, such as proliferation and migration, or functional roles, such as phagocytosis, are differentially modulated and must be assessed in the appropriate context. Similarly, studies with recombinant transforming growth factor (TGF)-β1 showed prominent connective (granulation) tissue induction. In contrast, transgenic animal models with TGF-β and Smad3 knockouts demonstrated improved wound healing in terms of epithelial closure. 10 In the latter, Smad3 knockout matrix-supported (skin wound) versus nonsupported (ear punch) showed strikingly opposite phenotypes attributed to fibroblast roles in matrix composition and mechanotransduction.11,12 Thus, this review outlines the value of examining specific cell responses in a modular manner and, where appropriate, employing multiplexed models to simulate more complex or interdependent cellular interactions for precisely designed, rationalized interventions. While wound healing shares conceptual parallels with embryogenesis and tumor biology, this review specifically interprets these biological principles within the framework of adult cutaneous wound repair. In skin, healing is characterized by keratinocyte-driven re-epithelialization, barrier restoration, and dermal matrix remodeling, and it proceeds predominantly through repair mechanisms rather than complete tissue regeneration.
TRANSLATIONAL RELEVANCE
Beyond its organizational value, this review aims to serve as a practical reference for researchers designing experimental approaches in wound healing. Whether applying simple monoculture systems or complex coculture models involving multiple cell types, researchers can leverage the framework for both fundamental discoveries and clinical translational goals. 13 The modularity of these systems fosters the fundamental tenets of necessity (gain or loss of function—partial or exaggerated phenotypes) with sufficiency (loss of phenotype). For example, a novel drug candidate might be assessed in a keratinocyte migration assay during the epithelialization phase, followed by fibroblast matrix assay to further evaluate long-term wound remodeling. These in vitro wound models would enable integration into robotic automation and artificial intelligence (AI) and machine learning-driven platforms for iterative development through rapid, high-throughput screening.4,14,15 A significant challenge in AI-driven biological discovery is the “leap of uncertainty” where the poorly labeled or context-less data result in uninterpretable or unreliable predictions. 16 Hence, offering well-defined, phased or time-staged, and functionally categorized in vitro wound models appears to be ideal testbeds for training and optimization of novel, precision wound therapeutics. Findings from these models must be validated in animal models and clinical studies before extrapolating to whole-wound healing outcomes.
CLINICAL SIGNIFICANCE
Wounds represent a major public health concern. A 2018 study estimated that approximately 8.2 million Medicare beneficiaries experienced at least one wound-related condition, and the total health care spending on all wound types ranges from 28.1 to 96.8 billion dollars. 17 These clinical and economic burdens emphasize the urgent need for better strategies to understand, prevent, and treat wounds. Although most wounds manifest in similar ways, their underlying etiologies, such as vascular insufficiencies, metabolic disorders, infections, or traumas, differ significantly, and each implies distinct cellular responses.18,19 Even within a typical wound-healing timeline, different cell types dominate during different stages, performing unique functions including hemostasis, inflammation, proliferation, and remodeling. 20 Currently, much of the clinical wound management relies on a broad empirical framework. While useful, this approach does not always reflect the temporal complexity of wound biology. For example, a moist environment is helpful in early healing stages, but excessive moisture can lead to maceration and is counterproductive or even harmful in latter healing phases. 21 A temporal, lineage-focused approach helps map specific cell behavior to healing contributions and can be used to align experimental models with real-world clinical challenges. By highlighting when each cell type is most active and when dysfunction is most likely to occur, this framework allows clinicians and researchers to better identify therapeutic opportunities. For example, a wound requiring primary closure would focus on epithelial migration and proliferation, while wound healing by secondary intention would focus on angiogenesis, granulation tissue, and extracellular matrix formation. Similarly, a stalled chronic could focus on modulating macrophage polarization or myofibroblast activation.
BACKGROUND
Wound healing represents an intricate and dynamic pathophysiological process that is integral in maintaining human health and wellness. Various injury agents can disrupt anatomical tissue integrity and functionality, causing wounds to develop. 22 Although the precise duration to categorize an acute versus chronic wound remains debatable, a general consensus is that a chronic wound persists over 30 days. 14 An acute wound occurs after a recent injury, regardless of its cause, and is expected to heal naturally by progressing through the normal phases of wound healing. In contrast, a chronic wound fails to follow this orderly process within the anticipated timeframe, hindering the restoration of anatomical and functional integrity.23–25 The cascading sequence of cellular and molecular events in the injured region facilitates a multistage process of healing that is broadly categorized into four phases: hemostasis, inflammation, proliferation, and remodeling. 4 Both soluble and insoluble compositional elements contribute to this process. Soluble factors include blood constituents, such as complements, plasma, and clotting factors, as well as tissue fluid. The insoluble factors include two major categories, namely, the matrix and cells of various lineages. This review focuses solely on the major cell constituents of the healing response and current in vitro model systems to examine their contributions. Although many in vitro wound models are broadly applicable across tissue types, this review focuses specifically on their application to cutaneous wound healing rather than true tissue regeneration that includes appendages and ancillary structures. Skin repair is characterized by keratinocyte-driven re-epithelialization and barrier restoration and proceeds predominantly through repair.
METHODS
We conducted a structured literature search across PubMed (n = 238) and EBSCO (n = 150) databases for the years 2020–2025 (Supplementary Fig. S1). Boolean search strings combined in vitro wound healing or wound model with individual cell types (epithelial cells, keratinocytes, fibroblasts, macrophages, platelets, and endothelial cells) and functional responses (proliferation, migration, collagen synthesis, tubulogenesis, phagocytosis, and clot formation). Additional searches were performed for eosinophils, basophils, mast cells, lymphocytes, neutrophil extracellular traps (NETs), and polymorphonuclear neutrophils (PMNs), though these yielded few eligible studies. The combined search identified 388 records, of which 26 duplicates were removed, leaving 362 for screening. After title and abstract review (n = 222), 126 studies were included in the final analysis. Excluded studies were primarily due to the absence of a wound healing assay or model (39.5%), inaccessibility of full text (20.9%), inappropriate cell type (24.4%), lack of three-dimensional (3D) or organotypic culture systems (5.7%), or other reasons (10.5%). Results were organized by individual cell types and their temporal functional roles (Supplementary Figures S2, S3, S4, and S5). Notably, only 16 studies specifically employed 3D or organotypic culture systems, revealing a gap in physiologically relevant in vitro wound models (Supplementary Fig. S6). We noted several models of other tissue wounds, such as muscles, nerves, and other tissues/organ systems that offer good insight into anatomical differences in the healing process, but these were excluded from the current analysis.
RESULTS AND DISCUSSION
Etiological and microenvironmental determinants of wound injury agents
Wound healing is taught in the textbook of pathology, not physiology. The fundamental premise is that there needs to be an injurious, pathological agent to evoke physiological wound responses. These factors can be broadly categorized as damage or biological, or more specifically microbiological agents. The former group of agents are collectively termed.
Damage-associated molecular patterns
These involve biophysical agents, such as physical injuries from erosion, contusions, incisions or excisions, ionizing radiation, thermal, or cryoinjuries26–28 (Fig. 1). They also include biomechanical injuries, such as pressure or shear stress, as well as biochemical agents, such as hypoxia, pH, hyperglycemia, and chemotherapeutic agents. The second category of injury agents is collectively termed.
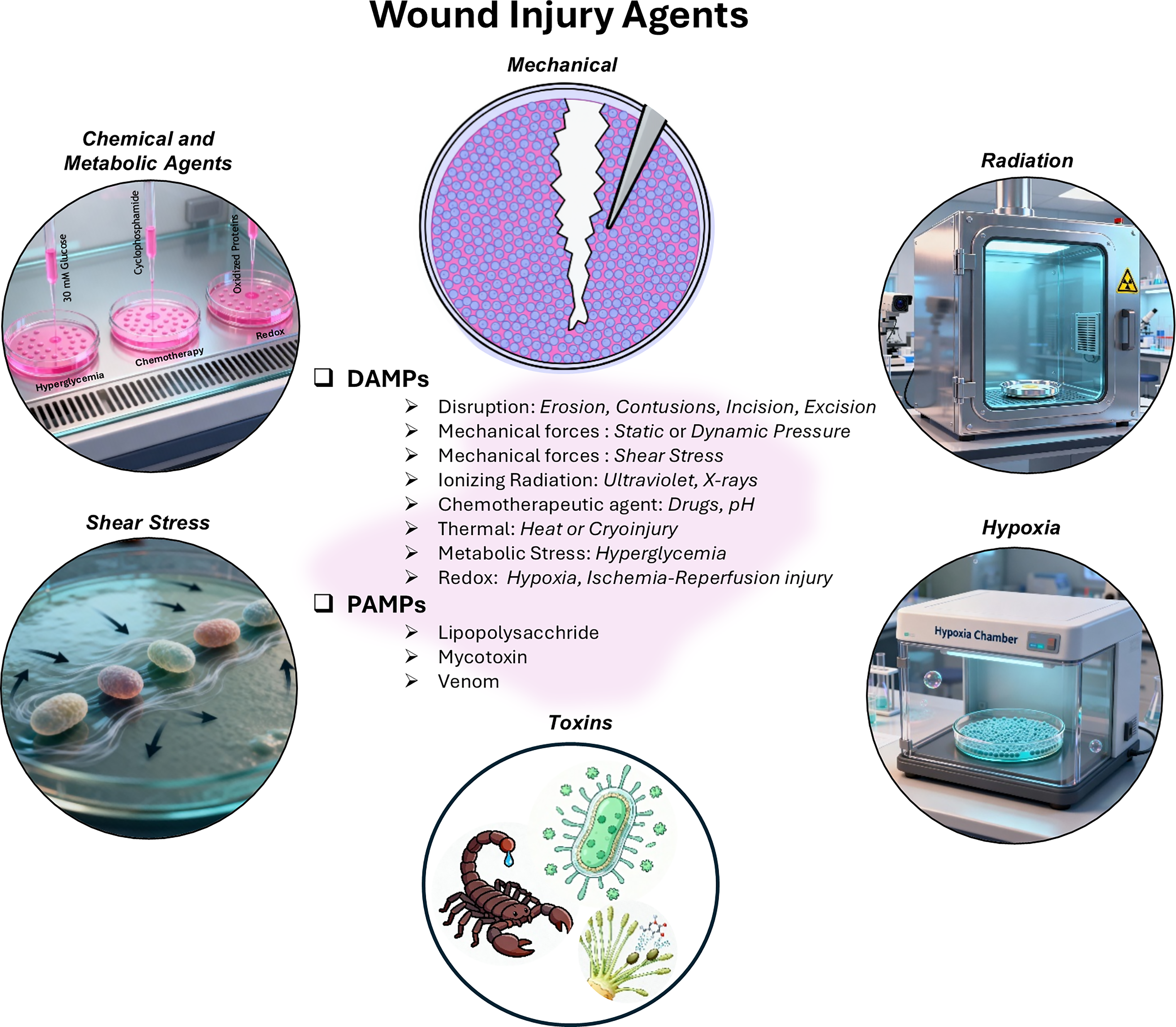
Wound injury agents driving acute and chronic wound healing responses: injury stimuli, cellular phases, and associated in vitro assays. Summary of wound-healing triggers grouped as surgical, DAMPs, and PAMPs, with corresponding primary cell types and their major functional roles such as aggregation, activation, migration, proliferation, and matrix synthesis across the initial, acute, subacute, and chronic phases of repair. DAMP, damage-associated molecular pattern; PAMP, pathogen-associated molecular pattern.
Pathogen-associated molecular patterns
This includes microbial biofilms harboring bacterial or fungal colonies that can be simulated in vitro through lipopolysaccharides (LPSs) and mycotoxins. Venom from scorpions and poisonous snakes, such as the Bothrops species, is known to generate significant wounds in vivo and has therefore been employed to simulate in vitro injuries.29–32
It is prudent to emphasize that the mere presence of these injurious agents lends themselves as pathophysiological in vitro wound models, compared with the simpler, physiological functional assays they represent. Acute wounds demonstrate well-regulated oxygen tension, leading to a mildly hypoxic environment, which, in turn, promotes tissue regeneration through controlled angiogenesis and extracellular matrix (ECM) remodeling. 33 In contrast, chronic wounds develop persistent hypoxia that disrupts these reparative processes. 33 pH levels further differentiate wound types: acute wounds maintain near-neutral pH (mean 7.4), while chronic wounds show alkaline shifts ranging from 7.4 to 8.9.34,35 Both wound types exhibit upregulated glycolysis to meet energy demands, although this metabolic adaptation proves insufficient for restoring homeostasis in chronic conditions. 36 Neutrophils exposed to hypoxia and acidic conditions demonstrate altered NET formation, reduced phagocytic efficiency, and delayed clearance, which leads to persistent inflammation evident in chronic wounds.37,38 Macrophages under similar conditions show skewed polarization and excessive proinflammatory cytokine production, impairing their transition from M1 to M2 phenotypes and stalling tissue repair. 7 Keratinocytes are sensitive to early environmental shifts, such as hypoxia and LPS, that inhibit their proliferation and migration, two key aspects of re-epithelialization. 39 Thus, these agents provide critical insight into the interplay between early microenvironmental changes and cellular dysfunction driving healing response.
Karichronos: temporal framework of lineage-specified wound models
The progression of the healing response provides a kinetic timeline to logically categorize the wound models as either acute (initial, early, and late) or chronic (delayed) (Fig. 2A). Each of these phases has specific cell types that drive the primary tissue healing response, while simultaneously involving other cells with supportive roles (Fig. 2B). The crosstalk and interplay among these cell types, their soluble cytokines and growth factors, as well as their insoluble ECM have important roles in determining the integrated healing response. In the following sections, each cell type and its associated functions are outlined to elaborate on their contributions to tissue healing. The Greek term Karios refers to qualitative opportune time, indicating that wounds, despite their initiating etiopathology, would heal uneventfully in the right environment. This shows that cell responses in the acute models of wound healing, namely, initial, early, and late, could be investigated to further understand and intervene in a rationalized, phase-specific, and timed manner for optimal clinical therapeutic responses. The Greek term Chronos refers to quantitative sequential time, indicating that the persistent or exaggerated nature of the injurious agent or perturbed cell responses would stall the healing responses, thereby leading to chronic wounds. These pragmatic in vitro wound models provide an opportunity to develop directed therapeutics for the significant health issues chronic wounds represent in current health care. A major goal of this review is to identify existing current assay systems, their predictive validity, and gaps in the technical and knowledge base to generate new rationalized wound management strategies.
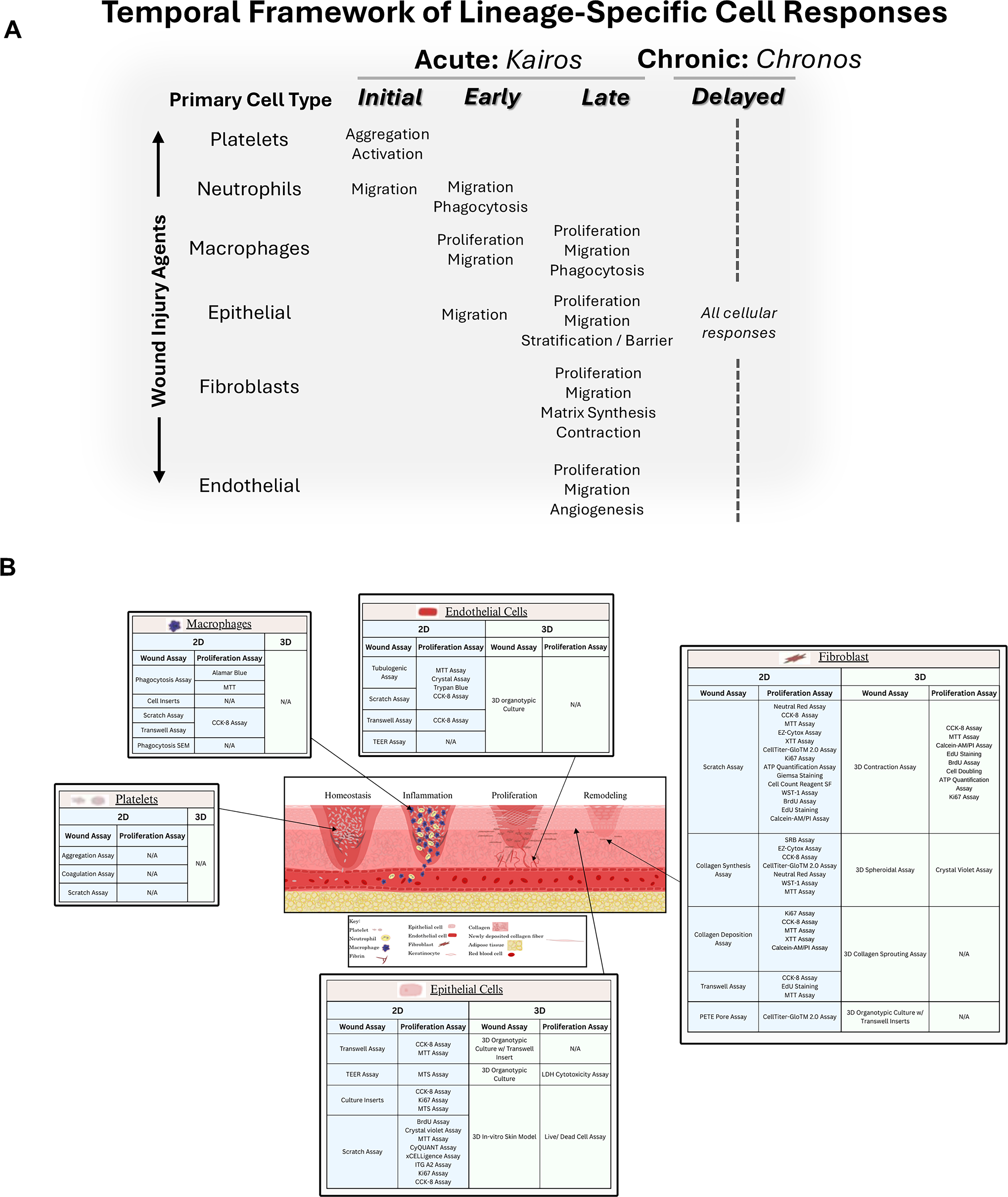
Temporal framework of lineage-specific cell responses in acute and chronic wound healing.
Initial in vitro model of acute wound healing: platelets
Hemostasis is the first stage of wound healing and refers to the local responses of tissues to stabilize the surrounding damaged microenvironment and ensure optimal recovery. 40 Tissue damage triggers immediate clot aggregation through platelet activation and fibrin deposition, creating a stable matrix that seals the wound and prevents further blood loss. Hemostasis is initiated rapidly, within minutes to hours, postinjury. Clotting efficiency can be impaired by the relevant underlying platelet disorders in both acute and chronic wounds. Both quantitative abnormalities, such as thrombocytopenia, and qualitative defects, such as Glanzmann thrombasthenia, storage pool defects, and Bernard–Soulier syndrome, compromise platelet adhesion, aggregation, and secretion, which ultimately leads to the different phases of wound healing presented here. 41 These defects further disrupt early matrix formation and impair the recruitment of inflammatory and regenerative cell populations needed for downstream repair.
Several in vitro models have been developed to study platelet function and aggregation under controlled conditions. These include light transmission aggregometry, impedance-based assays, and microfluidic platforms, which function to simulate physiological flow conditions. More advanced systems further integrate the 3D fibrin matrices or coculture with endothelial cells to better replicate the wound microenvironment and access platelet-mediated hemostasis in a relevant context. These models are increasingly being used to evaluate and optimize platelet-targeted therapeutics in patients with bleeding disorders or impaired wound healing through platelet-rich biomaterials, synthetic agonists, and engineered nanoparticles designed to restore hemostatic function. 37 Despite their advantages, limitations to in vitro models studying platelet function and aggregation include poor standardization, high levels of technical variability, and an oversimplified vessel geometry, which can cause incomplete recapitulation of the cellular complexity of in vivo wound homeostasis and repair.42,43
Early in vitro model of acute wound healing: inflammation
The inflammation phase is the second stage of wound healing that is initiated concurrently with hemostasis through vasodilation and increased vascular permeability. 44 The primary cell types operative in this phase are the PMNs, macrophages, and the epithelial keratinocytes. The latter two cell type responses appear to predominantly determine “wound closure” and could be considered the true bridging coordination between the initial tissue damage responses and ultimate healing and regeneration.
PMNs (or neutrophils)
A key driver of the inflammatory responses is the PMNs (or neutrophils). These are the primary cells recruited to the wound site and play a critical role in orchestrating the inflammatory response. In acute wounds, neutrophils are recruited in high numbers to clear tissue debris and pathogens through phagocytosis. 38 In addition to phagocytosis, neutrophils secrete potent antimicrobial proteins, including neutrophil elastase, gelatinase, cathepsin G, myeloperoxidase, lactoferrin, and histones, which have high affinities for DNA. They also form NETs consisting of extracellular fibrous structures of nucleic acids and associated proteins that enhance antimicrobial activity. The process of NETosis involves NADPH oxidase activation of PAD4 and histone citrullination. This leads to chromatin decondensation and addition of granule proteins such as myeloperoxidase and neutrophil elastase. This process is regulated by the lipoxygenase pathway and occurs in two forms, suicidal and vital NETosis. The former is triggered by toll-like receptors, Fc receptors, and complement receptors over several hours, ultimately resulting in neutrophil death through plasma membrane rupture and extracellular release. Vital NETosis occurs quickly, in minutes, after being triggered by bacterial LPS, TLR4-activated platelets, or complement proteins. NETs are released through nuclear blebbing and exocytosis to retain viable and functional neutrophils. In latter healing phases, the macrophages clean up released NETs through phagocytosis. Neutrophils not only act as first responders but also play a pivotal role in orchestrating the transition from the inflammatory phase to the proliferative phase of wound healing. 4
Studies on the neutrophil cells’ role in wound healing have been limited because of a lack of immortalized cell lines. Immortalized cell lines require primary cell isolation from donors, which is not only cumbersome but also introduces variability.37,45 Functional assays to study primary neutrophil include NET assays, phagocytosis assay, and oxidative burst assays. NET assay includes immunofluorescence detection of citrullinated histone H3 or Sytox Green staining of extracellular DNA,38,46,47 whereas phagocytosis assay includes the uptake of fluorescence beads or labeled bacteria, typically measured through microscopy or flow cytometry. 48 In acute wound scenarios, these assays show rapid pathogen clearance and controlled NET formation. 44 Neutrophils release reactive oxygen species (ROS) in the wound bed to eliminate pathogens. In acute wounds, ROS levels are balanced, promoting healing without causing harm. In contrast, chronic wounds exhibit excessive ROS production, resulting in oxidative stress, tissue damage, infection, and impaired healing.26,27 Therefore, in vitro models that replicate these dysfunctional PMN behaviors are critical for understanding stalled wound healing, as well as for developing therapeutic strategies that specifically target neutrophil activity. However, these neutrophil functional assays are limited by their reductionist nature, reliance on isolated readouts and artificial stimuli, and inability to replicate the dynamicity of the neutrophil complex behavior occurring in in vivo wound healing and multiorgan involvement in recruitment. 49
Several in vitro wound models have been developed to examine the redox aspect of healing in specific cell types. For example, keratinocytes and fibroblasts cultured under hypoxic conditions or exposed to hydrogen peroxide have been used to assess ROS-driven changes in proliferation, migration, and apoptosis.26,39 Endothelial cell tube formation assays under oxidative stress provide insight into how the ROS level regulates angiogenesis. 50 More advanced systems employ microfluidic platforms or 3D coculture models that allow controlled oxygen gradients and ROS exposure, enabling studies to investigate the influence of redox imbalances on multiple cell lineages simultaneously.51,52 These models collectively highlight how balanced ROS activity is beneficial for wound repair, whereas dysregulated ROS contributes to chronicity. These in vitro redox-focused wound models are limited by their reliance on nonphysiological ROS sources and simplified culturing conditions, which fail to capture the multicellular and heterogeneous oxidative environment of wound healing in vivo. 53
Macrophages
Inflammation and immune responses represent a critical wound healing milestone. Although they are frequently used interchangeably, it is important to distinguish between inflammation and immune responses in wound healing. Inflammation refers primarily to the immediate vascular and cellular reactions following injury, including vasodilation, permeability changes, and the rapid recruitment of innate immune cells such as neutrophils and mast cells. 54 In contrast, immune responses represent the broader, coordinated activity of leukocytes (macrophages, lymphocytes, and dendritic cells) that shape both innate and adaptive phases of repair. While these processes overlap temporally, inflammation establishes the early wound milieu, whereas immune responses drive resolution and facilitate transition into tissue regeneration. 44 All cells in the wound milieu have roles in mediating inflammation and immune responses, some doing so directly (effectors), while others through modulating or supporting roles. However, the major effector cell driving the transition from initial injury responses to resolution is the macrophage.
This cell type undergoes phenotypic changes that reflect this molecular transition from early proinflammatory (M1) to later anti-inflammatory (M2) states. 55 Macrophages display remarkable plasticity and exist along a continuum of activation states, rather than as two rigidly defined M1 and M2 populations. The classical M1 (classically activated) and M2 (alternatively activated) labels are best viewed as operational extremes of a spectrum, with many intermediate and context-dependent phenotypes emerging in response to distinct microenvironmental cues such as cytokines, pathogens, and tissue-derived signals. M1-like macrophages are typically associated with strong inflammatory and microbicidal functions, whereas M2-like macrophages are linked to immunoregulation, tissue remodeling, and fibrosis, but both extremes can become maladaptive when dysregulated in time or space. However, an overrepresentation of M2-like macrophages can impair microbial and necrotic debris clearance and promote excessive ECM deposition, contributing to fibrosis and aberrant remodeling in certain pathological settings. Thus, optimal healing requires a balanced and temporally regulated macrophage response rather than a simple M1 to M2 switch.56–59 Several other subtypes have been described attributed to specific stimuli and responses.60,61 In acute wounds, approximately 85% of macrophages are M1 during the early stages of inflammation; this proportion decreases to 15–20% within 5 days as M2 macrophages become predominant.62,63 This seamless transition facilitates wound bed preparation for healing, by clearing debris and bacteria, while simultaneously promoting tissue repair. During the early inflammation phases, there is a balanced release of proinflammatory and anti-inflammatory cytokines. However, after the M1-to-M2 transition, a switch to a predominantly anti-inflammatory state supports wound progression to the proliferative phase. This transition has been noted to be dysregulated in chronic wounds, where approximately 80% of macrophages remain in the M1 state. 7 Dysregulated macrophage polarization leads to sustained proinflammatory cytokine activity and reduced anti-inflammatory signaling. Elevated levels of tumor necrosis factor (TNF)-α, interleukin (IL)-1β, IL-17, and inducible Nitric Oxide Synthase are characteristic of chronic wounds, while activity of growth factors such as TGF-β, vascular endothelial growth factor (VEGF), IL-10, and Insulin Growth Factor-1 remains low.7,64–67 The persistent inflammatory state stalls healing progression by delaying both proliferation and remodeling.68,69
Macrophages have been extensively investigated as essential regulators of inflammation and resolution during wound healing. In two-dimensional (2D) systems, their proliferation and viability are quantified using the live/dead assays and crystal violet assays, which provide insights into survival under wound-like stressors. 70 Migration is modeled by scratch assays and droplet spreading assays, both of which quantify directional movement across a defined gap or radial expansion from a droplet site. 71 Phagocytotic function is examined through the uptake of fluorescent beads or labeled apoptotic cells, while polarization is induced by cytokines (e.g., interferon [IFN]-γ/LPS for M1 and IL-4/IL-13 for M2) and confirmed by Quantitative polymerase chase reaction or enzyme-linked immunosorbent assay (ELISA). 61 In 3D models, physiological relevance has been investigated by embedding macrophages in a hydrogel such as VitroGel, where chemoattractant gradients guide macrophage migration and invasion. Confocal z-stack imaging in these systems captures altered phagocytic clearance under hypoxia or oxidative stress, recapitulating chronic wound pathology. 26 Organotypic cocultures of macrophages with keratinocytes and fibroblasts have further revealed impaired M1 to M2 polarization in diabetic wound conditions, a hallmark of stalled healing. 72 Together, these platforms reflect how macrophage dysfunction contributes to chronic wound persistence. However, these models are limited by their use of oversimplified stimuli and culture conditions that do not full capture the complex tissue-level signaling networks that control macrophage behavior and ECM components in vivo during wound repair. 73
Other immune-inflammation cell types in wound healing
Beyond neutrophils and macrophages, lymphocytes are involved in regulating the later stages of inflammation and coordinating adaptive immune responses.33,34 T and B lymphocytes regulate adaptive feedback on fibroblast proliferation and keratinocyte migration through cytokines such as IFN-γ and IL-10. Helper T (Th) cells shape cutaneous wound healing through distinct functional subsets, each contributing to the balance between inflammation and repair. Th1 cells drive early pro-inflammatory responses through IFN-γ and TNF-α, promoting macrophage activation and microbial clearance but potentially exacerbating tissue damage if sustained. Th2 cells secrete IL-4, IL-5, and IL-13, supporting M2-like macrophage polarization, angiogenesis, and matrix deposition, which can facilitate repair but also predispose to fibrosis when dysregulated. Th17 cells produce IL-17 and IL-22, amplifying neutrophil recruitment and epithelial responses that aid barrier restoration may contribute to chronic inflammation and impaired healing when overactive. Regulatory T cells (TRegs) accumulate in skin during the late inflammatory and proliferative phases, where they suppress excessive Th1/Th17-driven inflammation, modulate Th2 type responses, and directly support re-epithelialization and stem cell-mediated regeneration, thereby promoting resolution and preventing pathological scarring. Overall, coordinated Th1, Th2, Th17, and Treg activity ensures timely progression through wound healing phases, whereas imbalance among these subsets is linked to delayed healing or fibrotic outcomes.74–77 Readily available lines including Jurkat (T-cells), natural killer (NK)-92 (NK-cells), and B-LCL (B cells) can be used to explore these effects. In addition, mast cells promote vascular permeability and release histamine and proteases that influence fibroblast and endothelial activity. 35 Mast cells (human mast cell [HMC]-1 and LAD2) release histamine, tryptase, and VEGF that modulate vascular permeability and angiogenesis. There have been few directed studies for the role of these cells in wound healing, which are largely in in vivo animal studies. Eosinophils (EoL-1 and HL-60) and basophils (KU812 and PBL-2H3) provide IL-4 and IL-13 to drive macrophage polarization and ECM remodeling. 36 Incorporating these cell types into 3D hydrogel or microfluidic coculture systems will more accurately reproduce the persistent inflammation seen in chronic wounds and enable evaluation of immunomodulatory therapeutics.13,15,25
A comprehensive approach addressing the contributions of diverse immune cell populations, including lymphocytes, mast cells, eosinophils, and basophils, is essential for a thorough mechanistic understanding of inflammation and tissue remodeling during wound healing. 37 Lymphocytes, such as T cells, serve as conductors of the cellular response, often attenuating dermal scarring, while NK cells exert a negative regulatory function by contributing to proinflammatory cytokine expression, including IFN-γ and TNF-α.37,38 Basophils are confirmed as key proresolution cells in murine skin wound healing, driving the shift of monocytes/macrophages (Mo/MΦ) toward a reparative M2-like phenotype through the action of IL-4 and colony-stimulating factor 1. 39 Furthermore, mast cells are identified as pivotal players in the early and proliferative phases, releasing mediators such as tryptase and growth factors that actively promote keratinocyte migration and wound contraction.40,41
To simulate the complex immunological environment and adaptive immune responses, incorporating established cell lines within coculture systems is recommended. 42 Specifically, the HMC-1 can be utilized to study the prohealing effects of mast cell secretions on keratinocytes, demonstrating a migratory effect that is tryptase-dependent. 43 Moreover, coculture of Jurkat (lymphocytes) and THP-1 (monocytes) cell lines offers a reliable in vitro platform for assessing immunosuppressive actions by quantifying the release of cytokines such as IL-2, IL-8, and TNF-α. 41 In addition to coculture models, integrating organotypic or microphysiological systems (MPSs) would enhance the physiological relevance of findings, as these systems effectively recapitulate complex tissue architecture and function, which is crucial when modeling skin diseases and aging-related wound healing pathologies. 44 Utilizing these diverse methodologies, including primary cells and established cell lines such as Jurkat and HMC-1, within both coculture and organotypic platforms will facilitate a deeper exploration of immune cell function in inflammation and remodeling.41,44 However, even advanced coculture, organotypic, and MPS-based skin models remain limited by their inability to fully encapsulate in vivo skin complexity, lacking components of vasculature, appendages, and fully integrated inflammatory responses. 78
Late in vitro model of acute wound healing: reconstitution
The resolution of the inflammation-immune phases transitions cells in the wound milieu to enter a tissue reconstitution phase. Depending on the extent and nature of the wound, it can involve multiple cell types, predominantly keratinocytes, fibroblasts, and endothelial cells. This phase involves dynamic cellular processes, including re-epithelization, angiogenesis, ECM synthesis, and wound contraction. 20 Thus, the concerted responses of these cells prime wound tissues structurally, metabolically, and mechanically for successful repair.4,79
Biological responses: proliferation, migration, and functional reconstitution
Cell proliferation is fundamental in the process of regeneration during the reconstitution phase. It ensures an adequate supply of cells required for barrier restoration, ECM production, and the establishment of new vascular networks that support the metabolic demands of the healing tissue.80,81 Proliferation in each of the cell lineages is phased to initiate and promote specific aspects of tissue reconstitution and remodeling. The prominent role of nutrition and supplements in mediating clinical wound healing responses could be directly correlated with cell proliferative responses. Cells not only divide but also migrate directionally toward the wound site in response to chemotactic gradients and ECM cues. 71 At the wound margins, cells undergo cytoskeletal reorganization and transiently loosen cell adhesions to migrate. 79 As migration proceeds, ongoing proliferation ensures a continuous supply of cells for barrier formation and tissue rebuilding. Proliferation-coupled migration is a key mechanism during the reconstitution phase of wound healing, enabling efficient repopulation and restoration of tissue structure. Disruptions in this coupling can compromise wound closure and result in chronic, nonhealing, or fibrotic wounds.
Independent of the proliferative response, migration alone is also a key aspect of the healing response for all three cell types. This may be investigated by examining the initial cell movement prior to cell doubling times (usually 24–36 h for epithelial and endothelial cells and 36–48 h for fibroblasts) or by inhibiting proliferative responses with small molecules such as Mitomycin C. Thus, investigating proliferation or migration alone as well as the coupled responses in each cell type can provide valuable insights into healing outcomes. Functional reconstitution of individual cell types includes epithelial stratification and maturation, leading to restoration of barrier functions. In fibroblasts, this involves ECM deposition, contraction, and reorganization to its preinjury, pristine architecture. In endothelial cells, the functional restoration of blood supply is driven by reorganization of the cell budding networks, tubulization, and restoring blood flow. These terminal functional responses in each cell type are key determinants of healing completion and degree of regenerative outcomes that can be examined with in vitro wound models.
Keratinocytes
Keratinocytes at the wound margins initiate the re-epithelialization by migrating across the wound bed and proliferating to further establish the epidermal barrier. 39 There have been several key observations on the pattern of these epithelial cell closure responses.82,83 Paul Martin and colleagues noted a “purse string” phenomenon, whereby actomyosin fibers at the wound edge generate circumferential tension to draw wound margins inwards. 84 A “tractor-tread or sliding” model has been proposed with basal keratinocytes migrating over the wound bed, while maintaining connections and pulling the epidermis forward. 85 Proliferating cells push suprabasal cells upward through mitotic pressure that reconstitutes the latter epithelial barrier. A “leap-frogging” or “rolling” theory outlines the role of distant (nonleading edge) suprabasal keratinocytes sliding over basal cells at the wound edge to convert to a basal phenotype upon reaching the wound bed. This enables effective re-epithelialization when wound margins are far apart. Usui et al. have proposed a combination of both theories, suggesting basal cells lead migration while suprabasal cells migrate and outnumber them, allowing rapid wound coverage. 86 Safferling et al. outlined a 3D model of concentric waves of keratinocyte production moving toward the wound center, with basal cells migrating collectively and suprabasal cells forming a protective shield, creating a triangular multilayered epithelium. 87 These discrete epithelial cell populations have not been carefully investigated individually in current wound models.
To assess cell proliferation in 2D assays, both direct approaches, such as crystal violet colony staining and Ki-67 immunolabeling, and indirect approaches, such as AlamarBlue, MTT, and XTT, have been investigated (Fig. 3). 88 Among the indirect methods, the XTT and MTT assays are terminal, while AlamarBlue (Resorufin-based) assays include multiple kinetic assessments of a given cell condition as well as 3D probing of cell health. Cell migration is assessed using “wound scratch” modeling, where cell sheet migration occurs across a linear wound (Fig. 4). Another format for cell migration is a cell outgrowth assay that captures radial migration from confined spots. 71 The major advantage of this latter assay is the ability to seed cells on various matrix components in order to assess cell migration behavior. This is not feasible with the scratch assay.
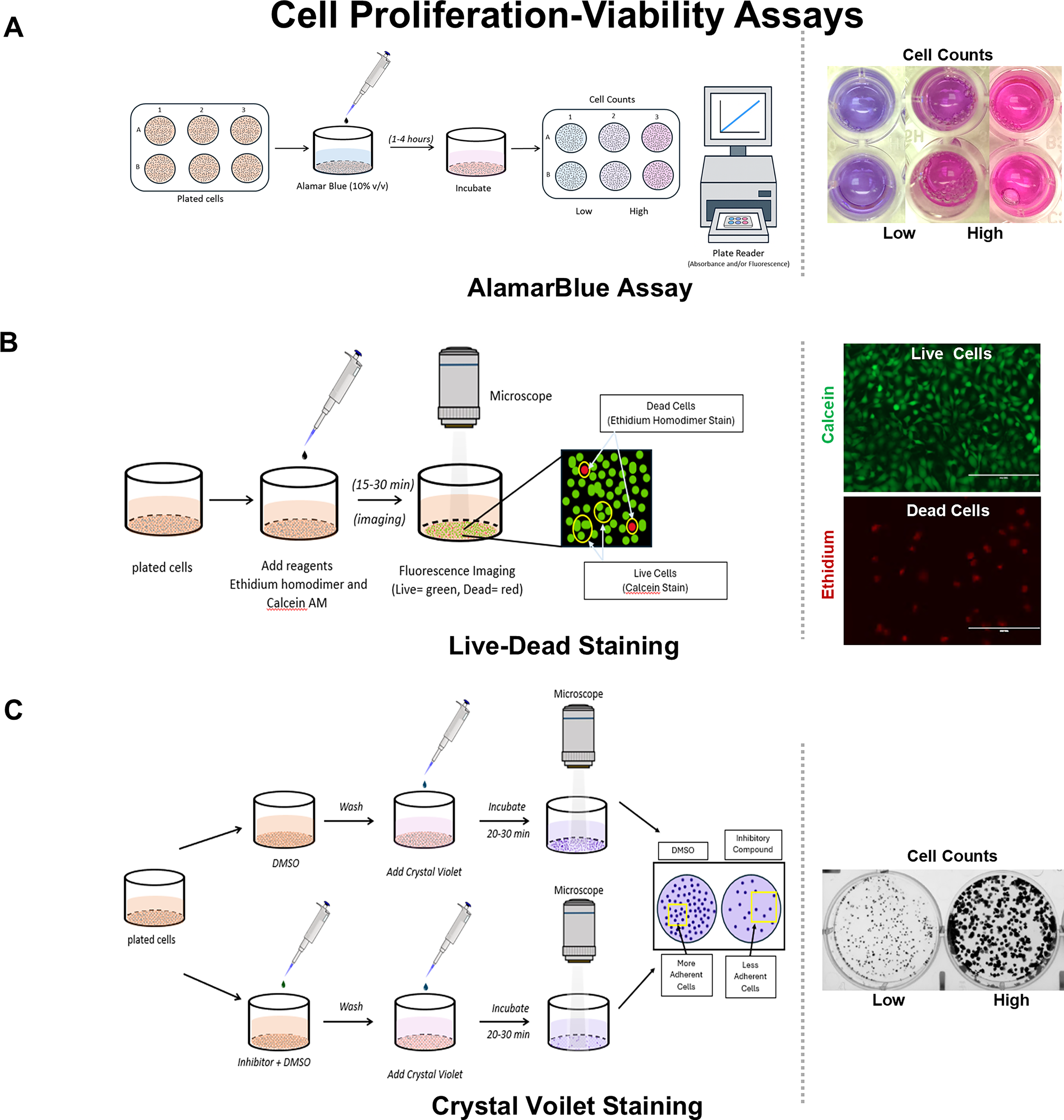
Cell proliferation and viability assays.
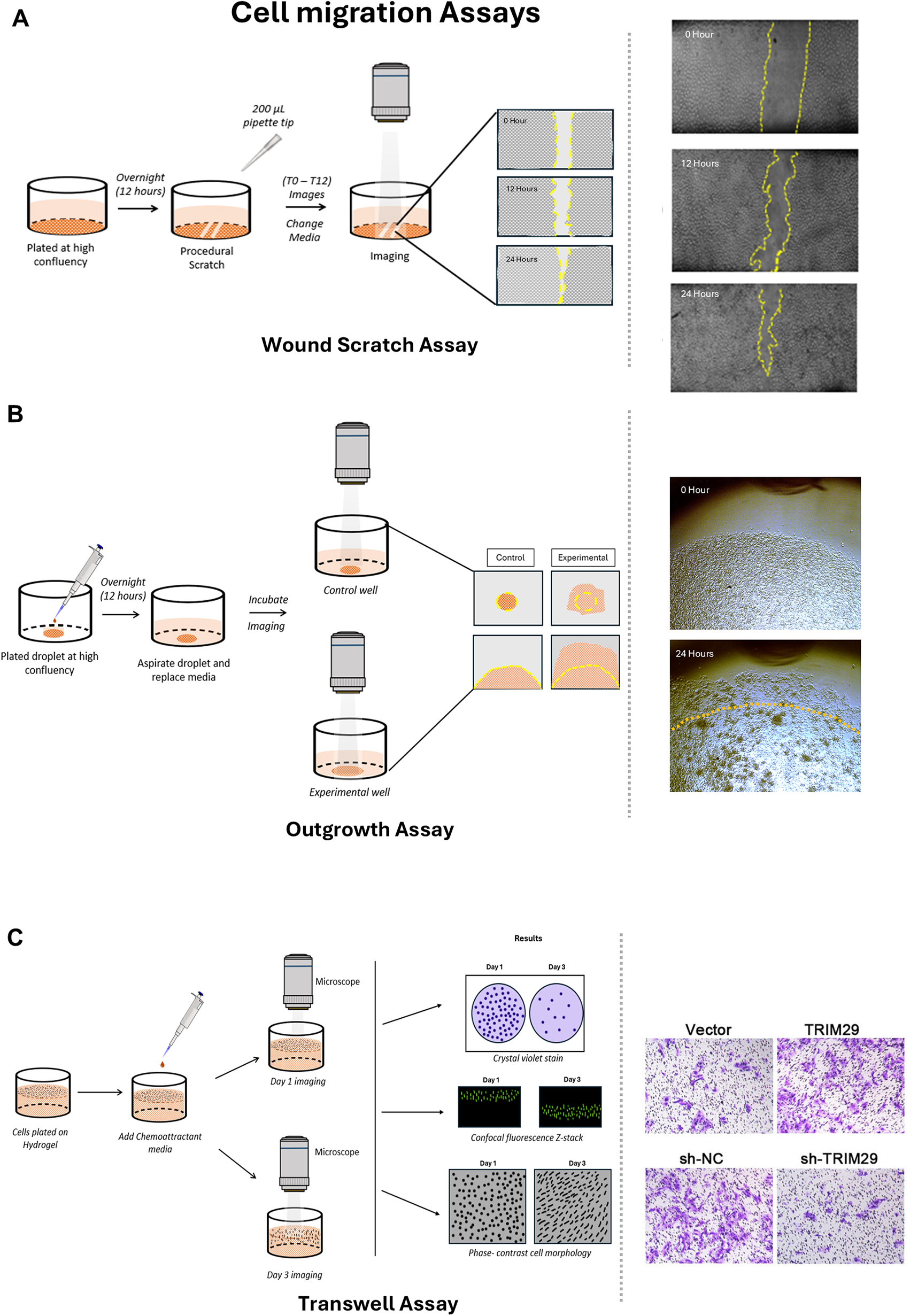
Cell migration assays in 2D and 3D model systems.
Keratinocyte 3D graft or stratification assays are used to model the formation of a multilayered epidermis in vitro (Fig. 5A). 89 Air–liquid interface (ALI) cultures promote stratification into basal, suprabasal, and cornified layers, enabling studies of differentiation and barrier repair. 89 Organotypic skin equivalents and coculturing keratinocytes with fibroblasts reproduce the dermal-to-epidermal junction, allowing analysis of integrin- and matrix metalloproteinase (MMP)-mediated migration. 90 Barrier function is evaluated using transepithelial electrical resistance (TEER) or dye penetration assays. 91 In these systems, keratinocytes are cultured on a dermal equivalent or scaffold at an ALI, which promotes their differentiation and stratification into basal, suprabasal, and cornified layers resembling native epidermis.89,90 These assays allow for the assessment of keratinocyte proliferation, differentiation, and barrier formation under conditions that mimic the in vivo environment. 90 They are commonly used to evaluate the effects of growth factors, cytokines, or biomaterials on epidermal regeneration and to study wound re-epithelialization in a physiologically relevant 3D context.14,39 These 3D keratinocyte graft and stratification models are constrained by their limited representation of adnexal structures, vasculature, immune components, and long-term remodeling. 92
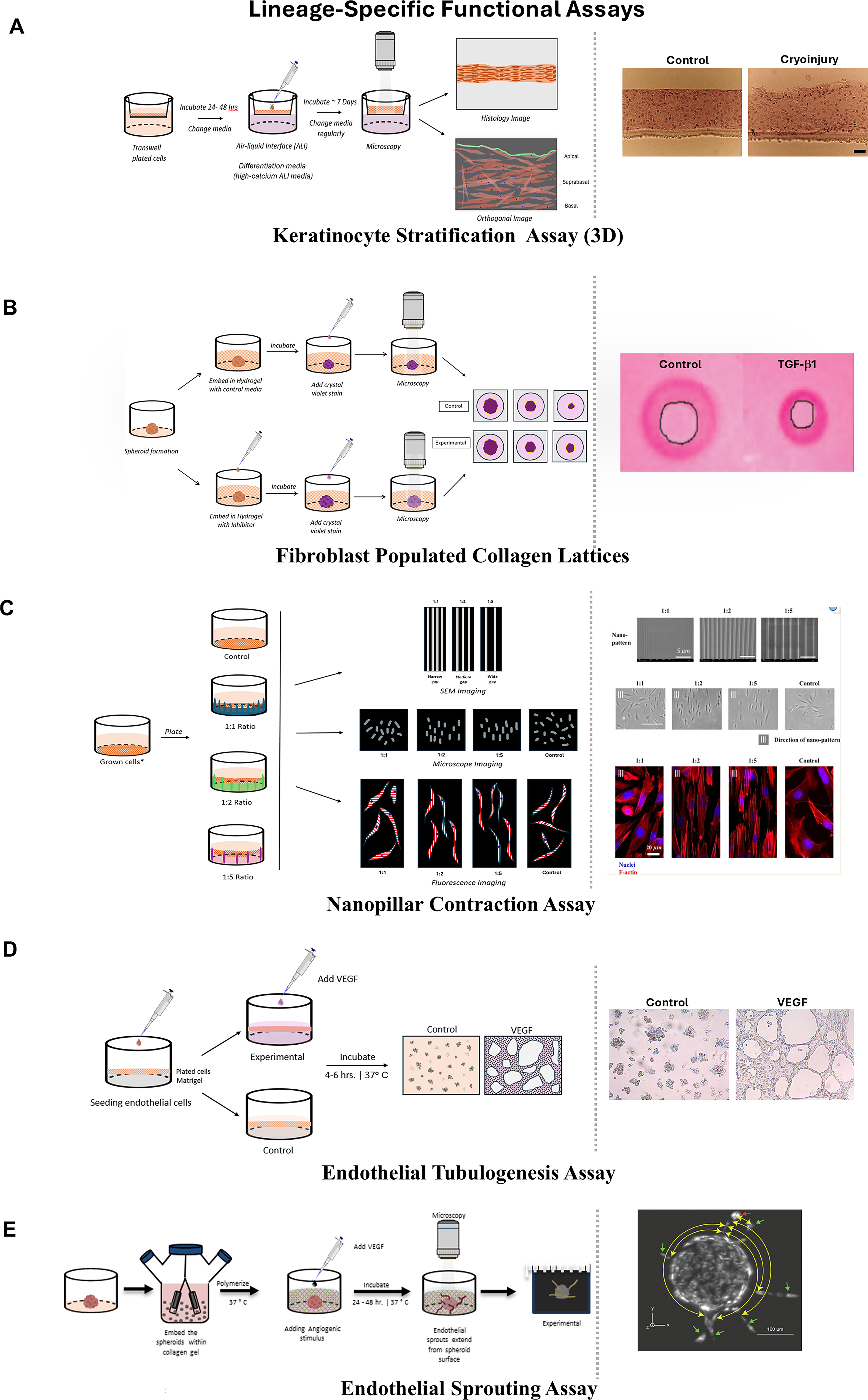
Lineage-specific functional assays for tissue reconstitution and remodeling.
Fibroblasts
Fibroblasts are recruited from adjacent and underlying connective tissues and mediate ECM synthesis, wound contraction, and remodeling. 39 Concurrently, fibroblasts secrete ECM structural proteins, such as collagen and fibronectin, and contribute to wound contraction by generating α-SMA positive, contractile phenotype (myofibroblasts) that is predominantly driven by platelet-derived growth factor (PDGF) and TGF-β signaling.11,93,94 The anatomical location of the skin fibroblasts appears to reflect discrete subpopulations. The papillary (upper dermis) exhibits more proliferative and migratory activity, generating type 3 collagen capable of epithelial–mesenchymal transition and active roles in early wound responses.95–98 The reticular (deep dermis) fibroblasts (CD36+) generate thicker collagen and more mature ECM that primarily contributes to the mechanical strength and pathological scar formation. There have been few efforts to examine these individual populations in directed investigations, presenting opportunity for innovation.
In 2D wound models, migration is measured with scratch and droplet assays, while fibroblast-populated collagen lattices (FPCLs) serve as contraction assays to quantify mechanical forces (Fig. 5B). 99 Collagen and fibronectin production are commonly assessed by staining or immunodetection. 100 3D systems provide higher fidelity by embedding fibroblast spheroids in hydrogels, where contraction over time reflects wound closure dynamics. 101 Nanopatterned substrates with ridge-to-gap geometries (1:1, 1:2, 1:5) test how matrix topography regulates fibroblast cytoskeletal alignment and mechanosensitive processes (Fig. 5C). 102 Chronic wound fibroblasts often exhibit senescence-like phenotypes, with impaired migration and persistent myofibroblast activity, which can be reproduced in advanced 3D scaffolds. 103
Fibroblast matrix synthesis and remodeling assays are designed to evaluate the ability of fibroblasts to deposit, organize, and remodel ECM components, such as collagen and fibronectin. 101 These assays may involve culturing fibroblasts in 3D collagen gels or on biomaterial scaffolds, allowing the cells to produce and align new ECM fibers over time. 99 Quantification can be performed by measuring total collagen deposition, visualizing fibril alignment using microscopy, or analyzing expression of key ECM proteins.5,100 Remodeling activity is often assessed by gel contraction assays, in which the extent of fibroblast-mediated gel compaction reflects their contractile and matrix organizing functions.93,94 One limitation of fibroblast matrix synthesis and remodeling assays is that they typically isolate fibroblast–ECM interactions in simplified 3D gels or scaffolds and therefore do not fully capture the influence of other wound-resident cell types, immune mediators, and mechanical cues that shape fibroblast behavior in vivo.104,105
Endothelial
Endothelial cells are central to angiogenesis, forming new capillaries from existing vessels in response to hypoxia-induced VEGF signaling. In larger wounds with appreciable tissue loss, adjacent fibroblasts and endothelial cells migrate into the wound to generate the fibrovascular tissues, termed granulation tissue. The comigration of endothelial cells forms solid cords of cells that undergo lumenization with the leaky lining, contributing to clear wound fluids apparent clinically. The early wound is mildly hypoxic, driving the angiogenesis process through VEGF signaling. 106 Proliferative activity is regulated by a network of growth factors, including epidermal growth factor, PDGF, and VEGF, while migration is promoted by VEGF, Fibroblast Growth Factor-2, and TGF-β.107,108 The maturation phase of these newly formed blood vessels includes migration and close association of smooth muscle cells. The newly formed vascular network ensures oxygen and nutrient delivery to the metabolically active wound bed, thereby sustaining keratinocyte and fibroblast activity and supporting granulation tissue formation. 106 Due to their richly vascular environment, granulation tissue is highly susceptible to infections, and it is recommended that all diseased tissues be removed. 109
Endothelial cells regulate angiogenesis, which restores perfusion to regenerating tissues. In 2D assays, tube formation on matrigel remains the gold standard for evaluating endothelial sprouting and capillary-like networks. 110 Scratch assays assess collective migration, while TEER assays measure barrier properties of confluent monolayers. 91 3D models extend physiological relevance utilizing endothelial cell-coated beads in collagen or matrigel hydrogels to assess new sprouting capillary-like vessels in response to VEGF gradients (Fig. 5D and E). 111 Microfluidic vasculature-on-chip platforms generate perfused vascular beds under flow conditions. 112 Organotypic cocultures with smooth muscle cells and fibroblasts stabilize vessel formation and mimic granulation tissue. 113 These advanced systems reproduce the fragmented or immature angiogenesis seen in chronic wounds, supporting evaluation of proangiogenic therapies.106,114 However, despite incorporating 3D matrices, flow, and coculture, they are currently only capable of including a restricted set of cell types and less complex microenvironmental cues. 115
Chronic in vitro model of wound healing: lack of healing
The wound-tumor to tumor-development dichotomy remains a prototypical feature of biological regulation.116,117 The prominent disorganization evident in early wounds involves profuse proliferation, migration, adhesion, and deadhesion of cells, angiogenesis, and matrix synthesis, along with mechanical changes that mimic tumor processes. A key intermediate step during the healing continuum involves the regulatory flip, where remodeling and reorganization to an original, development-like anatomical and functional state are accomplished. The lack or delay of this regulatory step is a key feature of chronic wounds that are stalled or undermined from progressing to later states. While the in vitro wound models from the acute stages with specific injury agent can be employed to investigate chronic wounds, a major difference is the frequency (persistence), amplitude, unique characteristic(s), or complexity of the injurious agent.118–121 For example, a burn wound is more complex to manage than a sterile, surgical wound that has prominent inflammation and preponderance to wound infection. A sulfur wound is more challenging to manage as its unique aspects of chemotoxic injury necessitate responses and repair of additive redox and genotoxic damage. Thus, simulating these discrete chronic in vitro wound models offers significant utility in investigating their etiopathogenesis and developing directed therapies.
The ideal outcome of wound healing is tissue regeneration, where the extent of remodeling and reorganization restores pristine anatomical form and function. The acute phase can span weeks to months depending on the type and extent of the wound (Fig. 6A and B). These pathophysiological phases are critically dependent on intrinsic cellular responses and suitable extrinsic microenvironment that provides the optimal opportunity (Karios) for tissue healing. Current clinical assessment deems any delay in epithelial closure beyond 3 weeks as chronic wounds. Dysfunction of individual cell responses or lack of progression to the next phase can protract the healing response and result in a chronic, nonhealing wound that persists for months to years. A few key characteristics of nonhealing wounds are the prominent, persistent inflammation. In acute wounds, the cells mediating inflammation are evident within the first 24 h of postinjury that typically resolve within 2 weeks.24,107 Conversely, chronic wounds exhibit a prolonged presence of neutrophils and macrophages that persist for extended periods.15,25 In acute wounds, timely PMN apoptosis and clearance further initiate recruitment of macrophages, activation of fibroblasts, and proliferation of keratinocytes, ensuring progression into the next stage of repair. 55 In contrast, neutrophils in chronic wounds often remain in a dysfunctional state, characterized by impaired migration, excessive or dysregulated NETs, ineffective phagocytosis, and persistent oxidative stress, all of which prolong inflammation and tissue damage, further stalling healing and driving chronicity.7,26,37,38,122,123 Moreover, excessive NET accumulation further damages tissues and persists during inflammation.
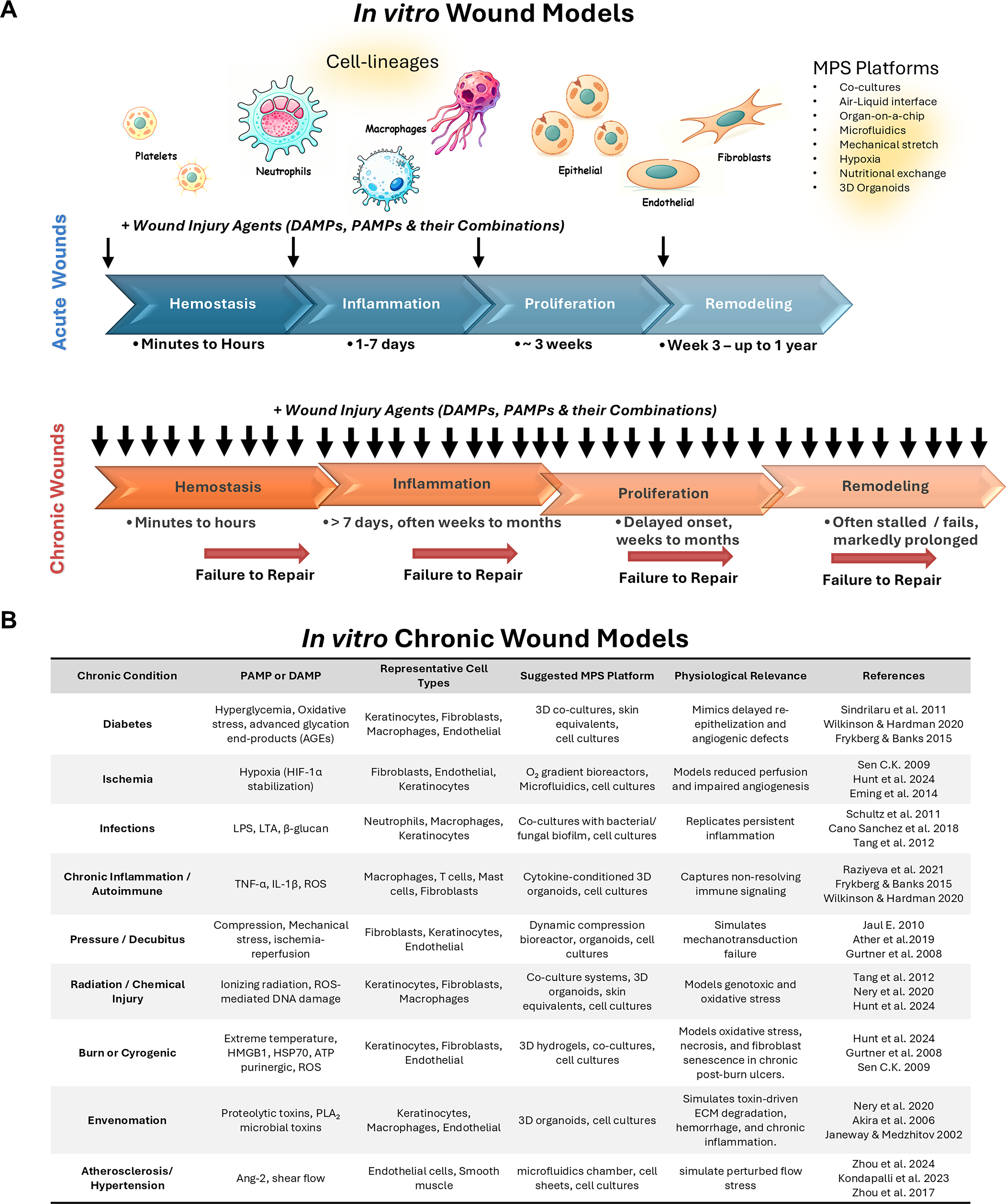
Temporal divergence of cellular processes in acute versus chronic wounds.
Acute wounds require targeted phagocytic activity that efficiently clears pathogens and cellular debris, while chronic wounds suffer from dysregulated immune responses that perpetuate biofilm formation and sustained inflammation.44,124 Prolonged inflammation and ECM disorganization generate a hostile microenvironment with disrupted chemotactic gradients, oxidative stress, and excessive protease activity—a pathognomonic biomarker of wound chronicity. 125 Cytokine profiles mirror this imbalance: acute wounds maintain low inflammatory cytokines with elevated growth factor concentrations, whereas chronic wounds show inverse patterns with heightened proinflammatory signaling and diminished growth factor availability.15,107 Deficiencies in proliferation or migratory capacity of cells, particularly in keratinocytes and fibroblasts, delay repair. 126 It is further characterized by the ECM maturation, collagen remodeling, and the vascular regression, which initiate and contribute to the formation of scar tissue. 4 In an acute wound, the type 3 collagen first deposits during proliferation and is then gradually replaced with type 1 collagen. This improves the tensile strength and structural integrity of the new forming tissue.127,128 However, in chronic wounds, this collagen transition is often disrupted, resulting in an abnormal ECM composition that compromises wound strength and function. 4 Myofibroblasts undergo apoptosis once their goal is achieved in contraction and remodeling, similar to the process in the proliferation phase, preventing excessive fibrosis. 11 However, chronic wounds rarely progress efficiently into this phase. The continued inflammation, cellular senescence, and the partially impaired ECM remodeling create a fibrotic environment. 129 This is marked by abnormal collagen cross-linking and poor tissue organization.130,131 This leads to a wound that remains open, is vulnerable to infection, and lacks any biomechanical integrity required for the full restoration.107,132 The early angiogenic responses formed during the proliferative phase are selectively pruned during this phase, leading to restore preinjury vascular architecture. 106
In chronic wounds, a sustained inflammatory environment interferes with keratinocyte migration and induces MMP overexpression, leading to excessive degradation of the ECM and a failure for full epithelization. 133 Chronic wounds show fragmented or immature vascularization, due to the disrupted VEGF gradients and the continued oxidative stress, thereby impairing tissue perfusion and stalling the repair process. 114 Excessive MMP activity, unresolved oxidative stress, and deficient growth signaling of TGF-β and IGF1 prevent structural maturation with smooth muscle coassociations.107,134 The balance between tissue inhibitors of metalloproteinases (TIMPs) and MMPs is critical for proper ECM turnover during wound healing. 135 TIMP–MMP assessment assays measure the expression, activity, and inhibitory interactions of these molecules. 135 MMP activity can be quantified using gelatin zymography, fluorogenic peptide substrates, or ELISAs, whereas TIMP levels are typically measured by ELISA or western blotting.136–138 These assays help determine whether matrix degradation and deposition are balanced, as excessive MMP activity can impair healing by degrading the new ECM and limiting necessary remodeling.139,140 Chronic wounds, however, exhibit dysregulated clot formation characterized by deficient fibrin matrices and failed stabilization of the wound bed.4,26,27,41 Collectively, the chronic wound models indicate that nonhealing wounds are not driven by novel cellular constituents but rather by failure of the same acute mechanisms to resolve in a timely and coordinated fashion. By prolonging exposure to injurious stimuli, altering environmental parameters such as hypoxia or pH, and incorporating multiple cell types in coculture, these in vitro systems recapitulate key features of chronicity, including persistent inflammation, aberrant matrix remodeling, delayed re-epithelialization, and impaired angiogenesis. Although individual assays such as scratch migration, FPCLs, redox-manipulated cultures, or organotypic skin equivalents interrogate discrete components of this process, they appear to constitute a modular toolkit for deconstructing complex wound phenotypes into tractable cellular responses. This modularity also enables alignment of specific model readouts with defined clinical bottlenecks, such as stalled epithelial closure, excessive fibrosis, or unstable neovascularization. Considered within a temporal framework, these models underscore that chronic wounds arise when regulatory checkpoints between phases are missed or overwhelmed, rather than from a single defective pathway. In this context, adaptations of acute in vitro platforms can be rationally combined to approximate chronic wound pathophysiology and to guide the development of phase- and lineage-targeted therapeutic strategies.
To move beyond descriptive pathophysiology, chronic in vitro wound models must incorporate simulation platforms that reproduce persistent hypoxia, hyperglycemia with advanced glycation end-products (AGEs), biofilm formation, and sustained inflammatory signaling as outlined in the “Validation and standardization gap” section. Controlled hypoxia systems, oscillating glucose gradient models, and biofilm-integrated microfluidic platforms provide experimentally tractable systems to test proangiogenic agents, macrophage reprogramming strategies, antioxidant therapies, MMP inhibitors, and antimicrobial interventions. Admittedly, these models still remain as simplified representations of in vivo chronic wounds and lack full immune, lymphatic, and appendageal complexity. They provide critical early-stage platforms to identify therapeutic targets, optimize dosing strategies, and screen candidate interventions prior to animal or clinical translation.
MPS: the platform for wound models
MPSs are sophisticated, in vitro platforms that mimic advanced, complex cell–cell and cell–matrix interactions in functional pathophysiological contexts. These advanced platforms represent the high-complexity end of the in vitro wound model spectrum and are essential for evaluating therapeutic efficacy in physiological relevant environments. These in vitro engineered platforms integrate living cells within biomimetic 3D architecture, often incorporating microfluidic flow, mechanical cues, and controlled biochemical gradients to recapitulate key structure and functional aspects of human tissue or organ for disease modeling and drug response assessment. 141 This includes multilineage cell cocultures, 3D coculture platforms such as organoids, organotypic “skin-on-chip” models, and ALI cultures, microfluidics systems, and a broad category termed physiological mimicry that includes mechanical stretch, pressure changes, and interorgan crosstalk. Coculture models are in vitro cell culture systems in which two or more distinct cell types are maintained together to examine intercellular interactions, including direct cell–cell contact and paracrine signaling, enabling controlled investigation of functional crosstalk between populations. 142 Organotypic models are in vitro culture systems that preserve key aspects of a tissue’s native 3D architecture, cellular composition, and cell–cell and cell–matrix interactions, enabling more physiologically relevant investigation of tissue-level structure and function than the conventional monolayer cultures. 143 These platforms recreate the complex biochemical and biomechanical gradients that enable more appropriate predictors in vivo responses. In wound healing, these systems enable real-time control of oxygen, nutrient, and cytokine microenvironments and are adaptable to both acute and chronic wound contexts. Only 16 of 126 studies included in this review noted utilization of 3D or organotypic designs, underscoring the limited physiological relevance of findings and the urgent need for broader adoption through funding and training initiatives. These MPSs can be modified for wound research by incorporating controlled hypoxia chambers (1–10% O2) to mimic ischemic conditions, oscillating glucose gradients (25–30 mM) with AGEs for diabetic wounds, and biofilm microchannels for infection simulation, integrating immune-stromal cocultures of keratinocytes, fibroblasts, macrophages, and endothelial cells. These platforms mimic selected aspects of the wound microenvironment (e.g., hypoxia and cytokine gradients) but cannot reproduce systemic immune responses, vascular perfusion, or neuroendocrine regulation.
While coculture, organotypic, and MPS models all aim to increase physiological relevance beyond traditional monolayered culture, they differ in biological complexity and functional scope. Coculture models primarily enable the study of intercellular communication between defined cell populations, focusing on cell–cell and paracrine interactions. Organotypic models extend this approach by preserving the native architecture and multicellular organization, allowing investigation of tissue-level structure and function. MPS further advances physiological fidelity by integrating dynamic mechanical biochemical and fluidic cues to recapitulate organ-level functions, making them suitable for disease modeling and drug response studies.
ALI models
ALI cultures represent a critical yet underutilized MPS approach for modeling epithelial wound healing. 144 ALI systems promote proper keratinocyte stratification, differentiation, and barrier formation that cannot be achieved in submerged cultures. Circular mechanical wounds applied to ALI-cultured primary airway epithelial cells enabled real-time assessment of wound closure rates under exposure to Wnt activators and TGF-β inhibitors, demonstrating modulation of re-epithelialization kinetics.144,145 For cutaneous wound applications, ALI models support mucus production, appropriate lipid lamellae formation, and establishment of functional tight junctions that represent barrier recovery. However, one limitation of ALI-based models of wound healing is that epithelial monolayers grown at an air-exposed interface typically lack key nonepithelial components, including cells of the stroma, immune cells, and vascular cells, and a naïve ECM. 144 Further, these models are currently limited in their ability to absorbency of biomaterial dressings, which could potentially be explored in the future with reverse TEER and wet weight assessments.
Skin-on-chip systems
Recent diabetic wound-on-a-chip (DWOC) model successfully replicated hallmark pathologies, including impaired ECM remodeling, disrupted dermal-vascular crosstalk, defective angiogenesis, and endothelial-to-mesenchymal transition under hyperglycemic conditions with AGEs.52,146 This platform demonstrated elevated proinflammatory markers (IL-1β, TNF-α, and MMP9) and reduced anti-inflammatory/angiogenic factors (IL-10 and VEGF-A), faithfully recreating the chronic inflammatory imbalance characteristic of nonhealing diabetic ulcers. One limitation of current skin-on-chip and DWOC systems is their restricted lifespan and incomplete incorporation of vascular and immune components, which limits their usability in long-term, chronic wound modeling. 147
Perfusion models
Perfusable skin-on-chip models represent the most physiologically relevant MPS platform for wound healing studies.148–150 These systems integrate vascularized dermal–epidermal constructs within microfluidic chambers that enable controlled perfusion, mimicking the dynamic fluid flow and nutrient delivery present in vivo. Vascularized blood vessel organoids cultured under continuous perfusion exhibited enhanced maturation, with transcriptomes most similar to in vivo transplanted organoids compared with static conditions. For wound healing applications, vascularized skin-on-chip systems enable investigation of angiogenic sprouting, endothelial–immune cell interactions, and real-time assessment of barrier function during re-epithelialization.52,148,151 Despite incorporating vascularized constructs and dynamic flow, they still face challenges such as limited lifespan, incomplete inclusion of all relevant skin and immune cell types, and technical/material constraints, such as medium absorption and oxygen differences. 147
3D coculture platforms
Multicellular 3D coculture constructs offer critical advantages over monoculture systems by recapitulating essential cellular crosstalk. 152 Keratinocyte–fibroblast cocultures embedded in collagen–fibrin matrices demonstrated accelerated migration rates, with keratinocytes reaching the wound center by day 3 compared with day 7 in monocultures. Critically, the presence of fibroblasts modulated keratinocyte proliferation dynamics and altered matrix mechanical properties, underscoring the importance of reciprocal cell–cell signaling that is absent in simplified models. 153 Macrophage–endothelial cocultures have revealed phenotype-specific regulation of angiogenesis during wound repair. Live imaging studies demonstrated that inflammatory (M1) macrophages associate with endothelial tip cells during early sprouting phases through TNF-α signaling, while anti-inflammatory (M2) macrophages dominate during anastomosis and vessel maturation. Macrophage depletion resulted in poorly directed filopodial sprouting and failure of anastomosis, highlighting that MPSs incorporating immune cells are essential for modeling the complete wound healing cascade. Live imaging studies demonstrated that inflammatory (M1) macrophages associate with endothelial tip cells during early sprouting phases through TNF-α signaling, while anti-inflammatory (M2) macrophages dominate during anastomosis and vessel maturation. Macrophage depletion resulted in poorly directed filopodial sprouting and failure of anastomosis, highlighting that MPSs incorporating immune cells are essential for modeling the complete wound healing cascade. Although these 3D cocultures offer significant insights into heterogenous multilineage interactions, they are not fully able to replicate vascular, mechanical, and spatial interactions evident in vivo.
Putative and emerging disease-specific modification for chronic wound modeling
This section explicitly distinguishes between disease-specific modifications that have been demonstrated with in vitro or MPS platforms, including those that remain conceptual or emerging requiring experimental validation.
Controlled hypoxia systems
Chronic diabetic wounds exhibit tissue oxygen tensions around 37 mm Hg compared with ∼60 mm Hg in acute wounds. Dual-sensor luminescent foils capable of simultaneous oxygen and pH imaging have revealed that chronic wounds display marked hypoxia without correlating oxygen gradients, contrasting with the dynamic oxygen fluctuations observed during normal healing. 154 MPS implementations should incorporate programmable oxygen tension control within the 1–10% O2 range combined with hyperglycemia (25–30 mM glucose). Under these conditions, human skin fibroblasts and keratinocytes exhibit HIF-1α overexpression, impaired migration, and reduced angiogenic factor secretion—pathological responses that were reversed by oxygen supplementation from photosynthetic microalgae-laden scaffolds. The specificity of oxygen concentration ranges, cell-type–specific responses, and validation methods strengthens the technical rigor beyond generic hypoxia chambers.155–157
Oscillating glucose gradients
Glucose-responsive hydrogel systems incorporating biosensors (glucose oxidase, concanavalin A, and phenylboronic acid) enable real-time detection and dynamic adjustment of glucose concentrations. However, the critical pathological feature is not merely elevated glucose but the combination of hyperglycemia (30 mM), AGEs, and LPS that collectively activate the mtDNA-cGAS-STING inflammatory pathway.146,156,158 STING pathway activation under high glucose conditions drives sustained M1 macrophage polarization, elevated IL-1β and TNF-α secretion, and impaired wound closure. MPS models should therefore incorporate programmable multifactor diabetic stress conditions rather than glucose alone, with temporal modulation to simulate the fluctuating metabolic environment characteristic of poorly controlled diabetes. 156
Biofilm microchannels
Biofilm infections are present in more than 80% of chronic wounds yet only 6% of acute wounds. Multispecies biofilms of Staphylococcus aureus and Pseudomonas aeruginosa exhibit synergistic tolerance to antimicrobials and enhanced virulence through quorum sensing and genetic exchange.159–161 3D implantable biofilm-infected wound models have demonstrated that initial inoculation of 104 CFU can form biofilms exceeding 109 CFU/g tissue within 10 days, with gradients of PMN infiltration at wound edges and cellular inhibition in wound beds. MPS implementations should incorporate microfluidic perfusion channels that enable controlled bacterial seeding, biofilm maturation under flow conditions, and antimicrobial susceptibility testing within physiologically relevant time scales. The chronic inflammatory response to biofilm—characterized by elevated IL-6, IL-10, IL-17A, TNF-α, and MMPs—can only be accurately modeled when bacterial burden, host immune response, and ECM degradation are captured simultaneously.159–162 These models may offer significant utility to be adapted for biomaterial antimicrobial assessments.
Real-time biosensing integration
Static endpoint analyses cannot capture the temporal dynamics of inflammatory phase resolution, macrophage phenotype switching (M1 → M2), or the biphasic patterns of MMP expression that govern ECM remodeling. Real-time biosensing is not merely advantageous but essential for mechanistic understanding of phase transitions in wound healing.152,154,163 Luminescent dual-sensor foils utilizing time-domain dual-lifetime referencing for pH and luminescence-lifetime imaging for oxygen enable noninvasive, real-time 2D mapping with spatial resolution < 5 μm and temporal stability over 30 min. These sensors revealed centripetally increasing pH gradients (pH 6.5–8.0) on chronic wound surfaces that disrupted keratinocyte migration and proliferation. 154 Electrochemical biosensors with gold nanoparticle-modified electrodes achieve submicromolar detection limits for inflammatory cytokines (IL-6, IL-1β, and TNF-α) with > 95% repeatability over 60 days. Wound biosensors can monitor pH, temperature, moisture, glucose, lactate, uric acid, and trimethylamine in real-time, enabling early infection detection and personalized therapeutic interventions. High-throughput automated imaging systems with nucleus tracking and automated wound closure analysis algorithms enable cost-effective screening but require validation against manual quantification and standardization across different imaging platforms. Integration of such sensors within MPS platforms transforms them from endpoint assessment to dynamic monitoring systems capable of capturing healing kinetics.164,165
Modulus
It is also known that ECM stiffness influences fibroblast activation into myofibroblasts, which influences chronic wound healing processes. Insufficient stiffness impairs myofibroblast action and delays wound closure, while excessive stiffness locks myofibroblasts in a contractile, apoptosis-resistant state, retaining inflammation. 166 In vitro models using collagen gels of varying moduli reveal a “lockstep” contraction mechanism, in which isometric forces permit the ECM to remodel. 167 Another approach to represent varying moduli of the ECM is to grow the cells on culture substrates with different adhesive capacities, thereby increasing their mechanical challenge on contractile behavior. 168
CURRENT LIMITATIONS AND FUTURE DIRECTIONS
A critical gap in wound healing research is the absence of unified benchmarking framework to compare model performance across studies, compromising rigor and reproducibility. The Standards Coordinating Body for MPSs has identified critical gaps in terminology standardization, sensor calibration protocols, and quality control criteria specific to MPS. International Organization for Standardization and American Society for Testing and Materials standards for MPS terminology and materials are emerging but remain incomplete for wound-specific applications. The standardization and implementation of an optimal workflow of MPS holding much promise for reproducible benchmarks for community-wide validation studies establish interlaboratory reproducibility metrics (Supplementary Fig. S9).169,170 Pneumatically controlled microfluidic wound healing assays achieve wound area reproducibility with relative standard deviations of 2–4%, representing 9- or 10-fold improvement over manual scratch assays (19% RSD). However, this technical reproducibility does not address biological variability arising from primary cell donor differences, passage number effects, or matrix batch variations.144,171 Nutrient consumption modeling and medium exchange protocols must be rationally designed based on cell density, metabolic demands, and desired concentration ranges to maintain quasi-steady-state conditions. 172 Further, embedding microsensors for oxygen tension, pH, or cytokine release within these chips enables longitudinal assessment of healing kinetics that are difficult to capture in static cultures. By merging these features with automated imaging and standardized perfusion protocols, MPS can provide reproducible, physiologically relevant benchmarks for both acute and chronic wound-healing studies.5,11,14,145,159,169,173 Addressing these limitations requires integration of emerging technologies that improve standardization, scalability, and predictive validity. Artificial intelligence (AI), machine learning (ML), automated imaging, and high-content analytics offer practical tools to overcome these translational gaps.
The major utility of this temporal approach to utilizing in vitro wound models is to recognize that the opportunities (Karios) for therapeutic intervention that most effectively revert or prevent chronicity (Chronos). Moving beyond the cells themselves to individual locations or function-specific subpopulations offers an innovative future aspiration for these wound models. For example, using CD44 (basal) versus Keratin 10 (spinous) positive epithelial cells to examine barrier reconstitution would be clinically relevant. Another example would be to utilize CD36-positive fibroblast subpopulations to examine myofibroblast transformation and scar generation. Further, the ability to study more complex interactions by addressing crosstalk in multiplexed models, as well as building more sophisticated skin-on-a-chip multicompartmentalized analytical systems, is particularly attractive for current AI–ML robotic automation. Integration of in vitro and microphysiological data with AI and ML tools can greatly enhance model predictability and reproducibility. Though AI-driven predictions from in vitro datasets reflect cellular behaviors, it does not complete wound closure trajectories in patients. Structured, time-stamped datasets capturing quantitative outputs, such as migration rate, ECM synthesis, cytokine levels, and morphological features, can train algorithms such as random forest classifiers, transformer networks, or graph neural networks to predict healing trajectories. AI-based analytics combined with automated imaging and liquid handling can standardize assay benchmarking and accelerate discovery of therapeutic combinations for both acute and chronic wounds. In addition, establishing open, annotated repositories linking imaging, molecular, and functional readouts from wound models would enable cross-study validation and foster reproducible AI pipelines. Integrating multimodal data from omics, microscopy, and biomechanical sensors could further refine predictions of healing phases and therapeutic response, ultimately guiding personalized treatment strategies.4,14,16
Like many other applications, AI–ML appears to have transformative potential for wound healing research. However, concrete datasets and pipelines remain underdeveloped. Recent studies demonstrate feasibility of DeepLabv3+ models trained on 4,000+ prospective/retrospective wound images achieved 92% dice coefficient for automated segmentation and 78% tissue classification accuracy, enabling real-time mobile deployment. 174 XGBoost algorithms applied to Vascular Quality Initiative registry data (n = 235,677) predicted 1-year major adverse limb events with Area Under the Receiver Operating Characteristic curve (AUROC) 0.94, surpassing logistic regression (AUROC 0.67). 175 Time-series analysis of in vitro wound closure using enhanced UNet++ segmentation combined with temporal convolutional networks accurately modeled healing trajectories across epithelial phenotypes. 176 Future studies should generate structured, time-staged datasets from MPS platforms incorporating live-cell imaging, multiplexed biosensors (pO2, pH, and cytokines), and single-cell RNA-seq at defined healing phases (days 0, 3, 7, and 14). These AI-ready datasets, formatted as longitudinal CSV/JSON arrays with standardized ontologies, will enable predictive modeling of healing failure and therapeutic response through ML algorithms including XGBoost, random forests, and deep learning architectures. Computational collaborations should prioritize federated learning frameworks to integrate multilab MPS data while preserving proprietary models, establishing reproducible pipelines from in vitro systems to clinical translation.169,170
Another example is the distinction between migration and proliferation-coupled migration of each cell type in healing that should be separated and quantified by examining responses over time and use of inhibitors, such as Mitomycin C. Each cell type not only has a different rate of proliferation (turn over) but also migrates at different rates. A keratinocyte migrates and proliferates rapidly (within 12 and 24 h, respectively), while fibroblasts are slower in both migration and proliferation. Endothelial cells predominantly migrate at 48 h, while smooth muscle cells transition from migration-dominant to proliferation-dominant mechanisms after 24 h. To standardize these, a core assay panel capturing each phase over specific time periods is critical using clear and objective outcomes (Supplementary Fig. S10). The outcomes themselves must be functionally valid and easily measurable to be clinically relevant. For example, keratinocyte migration is assessed by percentage wound area reduction and re-epithelialization rate in μm/h, while fibroblast proliferation and collagen deposition are assessed through hydroxyproline quantification and PCNA/Ki-67 immunofluorescence as secondary outcomes. ECM remodeling through quantitative MMP-2/TIMP-2 and MMP-9/TIMP-1 ratios using gelatin zymography can serve as tertiary endpoints.
Another resource for benchmarking these in vitro models is to utilize the recently described clinical wound scoring systems such as the HEALS-A histological scale (Healing, Epithelialization, Angiogenesis, Leukocytes, Scar tissue, Appendages) and the GRADES clinical scale (Granulation tissue, Redness/edema, Appearance of wound, Devitalized tissue, Exudate, Surrounding skin). Cross-study benchmarking comparisons could stratify wound etiology as key factor distinguishing acute, diabetic, ischemic, and infected wounds—as healing kinetics differ substantially among them necessitating significant adjustment of sample sizes. For the models of wound healing described here, community adoption of standardized protocols including defined cell passage numbers, validated matrix compositions, standardized perfusion rates, and predetermined sample sizes could enable deposition of raw datasets into open repositories (e.g., GigaScience, Zenodo), facilitating meta-analyses, reproducibility validation, and statistical comparison across multilaboratory studies with rigorous benchmarking standards.
CRITICAL LIMITATIONS AND IMPLEMENTATION BARRIERS
Technical complexity and cost
These approaches require specialized fabrication (soft lithography and 3D bioprinting), perfusion control systems, environmental chambers with programmable oxygen/glucose control, and integrated biosensing—infrastructure beyond the capacity of most labs. Initial costs for organ-on-chip systems and smart biosensors create substantial barriers, particularly for resource-limited settings. 171
Insufficient training
Specialized biomedical engineering expertise is necessary for many of these platforms beyond cell biology and bioanalytical chemistry. Training programs and masterclasses in MPS are emerging but remain limited in availability. 172
Validation and standardization gaps
Lack of standardized protocols for cell sourcing, matrix composition, perfusion rates, and validation metrics impedes cross-study comparisons. Quality control criteria specific to wound healing MPS remain undefined.173,177
Scalability challenges
While high-throughput MPS platforms with 12–96 independently addressable tissue units have been demonstrated, scaling to pharmaceutical screening throughput while maintaining physiological relevance remains challenging. Vascularization of large constructs, achieving uniform perfusion across all units, and minimizing edge effects require continued engineering optimization.149,178,179
Regulatory uncertainty
Acceptance of MPS data by regulatory agencies for therapeutic development remains inconsistent, limiting industry adoption. FDA Modernization Act 2.0 supports MPS as reduced alternatives to animal testing, but validation requirements and acceptance criteria remain under development. 171
Biological complexity
Even advanced MPS platforms remain simplified relative to in vivo wound healing. Integration of lymphatic drainage, innervation, appendages (hair follicles and sweat glands), and the complete immune cell repertoire (neutrophils, macrophages, T cells, and mast cells) remains technically challenging. Current models capture selected aspects of wound biology but cannot yet recapitulate the full spatiotemporal complexity.150,172,180
Important limitation
In vitro models are reductionist systems. They allow controlled study of mechanistic components such as cell migration, proliferation, ECM synthesis, and angiogenesis but cannot replicate the full spatiotemporal complexity of wound healing in vivo, which involves systemic immune regulation, vascular perfusion, and multiorgan interactions.
AUTHORS’ CONTRIBUTIONS
D.K.: Conceptualization, literature search, drafting the article, figure preparation, and data extraction. M.A.: Conceptualization, literature search, drafting the article, table preparation, and data extraction. R.V.: Data extraction, drafting the article, figures, and tables preparation. A.N.: Data extraction, table development, and drafting the article. S.C.: Illustration and figure preparation and drafting the article. I.G.: Data extraction and drafting the article. A.L., J.L., and S.S.: Reviewing and drafting the article. P.A.: Conceptual oversight, guidance on structure and content, critical review, and final approval of the article.
Footnotes
ACKNOWLEDGMENTS AND FUNDING SOURCES
This work was supported by Commercialization of Advanced Technologies (CAT), Business Entrepreneurship Partnership,
AUTHOR DISCLOSURE AND GHOSTWRITING
The authors declare no conflicts of interest. No ghostwriting was involved in the preparation of the article; all contributing authors agree to the final submitted version of the article. Artificial intelligence tools were used for language and grammar editing but not for data analysis or interpretation.
ABOUT THE AUTHORS
Supplemental Material
Abbreviations and Acronyms
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.