
Abstract
Rumination is an established cognitive vulnerability for depression. Despite substantial work on the environmental origins of rumination, the heritability of rumination has not been examined, and it is not known whether rumination accounts for some of the genetic vulnerability associated with depression. A total of 756 adolescent twins aged 12 to 14 years completed the Response Styles Questionnaire and multiple measures of depressive symptoms. Brooding correlated positively, and distraction negatively, with concurrent depressive symptoms. Estimated heritabilities were 54% for depression, 21% for brooding, 37% for reflection, and 30% for distraction. Bivariate genetic analyses suggested that (a) individual differences in distraction share both genetic and environmental sources of variation with depression and (b) although the heritable influences on brooding are small, they account for the majority of the relationship between brooding and depression (h2 = .62).
Roughly 1 in 10 adolescents will experience major depression or dysthymia by age 18 (Merikangas et al., 2010). Adolescents with depressive symptoms who do not meet Diagnostic and Statistical Manual of Mental Disorders (fourth edition) criteria for depression nonetheless experience significant impairment (e.g., González-Tejera et al., 2005) and are at elevated risk for later development of major depression and suicidal behaviors (Fergusson, Horwood, Ridder, & Beautrais, 2005; Klein, Shankman, Lewinsohn, & Seeley, 2009). To inform intervention for depression, we must rigorously examine risk factors and their developmental origins during this critical adolescent stage. Rumination, “the process of thinking perseveratively about one’s feelings and problems” (Nolen-Hoeksema, Wisco, & Lyubomirsky, 2008, p. 400), is an established cognitive vulnerability for depression. Higher levels of rumination are associated with greater risk for onset of depression, longer duration, and increasing severity of symptoms (Abela & Hankin, 2011; Nolen-Hoeksema et al., 2008). Given rumination’s predictive power for the onset of depression, researchers have focused on the origins of rumination. Despite a growing body of research on the environmental origins of rumination (Hankin et al., 2009), little research on possible genetic origins of rumination exists. The lack of attention to genetic factors is understandable, given that the study of rumination began within the coping framework, which was not biologically oriented. However, the evidence that adolescent depressive symptoms are moderately heritable (Lau & Eley, 2006, 2010; Rice, Harold, & Thapar, 2002) invites the question of whether rumination differences account for some of the genetic vulnerability associated with depression. Thus, we aimed to illuminate the genetic and environmental influences on rumination and its relationship to a composite measure of depressive symptoms using a community sample of adolescent twins. Understanding the magnitude of the genetic and environmental overlap between rumination and depression first requires analysis of the heritability of rumination. Initial analyses are best conducted in a community sample, which provides a baseline for future genetic study in diagnostic groups.
We have observed the recent refinement in studies of rumination to remove depression-confounded items and to recognize subcomponents that differentially relate to depression: brooding and reflection (Armey et al., 2009; Joormann, Dkane, & Gotlib, 2006; Treynor, Gonzalez, & Nolen-Hoeksema, 2003). The brooding items reflect “moody pondering” (Treynor et al., 2003, p. 252), or passively focusing on one’s problems and their consequences, whereas the reflection items represent a purposeful attempt to understand and overcome one’s problems. Brooding is more strongly associated with concurrent depression than is reflection and also predicts the development of depressive symptoms and diagnoses (Armey et al., 2009; Gibb, Grassia, Stone, Uhrlass, & McGreary, 2012; Treynor et al., 2003).
The literature on the development of rumination has focused on environmental factors. Children who experience overcontrolling parenting and a negative-submissive style of family interaction tend to brood more as adolescents (Hilt, Armstrong, & Essex, 2012). Parenting in both childhood and adolescence appears to play an important role in the development of rumination. Adolescent girls whose mothers encourage emotional expression are more likely to ruminate (Cox, Mezulis, & Hyde, 2010). Although children of depressed mothers brood more than children of nondepressed mothers, this difference does not appear to be due to modeling of the mother’s rumination (Gibb et al., 2012). However, examinations of familial resemblance for rumination that are not genetically informative neglect possible passive and evocative gene-environment correlations (e.g., Lau & Eley, 2008b). Additionally, the inconsistent nature of findings regarding two specific genetic variants and rumination (Beevers, Wells, & McGeary, 2009; Hilt, Sander, Nolen-Hoeksema, & Simen, 2007; cf. Clasen, Wells, Knopik, McGeary, & Beevers, 2011; Gibb et al., 2012) points to a need to establish the overall magnitude of the genetic influences on rumination prior to embarking on candidate gene studies or at least to provide a framework for interpretation of candidate genes and other more comprehensive genomic approaches.
Although rumination has garnered substantial empirical support as a risk factor for depression, the relationship between other response styles and depression has been inconsistent. In the original response styles theory, Nolen-Hoeksema (1991) argued that the use of positive distractions was an adaptive alternative to rumination. However, distraction and depression correlate inconsistently in both cross-sectional and longitudinal studies (Nolen-Hoeksema et al., 2008; Rood, Roelofs, Bögels, Nolen-Hoeksema, & Schouten, 2009). A ratio approach (rumination:distraction) appears to be more predictive of changes in depressive symptoms than either of the subscales alone (Abela, Aydin, & Auberach, 2007; Hilt, McLaughlin, & Nolen-Hoeksema, 2010), suggesting that individuals who have both a greater tendency to ruminate and a lesser tendency to distract are at the greatest risk for experiencing depressive symptoms. Some researchers have speculated that the more specific reflection construct may be an adaptive form of rumination (Watkins, 2008). Although the relation between reflection and depression is positive concurrently (Joormann et al., 2006), it is negative longitudinally (Treynor et al., 2003), suggesting that reflection may be adaptive in the long term. Although no other study has examined the heritability of distraction or reflection as measured by the Response Styles Questionnaire (RSQ; Nolen-Hoeksema & Morrow, 1991), two twin studies of distraction based on the Coping Inventory for Stressful Situations (Endler & Parker, 1990) do exist. However, these studies report conflicting findings: Data from Polish adult twins showed that one third of the variance in the Distraction subscale score was associated with genetic differences (Kozak, Strelau, & Miles, 2005), whereas data from Canadian adult twins on the same subscale showed exclusively environmental effects (Jang, Thordarson, Stein, Cohan, & Taylor, 2007). We use the RSQ, which is central in the literature and which allows us to study reflection and distraction as well as rumination, and we focus on the developmentally crucial early adolescent period.
In summary, our main objective was to analyze brooding, reflection, distraction, and depressive symptoms in adolescent twins to determine whether response styles account for some of the genetic vulnerability associated with depression. Integrating the genetics of response styles with the genetics of depression is paramount to understanding the etiology of depression.
Methods
Participants
Participants were drawn from the Wisconsin Twin Project, a statewide, birth register–based twin sample that was mildly enriched for internalizing and externalizing symptoms when the twins were 7 years old (Lemery-Chalfant, Goldsmith, Schmidt, Arneson, & Van Hulle, 2006). Families with twin births in Wisconsin from 1989 to 2004 joined the project by responding to one of two recruitment letters. Eighty-one percent of families contacted responded favorably. The University of Wisconsin Institutional Review Board approved this study. Parents provided consent, and adolescents assent, for their participation. All participants were paid. Our sample comprised 756 adolescent twins aged 12 to 14 years (M = 13.1, SD = 1.3; 53% female). The sample was 88.6% Caucasian, representative of Wisconsin’s population. Furthermore, 35.8% of the sample was monozygotic (MZ); 33.8%, same-sex dizygotic (DZ); and 30.4%, opposite-sex DZ.
Measures
Depression composite
We created a composite of depressive symptoms based on four depression-relevant self-report scales or subscales. The Children’s Depression Inventory (Kovacs, 1985) measures negative mood, interpersonal difficulties, negative self-esteem, ineffectiveness, and anhedonia during the previous 2 weeks and differentiates emotionally distressed children from nondistressed children (Kovacs, 1981; Saylor, Finich, Spirito, & Bennett, 1984; Smucker, Craighead, Craighead, & Green, 1986). The Early Adolescent Temperament Questionnaire–Revised (Capaldi & Rothbart, 1992; Ellis & Rothbart, 2001) measures social-emotional functioning and aspects of temperament related to self-regulation, reactivity, and emotionality. The Depressed Mood subscale of the questionnaire measures unpleasant affect, lowered mood, and loss of enjoyment and interest and is associated with higher levels of temperamental negative affectivity (Ellis & Rothbart, 2001). The Depression subscale of the Health and Behavior Questionnaire (Armstrong, Goldstein, & the MacArthur Working Group on Outcome Assessment, 2003) measures mental health symptoms, physical health, and academic and social functioning; it discriminates between clinic-referred children and controls (Ablow et al., 1999) and corresponds well with Diagnostic and Statistical Manual of Mental Disorders (fourth edition) symptoms and diagnoses in children (Lemery-Chalfant et al., 2007). Finally, we included symptoms of major depressive disorder from the Diagnostic Interview Schedule for Children, Version IV (Fisher et al., 1997). Of the 756 participants, 97% had data from at least three measures (with 79% having data from all four measures). Each measure was standardized, and all available measures for each participant were averaged to create a depression composite.
Response styles
Response styles were assessed with the 22-item Ruminative Response Scale (brooding, reflection, and depression-confounded items) and the 11-item Distraction Scale from the RSQ. Participants indicate how frequently they engage in ruminative or distractive thoughts/activities when they are upset (1 = almost never, 4 = almost always). The 5 brooding items assess moody pondering (e.g., “I think ‘What am I doing to deserve this?’”). The 5 reflection items assess neutral pondering (e.g., “I go away by myself and think about why I feel this way”). The 12 depression-confounded items are items similar to depressive symptoms (e.g., “I think about how hard it is to concentrate”); given the focus of this article, we dropped the depression-confounded items from our analyses. The distraction items assess actions or plans to engage in actions that divert one’s attention when upset (e.g., “I do something fun with a friend”). Internal consistencies were .71 for brooding, .72 for reflection, and .87 for distraction.
Zygosity
Zygosity was determined at multiple phases on the basis of multiple methods, including maternal report on the Zygosity Questionnaire for Young Twins (Goldsmith, 1991), observer ratings, and genotyping.
Socioeconomic status
We standardized and averaged mother and father years of education and annual family income to create a composite socioeconomic status score. Mothers had an average education of 14.6 years and fathers, 14.0 years; median family income was $60,000 to $70,000.
Genetic analyses
Twin studies utilize the varying degree of genetic similarity of MZ and DZ twins to parse genetic and environmental contributions to individual differences in behavior. Covariation between twins is due to correlated genetic factors (coefficient of 1.0 for MZ and 0.5 for DZ twins) and correlated shared environments (coefficient of 1.0 for both MZ and DZ twins). If MZs are more similar than DZs on the variable of interest, genetic variance is implicated. If, however, MZ and DZ twins are comparably similar, shared environmental influences are implicated. Nonshared environmental influences are nongenetic factors that make siblings different and are implied to the degree that MZ twins are dissimilar (Plomin, DeFries, McClearn, & Rutter, 2008).
We used maximum likelihood estimation techniques to fit structural equation models (Neale & Cardon, 1992) to partition the observed variation into latent additive genetic components (A; additive effect of multiple genes), shared environmental components (C), and nonshared environmental components (E; also includes measurement error), including 95% confidence intervals (CIs) for each component. Model fitting proceeded in a stepwise fashion: Individual parameters were dropped and the submodel fit compared to the fit of the full model based on the difference in log-likelihoods (LLs), which is distributed as a chi-square (χ2 difference = −2LLsubmodel – 2LLfullmodel), with degrees of freedom equal to the difference in the number of estimated parameters. A significant change in chi-square indicates that the submodel should be rejected in favor of the full model. In general, the model that explains the most variance with the fewest parameters is preferred (Neale & Cardon, 1992). All models were fit per the statistical program OpenMx (Boker et al., 2011).
Results
Phenotypic analyses
Means and standard deviations for the depression composite and response styles are included in supplementary materials (http://cpx.sagepub.com/content/by/supplemental-data). There was a modest gender difference in the depression composite, with girls (M = 0.06, SD = 0.88) scoring higher than boys (M = −0.06, SD = 0.75), Welch’s t(752) = 1.94, p = .052, d = .15. There was a moderate gender difference in reflection, with girls (M = 2.17, SD = 0.68) reflecting significantly more than boys (M = 1.92, SD = 0.58), Welch’s t(746) = 5.36, p < .001, d = .38. The lack of a gender difference in brooding is unsurprising, given that the gender difference in rumination may not emerge until age 15 (Cox et al., 2010). Subscale intercorrelations were run with one randomly selected twin from each pair (full results are in supplementary materials). The depression composite correlated with all response styles: r = .47, p < .001, for brooding; r = .14, p < .05, for reflection; r = −.40, p < .001, for distraction; and r = .66, p < .001, for the brooding:distraction ratio. Reflection correlated positively with both brooding and distraction (r = .48 and r = .45, respectively, p < .001), but brooding and distraction were not correlated (r = .09, ns). Age correlated positively with brooding and the brooding:distraction ratio (r = .12, p < .05, for both correlations). Socioeconomic status correlated negatively with the depression composite (r = −.47, p < .001).
Genetic analyses
Prior to genetic analyses, all measures were log-transformed to correct for nonnormal distributions. Residualized scores accounted for age, sex, and socioeconomic status effects (McGue & Bouchard, 1984). Twin intraclass correlations for the residualized variables were as follows: depression composite (rMZ = .53, rDZ = .26, p < .01), brooding (rMZ = .22, p < .05; rDZ = .10, ns), reflection (rMZ = .40, p < .001; rDZ = .14, p < .05), and distraction (rMZ = .40, p < .05; rDZ = .02, ns). Doubling the difference between MZ and DZ intraclass correlations roughly estimates heritability (Falconer & Mackay, 1996).
Univariate models examined the depression composite, brooding, reflection, and distraction independently (see Table 1 for model fit). We found no evidence for shared environmental influences on any of the variables. Significant additive genetic influences were implicated for all variables. The estimated heritability of the depression composite was 54% (95% CI: 42%–63%); brooding, 21% (95% CI: 7%–34%); reflection, 37% (95% CI: 24%–49%); and distraction, 30% (95% CI: 17%–42%). The remainder of the variability could be accounted for by nonshared environmental factors: 46% for depression composite (95% CI: 37%–58%), 79% for brooding (95% CI: 66%–93%), 63% for reflection (95% CI: 51%–76%), and 70% for distraction (95% CI: 58%–83%).
Fit Statistics for Univariate Models
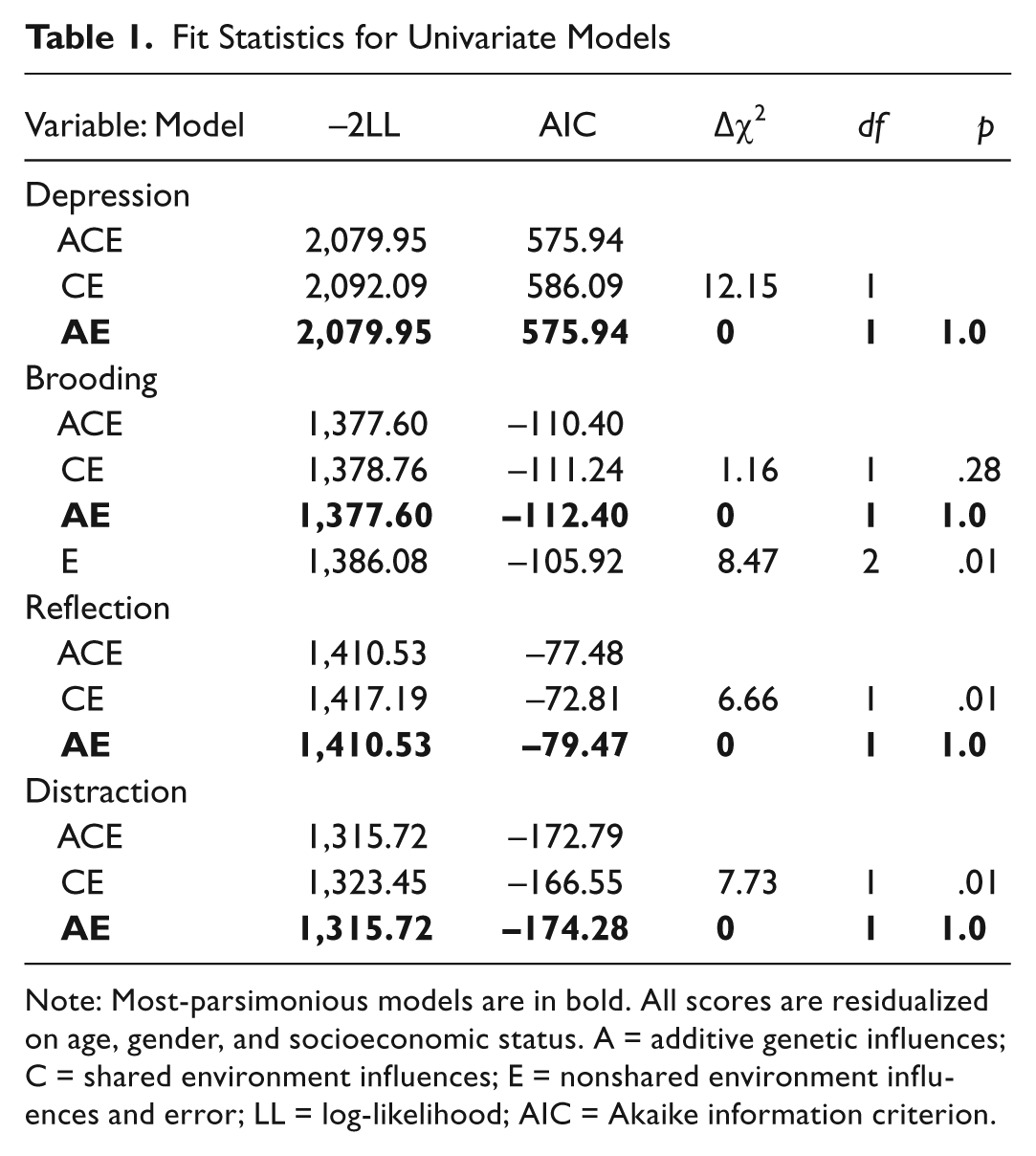
Note: Most-parsimonious models are in bold. All scores are residualized on age, gender, and socioeconomic status. A = additive genetic influences; C = shared environment influences; E = nonshared environment influences and error; LL = log-likelihood; AIC = Akaike information criterion.
Bivariate genetic analyses
Bivariate genetic analyses examined the unique and shared genetic and environmental influences on two correlated variables. Because reflection did not strongly correlate phenotypically with the depression composite, we limited our analyses to two models: (a) brooding and the depression composite and (b) distraction and the depression composite (see Fig. 1). Fitting the bivariate Cholesky model revealed substantial additive genetic influences (h2 = .62) and nonshared environmental influences (e2 = .38) on the relationship between brooding and the depression composite, as well as unique genetic and nonshared environmental influences on each (−2LL = 3,306.50, Akaike information criterion = 314). The genetic correlation was .83 and the environmental correlation, .27, indicating substantial genetic contributions to the covariance between brooding and depression. Model fitting also revealed substantial additive genetic influences (h2 = .48) and nonshared environmental influences (e2 = .52) on the relationship between distraction and the depression composite, as well as unique genetic and nonshared environmental influences on each (−2LL = 3,304.10, Akaike information criterion = 312). The genetic correlation was −.40 and the environmental correlation, −.30.
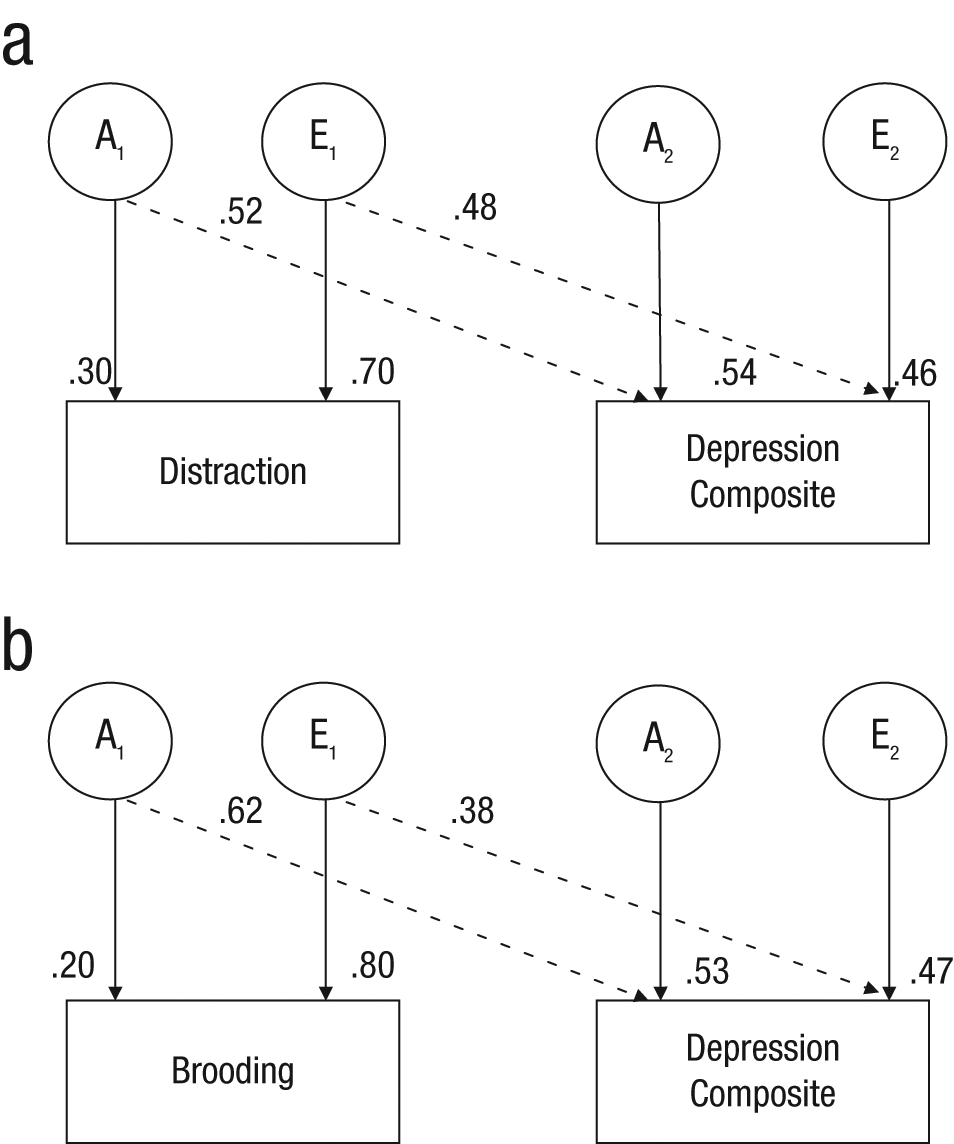
Bivariate genetic models of (a) distraction and depression and (b) brooding and depression.
Discussion
We estimated the magnitude of the genetic and environmental effects on three components of response styles (brooding, reflection, and distraction) and the magnitude of the genetic and environmental contributions to the covariance between response styles and depression. Brooding, reflection, and distraction have small to moderate heritable components but are mostly influenced by individual-specific environmental factors (h2 = .21 for brooding, .37 for reflection, and .30 for distraction). Consistent with our results, many twin studies of psychopathology report no evidence for shared environmental effects by adolescence (Kendler et al., 2008; Rhee & Waldman, 2002). Our estimates of genetic and nonshared environmental influences on depression were similar to those reported by Lau and Eley (2006), suggesting that our depression findings are representative.
Our key contribution to understanding pathways to a depressive phenotype is our estimation of the magnitude of the genetic and environmental overlap between response styles and depression. The bivariate analysis of distraction and depression, coupled with the negative correlation between distraction and depression, suggests that the same genetic factors that contribute to distraction may protect against concurrent depression (h2 = .48). Likewise, nonshared environmental influences that lead one to use distraction could also protect against depression (e2 = .52). Showing that this developmental interpretation of our cross-sectional findings is valid should be a priority for future research. Although brooding has a low heritability (i.e., is most strongly influenced by environmental factors unique to each individual), the bivariate analysis indicated that most of the covariance between brooding and depression is due to a common set of underlying genetic influences (h2 = .62, genetic correlation = .83). One interpretation of this result is that part of the genetic effect on depression is mediated by the maladaptive tendency to brood when confronted with loss and goal blockage.
The results of this study suggest four potential avenues for future research. First, given brooding’s low heritability, candidate gene studies of brooding should not be a high priority, at least for the measures and type of sample that we used. However, the strong genetic correlation between brooding and depressive symptoms suggests that a novel candidate gene design targeting depressive brooders (individuals high on both depression and brooding compared with those low on both) might illuminate the shared genetic vulnerability.
Second, as noted earlier future research should examine genetic and environmental contributions to the relationship between response styles and depression longitudinally. Because all constructs in this study were measured concurrently, it remains unclear from our results alone whether brooding is a risk factor for depression or is merely associated with depressed mood concurrently. However, our demonstrating genetic underpinnings of this association does support the argument that the association is not an artifact of measurement. A longitudinal approach would also illuminate how the heritability of response styles and their relationships to depression might change at different developmental stages (see, e.g., Kendler et al., 2008; Lau & Eley, 2006).
Third, it will be important to conduct genetically informative analyses with more nuanced measures of rumination. The entire realm of brooding-like behavior is likely not captured by five items on the RSQ. The RSQ brooding items, which focus on typical responses to being upset, are qualitatively different from ruminating about depressed affect (Nolen-Hoeksema, 1991) and ruminating about negative life events (Mezulis, Abramson, & Hyde, 2002; Robinson & Alloy, 2003).
Finally, future research should identify the unique and shared relationships between distinct cognitive vulnerabilities and depression. Attributional style is also heritable (Lau & Eley, 2008a) and associated with the BDNF Val66Met genetic polymorphism (Haeffel, Eastman, & Grigorenko, 2012). To inform both etiological understanding and intervention, it will be important to determine whether different cognitive vulnerabilities share distinct or overlapping genetic variance with depression; this could be determined via multivariate Cholesky analyses. Extended family designs could also serve to provide added strength to identify shared environmental influences, as well as to help clarify possible gene-environment interactions and correlations.
Footnotes
Declaration of Conflicting Interests
The authors declared that they had no conflicts of interest with respect to their authorship or the publication of this article.
Funding
This work was supported by the National Institute of Mental Health (R01 MH59785 to Goldsmith and Lemery-Chalfant; T32 MH018931 to Moore), the Conte Neuroscience Center (P50 MH084051), and the National Science Foundation (DGE-071823 to Salk). Infrastructure support was provided by the Waisman Center (P30 HD03352). Any opinion, findings, and conclusions or recommendations expressed in this material are those of the authors and do not necessarily reflect the views of the National Science Foundation.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.