
Abstract
Many treatments for anxiety utilize associative learning theory and involve mental imagery components. However, the roles associative learning and imagery play in anxiety disorders and their treatments are still largely unknown. Here we show that voluntary mental imagery can undergo associative learning in the same manner as normal sensory perception. After conditioning voluntary mental images with emotion-evoking stimuli, perceptual stimuli of the same content were found to produce the associated emotional response—indicating generalization from imagery to perception. This associative learning with imagery was found to be orientation selective and could not be accounted for by alternative, non-imagery-based interpretations of the data. These results support pictorial theories of mental imagery by indicating the interchangeability of imagery and perception in associative learning and demonstrate that voluntary mental images can drive associative learning, providing a mechanistic account of clinical practice involving mental imagery.
Keywords
In recent times the field of learning has seen rapid growth in the areas of fear and hedonic processes (Lissek et al., 2005), perceptual learning (Sasaki, Nanez, & Watanabe, 2010), motor learning (Jeannerod, 1995), and learning driven by neural-feedback signals (Shibata, Watanabe, Sasaki, & Kawato, 2011). The learning paradigms of conditioning, counterconditioning, and extinction form the backbone of the underlying mechanisms of many psychological disorders and their treatments (Milad, Rauch, Pitman, & Quirk, 2006; Myers & Davis, 2002; Rauch, Shin, & Phelps, 2006). Many behavioral interventions for psychological disorders rely on counterconditioning or extinction-like approaches as part of cognitive behavioral therapy (CBT) to produce behavior change. Exposure, for example, is a key component of many effective treatment strategies for anxiety disorders. A clinically anxious individual may receive real or imagined exposure to his or her feared stimulus or situation with the goal of reducing fear and anxiety (Holmes & Mathews, 2010; Hunt & Fenton, 2007). Progress in understanding the many aspects of learning has led to great improvements in the treatment of many mental illnesses, such as anxiety disorders and various phobias, which are often modeled on fear learning (Lissek et al., 2005; Milad et al., 2006; Rauch et al., 2006; Watson & Rayner, 1920).
Treatments for disorders of anxiety often focus on their imagery-like symptoms (Edwards, 2007; Holmes & Mathews, 2010). One such treatment is CBT with imagery rescripting (Arntz, Tiesema, & Kindt, 2007; Holmes, Arntz, & Smucker, 2007; Smucker, Dancu, Foa, & Niederee, 1995), wherein patients are directed to focus on and then transform the content of their distressing mental imagery. Though such techniques have been effective for anxiety sufferers, current imagery-based treatments show room for improvement (Bradley, Greene, Russ, Dutra, & Westen, 2005; Foa & Meadows, 1997; Harvey, 2003). A greater understanding of the mechanisms of these disorders and their treatment options is pivotal for the development of new treatment methods that more directly address their underlying causes.
Mental imagery, a perceptual-like experience without corresponding sensory input, is typically based on the memories of previously perceived stimuli. The generation of mental images can be voluntary, explicitly crafted by the observer, or involuntary, automatically cued by some external stimulus (Albright, 2012; Brewin, Gregory, Lipton, & Burgess, 2010; Holmes & Mathews, 2010). For anxiety sufferers these images are typically involuntary and related to the individual’s specific fears or concerns (Brewin et al., 2010; Dadds, Hawes, Schaefer, & Vaka, 2004; Hirsch & Holmes, 2007). Such imagery is predominantly visual, but can also be auditory and tactile (Ehlers & Steil, 1995). Instances of distressing mental imagery tend to precede or amplify other nonimagery symptoms (Hirsch, Mathews, Clark, Williams, & Morrison, 2006; Holmes, Geddes, Colom, & Goodwin, 2008; Holmes & Mathews, 2010; Pitman et al., 1990), indicating that they play a key role in the maintenance of the disorder. Unfortunately, a comprehensive understanding of what mental imagery is and how it fits into current theories of associative learning is still missing.
Associative learning has been well documented with various perceptual stimuli (De Houwer, Thomas, & Baeyens, 2001; Martin & Levey, 1978; Milad et al., 2006). A review by Dadds, Bovbjerg, Redd, and Cutmore (1997) suggested that mental imagery and perception are highly similar processes that might play interchangeable roles in associative learning. This view has gained support from the finding that the processes involved in imagery and perception can share similar patterns of neural activity (Behrmann, 2000; Slotnick, Thompson, & Kosslyn, 2005). More recently, it has been found that repeated generation of a mental imagery stimulus will subsequently improve an observer’s ability to discriminate between perceptual stimuli (Tartaglia, Bamert, Mast, & Herzog, 2009), but repeated training of imagery does not lead to improvements in imagery itself (Rademaker & Pearson, 2012). In general, the evidence seems to indicate that mental imagery can act like perception (Kosslyn, Ganis, & Thompson, 2001; Pearson, Clifford, & Tong, 2008) and the relative strength of imagery is closely tied to the observer’s subjective reports of its vividness (Pearson, Rademaker, & Tong, 2011).
In the current study we sought to determine whether voluntary mental imagery could undergo associative learning as visual perception does. Specifically, we first sought to verify that our task could produce significant associative learning effects for perceptual stimuli (Experiment 1); we then utilized this same task to associate specific content of mental imagery with a set of emotion-evoking photographs (Experiment 2). The formation of these kinds of associations can result in evaluative conditioning (De Houwer et al., 2001; Martin & Levey, 1978), wherein the perceptual or imagery content will gain new emotional significance subsequently inducing fear-indicant responses, much like the original emotion-evoking photographs.
To test the emotional valence of the voluntarily generated mental images, we used an affective priming method (Fazio, Sanbonmatsu, Powell, & Kardes, 1986; Hermans, De Houwer, & Eelen, 1994). If the imagery patterns become associated with a specific emotion, they should facilitate the recognition of subsequently presented photographs with matching emotional content. The magnitude of this facilitation should be indicative of the strength of associative learning with imagery.
In addition, we sought to determine whether this associative learning can occur without color information (Experiment 3). It is has been suggested that various colors can have strong emotional associations (Gao et al., 2007; Kaya & Epps, 2004). Hence, we investigated associative learning with the same paradigm using achromatic imagery. Two additional tests were also conducted to rule out a potential nonmental imagery account of our results.
Finally, we assessed the orientation specificity of the associative learning with mental imagery (Experiment 4). Evidence shows that neurons within low-level visual cortex are sensitive to slight spatial rotations of visual stimuli, whereas activity in higher-level brain areas appears to be relatively insensitive (Blakemore & Campbell, 1969; Fang, Murray, Kersten, & He, 2005; Hubel & Wiesel, 1968). To show that associative learning for mental imagery is similarly orientation specific would indicate that such learning is contingent on activity within these low-level visual areas, such as primary visual cortex. Consequently, this would improve our understanding of the mechanisms underlying disorders involving mental imagery symptomatology. This would also suggest that the effectiveness of treatments for anxiety might be enhanced by focusing on the low-level sensory aspects of the distressing imagery symptoms. In addition, showing that imagery and perception have shared neural processing in relation to associative learning in early visual cortex would provide compelling evidence that visual mental imagery can be pictorial in nature.
Materials and Method
Participants
A total of 143 University of New South Wales students were recruited for the current study (nExperiment 1 = 79, nExperiment 2 = 30, nExperiment 3 = 19, nExperiment 4 = 15). All participants were students from the University of New South Wales who gave written, informed consent before the start of each experimental session. New participants were recruited for each experiment via Web advertisements. Fewer participants were recruited for each successive experiment in an attempt to produce a more conservative replication of the original results and efficient use of resources. Course credit or money was awarded upon completion of the study. The current study received ethical approval from the local committee.
Stimuli
All stimuli were created and presented using Psychtoolbox (Brainard, 1997) for Matlab (Version 7.10.0 R2010a) on a Mac mini computer and displayed on a 27 × 35.5 cm Philips 109P4 monitor at a resolution of 1152 × 870 at 75 Hz. Stimuli were presented on a black background with a circular fixation mark (diameter = 0.62°) in the center of the screen.
In Experiments 1, 2, and 4, two different Gabor patterns (σ = 4°; SF = 1 cycles/degree) were used as conditioned stimuli: a red vertical pattern (CIE color space x = 0.6, y = 0.37) and a green horizontal pattern (CIE x = 0.27, y = 0.62). In Experiment 3, the achromatic imagery experiment, both Gabor patterns were grayscale (contrast = 100%; max luminance = 45.0 cd/m2). In all experiments the background was black (max luminance = 0.01 cd/m2), as background luminance has been shown to attenuate imagery generation (Keogh & Pearson, 2011; Pearson et al., 2008; Sherwood & Pearson, 2010). Photographs from the International Affective Picture System (IAPS; Lang, Bradley, & Cuthbert, 2008) were used as unconditioned stimuli. These photographs were split into two categories based on their rating norms. “Pleasant” IAPS photos were those rated low on arousal and high on pleasantness. “Aversive” IAPS photos were those rated high on arousal and low on pleasantness. Photos rated high on both arousal and pleasantness were not used, as they are mostly sexually explicit in nature. Photos rated low on both scales were not used as they had no emotional content. In the associative learning phase a set of 12 photos (6 pleasant, 6 aversive) was used, and in the test phase a different set of 12 was used.
In Experiment 4, the orientation specificity experiment, the two original Gabor patterns (red = 90°, green = 0°) were accompanied by similar patterns that had been rotated 22° and 45°. Also, the number of IAPS photos was increased; 12 photographs were added to the associative learning phase (6 pleasant, 6 aversive) and 36 were added to the test phases (18 pleasant, 18 aversive). These additional photos were added to accommodate an increase in the total number of trials.
Procedure
All participants were seated in a dark room with their heads stabilized on a headrest 57 cm from the monitor. Participants undergoing associative learning for mental imagery had previously been shown both Gabor patterns and instructed to imagine them when a corresponding letter cue was shown. In Experiments 2 and 4 the letter cue “R” corresponded to the red vertical Gabor and “G” corresponded to the green horizontal Gabor. In Experiment 3, the achromatic imagery experiment, the letter cue “V” corresponded to the vertical Gabor and “H” corresponded to the horizontal Gabor. Participants were instructed to fixate on a circular fixation mark in the center of the screen for the duration of the experimental session. Figure 1 shows the trial designs for the two associative learning phases and three test phases used in the current study.

Experimental timelines. (a) Experiment 1: associative learning with perceptual stimuli. (b) Experiments 2, 3, and 4: associative learning with mental imagery. (c) All Experiments: test of association between perceptual Gabor and emotional photograph. (d) Experiment 3: test of association between letter cue and emotional photograph. (e) Experiment 3: test of association between perceptual Gabor and letter cue.
It is important to note that voluntary mental imagery was used only in the associative learning phase and not in the test phase. Hence, any effect of learning in this design is inherently a generalization, or crossover, from an imagined representation to a perceptual one.
Associative learning phase
At the start of each experimental session participants completed multiple trials of the associative learning task. As shown in Figure 1a, the perceptual trials started with the presentation of one of the two randomly chosen perceptual Gabors for 1.5 s, followed immediately by the emotional photograph for 1.5 s. The screen then remained blank for a 2-s intertrial interval. As shown in Figure 1b, the mental imagery trials started with the presentation of one of the two randomly chosen letter cues for 1.5 s. This was followed by a blank screen for 5.5 s, during which the participants were to imagine the pattern indicated by the letter cue. Then either a pleasant or aversive photograph was displayed for 1.5 s, followed by a 2-s intertrial interval wherein the screen remained blank. In both learning tasks the participants were required to press the space bar as soon as the photograph was presented to help them maintain vigilance. This kind of associative learning technique is commonly used across the literature to associate emotional content with neutral sensory stimuli (De Houwer et al., 2001).
Throughout the associative learning phase the relationship between the Gabor patterns and their associated emotion-evoking photographs remained constant. The stimulus associations were counterbalanced across participants: For half of the participants the vertical Gabor patterns were paired with aversive photos and the horizontal Gabors with pleasant photos, and vice versa for the other half. Each Gabor was paired with all of its corresponding emotion-evoking photographs an equal number of times. These Gabor–photograph pairs were presented in a random order.
In Experiment 1 a total of 40 associative learning trials were conducted. In Experiments 2 and 3 a total of 84 learning trials with mental imagery were conducted. In Experiment 4 the number of learning trials was increased to 144, with 102 trials at the beginning of the study and another 42 halfway through the test phase. This “top-up” was done to accommodate for any potential extinction due to the increased number of test trials.
Gabor pattern: test phase
Participants in all experiments then completed 120 trials of a choice reaction time task on a new set of emotion-evoking photographs to test the strength of the previously learned associations. Figure 1c shows the design of a Gabor pattern test trial. One of the two previously imagined Gabor patterns was presented for 100 ms, immediately followed by the presentation of an IAPS photograph for 1 s. Participants were required to judge the emotional content of the photographs upon their presentation as quickly and accurately as possible. They could judge the photos as either “Pleasant” or “Aversive” by pushing either the 1 or 3 button on the keyboard. Participants were instructed to respond as fast as possible without sacrificing accuracy. If conditioning did occur, then reaction times should be faster when the pattern’s associated emotion accurately predicts the emotional content of the subsequent photograph.
In Experiment 4 the number of Gabor test trials was increased to 288, split evenly among the six Gabor pattern orientations (red 90°, red 68°, red 45°, green 0°, green 22°, and green 45°) and presented in random order.
Letter cue: test phase
Participants in Experiment 3 completed an additional priming task to test the strength of any learned associations between the emotional content and the letter cues used in the associative learning task. As Figure 1d shows, this letter cue test phase was identical to the Gabor test phase except that the Gabor patterns were replaced with their corresponding letter cues. The order of test phase presentation was counterbalanced between participants, with half of the participants completing the Gabor test first and the other half completing the letter cue test first.
Gabor cue: test phase
Participants in Experiment 3 also completed a third test to directly measure the strength of any learned association between the Gabors and their corresponding letter cues. Figure 1e shows the trial design for testing the priming from the Gabor patterns for the letter cues. Participants completed a total of 40 trials, consisting of 10 presentations for each combination of Gabor and letter cue. In each trial one of the two previously imagined Gabor patterns was physically presented for 100 ms, followed immediately by one of the two letter cues for 1 s. Upon presentation of a letter cue, participants were required to press the corresponding key on the keyboard as quickly and accurately as possible. This Gabor cue test was always completed last.
Data preparation
Incorrect emotionality judgments were removed from the data set, as were individual participant reaction times that were shorter than 250 ms, longer than 2,500 ms, or beyond 2.5 SD from that participant’s mean. In addition, 2 outlying participants were excluded from analysis in Experiment 3 due to having average mismatch-match scores beyond 2.5 SD of the overall group mean.
To measure the strength of associative learning for the voluntary mental imagery, each participant’s reaction-time data were split into matched and mismatched trials. Matched trials included the stimulus pairs that were presented during the associative learning, whereas mismatched trials consisted of stimulus pairs that were not. If matched trials produced significantly faster reaction times, this would be indicative of affective priming, suggesting that the previously neutral Gabor patterns had gained emotional content due to associative learning.
Results
Experiment 1: perceptual stimuli
Figure 2a shows the data from the Gabor pattern test phase following associative learning with perceptual Gabor patterns. A paired-samples comparison shows that reaction times were significantly faster when the preceding pattern’s associated emotion matched the emotional photograph (M = 563.9 ms, SEM = 7.84 ms) than when it was mismatched (M = 572.5 ms, SEM = 8.05 ms), t(78) = 3.29, p = .002, η2 = .12. This shows that our task can produce associative learning for perceptual stimuli.
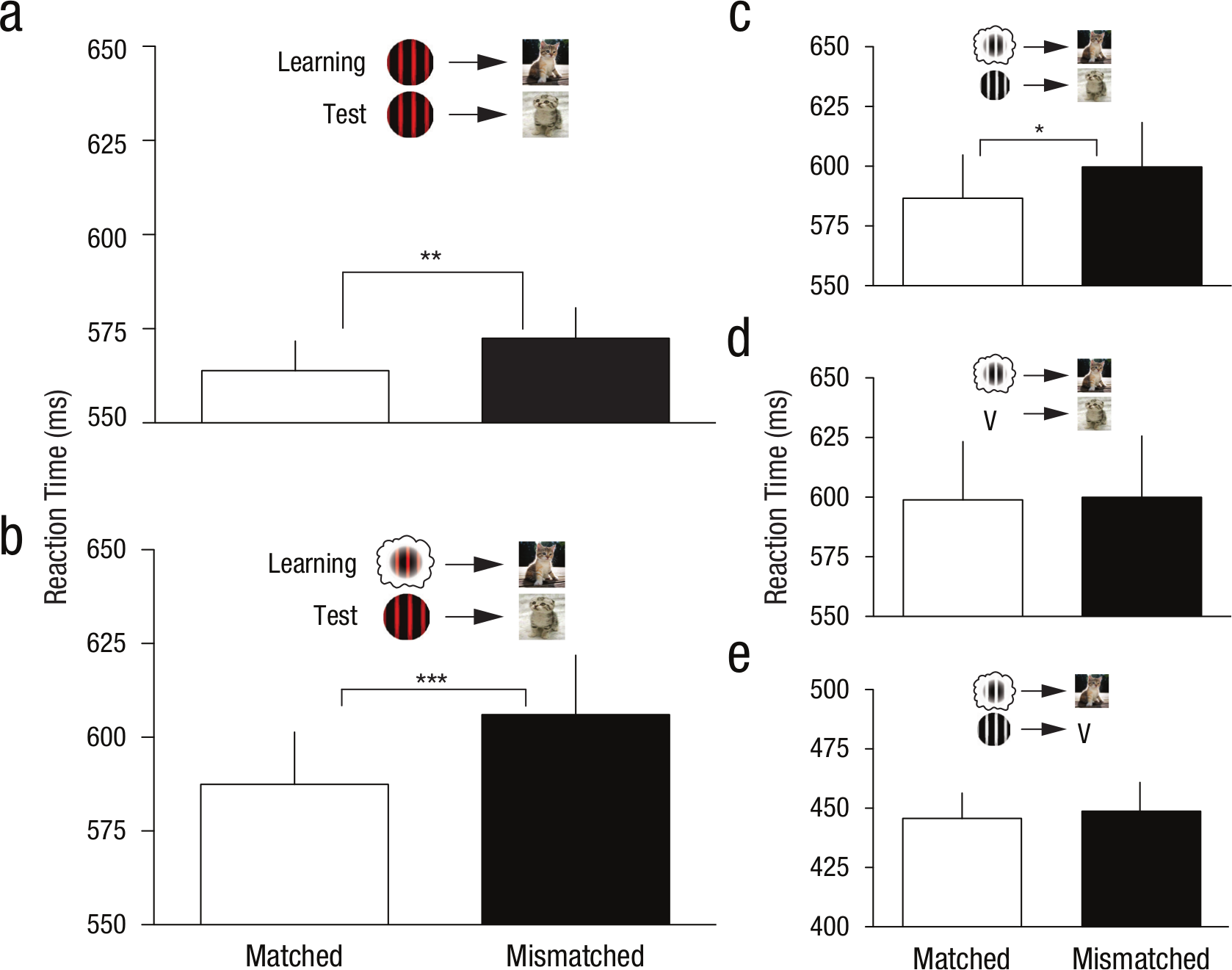
Mean reaction times for matched and mismatched stimuli during the test phases. Data indicate associative learning effects from sensory stimuli and imagined stimuli. (a) Experiment 1: Gabor-photo test preceded by perceptual conditioning (n = 79). (b) Experiment 2: Gabor-photo test preceded by imagery conditioning (n = 30). (c) Experiment 3: Achromatic Gabor-photo test preceded by achromatic imagery conditioning (n = 17). (d) Experiment 3: cue-photo test preceded by achromatic imagery conditioning (n = 17). (e) Experiment 3: achromatic Gabor-cue test preceded by achromatic imagery conditioning (n = 17).
Experiment 2: mental imagery
Figure 2b shows the data from the Gabor pattern test phase following associative learning with mental imagery. A paired-samples comparison shows that reaction times were significantly faster when the preceding pattern’s associated emotion matched the emotional content of the photograph (M = 587.44 ms, SEM = 13.95 ms) than when it was mismatched (M = 606 ms, SEM = 15.82 ms), t(29) = 4.14, p < .001, η2 = .37. This finding suggests that associative learning can occur between mentally generated stimuli and the emotional content of photographs.
Experiment 3: achromatic mental imagery
Figure 2c shows the data from the Gabor test phase following associative learning with achromatic mental imagery. A paired-samples comparison shows significantly faster reaction times when the pattern’s previously associated emotion matched the emotional photograph (M = 586.7 ms, SEM = 18.05 ms) than when it was mismatched (M = 599.7 ms, SEM = 18.6 ms), t(16) = 2.50, p = .023, η2 = .28. These data once again show a significant effect of associative learning with mental imagery. In addition, this replication provides further evidence that mental imagery can undergo associative learning, and also suggests that color information is not necessary for such associative learning.
However, one possible alternative explanation of the data is that during the test phase the perceptual Gabor patterns induced a representation of the corresponding letter cues, and these letter cues were actually driving the affective priming shown in Figure 2c. To assess this alternative account, each participant as part of Experiment 3 completed two additional test phases. These tests measured the strength of the associations between the letter cues and the photographs, and between the perceptual Gabor patterns and the letter cues. If both tests show significant priming, this would suggest an alternative explanation of our data and that mental imagery did not necessarily undergo associative learning.
Figure 2d shows the data from the letter cue test phase following associative learning with achromatic mental imagery. This test was conducted to determine if there was any direct link between the imagery cues and the emotion-evoking photographs. Associative learning found in this task would be attributable to either conditioning between the cue and the emotional photographs, or indirect association via the mental imagery Gabors. However, a paired-samples comparison showed no significant difference in the reaction times between matched (M = 598.9 ms, SEM = 24.44 ms) and mismatched pairs (M = 599.9 ms, SEM = 25.69 ms), t(16) = 0.21, p = .838. These data indicate that the letter cues did not undergo significant associative learning according to our measure.
Figure 2e shows the data from the Gabor-cue test phase following associative learning with achromatic mental imagery. This test was conducted to determine if the Gabor patterns were associatively linked to the letter cues. A paired-samples comparison showed no significant difference in reaction times between the matched (M = 445.7 ms, SEM = 10.73 ms) and mismatched conditions (M = 449.1 ms, SEM = 11.78 ms), t(16) = 0.43, p = .675. This indicates that participants did not reliably associate the Gabor patterns with their letter cues, despite being shown physical versions of the Gabors at the start of the experiment and being instructed to imagine them when the appropriate letter cue was shown.
Experiment 4: orientation specificity of mental imagery
Figure 3 shows the mean reaction-time differences between the matched and mismatched trials for Gabor patterns rotated 0°, 22°, and 45°. A significant change was found across the three levels of orientation, repeated measures ANOVA, F(2, 28) = 4.11, p = .042, η2 = .23, n = 15. Post hoc analyses showed that the difference scores for Gabors rotated 0° (M = 18.05 ms, SEM = 7.89 ms) and 22° (M = 16.76 ms, SEM = 7.43 ms) were significantly greater than Gabor patterns rotated 45° (M = −12.06 ms, SEM = 11.75 ms), p < .05. These results show that the associative learning was specific to the given spatial orientation of the Gabor patterns.
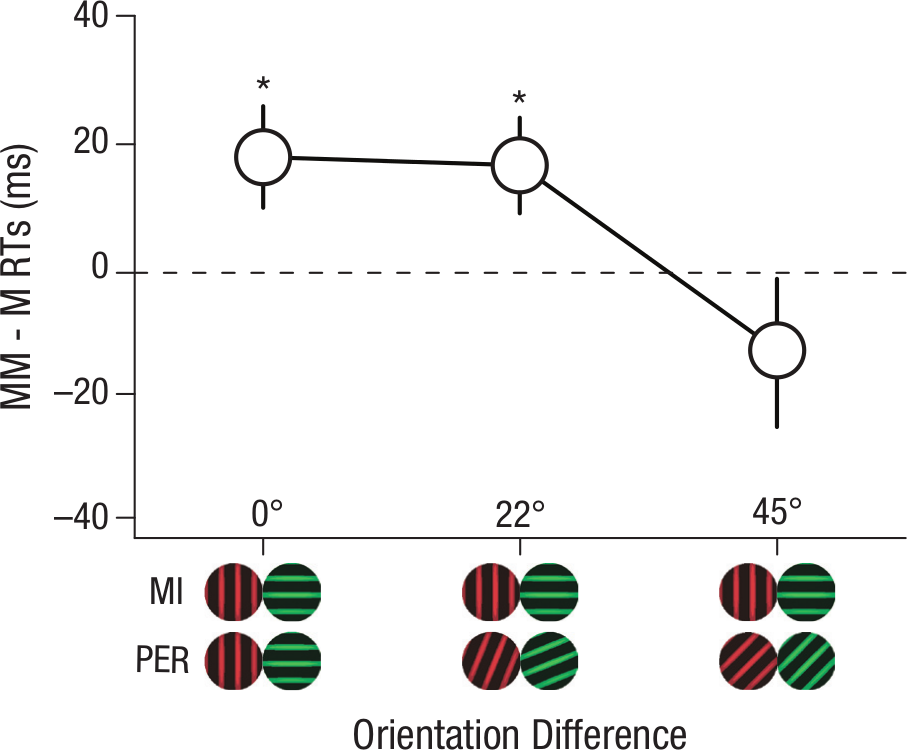
Reaction time difference between mismatched and matched pairs using Gabors rotated 0°, 22°, and 45° from those used in the associative learning phase.
We then sought to determine if a significant amount of associative learning had occurred in any of the three orientation conditions. For each test pattern orientation, the matched and mismatched reaction times were compared in the same manner as the previous experiments. Reaction times for 0° rotated Gabors were significantly faster for matched pairs (M = 640.98 ms, SEM = 13.52 ms) than mismatched pairs (M = 659.03 ms, SEM = 16.51 ms), t(14) = 2.29, p = .038, η2 = .27, providing yet another replication of our findings from Experiments 2 and 3. Reaction times for 22° rotated Gabors were also significantly faster for matched pairs (M = 646.86 ms, SEM = 12.13 ms) than mismatched pairs (M = 663.62 ms, SEM = 16.45 ms), t(14) = 2.26, p = .041, η2 = .27, indicating a degree of generalization between similarly oriented Gabors. However, when the patterns were rotated 45°, the reaction times for matched pairs (M = 656.66 ms, SEM = 17.26 ms) were not significantly different from mismatched pairs (M = 644.60 ms, SEM = 13.13 ms), t(14) = 1.03, p = .322. In addition, these data also show that the Gabor pattern colors were unable to independently elicit the learned emotion, as only the spatial orientation changed and not the colors. This result supports that of Experiment 3 (achromatic stimuli), suggesting that associative learning with mental imagery can be feature specific in the domain of vision.
General Discussion
We have shown evidence that associative learning can occur for voluntary mental imagery. Perceptual Gabor patterns were found to elicit a measurable emotional response after voluntary mental imagery of the same Gabors was subjected to associative learning. Hence, by definition, our dependent measure includes generalization from an internally generated mental image to sensory perception. The observed effects were substantial across all studies, similar to previous findings of associative learning with perceptual stimuli (De Houwer et al., 2001; Martin & Levey, 1978; Milad et al., 2006).
Our results cannot be accounted for by associative learning involving the imagery cues, as opposed to the imagery content. We tested an alternative hypothesis consisting of associative learning for the imagery cues in conjunction with the test phase Gabor patterns inducing recall of these imagery letter cues. This compound-alternative hypothesis was not supported by our two control experiments.
If this nonimagery hypothesis were valid, then the imagery cues should be able to prime the emotion- evoking IAPS images, and the perceptual Gabors should be able to prime the imagery cues. However, we found that the imagery cues did not prime the emotion-evoking photographs (Figure 2d), and the perceptual Gabor patterns did not prime the imagery cues (Figure 2e), thus ruling out this alternative account. In addition, the failure of this alternative account may be explained by the temporal relationships between the stimuli. It has been shown that the strength of a learned association between two stimuli is negatively correlated with the temporal distance and number of stimuli presented between them (Balsam, Drew, & Gallistel, 2010). Hence, the lack of associative learning for the letter cues may simply be due to the large amount of time between the cues and the emotion-evoking photographs.
In primates, a strong candidate for the neural mechanism of associative learning is the inferior temporal (IT) cortex (Jones, 1974; Meyer & Yates, 1955; Miyashita, 1993; Sakai & Miyashita, 1991; Yakovlev, Fusi, Berman, & Zohary, 1998). Both single cell physiology and human neuropsychological studies demonstrating that damage to the IT cortex can lead to impairments in visual recognition and learning together suggest associative learning occurs in high-level IT cortex. Here, we have demonstrated that the content of voluntary mental imagery can undergo a form of associative learning that is selective for spatial orientation. These findings support previous research showing orientation specificity for mental imagery (Pearson et al., 2008), though the present findings show a broader degree of orientation tuning than previously reported. Orientation specificity has been shown to be a hallmark of early levels of processing in the visual cortex (Blakemore & Campbell, 1969; Hubel & Wiesel, 1968; Ling, Pearson, & Blake, 2009). Our findings show that associative learning with voluntary mental imagery is sensitive to spatial orientation, indicating the involvement of these early visual processing areas. This is a rather surprising finding as associative learning is thought to involve processing within higher-level areas (Jones, 1974; Meyer & Yates, 1955; Miyashita, 1993; Sakai & Miyashita, 1991; Yakovlev et al., 1998).
Mental imagery and perception are similar processes
These results provide further evidence that mental imagery and sensory perception can involve overlapping mechanisms. However, imagery is an inherently private affair, making it difficult to study using traditional research methods, which has resulted in many compelling criticisms of the research. The founder of the behaviorism movement, John B. Watson (1913), argued that mental imagery plays no role in thought or behavior—the generation of mental images simply does not occur. This idea was revisited by B. F. Skinner (1977), who claimed that everything required to explain human behavior can be found within the environment. Zenon Pylyshyn (1973, 1981) conceded that mental imagery can occur but argued that the term is misleading, as what people refer to as imagery is actually the generation of verbal descriptions and not pictorial representations.
With the advent of modern brain imagining techniques it has become possible to observe the neural processes involved in mental imagery tasks. Studies using functional magnetic resonance imaging (Ganis, Thompson, & Kosslyn, 2004; Knauff, Kassubek, Mulack, & Greenlee, 2000; Mellet, Petit, Mazoyer, Denis, & Tzourio, 1998; O’Craven & Kanwisher, 2000), single cell neurophysiology (Kreiman, Koch, & Fried, 2000), and transcranial magnetic stimulation (Ganis, Keenan, Kosslyn, & Pascual-Leone, 2000; Sparing et al., 2002) have provided a wealth of neural evidence that common brain areas can be used for both visual imagery and visual perception. In addition, individuals with cortical damage specific to perceptual areas tend to experience deficits in the ability to generate mental imagery (Barton & Cherkasova, 2003; Farah, 1984; Farah, Levine, & Calvanio, 1988), but this is not always the case (Bridge, Harrold, Holmes, Stokes, & Kennard, 2012). For normal observers, perception has been shown to interfere with concurrent mental image generation (Pearson et al., 2008; Sherwood & Pearson, 2010), and the content of mental imagery has been shown to bias subsequent visual perception in a manner specific to early sensory processes (Pearson et al., 2008). The current study builds on this literature by showing that mental imagery can be used in place of perceptual stimuli in an associative learning task, and that this learning is specific to the spatial orientation of the imagined stimuli. Taken together, these studies provide compelling evidence that mental imagery can indeed be pictorial.
Clinical relevance
Fear learning is a type of associative learning wherein a neutral stimulus will come to display fear-indicant responses after being paired with an aversive stimulus (Öhman & Mineka, 2001). Influential models for numerous mental illnesses tend to include fear learning components, such as posttraumatic stress disorder, obsessive- compulsive disorder, and various phobias (Büchel & Dolan, 2000; Lissek et al., 2005; Milad et al., 2006; Rauch et al., 2006; Rosen & Schulkin, 1998; Watson & Rayner, 1920). These disorders often involve instances of intrusive and harmful mental imagery (Hackmann, Clark, & McManus, 2000; Hirsch & Holmes, 2007), which seem to cause or amplify the distress experienced (Hirsch et al., 2006; Holmes et al., 2008; Holmes & Mathews, 2010; Pitman et al., 1990). CBT and other treatments for anxiety often include mental imagery components (Holmes & Mathews, 2010), such as imaginal exposure or imagery rescripting (Arntz et al., 2007). Unfortunately the underlying mechanisms of these imagery-based treatments are not fully understood.
Our findings show that the content of voluntary mental imagery can gain new (emotional) meaning through association with emotion-evoking stimuli. Hence, the present data suggest that the efficacy of these imagery-based treatments can be explained in part by associative learning theory. These treatments involve the manipulation of mental imagery content to alter learned associations. In imaginal exposure, the distressing mental images are repeatedly generated in a nonaversive setting, resulting in the gradual decrease of anxiety symptoms (Tarrier et al., 1999). In addition, in imagery rescripting the patient actively attempts to change the content of their distressing mental images by associating them with pleasant or neutral mental images (Smucker et al., 1995). The orientation specificity observed in the current study suggests that the associative learning components of these treatments might involve activity within low-level sensory areas of the brain.
The effectiveness of imagery-based treatments for anxiety might actually be enhanced by focusing on the low-level sensory aspects of the distressing imagery symptoms as opposed to the semantic content. The orientation specificity observed in the current study supports this suggestion by providing evidence that it is the sensory content of the voluntary imagery that is driving the mechanisms of learning, which likely involves low-level sensory areas of the brain. Additional support comes from the recent demonstration that engaging low-level sensory systems, not high-level semantic systems, can help prevent the formation of distressing flashbacks after experiencing trauma (Holmes, James, Kilford, & Deeprose, 2010). Though this particular study did not use mental imagery stimuli, the current study and many others have shown that visual imagery can act in a similar manner to visual perception (Kosslyn et al., 2001; Pearson et al., 2008). Therefore, it seems likely that highly effective treatment methods could be developed that focus on engaging low-level sensory systems.
Conclusion
We have shown that associative learning can occur between a voluntarily generated mental image and the emotional content of a photograph, providing further support for the pictorial theory of mental imagery. For the first time we have demonstrated that a stimulus not physically present can become emotionally charged after its voluntarily generated mental image is paired with an emotion-evoking photograph. Associative links will form between these sensory representations even though they are never physically presented together in the world. This novel finding adds to the previous research on associative learning for mental imagery (Dadds et al., 1997) by showing that mental images can be effectively used as conditioned stimuli, thus providing a unique contribution to the rapidly growing field of learning (Jeannerod, 1995; Lissek et al., 2005; Sasaki et al., 2010; Shibata et al., 2011).
It will be exciting for future work to compare the relative strength of associative learning with voluntary imagery and normal perception. There have been claims that triggering memories perceptually prior to learning regimes can boost memory reconsolidation, which in turn modulates the effects of associative learning (Monfils, Cowansage, Klann, & LeDoux, 2009; Schiller et al., 2009; Schiller & Phelps, 2011). If simply activating memories in this manner has the power to change the outcome of learning, it will be of particular interest to see if mental imagery, a process that inherently involves memory retrieval, initiates similar or even stronger reconsolidation of memories. The prospect that mental imagery could be utilized to simultaneously target memories and make them malleable and more receptive to long-term change is an exciting possibility as many psychological disorders are rooted in traumatic memories. Such breakthroughs would have a profound impact on the outcome of clinical practice that utilizes extinction-like paradigms.
Our findings suggest that the sensory components of imagery are instrumental to its role in associative learning. On a general level this has some interesting implications. All the characteristics known to modulate imagery should logically be applicable to regulating associative learning with mental imagery. For example, research has shown that uniform passive perception, simply being in a bright environment, disrupts the generation of mental images (Keogh & Pearson, 2011; Pearson et al., 2008; Sherwood & Pearson, 2010); the brighter the environment, the greater the attenuation of imagery. It will be intriguing for future research to investigate the role of the perceptual environment in associative learning with mental imagery, and how this extends to clinical practice. For example, is imagery rescripting more effective in dark environments or with the eyes closed because imagery is stronger?
In summary, the current study demonstrates that voluntary mental imagery can drive the mechanisms of associative learning. This provides strong laboratory support for clinical practice that utilizes both mental imagery and regimes based on associative learning.
Footnotes
Acknowledgements
We thank Richard Bryant and Gabrielle Weidermann for helpful discussions and Franco Caramia for programming assistance.
Declaration of Conflicting Interests
The authors declared that they had no conflicts of interest with respect to their authorship or the publication of this article.
Funding
This work was supported by National Health and Medical Research Council Project Grants APP1024800 and APP1046198 and a Career Development Fellowship APP1049596 held by JP.