
Abstract
Biased attention toward negative stimuli is a known vulnerability for affective psychopathology. However, factors that contribute to the development of this cognitive bias are largely unknown. Variation within the serotonin transporter gene (i.e., 5-HTTLPR) is associated with increased susceptibility to environmental influence and biased processing of negative stimuli. Using a passive viewing eye-tracking paradigm, this study examined gaze fixation for emotion stimuli in 91 U.S. Army soldiers before and after deployment to Iraq. In addition, participants underwent genetic assay and provided in situ measures of war zone stress exposure. 5-HTTLPR short allele homozygotes were more likely than other genotype groups to develop a gaze bias toward negative stimuli as a function of increasing war zone stress, even when controlling for postdeployment posttraumatic stress disorder and depression severity. Short allele homozygotes appear especially sensitive to environmental influence, which likely contributes to the development of cognitive vulnerability to anxiety and mood disorders.
Soldiers returning from a war zone environment are at high risk for the development of psychopathology, particularly anxiety and mood disorders (Boscarino, 1995; Fontana & Rosenheck, 1994; Foy, Sipprelle, Rueger, & Carroll, 1984; Kessler, Sonnega, Bromet, Hughes, & Nelson, 1995; Mellman, Randolph, Brawman-Mintzer, Flores, & Milanes, 1992; Seal, Bertenthal, Miner, Sen, & Marmar, 2007). Approximately 28% of U.S. veterans serving in Operation Enduring Freedom or Operation Iraqi Freedom (OIF) reported clinically significant levels of depression or anxiety following deployment (Hoge et al., 2004). Prevalence of clinical depression in veterans deployed to a war zone is nearly 50% higher than in nondeployed soldiers (Hoge et al., 2004), whereas incidence of posttraumatic stress disorder (PTSD) jumps more than 300%, from 2.3% in nondeployed personnel to 7.6% for soldiers exposed to war zone stress (Smith et al., 2008). These figures likely underestimate the degree of impairment for veterans, considering that roughly 60% of veterans who meet criteria for mental illness do not seek mental health care (Hoge et al., 2004).
Cognitive theorists suggest that altered cognitive processing of emotion stimuli serves as a putative foundation for the onset and maintenance of such disorders (Bar-Haim et al., 2010; Bar-Haim, Lamy, Pergamin, Bakermans-Kranenburg, & Van IJzendoorn, 2007; Beck & Clark, 1997; Eysenck, Derakshan, Santos, & Calvo, 2007; Mathews & MacLeod, 1994). Specifically, biased attention for mood-congruent negative stimuli (relative to neutral or positive stimuli) has been consistently observed across anxious and depressed populations, yet does not appear in nonclinical control groups (Bar-Haim et al., 2007; Bradley, Mogg, & Lee, 1997; Bradley, Mogg, White, Adams, & Bono, 2010; Bryant & Harvey, 1997; Koster, De Raedt, Goeleven, Franck, & Crombez, 2005; Mogg, Bradley, & Williams, 1995; Vythilingam et al., 2007).
More specifically, in anxiety disorders, biased attention can have divergent influences depending on context. In stressful situations, gaze bias toward fear-related stimuli can be adaptive, whereas gaze bias away from fear-related stimuli can predict future onset of anxiety disorders (Bar-Haim et al., 2010; Wald et al., 2011). However, in a nonstressful environment, attention biased toward fear-related stimuli appears to represent a vulnerability factor for anxiety disorders. A meta-analysis involving 4,031 participants across 172 studies found bias toward fear-related stimuli to be consistently observed in patients suffering from PTSD, generalized anxiety disorder, panic disorder, obsessive-compulsive disorder, social phobia, and specific phobia, as well as those who self-report with high trait anxiety (Bar-Haim et al., 2007). There were no significant differences in attentional bias across disorder type, experimental paradigms (e.g., emotional Stroop task, the dot-probe task, and emotional spatial cuing task), or levels of conscious awareness (e.g., subliminal or supraliminal presentation; Bar-Haim et al., 2007). Of importance, biased attention has been observed in veterans with PTSD following service in Iraq, who show increased gaze fixation and greater pupillary response when viewing fear-related images (Kimble, Fleming, Bandy, Kim, & Zambetti, 2010). Biased attention toward fear-related stimuli appears to be a robust and generalized finding across numerous domains of anxiety.
In addition, for patients diagnosed with mood disorders such as major depressive disorder (MDD), attention appears to be consistently biased toward sad stimuli (Koster et al., 2005), although the findings are, in general, less consistent than those in anxiety disorders (Mogg & Bradley, 2005). Unlike anxiety disorders, where attentional biases are observed at 500 ms or less, biases in MDD generally appear in later stages of processing, often between 500 and 1,000 ms (Caseras, Garner, Bradley, & Mogg, 2007; Donaldson, Lam, & Mathews, 2007; Gotlib, Krasnoperova, Yue, & Joormann, 2004).
It is theorized that bias in MDD is not characterized by rapid orientation toward negative stimuli (as with anxiety), but rather with delayed disengagement away from negative stimuli (Gotlib & Joormann, 2010). This hypothesis is supported by visual search results in which MDD patients were no more effective at detecting negative words, but were more easily distracted once they did attend to negative words (Rinck & Becker, 2005). Furthermore, eye-tracking studies of depressed individuals have consistently shown longer gaze fixation toward sad stimuli compared to other emotional valences, which supports the hypothesis of delayed disengagement (Caseras et al., 2007; Eizenman et al., 2003; Kellough, Beevers, Ellis, & Wells, 2008). Although the underlying cognitive processes may differ, both depressed and anxious individuals demonstrate a significant and reliable attentional bias that increases exposure to negative stimuli.
In addition to co-occurring with mental illness, there is evidence to suggest that biased attention for negative stimuli may play a key role in the formation and maintenance of anxiety and mood disorders. Attentional biases toward negative stimuli are believed to stem from the activation of specific negative cognitive schemas (Beck, 1967, 2008; Clark, Beck, & Alford, 1999; Foa, Feske, Murdock, Kozak, & McCarthy, 1991; Foa, Steketee, & Rothbaum, 1989). When these schemas are activated, it likely interferes with adaptive cognitive mechanisms that would otherwise be used to integrate contextual information or facilitate reappraisal of incoming stimuli. As a result, salience for the negative stimuli is enhanced and coping may be attenuated (Beck, 2008; Clark et al., 1999; Foa et al., 1989; Foa et al., 1991).
Recent studies have identified the role of attentional biases in predicting psychopathology, particularly in the context of environmental stressors. Increased gaze toward negative stimuli has been shown to interact with life stress to prospectively predict increases in depression symptoms (Beevers & Carver, 2003). More specifically, prospective research in OIF soldiers has found that negative attentional bias moderates the relationship between war zone stress exposure and worsening of clinical symptoms reported during deployment (Beevers, Lee, Wells, Ellis, & Telch, 2011). For those exposed to war-related threat (including civilians), increases in the magnitude of biased attention predicted increases in overall psychological distress, including symptoms of PTSD, depression, and anxiety (Bar-Haim et al., 2010). These findings suggest that biased attention is more than a symptom or scar of mental illness, but rather is an indicator of cognitive vulnerability to anxiety and depression (De Raedt & Koster, 2010; Foa et al., 1989; Ingram, Bernet, & McLaughlin, 1994).
The presence and severity of attentional bias for negative stimuli may have a direct and possibly causal role in the etiology of emotional disorders. However, the etiological processes that contribute to the acquisition of attentional biases are not firmly understood. Childhood maltreatment appears to predict the development of attentional biases in children (Pine et al., 2005) as well as adults (Fani, Bradley-Davino, Ressler, & McClure-Tone, 2010; Gibb, Schofield, & Coles, 2009). In addition, negative peer experiences may predict attentional bias to stimuli associated with social threat (Haddad, Lissek, Pine, & Lau, 2010). However, research investigating the etiology of attentional biases is generally lacking.
In this study, we examined the association between level of exposure to war zone stressors and pre- to postdeployment changes in attention for negative stimuli. Soldiers who show increased gaze toward negative stimuli following deployment may be at increased risk for subsequent anxiety and depression symptoms (Beevers, Lee, et al., 2011). Greater understanding of the factors that moderate this cognitive vulnerability could help mitigate the risk of these negative outcomes.
Of particular interest is whether allelic differences in the serotonin transporter gene (SLC6A4) promoter polymorphism (5-HTTLPR), a region known to moderate cognitive vulnerability in anxiety and depression (Beevers, Scott, McGeary, & McGeary, 2009; Caspi et al., 2003; Schinka, Busch, & Robichaux-Keene, 2004; Xie et al., 2009), interact with the harmful effects of stressful environments. The 5-HTT gene, which regulates serotonin reuptake, has two common variants in the promoter region: a short allele (S) and a long allele (Heils et al., 1996). The presence of the short allele is associated with decreased available serotonin transporter and decreased cellular serotonin uptake (Heils et al., 1996; Lesch et al., 1996).
The 5-HTTLPR short allele has been associated with increased susceptibility to environmental factors (Belsky & Pluess, 2009). In the absence of life stress, this genetic plasticity predicts increased life satisfaction, decreased depression, and decreased neuroticism (Belsky et al., 2009; Kuepper et al., 2012; Taylor et al., 2006). However, the short allele has also been shown to predict negative cognitive, behavioral, and physiological outcomes for those experiencing significant life stress (Caspi et al., 2003; Xie et al., 2009), including acute, life-threatening situations (Mercer et al., 2012; Telch et al., 2013). Specifically, the short allele has been associated with greater attentional bias as a function of environmental influences. This has been shown in experimental manipulations, where attention bias modification yielded greater bias toward negative stimuli in short allele carriers (Fox, Zougkou, Ridgewell, & Garner, 2011). It is consistent with a recent meta-analysis (Pergamin-Hight, Bakermans-Kranenburg, Van IJzendoorn, & Bar-Haim, 2012) that showed that short allele homozygotes display significantly longer attentional fixation toward negative stimuli than other 5-HTTLPR genotype groups (see also Fox, Ridgewell, & Ashwin, 2009; Osinsky et al., 2008; Pérez-Edgar et al., 2010).
Based on these findings, we hypothesized that the soldiers’ 5-HTTLPR genotype would moderate the association between war zone stress exposure and pre- to postdeployment increases in attention for negative stimuli. In particular, we expected homozygous short participants who experience more war zone stress to show greater change in attention toward negative stimuli (e.g., sad and fearful faces) upon returning from Iraq. This finding would help explain the close relationship between war zone stress exposure and negative outcomes, and would shed light on mechanisms that underlie the development of cognitive biases that confer risk for emotional disorders.
Methods and Materials
Participants
Participants were 91 U.S. Army soldiers with no prior war zone experience who were scheduled for deployment to Iraq from Fort Hood, Texas. Participants were predominantly males in their early to mid-20s (80 male, 11 female, with a mean age of 23.9 years old). The military rank for the majority of participants was either private first class (33%) or private E-2 (29%). Participants were deployed for an average of 400 days, and time between predeployment and postdeployment assessments was 515 days. Racial breakdown was as follows: 70.3% Caucasian, 11.0% African American, 9.9% American Indian/Alaska Native, 4.4% mixed race, and 2.2% each for Asian and Native Hawaiian/Pacific Islander. There were no significant differences in genotype distribution as a function of race (Table 1). In addition, all of the final analyses were consistent with analyses run in separate subpopulations consisting of Caucasian participants (n = 64) and non-Caucasian participants (n = 27), though subpopulations of this size lack the power to merit further discussion. Because of the lack of group differences, race will not be discussed further.
Distribution of 5-HTTLPR Genotype as a Function of Race

Note: This table presents the frequency breakdown across race for each of the three genotype variants—homozygous long allele (LL), heterozygous (SL), and homozygous short allele (SS). There was no significant difference in genotype frequency across race
Participants were selected from a sample of 184 soldiers who provided informed consent. Of the excluded participants, 6 were not deployed to Iraq and 1 withdrew consent. Of the remaining 177, genetic data were not available for 31 soldiers, 17 soldiers failed to come in for postdeployment assessment, and eye tracking was unsuccessful for 38 soldiers (i.e., fewer than 7 of the 8 eye-tracking trials with 60% or more data points available on either assessment). The participants who were excluded from the final sample did not differ in age (p = .37) or stress exposure (p = .65), nor did they differ on in situ measures of PTSD (p = .80) or depression symptoms (p = .80) compared to those who completed all study assessments.
Assessments
War zone stressors
War zone stress was recorded using the Combat Experience Log (CEL). The CEL is a Web-based system that allows soldiers to select from a list of 18 well-validated war zone stressors (e.g., received hostile incoming fire, observed homes or villages being destroyed, received bad news from home; Lee, Goudarzi, Baldwin, Rosenfield, & Telch, 2011). These stressors were chosen from a modified version of the Deployment Risk and Resilience Inventory (King, King, Vogt, Knight, & Samper, 2006). Soldiers could list up to two unique stressors not covered by the 18 standard items. Because of the nature of military service and intermittent Internet access during active combat, frequency of CEL completion was highly variable. The CEL prompts participants to log the number of war zone stressors they experienced since their previous CEL assessment. Soldiers could therefore be reporting across different lengths of time, depending on their access to the CEL system. To control for this variability in reporting time, the sum total of war zone stressors endorsed across all CELs was used to estimate level of war zone stress exposure during deployment. Number of CEL assessments was entered as a covariate in all analyses.
PTSD symptoms
The 17-item military version of the PTSD Checklist (PCL) is a self-report scale designed to assess the presence and severity of PTSD symptoms in response to stressful military experiences (Weathers, Litz, Herman, Huska, & Keane, 1993). It is used for screening, diagnosing, and monitoring symptom change for active service members and veterans of military service. Questions are divided into three domains: re-experiencing, avoidance/psychic numbing, and hyperarousal. A total score of 50 is commonly used as a cutoff for PTSD diagnosis (Blanchard, Jones-Alexander, Buckley, & Forneris, 1996; Perrin et al., 2007). The PCL was administered before, during, and after deployment.
Depression symptoms
The Center for Epidemiologic Studies–Depression Scale (CES-D) is a 20-item, self-report scale designed to assess the presence and severity of depressive symptoms over the previous week (Radloff, 1977). Questions cover a range of potential symptoms, including sad mood, self-worth, motivation, and appetite. The CES-D was administered before, during, and after deployment.
Genetic assessment
Assays of 5-HTTLPR were conducted by sampling epithelial cells collected via passive drool of whole saliva. Previously reported methods were used to isolate genomic DNA from buccal cells (Freeman et al., 1997; Lench, Stanier, & Williamson, 1988) and for assaying the 5-HTTLPR (Pooley, Houston, Hawton, & Harrison, 2003). The primer sequences are forward 5′-GCG TTG CCG CTC TGA ATG C-3′ and reverse 5′ GGA CTG AGC TGG ACA ACC AC-3′. Genotyping yielded three distinct SNP iterations: S, LA, and LG. As per standard convention (Hu et al., 2005; Zalsman et al., 2006), LG was treated as equivalent to S. Allelic frequency was in Hardy-Weinberg equilibrium (Pearson χ2 = 1.20, df = 1, p = .27). Participants were subdivided into one of three groups depending on their genotype: homozygous long (LL; n = 24), heterozygous (SL; n = 50), and homozygous short (SS; n = 17).
Eye-tracking paradigm
Participants completed a passive viewing task consisting of simultaneous presentation of four images selected from the Pictures of Facial Affect (Ekman & Friesen, 1976). Stimuli were faces from six male and six female actors. Each stimulus consisted of an actor with one of four possible facial expressions: happy, sad, fear, and neutral. Participants viewed four simultaneous images of a single actor displaying all four emotions. Facial expressions were randomly assigned to each quadrant with equal frequency.
Participants underwent eight critical trials (using happy, sad, fear, or neutral faces) and four filler trials (using faces from other emotion categories). Each trial began with a central fixation cross, followed by a 30-s presentation of the face stimuli. Gaze orientation was assessed using a remote optics eye-tracking system model R6 from Applied Science Laboratories (Bedford, MA, USA). Coordinates for gaze were sampled every 16.7 ms (60Hz), producing 1,796 gaze location measurements for each 30-s trial. Eye movements that were stable for more than 100 ms within 1° of visual angle were classified as fixation. In critical trials, total duration of fixation periods for each valence was summed. Total fixation duration as a measure of sustained attention has been employed repeatedly in various domains of attention research (e.g., Green, Williams, & Davidson, 2003; Horley, Williams, Gonsalvez, & Gordon, 2004; LaBar, Mesulam, Gitelman, & Weintraub, 2000). Within each valence, total fixation duration in predeployment trials was subtracted from total fixation duration from postdeployment trials to yield change in fixation duration, which serves as our dependent measure.
This task represents one of many ways to operationalize attentional bias. Others, such as the dot probe or the emotional Stroop task, require active task involvement and assess sustained attention by examining behavioral response time. These tasks can provide precise measurement of acute attentional biases and are capable of detecting subtle differences in response time. The passive viewing task complements these other reaction time approaches in that it may capture more sustained and effortful viewing patterns of participants. Sustained attention for affective stimuli has been identified as an important marker of affect regulation (Todd, Cunningham, Anderson, & Thompson, 2012). Although the parameters and scale of the tasks differ considerably (passive viewing, for example, is measured in seconds, whereas dot probe is measured in milliseconds), the use of eye-tracking methodology also avoids many of the confounds inherent to reaction time data (Hermans, Vansteenwegen, & Eelen, 1999).
Procedure
Approximately 3 months prior to deployment, participants underwent a full day of assessment at the University of Texas at Austin. Following informed consent, participants provided demographic information, completed questionnaires, and were interviewed for the presence of diagnoses based on the fourth edition, text revision of the Diagnostic and Statistical Manual of Mental Disorders (DSM-IV-TR; American Psychiatric Association, 2000). Study procedures included a number of assessments that are not the focus of the current study but are reported elsewhere (Beevers, Lee, et al., 2011; Beevers, Marti, et al., 2011; Josephs et al., 2012; Telch et al., 2013; Telch, Rosenfield, Lee, & Pai, 2012).
Eye tracking generally occurred in the early afternoon. Participants were calibrated to the device and instructed to look at a fixation cross prior to each trial to standardize starting location. Participants were told to view images that seemed interesting, as if they were watching television or viewing a photo album.
Self-report assessments, such as the CEL, PCL, and CES-D, were obtained before, during, and after deployment via a Web-based system developed by study personnel (Lee et al., 2011). Upon return from Iraq, participants were invited to return for a postdeployment assessment, which followed the same overall procedures as the predeployment assessment.
Statistical analyses
The primary analysis examined change in total fixation duration from pre- to postdeployment within each emotional valence. 1 Nonparametric robust regression models were used to examine the influence of 5-HTTLPR genetic variation and total war zone stress on change in fixation duration for emotion stimuli. Nonparametric robust regression was conducted using Stata 12 (StataCorp, College Station, TX, USA) and was selected over standard multiple regression because of nonnormative distribution of residuals resulting from ordinary least squares regression. The technique eliminates any observation with Cook’s D greater than 1 and then utilizes iterative Huber and biweight estimation to converge on the best model. All results presented from robust regression models were consistent with results from standard regression techniques. For regression models, the 5-HTTLPR variable was dummy coded to form three variables. To explore the influence of genotype, models either used the LL group for comparison (i.e., SS vs. LL and SL vs. LL) or used the SL group for comparison (i.e., SS vs. SL). Subsequent analyses controlled for each participant’s number of CEL assessments.
Results
Clinical symptoms
Among the soldiers included in this sample, postdeployment PTSD symptom severity, measured using the PCL, was relatively few in number (range = 17–59, M = 25.00, SD = 8.67), with an average PCL increase of 7.34 (SD = 9.39) from pre- to postdeployment. Two individuals exceeded the cutoff for clinically significant PTSD symptoms at postdeployment, compared to one at predeployment. Postdeployment depression symptom severity, measured using the CES-D, was also few in number (range = 0–47, M = 11.41, SD = 9.09), with an average CES-D increase of 0.51 (SD = 8.92) from pre- to postdeployment. A total of 24 individuals exceeded the cutoff for clinically significant depression symptoms at postdeployment, compared to 23 at predeployment.
Prediction of change in fixation duration
Mean fixation duration within each genotype group, time point, and emotional valence can be found in Table 2. Two sample mean comparison t tests were used to compare pre- and postdeployment fixation duration. There were no significant differences across time points for sad stimuli, t(90) = −0.20, p = .84, fear stimuli, t(90) = −0.63, p = .53, happy stimuli, t(90) = −1.71, p = .09, and neutral stimuli, t(90) = 0.66, p = .51.
Gaze Fixation (in seconds) Across Genotype, Time Point, and Stimuli Valence
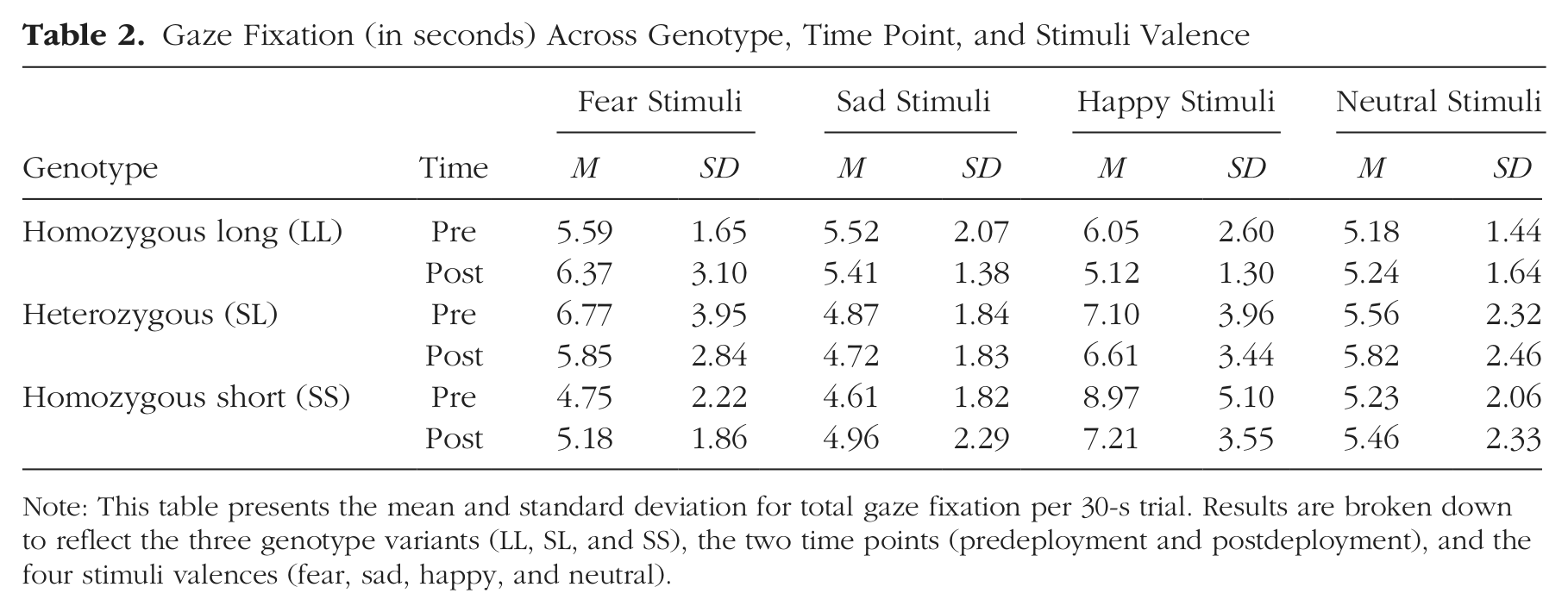
Note: This table presents the mean and standard deviation for total gaze fixation per 30-s trial. Results are broken down to reflect the three genotype variants (LL, SL, and SS), the two time points (predeployment and postdeployment), and the four stimuli valences (fear, sad, happy, and neutral).
Main effects
War zone stress exposure and 5-HTTLPR genotype were used to predict change in total fixation duration for each valence category. For war zone stress exposure, there were no significant main effects for fear, R2 = .03, F(1, 86) = 0.06, p = .80, or sad stimuli, R2 = .02, F(1, 86) = 0.43, p = .52. However, war zone stress did significantly predict increased attention toward neutral stimuli, R2 = .06, F(1, 86) = 6.22, p = .01, and decreased attention toward happy stimuli, R2 = .07, F(1, 86) = 7.62, p < .01. There were no significant main effects for 5-HTTLPR genotype across stimulus valences: fear, R2 = .03, F(2, 86) = 1.00, p = .37; sad, R2 = .02, F(2, 86) = 0.44, p = .64; happy, R2 = .07, F(2, 86) = 1.75, p = .18; and neutral, R2 = .06, F(2, 86) = 0.15, p = .86.
5-HTTLPR × war zone stress interaction effects
There was a significant gene-by-environment interaction for change in total fixation duration for fear stimuli. Specifically, increases in war zone stress exposure predicted greater increases in total fixation time for fear stimuli at postdeployment in the SS group, but not in the SL or LL groups, R2 = .07, F(2, 85) = 3.65, p = .03 (Fig. 1). The influence of war zone stress on attention for fear stimuli was significantly greater in the SS group compared to the LL (β = 0.18, SE = 0.07, t = 2.65, r = .28, p = .01) and SL groups (β = 0.15, SE = 0.06, t = 2.38, r = .25, p = .02). Exposure to war zone stress leads to increased postdeployment fixation duration for fear stimuli among the SS group compared to the LL and SL groups. The association between war zone stress and attention for fear stimuli did not differ for the SL and LL groups (β = 0.04, SE = 0.05, t = 0.79, r = .09, p = .43).
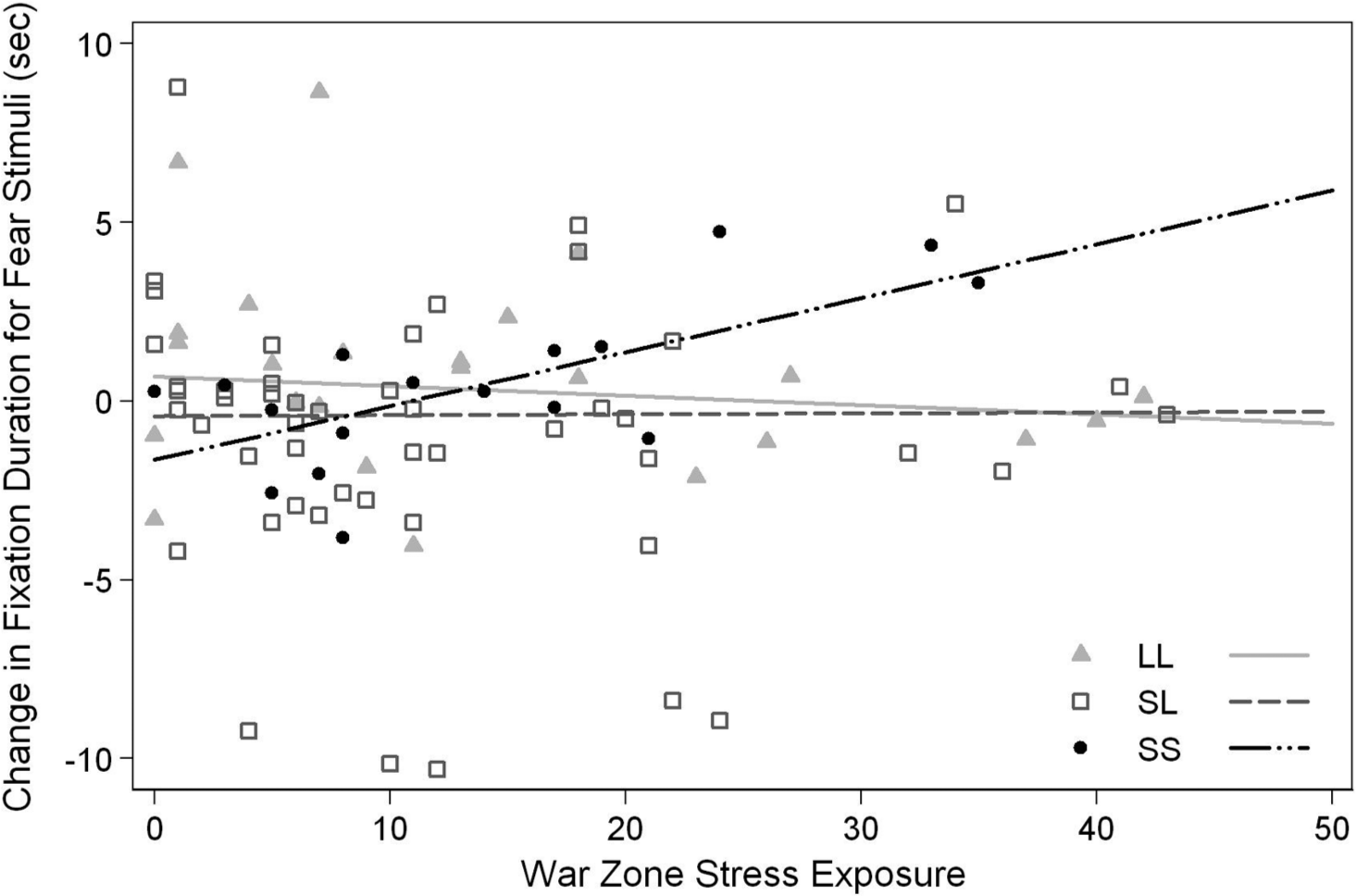
Change in attention toward fear stimuli as a function of war zone stress.
The same gene-by-environment interaction was observed for sad stimuli. Specifically, increases in war zone stress exposure predicted greater increases in total fixation time for sad stimuli at postdeployment in the SS group, but not in the SL or LL groups, R2 = .07, F(2, 85) = 3.42, p = .04 (Fig. 2). The influence of war zone stress on attention for sad stimuli was greater in the SS group compared to the LL (β = 0.12, SE = 0.05, t = 2.11, r = .22, p = .04) and SL groups (β = 0.13, SE = 0.05, t = 2.61, r = .27, p = .01). As with fear stimuli, the interaction between war zone stress and total fixation time for sad stimuli was not significantly different between the SL and LL groups (β = −0.01, SE = 0.04, t = 0.36, r = .04, p = .72). These findings suggest that total fixation duration for negative stimuli increases as war zone stress exposure accumulates in the SS group; however, attention for negative stimuli was relatively constant among SL and LL groups regardless of war zone stress exposure.
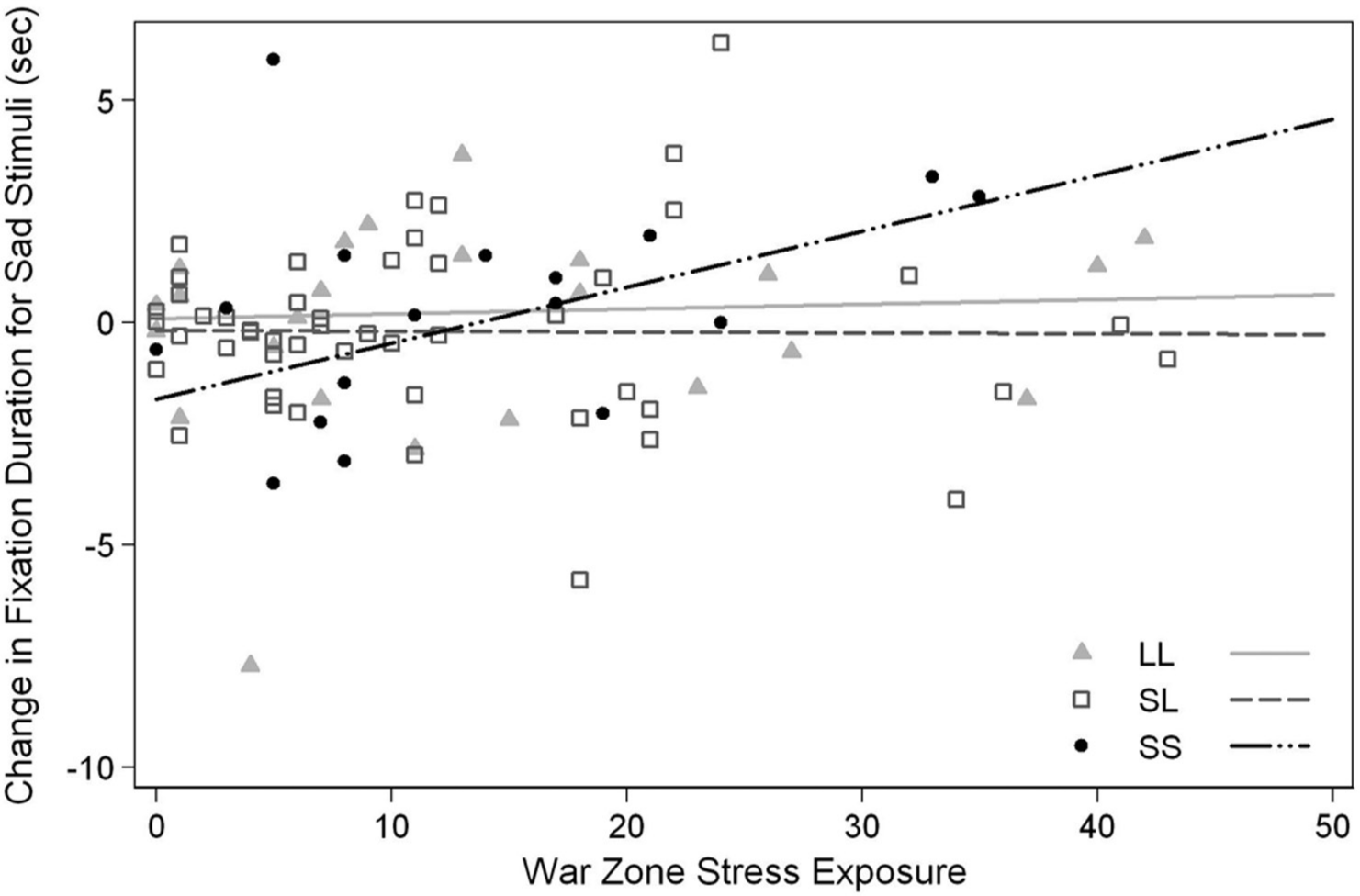
Change in attention toward sad stimuli as a function of war zone stress.
In contrast to the findings for negative stimuli, there was no significant gene-by-environment interaction for change in total fixation duration for happy or neutral stimuli in models comparing the SS and LL groups (happy: β = 0.09, SE = 0.09, t = 1.05, r = .11, p = .30; neutral: β = −0.05, SE = 0.07, t = 0.68, r = .07, p = .50) or models comparing the SS and SL groups (happy: β = 0.10, SE = 0.08, t = 1.28, r = .14, p = .20; neutral: β = −0.10, SE = 0.07, t = 1.50, r = .16, p = .14). Of importance, the gene-by-environment interaction observed for fear and sad stimuli in the SS group persisted when controlling for postdeployment PTSD symptoms (SS vs. LL: β = −0.18, SE = 0.07, t = 2.56, r = .27, p = .01; SS vs. SL: β = −0.15, SE = 0.06, t = 2.27, r = .24, p = .03) and depression symptoms (SS vs. LL: β = −0.11, SE = 0.05, t = 2.08, r = .22, p = .04; SS vs. SL: β = −0.13, SE = 0.05, t = 2.59, r = .27, p = .01), respectively. From this, we can infer that the increased gaze toward negative stimuli is independent of postdeployment PTSD and depression, and therefore cannot be accounted for by the presence of clinical symptoms.
Controlling for CEL assessment frequency
Participants completed an average of 7 CEL assessments during the course of their deployment (or approximately one every 57 days) and reported an average of 3.10 stressors per assessment (SD = 2.93). Given the variability in number of reports obtained from each soldier (M = 7.09, SD = 5.67), we controlled for number of completed CEL assessments as a main effect in the above analyses. However, we also examined whether number of completed CEL assessments moderated the interaction between genotype and war zone stress. The three-way interaction among genotype, total war zone stress, and number of CEL assessments was nonsignificant for change in fixation duration for fear stimuli, F(5, 79) = 0.49, p = .78, and change in fixation duration for sad stimuli, F(5, 79) = 0.78, p = .56. These models suggest that the main two-way interaction (5-HTTLPR by war zone stress) was not influenced by CEL frequency.
Discussion
We examined the interaction between 5-HTTLPR genotype and war zone stress exposure as it predicted change in attention for negative stimuli following deployment. We found that the impact of war zone stress on attention for emotion stimuli was strongest in short allele homozygotes. Specifically, the SS group showed greater increase in postdeployment gaze fixation toward negative stimuli (i.e., fearful and sad faces) as exposure to war zone stress increased during deployment in Iraq, whereas war zone stress was not linked to the increase of attention for any stimuli valence in the SL and LL groups. These findings are consistent with the outcome of a recent meta-analysis (Pergamin-Hight et al., 2012), but go further by longitudinally demonstrating that the development of a negative attentional bias depends on both 5-HTTLPR genotype and exposure to an adverse environment (i.e., war zone stress).
Biased attention for negative stimuli is uncommon in nonclinical populations compared to patients with anxiety and mood disorders (Bar-Haim et al., 2007; Bradley et al., 1997; Bradley et al., 2010; Bryant & Harvey, 1997; Koster et al., 2005; Mogg et al., 1995; Vythilingam et al., 2007). Although it is generally not thought to cause distress, biased attention can be viewed as a prodromal marker of future pathology. As such, these findings point to important factors that may contribute to the development of this cognitive vulnerability. Consistent with findings in other domains (Caspi et al., 2003; Mercer et al., 2012; Xie et al., 2009), the influence of significant environmental stressors appears to be more pathogenic for the SS group. Based on the role of attentional bias in the formation and maintenance of psychopathology (Beevers & Carver, 2003; Beevers, Lee, et al., 2011; De Raedt & Koster, 2010; Foa et al., 1989; Ingram et al., 1994), this interaction suggests that the SS group is at increased risk for emotional disorders following exposure to war zone stressors.
These findings are consistent with the diathesis-stress model of cognitive vulnerability, suggesting that the SS group is predisposed to respond poorly (e.g., with increased processing of negative stimuli) in response to negative environmental factors (e.g., war zone stress). These results are also consistent with the differential susceptibility perspective of Belsky and Pluess (2009), although it is difficult to support this connection considering that positive environmental factors were not measured. There are numerous positive factors associated with military service that could be viewed as adaptive or even protective. These include developing strong interpersonal relationships, team building, enhanced self-efficacy, and increased physical fitness. Further exploration into the influence of positive environmental factors would allow for a fuller understanding of how the environment combines with the 5-HTTLPR to enhance risk and resilience.
It may seem surprising that the interaction of 5-HTTLPR genotype and war zone stress can predict increased attention for negative stimuli independent of current clinical symptoms. The dissociation between attentional bias and clinical symptoms can be interpreted in several ways. It could be viewed as a credit to the psychological and social support available to veterans returning from deployment. There are a growing number of mental illness prevention treatments (Armfield, 1994; Feldner, Monson, & Friedman, 2007) and increased efforts to destigmatize psychological care in the armed forces (such as the U.S. Army’s Real Warriors Campaign). At-risk veterans have increased awareness for early signs of PTSD and depression symptoms and numerous mental health services available to them, which may prophylactically prevent symptom worsening.
Alternatively, another possibility is that increased attention for negative stimuli may be a residual effect of attention patterns that were adaptive and often used during deployment. In life-threatening situations, hypervigilance for threat-related stimuli can serve a protective function, allowing for quicker threat detection and increased time for decision making (Grillon et al., 2009; Robinson, Letkiewicz, Overstreet, Ernst, & Grillon, 2011). In threatening situations, it is attentional avoidance of negative stimuli that could be construed as maladaptive (Bar-Haim et al., 2010; Wald et al., 2011). However, characteristics that were adaptive in a war zone setting may be maladaptive upon returning from deployment. Because life-threatening situations are rare at home, sustained attention for fear-related stimuli is less likely to serve a protective role and more likely to increase the salience and awareness of negative elements in the environment.
Yet another alternative, however, is that the military trauma could manifest clinical symptoms that do not immediately follow the occurrence of war zone stress. Research has linked war zone trauma with delayed-onset PTSD (Gray, Bolton, & Litz, 2004; Prigerson, Maciejewski, & Rosenheck, 2001) and has estimated that 38.2% of all military PTSD cases fail to reach diagnostic threshold until 6 or more months following the trauma, compared to 15.3% in the civilian population (Andrews, Brewin, Philpott, & Stewart, 2007). Vulnerability factors may be present in soldiers, but lie dormant during periods of relative wellness. This cognitive vulnerability indicates that the SS group could be at greater risk of developing full-fledged emotional disorders following return from deployment, particularly once mental health services are less readily available. However, it will be impossible to test this hypothesis until additional prospective, longitudinal research that follows soldiers over time after deployment is completed.
Although speculative, the disparity between increased attention for negative stimuli and psychopathology is likely due to a combination of the previously discussed factors. Bias toward negative stimuli may be advantageous in a combat setting (Bar-Haim et al., 2010; Wald et al., 2011), and 5-HTTLPR short allele homozygotes may easily develop this bias in the military milieu. They may also be slower to develop symptoms of PTSD and MDD because of the comprehensive mental health services offered by the military and Veterans Administration. Ultimately, further research into the prevention and treatment of mental illness in returning veterans is clearly important and should be a high priority.
There are several features of this study design that merit further comment. First, the use of eye-tracking methodology to directly assess attention for emotion stimuli provides distinct advantages compared to indirect measures of attention that rely on behavioral reaction time. Second, the prospective design used in this study is more reliably interpreted compared to retrospective or cross-sectional approaches. By measuring attention for emotion stimuli before and after deployment, we can use temporal precedence to more clearly assess the impact of environmental influences. Third, our repeated, in situ use of the CEL allows for a more accurate and comprehensive assessment of war zone stress compared to postdeployment, retrospective reporting of stressors. In addition to these strengths, small sample size is arguably the most important study limitation. Replication with significantly larger samples is clearly warranted.
These findings represent a key link between genetic factors and potentially detrimental consequences of war zone stressor exposure. With increased war zone stress exposure, 5-HTTLPR short allele homozygotes are more likely to develop a negative attentional bias. Greater awareness for negative stimuli, in turn, likely contributes to the development of psychopathology at a later date (Beck, 2008; Clark et al., 1999; Foa et al., 1989; Foa et al., 1991). Thus, the 5-HTTLPR polymorphism in combination with negative life events, such as war zone stress exposure, may give rise to future psychopathology via increased attention for negative stimuli. Additional longitudinal research that tests this possibility is critically important, as it could help scholars to better understand factors that prevent the development of debilitating postdeployment symptoms. Investigating integrative models that incorporate etiological mechanisms across levels of analysis (e.g., genetics, environment, cognition), as in the current study, may facilitate a comprehensive understanding of vulnerability to affective psychopathology.
Footnotes
Declaration of Conflicting Interests
The authors declared that they had no conflicts of interest with respect to their authorship or the publication of this article.
Funding
This work was funded by the U.S. Army Research, Development, and Engineering Command Acquisition Center, Natick Contracting Division; and the U.S. Defense Advanced Research Projects Agency under Contract W911QY-07-C-0002 (PI: Telch). The sponsors were not involved in the design or conduct of the study; collection, analysis, management, or interpretation of the data; and preparation or approval of the manuscript. The views expressed in this publication are those of the authors and may not necessarily be endorsed by the U.S. Army.