
Abstract
Patients with and at risk for psychosis may have difficulty using associative strategies to facilitate episodic memory encoding and recall. In parallel studies, patients with first-episode schizophrenia (n = 27) and high psychosis risk (n = 28) compared with control participants (n = 22 and n = 20, respectively) underwent functional MRI during a remember-know memory task. Psychophysiological interaction analyses, using medial temporal lobe (MTL) structures as regions of interest, were conducted to measure functional connectivity patterns supporting successful episodic memory. During encoding, patients with first-episode schizophrenia demonstrated reduced functional coupling between MTL regions and regions involved in stimulus representations, stimulus selection, and cognitive control. Relative to control participants and patients with high psychosis risk who did not convert to psychosis, patients with high psychosis risk who later converted to psychosis also demonstrated reduced connectivity between MTL regions and auditory-verbal and visual-association regions. These results suggest that episodic memory deficits in schizophrenia are related to inefficient recruitment of cortical connections involved in associative memory formation; such deficits precede the onset of psychosis among those individuals at high clinical risk.
Although research on schizophrenia has traditionally focused on psychosis as the primary defining dimension of illness and intervention target, it is now widely recognized that neurocognitive deficits also represent core features of schizophrenia that greatly constrain patients’ social and occupational functioning and overall quality of life (Green, Kern, Braff, & Mintz, 2000). Of the domains of cognition affected in schizophrenia, those that affect long-term memory are among the most severe and debilitating (Aleman, Hijman, de Haan, & Kahn, 1999). Such deficits are present in patients during their initial episode (Saykin et al., 1994) and in adolescents at clinical (Fusar-Poli et al., 2012; Seidman et al., 2010) and genetic high risk for psychosis (Agnew-Blais & Seidman, 2013; Whyte et al., 2006), thereby indicating that they to some degree mark vulnerability to schizophrenia and may thus participate in the deterioration in functioning associated with onset of psychosis. Nevertheless, given that impairments in learning and memory tend to persist even after the successful amelioration of psychotic symptoms with antipsychotic drugs (Minzenberg & Carter, 2012), the mechanisms underlying these deficits must be to some extent independent of those involved in psychotic symptoms. Accordingly, it is imperative to identify the sources and timing of onset of learning and memory deficits in schizophrenia, given that this information will help in the development of interventions that target these deficits and related impairments in functioning as primary end points.
Several lines of evidence have implicated the medial temporal lobe (MTL) memory system as playing a key role in the learning and memory deficits in schizophrenia. Patients with schizophrenia show greater memory impairment when testing requires recollection of the learning context (as in episodic memory), and they perform better when recognition may be based on impressions of familiarity (Danion, Rizzo, & Bruant, 1999; Huron et al., 1995). Episodic memory has been shown to depend critically on structures in the MTL region, including perirhinal cortex, hippocampus, and parahippocampal cortex, whereas recognition via familiarity-based processes is less dependent on these MTL regions (Achim & Lepage, 2005; Eldridge, Engel, Zeineh, Bookheimer, & Knowlton, 2005; Yonelinas, Otten, Shaw, & Rugg, 2005). Of particular interest is the role of these regions during initial encoding. To lay the foundation for vivid or detailed memories, an integrative process must be active during the moment of learning to associate disparate aspects of the stimulus environment, a process referred to as “feature binding.” Such integration requires functional communication between the MTL regions implicated in long-term memory and regions involved in the active perceptual processing of the stimuli at hand (e.g., inferior frontal and superior temporal gyrus for auditory-verbal information or fusiform gyrus for pictorial-imagery information), along with prefrontal regions involved in selection of stimuli and maintenance of the present behavioral goal (Dudukovic & Knowlton, 2006; Gardiner & Java, 1990; Murray & Ranganath, 2007).
Given that disruptions in short- and long-range neural connectivity are thought to play key roles in the pathophysiology of schizophrenia (Karlsgodt et al., 2008), alterations in functional connectivity between the MTL and these other cortical regions may underlie the episodic memory deficits in these patients (Wolf et al., 2007; Zhou et al., 2008). Although some prior studies have demonstrated support for this hypothesis in patients with established illness (Glantz & Lewis, 2000; Heckers et al., 1998; Ragland et al., 2009; Ranganath, Minzenberg, & Ragland, 2008; van Erp et al., 2004), particularly in relation to the association of two or more stimuli during learning (Armstrong, Kose, Williams, Woolard, & Heckers, 2012; Lepage et al., 2006; Luck et al., 2009; Ragland et al., 2012), it remains to be determined whether such changes appear during the preonset or “prodromal” phase of the disorder. Such information is critical for establishing the temporal precedence of episodic memory deficits and associated neurophysiological changes in relation to onset of fully psychotic symptoms.
Individuals at risk for psychosis have been shown to exhibit functional abnormalities during memory encoding and recollection, including dysfunction in prefrontal regions and hippocampus/parahippocampal regions (Allen et al., 2011; Lencz et al., 2006), but it is not clear whether such abnormalities are restricted to or more severe in those individuals who later convert to psychosis. However, behaviorally, greater episodic memory impairment has been observed in at-risk individuals who later convert to psychosis (Fusar-Poli et al., 2012), and verbal declarative memory performance has also been shown to predict persistence of psychosis risk symptoms over baseline symptoms, functioning, or attention performance (Simon et al., 2012). Furthermore, individuals who later convert to psychosis show steeper rates of gray matter decline in prefrontal cortex (PFC) and parahippocampal gyrus during a 1- to 2-year interval (Mechelli et al., 2011; Pantelis et al., 2003; Sun et al., 2009). It thus seems likely that alterations in the memory circuitry predate and may even predict onset of psychosis among individuals at risk. Tests of this hypothesis can be facilitated by studying those individuals at clinical high risk (CHR) for psychosis in parallel with patients in their first episode (FE) of schizophrenia to elucidate continuities/discontinuities in neural and behavioral correlates of episodic memory dysfunction across the prodromal and fully psychotic phases of illness. Such a test also requires the use of a trial-based design that permits selective averaging of correctly remembered items at encoding and retrieval. Otherwise, impaired functional connectivity in episodic memory circuitry could reflect differences in performance, attention, or effort during encoding or retrieval rather than deficits in mnemonic processes per se.
In the present study, we used a variant of the remember-know paradigm in parallel trial-based functional MRI (fMRI) studies of (a) patients with FE schizophrenia versus control participants and (b) individuals at CHR for psychosis versus control participants. In keeping with prior fMRI studies of associative learning and memory in schizophrenia, we hypothesized that FE patients would show aberrant MTL connectivity with prefrontal cognitive control regions and with perceptual regions involved in stimulus processing. The key novel question addressed in this study is whether similar deficits would characterize CHR patients, especially those who later converted to psychosis. Such a pattern would indicate that altered functional connectivity of the memory circuitry constitutes a risk trait that precedes and predicts psychosis in at-risk individuals and may therefore participate in the deterioration in functioning associated with onset of psychosis. We also expected to extend behavioral evidence of a differential deficit in recollective versus familiarity-based memory, previously observed in patients with chronic schizophrenia (van Erp et al., 2008), to both of the patient samples. By studying patients in the preonset and early phases of schizophrenia, we eliminated many confounding effects of chronic illness, thereby enabling a clearer vantage point from which to identify when in the developmental course of schizophrenia episodic memory deficits first manifest.
Method
Participants—Sample 1 (FE schizophrenia patients)
The study was approved by the institutional review boards of the University of California, Los Angeles (UCLA) and Yale University, and all participants provided written informed consent. The sample comprised 27 outpatients with a diagnosis of FE schizophrenia (n = 23) or schizoaffective disorder–depressive type (n = 4), determined by administering the Structured Clinical Interview for DSM–IV Axis I Disorders (SCID-I; First, Spitzer, & Williams, 1997), and 22 demographically similar healthy control participants. All participants were between 18 and 35 years of age and understood spoken English sufficiently to comprehend testing procedures. Patients had their first psychotic episode within 2 years prior to participation in this study and were excluded from analysis if there was history of traumatic brain injury or clinically significant neurological disorder, evidence of substance abuse in the previous 6 months, or history of drug-induced psychotic episodes or if substance use was considered a dominant factor in the course of illness. Patients were recruited from Los Angeles public and private hospitals and clinics and were all being treated at the UCLA Aftercare Research Program. At the time of testing, all patients were clinically stabilized on oral risperidone.
The 22 healthy control participants were recruited through local newspaper and poster advertisements. Exclusion criteria for healthy control participants included a diagnosis of any DSM–IV Axis I psychotic disorder, bipolar disorder, obsessive-compulsive disorder, or posttraumatic stress disorder; recurrent or current major depression; or history of substance dependence or current substance abuse as assessed using the SCID-I. Potential control participants who met criteria for a prodromal state (Miller, McGlashan, Woods, & Stein, 1999); who had histories of neurological disorder, traumatic brain injury, or a first-degree relative with a psychotic disorder; or who were currently pregnant also were excluded from analysis. The FE and control groups did not differ significantly in terms of age, parental education, participant education, sex, or race (see Table 1 for sample demographic information, symptom characteristics, and behavioral performance).
Demographic Information, Symptom Characteristics, and Behavioral Performance for Samples 1 and 2
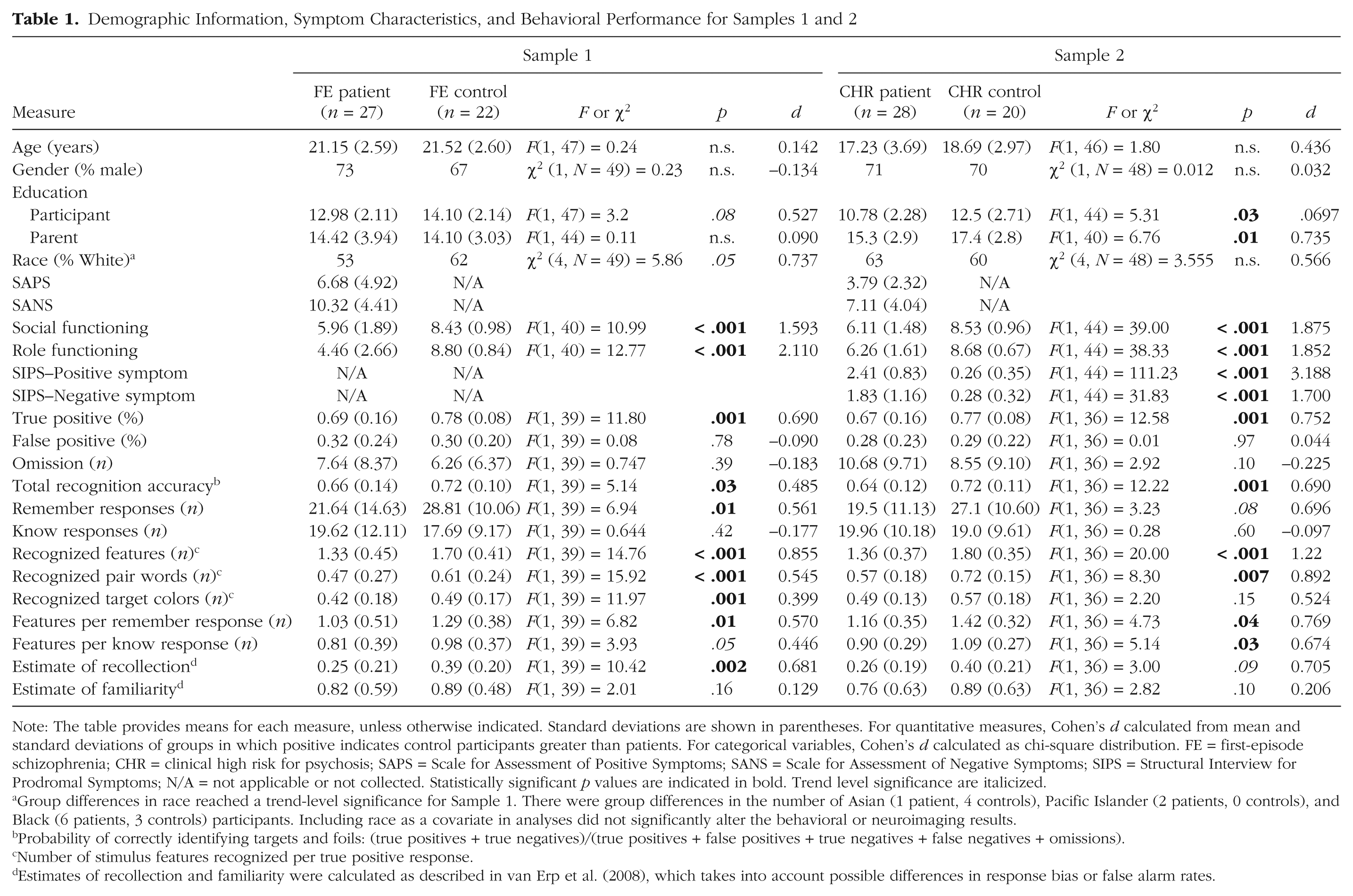
Note: The table provides means for each measure, unless otherwise indicated. Standard deviations are shown in parentheses. For quantitative measures, Cohen’s d calculated from mean and standard deviations of groups in which positive indicates control participants greater than patients. For categorical variables, Cohen’s d calculated as chi-square distribution. FE = first-episode schizophrenia; CHR = clinical high risk for psychosis; SAPS = Scale for Assessment of Positive Symptoms; SANS = Scale for Assessment of Negative Symptoms; SIPS = Structural Interview for Prodromal Symptoms; N/A = not applicable or not collected. Statistically significant p values are indicated in bold. Trend level significance are italicized.
Group differences in race reached a trend-level significance for Sample 1. There were group differences in the number of Asian (1 patient, 4 controls), Pacific Islander (2 patients, 0 controls), and Black (6 patients, 3 controls) participants. Including race as a covariate in analyses did not significantly alter the behavioral or neuroimaging results.
Probability of correctly identifying targets and foils: (true positives + true negatives)/(true positives + false positives + true negatives + false negatives + omissions).
Number of stimulus features recognized per true positive response.
Estimates of recollection and familiarity were calculated as described in van Erp et al. (2008), which takes into account possible differences in response bias or false alarm rates.
Participants—Sample 2 (CHR for psychosis patients)
The study was approved by the institutional review boards of UCLA and Yale University, and all participants (including parents in the case of minors) provided written informed consent. A total of 28 CHR adolescents, along with 20 age- and sex-matched healthy control participants, were included. The CHR participants met criteria for one of three prodromal syndrome categories, as assessed by the Structured Interview for Prodromal Symptoms (SIPS; McGlashan et al., 2001; Miller et al., 1999). Twenty CHR participants met criteria for attenuated positive symptoms, 6 for transient psychotic symptoms, and 2 for genetic risk and functional deterioration. Nine CHR adolescents (32%) converted to a psychotic disorder during the 2 years in which they participated in the study. Details regarding the SIPS criteria, reliability, and consensus procedures are described in a prior report (Meyer et al., 2005). The CHR control participants were free of any psychiatric disorders as determined by the SCID-I (First et al., 1997) or the Kiddie Schedule for Affective Disorders and Schizophrenia interview (Kaufman, Birmaher, Brent, Rao, & Ryan, 1996), did not meet criteria for any of the three prodromal syndromes, and did not have a first-degree family member with a psychotic disorder. Additional exclusion criteria for all the participants included the presence of a neurological disorder, drug or alcohol abuse or dependence within the last 6 months, pregnancy, insufficient English fluency, and IQ below 70.
All participants were recruited via community talks and advertising through the Staglin Music Festival Center for the Assessment and Prevention of Prodromal States at UCLA. The CHR patient and control groups did not differ significantly in terms of age, sex, race, and handedness but did differ in participant and parental education (see Table 1). Seven CHR patients were taking second-generation antipsychotics, 14 were taking selective serotonin reuptake inhibitors, and 5 were taking stimulants. There were no significant group differences for baseline demographic or functioning measures between CHR individuals who converted to psychosis (CHR-C) and those who did not convert (CHR-NC). However, consistent with general trends in the literature (e.g., Cannon et al., 2008), results showed that compared with CHR-NC patients, CHR-C patients had significantly higher baseline levels of total positive (CHR-C: M = 3.05, SD = 0.95; CHR-NC: M = 2.14, SD = 0.65), F(1, 26) = 8.74, p = .007, and negative (CHR-C: M = 2.56, SD = 1.19; CHR-NC: M = 1.51, SD = 1.03), F(1, 26) = 5.33, p = .03, symptoms, as assessed by the SIPS, and there was a trend toward lower baseline Global Assessment of Functioning scores in CHR-C participants relative to CHR-NC participants—CHR-C: M = 36.25, SD = 15.45; CHR-NC: M = 47.21, SD = 14.32; F(1, 26) = 3.12, p = .09.
Procedures common to both Sample 1 and Sample 2
Remember-know paradigm
The remember-know paradigm was administered using E-Prime (Psychology Software Tools; http://www.pstnet.com). Stimuli were presented as pairs of words and drawn pictures of the objects represented by the words (see Fig. 1). During the encoding phase, participants were shown 60 stimulus pairs. Participants were instructed to remember as much about the items on the screen as possible, including the target word (presented in capital letters), the color of the picture of the target word (an index of intraitem associative memory), and the paired word (an index of interitem associative memory), as well as encouraged to develop other associations. To check task engagement, we asked participants to use a button press to indicate whether the colored target picture appeared on the left or the right on each trial. During the item-recognition phase, participants were presented with the 60 previously presented targets and 20 new foil words. Participants were instructed to indicate, via a button press, whether they remember the word and could recall specific additional information presented (remember), know that the word was previously presented but could not recall additional information (know), or think the word was not previously presented (unstudied). After the item-recognition phase, participants were removed from the scanner. Next, participants completed the feature-recognition phase, in which they were first presented with a target word and asked to make a forced-choice recognition judgment between two possible paired words. A gray-scale target picture was then presented, and participants were asked to make a forced-choice recognition judgment between two possible picture colors. Both judgments also offered the “unstudied” option to indicate that the word or picture was not present in the encoding phase. The feature-recognition phase was self-paced.
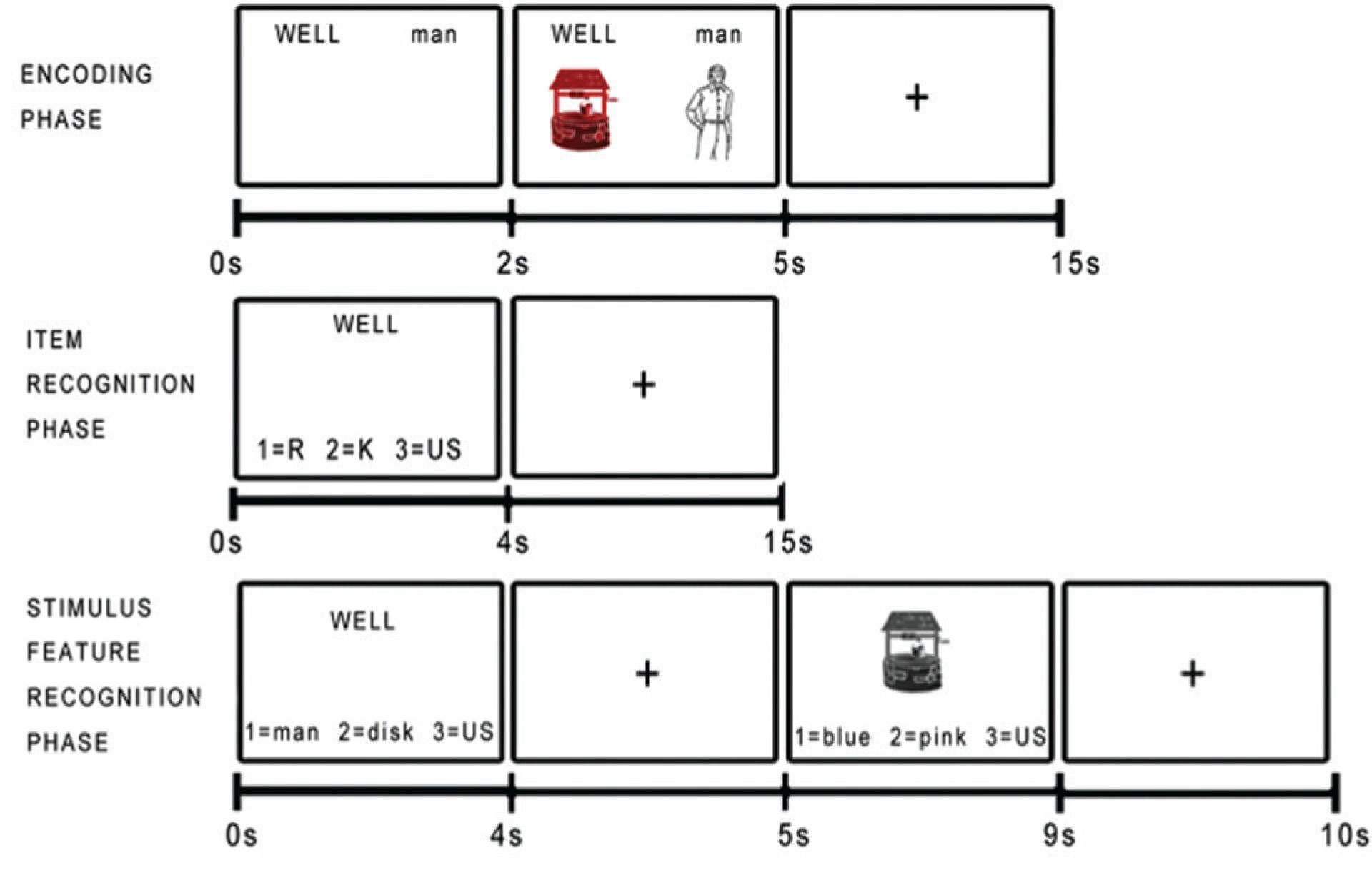
Example stimuli from the remember-know paradigm. During the encoding phase, stimuli were presented as pairs of words and pictures, and participants were explicitly asked to memorize as much about the items as possible: the words, pictures, color of the pictures, and their location on the screen. During the item-recognition phase, participants were presented with a word (either previously presented or a foil word) and asked to respond if they “remember” (R) studying the word, “know” (K) the word was previously presented, or believe the word was “unstudied” (US). In the stimulus-feature-recognition phase, participants were presented with a target word or target picture and asked to make a forced-choice recognition judgment between two possible paired words or picture colors. The encoding phase and remember-know item-recognition phase were administered in the scanner, and the stimulus-feature-recognition phase was administered outside the scanner, immediately subsequent to completion of the imaging portion.
The use of pairs of words and their corresponding pictures allows for multiple potential modes of associative binding during encoding, including a mode dependent primarily on the verbal (semantic or phonetic) content and a mode dependent primarily on the pictorial-imagery content. Before the task began, participants were given examples of the types of questions that would be asked in the item-recognition (Gardiner & Java, 1990) and associative memory recognition (Dudukovic & Knowlton, 2006) phases. As a pretest measure, participants were given six example sentences that described memories and asked whether each was reflective of a remember or of a know memory. Participants were required to get at least four out of six questions correct. Participants responded by pressing the “one,” “two,” and “three” keys on a Current Designs HH-1×4-L button box (http://www.curdes.com). E-Prime data (responses and reaction times) were scored using Excel Visual Basic macros.
Scan parameters
Scanning was conducted on a 3 Tesla Siemens Allegra MRI scanner with a standard radiofrequency head coil at the Ahmanson-Lovelace Brain Mapping Center at UCLA. Participants were given earplugs and noise-cancellation headphones (Resonance Technology, Inc.; http://www.mrivideo.com). Foam pads on each side of the headphones were used to minimize head motion during the scans. A three-plane localizer scan, followed by a sagittal scan, was used to align a high-resolution T2-weighted scan (repetition time = 5 s, echo time = 33 ms, flip angle = 90º, field of view = 200 mm2, 1.5625 × 1.5625 × 4 mm, slice thickness = 3 mm, 1-mm gap, 34 slices interleaved) and the functional echo-planar imaging scans (repetition time = 2,500 ms, echo time = 45 ms, flip angle = 80º, field of view = 200 mm2, 3.125 × 3.125 × 4 mm, slice thickness = 3 mm, 1-mm gap, 34 slices interleaved), both with a 45º slice angle (clockwise) from the longitudinal axis of the left hippocampus. The high-resolution T2-weighted scan had a readout bandwidth along the phase encoding direction identical to the functional runs such that the b0-related distortions were identical to those of the functional scans. The task included three encoding runs and four recognition runs with 120 repetition times (5 min) each.
Statistical analyses
We conducted behavioral data, functional activation, anatomically defined regions of interest (ROIs), and psychophysiological interaction (PPI) statistical analyses.
Behavioral data analyses
Group differences in accuracy and response type were analyzed using univariate mixed-model analysis of variance (PROC MIXED in SAS Version 8.2, SAS Institute, Inc., Cary, NC). Group differences in intra- and interitem associative memory were analyzed with a doubly repeated measures analysis of variance model, which treated response type (remember, know) and feature (target color, paired word) as within-subjects repeated measures variables. Group (patient, control) and sex (male, female) entered all the models as between-subjects variables, and age was included as a covariate. The statistical models included the interactions of all the between- and within-subjects class variables.
Functional activation analyses
The high-resolution T2-weighted scans and functional scans were skull stripped using the FSL Brain Extraction Tool (Smith, 2002). First-level analyses were performed with FEAT (fMRI Expert Analysis Tool) Version 5.98, part of FMRIB’s Software Library (FSL; http://www.fmrib.ox.ac.uk/fsl). Preprocessing included motion correction (6-parameter model) to the middle time point of each run using MCFLIRT (Jenkinson, Bannister, Brady, & Smith, 2002), slice-timing correction using Fourier-space time series phase shifting, spatial smoothing using a Gaussian kernel of full width at half maximum 5 mm, grand-mean intensity normalization, and high-pass temporal filtering (Gaussian-weighted least-squares straight line fitting, with sigma = 50 s). To ensure image quality, for each scan, we visually inspected the raw and preprocessed data for artifacts. Individual time points with artifacts, such as visible motion, radiofrequency coil spiking, or signal dropout, were tagged as covariates of noninterest, and any individual scans with significant motion (> 2 mm) or artifacts were not included in the analysis. Time series statistical analysis was carried out using FILM (FMRIB’s Improved Linear Model) with local autocorrelation correction (Woolrich, Ripley, Brady, & Smith, 2001). The design matrix included remember, know, and miss responses at encoding as well as remember, know, miss, correct-rejection, and false-positive responses at recognition, convolved with a hemodynamic response function (Gamma, phase = 0, SD = 3, mean lag = 6) as predictors, along with their first derivatives to allow for temporal adjustments. Motion parameters were included as covariates of noninterest. The functional scans were registered with the high-resolution T2-weighted image (7-parameter model) and the T2-weighted images with a group-average T2 template that was aligned to the MNI152 brain (12-parameter affine transformation). In the second-level analysis, first-level contrasts were combined using a fixed-effects model. Finally, third-level group analyses were conducted using mixed-effects FLAME 1 (FMRIB’s Local Analysis of Mixed Effects) with a z threshold of 2.3, a cluster p threshold of .05, and outlier deweighting (Worsley, 2001).
Anatomically defined ROIs
ROIs were selected on the basis of research that implicated divisions of the MTL with involvement in episodic memory (Diana, Yonelinas, & Ranganath, 2010; Spaniol et al., 2009). Anatomically defined hippocampus, perirhinal cortex (consisting of anterior parahippocampal gyrus), and parahippocampal cortex (consisting of posterior hippocampal gyrus) masks were defined using the Harvard-Oxford Structural Atlas (Kennedy, Lange, Makris, & Bates, 1998; Makris et al., 1999). Voxels with atlas-derived values corresponding to a greater than or equal to 25% probability of belonging to these regions were included, and each hemisphere was represented separately. FSL’s FLIRT (FMRIB’s Linear Image Registration Tool) was used to warp ROIs back into each participant’s space by applying the inverse of the transformation matrix used during the initial registration.
PPI analyses
A PPI analysis (Friston et al., 1997) was conducted to examine whether the patient groups and their matched control groups differed on the extent to which MTL regions covaried with other brain regions during remember trials versus know trials at encoding or retrieval. We chose to contrast remember trials with know trials because MTL involvement is expected to vary during encoding and retrieval of items identified as familiar or recollected. Furthermore, a remember response involves more associated information and context than does a know response and was thus predicted to require greater long-range functional connectivity.
The PPI analysis was carried out in FSL with regressors for response type, seed-region time series, and the interaction of task and time series. The psychological regressor modeled whether a given trial consisted of a remember response or a know response. The physiological regressor comprised the time series for the ROI. A third regressor modeled the interaction of the psychological regressor and the physiological regressor, such that it identified regions in which activation covaried in a task-dependent manner (remember > know) with the MTL region. Motion parameters were again included as nuisance regressors. First- and second-level analyses were conducted on individual scans as described earlier, and group-level analysis was carried out with FEAT Version 5.98, using a cluster-corrected z threshold of greater than 2.3 and cluster-corrected p threshold of less than or equal to .05 (Worsley, 2001). In-scanner motion has been shown to affect functional-connectivity measures (Satterthwaite et al., 2012); however, there were no group differences in any motion parameters, including absolute and relative root mean square or framewise displacement (Power, Barnes, Snyder, Schlaggar, & Petersen, 2012).
Results
Behavioral performance
Remember-know responses
In the analysis of behavioral performance on the remember-know task among FE patients versus healthy control participants, there was a significant overall multivariate analysis of variance main effect of group, F(1, 48) = 4.73, p = .03, d = 0.625. Compared with control participants, FE patients had significantly reduced percent true positives and total recognition accuracy, as well as significantly lower number of remember responses and lower estimate of recollection. FE patients and control participants did not differ in the number of know responses, false positives, or omissions or in reaction time for remember or know responses (see Table 1).
Parallel results were observed in the analysis of behavioral performance among CHR patients versus their matched sample of healthy control participants. The overall multivariate analysis of variance main effect of group was significant, F(1, 46) = 4.53, p = .04, d = 0.637. Compared with control participants, CHR patients had significantly reduced percent true positives and total recognition accuracy, as well as a trend toward reduced remember responses, but CHR patients did not differ from control participants in the number of know responses, false positives, or omissions or in reaction time measures (see Table 1). However, there were no significant differences on these behavioral measures between CHR-C and CHR-NC individuals (all ps > .14; data not shown).
Intra- and interitem associative memory
Among the FE patients and their matched control participants, there were significant main effects of group, F(1, 48) = 8.68, p = .005, d = 0.846, response type, F(1, 48) = 13.13, p = .0007, and feature, F(1, 48) = 7.67, p = .008, along with a significant Group × Feature interaction, F(1, 48) = 4.611, p = .037, on the number of recognized associated features per true positive response. Across response type, FE patients recognized fewer associated features per true positive response than did control participants, and across groups, the number of recognized associated features per true positive was larger for remember responses than for know responses and larger for the paired-word feature than for the target-color feature. Control participants recognized more features than did FE patients on remember responses in terms of both interitem (paired words) and intraitem (target colors) memory, whereas the groups did not differ on associative memory features for know responses.
The results for the CHR patients versus their matched control participants were largely consistent with those just described for the FE sample. There were significant main effects of group, F(1, 46) = 13.5, p < .0003, d = 1.08, response type, F(1, 46) = 13.95, p = .0003, and feature, F(1, 46) = 4.78, p = .03, along with a significant Group × Response Type interaction, F(1, 46) = 6.73, p = .01, on the number of recognized associated features per true positive response. Across response type, CHR patients recognized fewer associated features per true positive response than did control participants, and across groups, the number of recognized associated features per true positive was larger for remember responses than for know responses and larger for the paired-word than for the target-color feature. In this case, control participants recognized more overall interitems (paired words) than did CHR patients for both remember responses and know responses. As for the primary measures of remember-know performance, there were no significant differences in measures of intra- and interitem associative memory between CHR-C and CHR-NC individuals (all ps > .34; data not shown).
Relationships with functioning
Among the FE sample, there were significant positive relationships between ratings of social functioning and number of remember responses (r = .39, p = .05), overall recognition accuracy (r = .42, p = .03), and number of associated features recalled (r = .39, p = .05). No significant relationships between these performance measures and role functioning or symptoms were found, although there was a trend toward higher ratings on the Scale for Assessment of Negative Symptoms for individuals with lower recognition accuracy (r = –.33, p = .10) and fewer features recalled (r = –.36, p = .07). There was also a trend toward a relationship between lower number of remember responses and higher ratings on the Scale for Assessment of Positive Symptoms (r = –.36, p = .07).
Among CHR patients overall, there were no significant relationships between remember-know task performance measures and functioning or symptoms, although there was a trend toward a relationship between higher ratings of role functioning and number of associated features recalled (r = .38, p = .07). However, among CHR-C individuals specifically, there was a significant positive relationship between number of features recalled and ratings of role functioning (r = .88, p = .02), with a trend in the same direction for ratings of social functioning (r = .72, p = .10).
fMRI group activation (general linear model) results
Both FE patients and their matched control participants demonstrated higher levels of regional activation during the encoding of items subsequently “remembered” compared with items subsequently identified as familiar (“know”). At encoding, compared with control participants, FE patients demonstrated significantly greater differential activation between remember versus know trials in several regions, including precuneus, insula, superior temporal gyrus, inferior frontal gyrus, precentral gyrus, and superior frontal gyrus (see Fig. 2). During recognition, FE patients also demonstrated significantly greater differential activation, relative to control participants, between remember versus know trials in the middle frontal gyrus, inferior frontal gyrus, frontal pole, and occipital cortex (see Fig. 2). In contrast, there were no regions in which control participants showed significantly greater differential remember versus know activation than did FE patients at encoding or recognition (see Table S1 in the Supplemental Material available online).
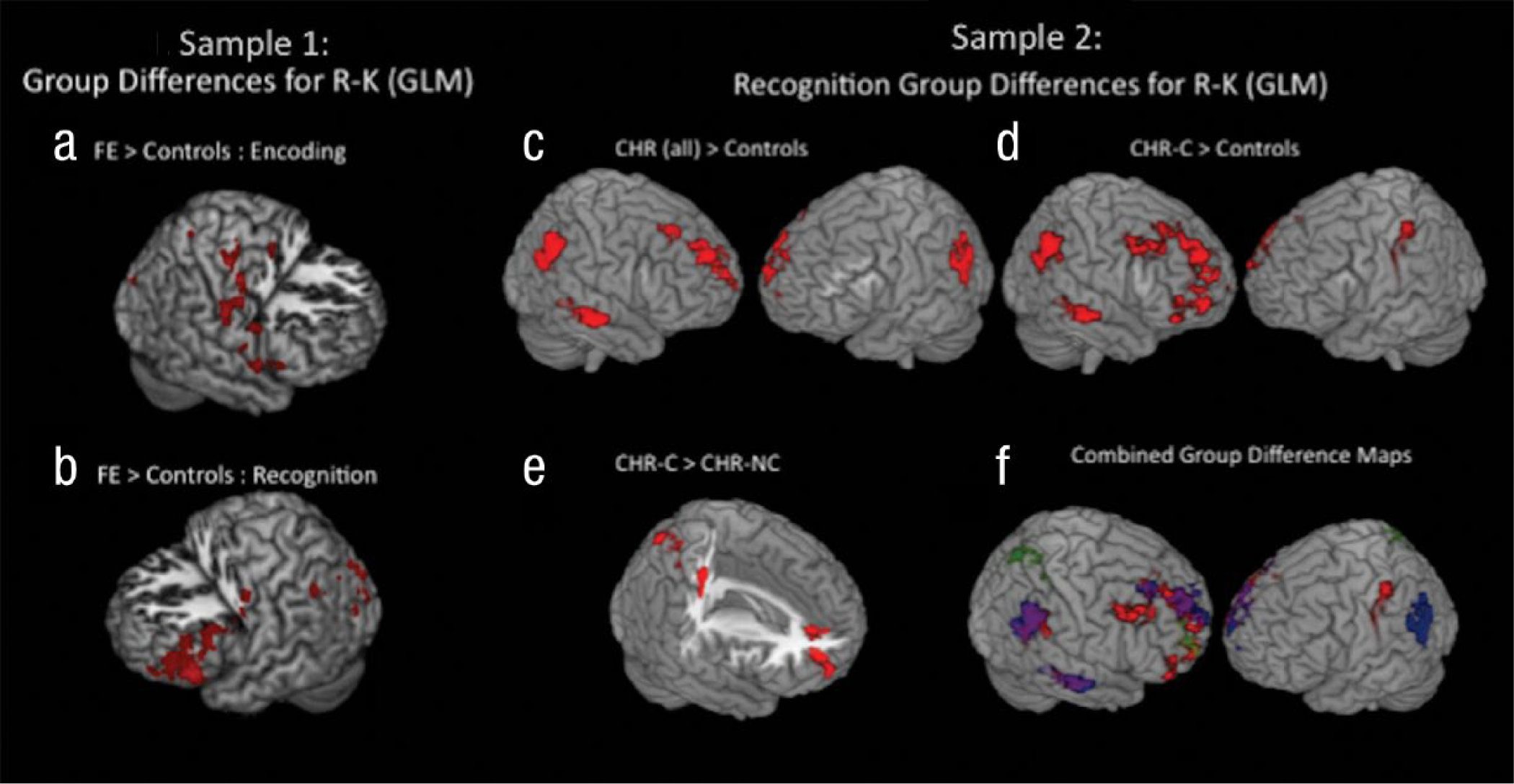
Results: Sample 1 and Sample 2 group differences. Sample 1 group differences in functional activation for remember versus know trials are shown at (a) encoding and (b) recognition. Regions in which patients with first-episode (FE) schizophrenia demonstrated greater functional activity than did control participants are shown in red. Compared with FE patients, control participants did not demonstrate significantly greater functional activity by trial type at encoding or recognition. Sample 2 group differences are shown in (c) regions in which remember produced more activation than did know in the clinical high risk (CHR) for psychosis sample during recognition (CHR individuals showed greater activation than did control participants when CHR individuals who converted to psychosis, CHR-Cs, and CHR individuals who did not convert to psychosis, CHR-NCs, were combined), (d) regions in which CHR-C patients showed more activation than did control participants during recognition, and (e) regions in which CHR-C patients showed greater remember-know activity than did CHR-NC patients during recognition (there were no significant differences between CHR-NC patients and CHR-C patients). An overlap (f) of the CHR sample group differences at recognition shows CHR patients with greater activation than control participants (red), CHR-C patients with greater activation than control participants (blue), and CHR-C patients with greater activation than CHR-NC patients (green). R-K = remember-know; GLM = general linear model.
Unlike the FE sample, there were no regions of significant activation differences during the encoding task among CHR patients and their matched control participants. However, during recognition, compared with control participants, the CHR patients demonstrated significantly greater differential activation on remember versus know trials in medial and superior frontal gyri, anterior cingulate, supramarginal gyrus, inferior parietal lobule, and middle temporal gyrus (see Table S2 in the Supplemental Material and Fig. 2). In addition, CHR-C patients demonstrated higher remember activation relative to know activation at recognition than did CHR-NC patients in several of these same regions, including the anterior cingulate, middle frontal gyrus, and superior frontal gyrus (see Table S2 in the Supplemental Material and Fig. 2).
fMRI functional connectivity (PPI) results
Encoding
As hypothesized, during encoding, FE patients showed significantly less functional connectivity than did control participants between the three MTL seed regions and regions implicated in auditory and visual association or executive functioning (see Fig. 3a). More specifically, the perirhinal cortex seed exhibited significantly less functional connectivity with auditory-language and visual-imagery regions, including middle occipital gyrus, fusiform gyrus, lingual gyrus, and cuneus (Brodmann’s area, BA, 18, 19, 31, 37, 39), and with superior and medial prefrontal gyrus regions (BA 8, 9, 10; see Table S3 in the Supplemental Material). A similar pattern of reduced connectivity in FE patients compared with control participants was seen for the hippocampal and parahippocampal cortex seeds in relation to prefrontal regions (BA 8, 9) but not in relation to regions implicated in auditory or visual association (see Fig. 3a). It is surprising that FE patients also demonstrated significantly greater functional connectivity than did control participants between the three MTL seeds and a number of brain regions, including many not generally differentially activated to a greater extent for remember trials relative to know trials, such as regions in parietal cortex and thalamus (see Table S3 in the Supplemental Material).
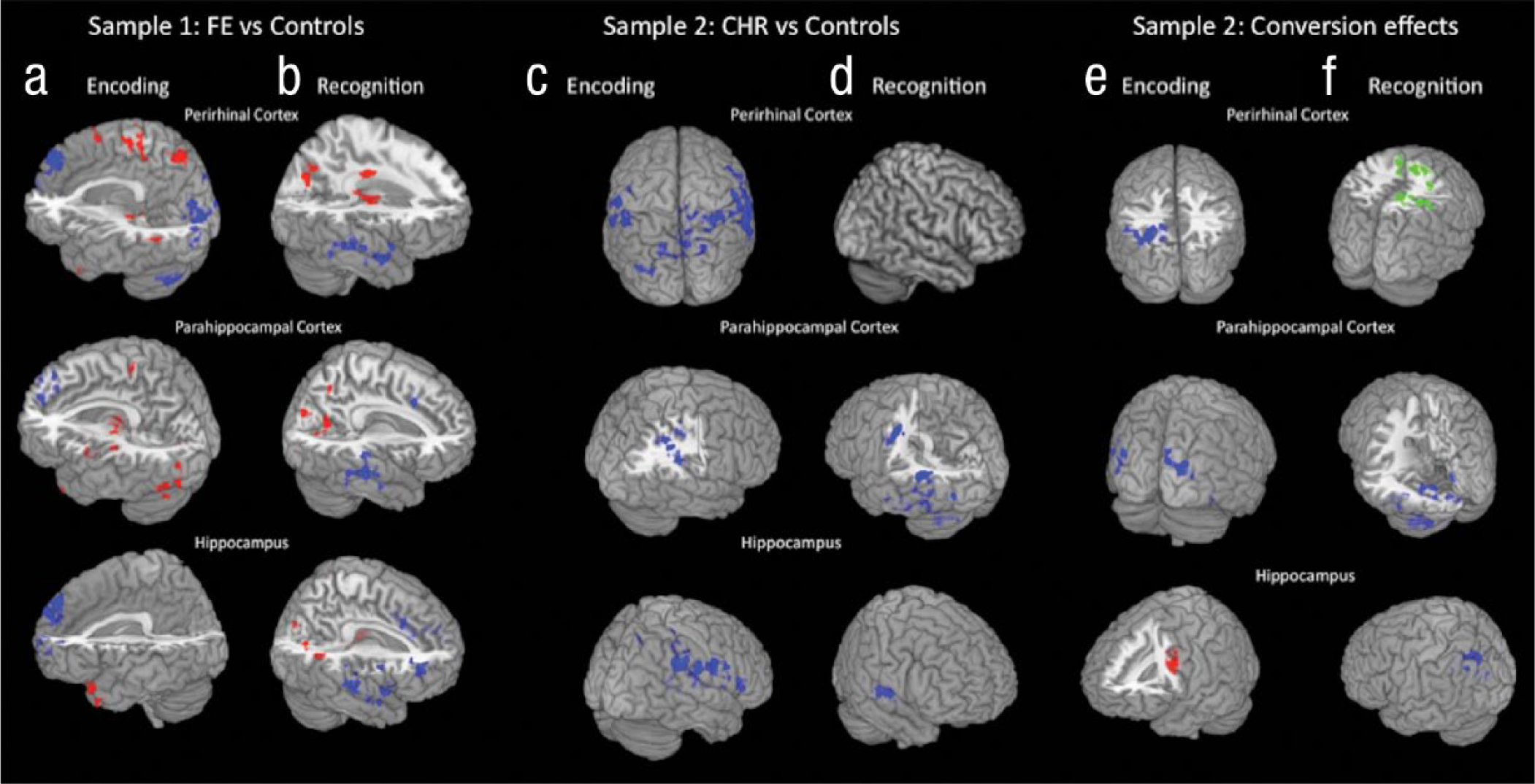
Group differences in functional connectivity for remember trials versus know trials. Sample 1 group differences in psychophysiological interaction (PPI) during (a) encoding and (b) recognition are shown with each medial temporal lobe (MTL) region of interest (ROI). Regions showing greater functional coupling in control participants are in blue, and those showing greater functional coupling in patients with first-episode (FE) schizophrenia are in red. Sample 2 group differences in PPI during (c) encoding and (d) recognition are shown with each MTL ROI. Regions showing greater functional coupling in control participants are in blue; no regions showed greater functional coupling in patients at clinical high risk (CHR) for psychosis. Sample 2 group differences in PPI during (e) encoding are shown with each MTL ROI, with those CHR patients who converted to psychosis (CHR-Cs) separated from those who did not convert (CHR-NCs). Regions showing greater connectivity such that control participants > CHR-NC patients > CHR-C patients are shown in blue. Regions showing greater connectivity such that CHR-C patients > CHR-NC patients are shown in red. Sample 2 group differences in PPI during (f) recognition are shown with each MTL ROI, with CHR-C patients separated from CHR-NC patients. Regions showing greater connectivity such that control participants > CHR-NC patients > CHR-C patients are shown in blue. Regions showing greater connectivity such that CHR-NC patients > CHR-C patients are shown in green.
The analyses for CHR patients versus their matched control participants showed parallel alterations in functional coupling between the three MTL seeds and associative cortical regions, but not with respect to MTL connectivity with frontal brain regions. As shown in Figure 3c, all three MTL seed regions exhibited significantly less functional connectivity in CHR patients than in control participants with auditory- and visual-association regions, including the superior temporal gyrus, supramarginal gyrus, posterior cingulate, and precuneus. The parahippocampal cortex and hippocampus seeds exhibited significantly less functional connectivity in CHR patients than in control participants in several additional regions, including the insula, transverse temporal gyrus, and putamen (see Table S5 in the Supplemental Material).
When CHR-C individuals were separated from CHR-NC individuals, a general pattern emerged such that connectivity of MTL seed regions with associative regions was reduced in CHR-C patients compared with CHR-NC patients and in CHR-NC patients compared with control participants (see Fig. 3e). This risk-gradient effect was observed for connectivity of the perirhinal and parahippocampal cortex seeds with auditory- and visual-association regions, including the fusiform gyrus, angular gyrus, lingual gyrus, cuneus, and middle temporal gyrus, thus paralleling the results observed for FE patients versus their matched control participants. However, the inverse pattern was observed with respect to connectivity of hippocampus and frontal brain regions such that CHR-C patients demonstrated greater functional connectivity than did both CHR-NC patients and control participants (see Table S5 in the Supplemental Material).
Recognition
At recognition, compared with control participants, FE patients showed significantly lower functional coupling of the MTL seeds with auditory-language-association regions (BA 13, 21, 22, 38), middle and superior frontal gyrus regions (BA 9, 10, 46), and regions previously implicated in decision making (BA 24, 32, 33; see Fig. 3b), but they showed significantly greater functional coupling of the MTL seeds with visual-imagery regions (BA 19, 30, 37). FE patients also exhibited greater connectivity between the perirhinal cortex and the precentral gyrus and regions in the parietal lobe, occipital lobe, and cerebellum, as well as increased functional connectivity of the parahippocampal cortex and hippocampus with the precuneus and lingual gyrus (BA 18, 19, 23, 30; see Table S4 in the Supplemental Material and Fig. 3b).
CHR patients as a group showed a pattern of reduced MTL connectivity with auditory-language and executive functioning regions at recognition similar to that observed for FE patients (see Fig. 3d), but CHR patients did not exhibit similar increases in functional connectivity of MTL with visual-association regions. In analyses that separated the CHR-C and CHR-NC patients, connectivity of the hippocampus and parahippocampal cortex seeds with auditory-language-association areas was significantly lower in CHR-C patients than in CHR-NC patients and significantly lower in CHR-NC patients than in control participants (see Table S6 in the Supplemental Material and Fig. 3f).
Discussion
CHR-C individuals showed many of the same deviations in functional connectivity of the memory-related circuitry seen in FE individuals. In particular, both FE patients and CHR-C patients showed altered functional coupling of MTL regions critically implicated in learning and memory with regions involved in auditory-language and visual-imagery processing at both the encoding- and the recognition-task stages. At the same time, there were important differences between the patterns of physiological and behavioral deviations between FE patients and CHR-C patients compared with their respective control participants. These differences relate primarily to the relatively greater deficits in FE patients compared with their control participants in terms of episodic memory performance and MTL coupling with prefrontal regions involved in organization and selection of stimuli and the apparent “recruitment” of additional regions not differentially active for remember trials versus know trials. These differences provide important clues as to the components of the learning and memory system already compromised in the at-risk state versus those that may deteriorate during the emergence and early course of psychosis.
Because the stimuli used in this study included verbal and pictorial representations, participants could use both types of information to fuse the information for later recall. This design feature was incorporated to ensure that FE patients would achieve a sufficient number of remember responses to allow for meaningful statistical comparison, but it also provided for testing the hypothesis of disrupted MTL connectivity in relation to more than one channel of perceptual representation. Consistent with prior studies of associative learning in schizophrenia, results showed that the FE patients in this study had reduced functional coupling of MTL regions with regions involved in auditory-verbal and visual-imagery stimulus representation and cognitive control during encoding (Armstrong et al., 2012; Lepage et al., 2006; Luck et al., 2009; Ragland et al., 2012). Parallel results were observed for CHR individuals overall, and particularly for CHR-C individuals, with respect to reduced connectivity of the MTL seeds with regions involved in auditory-verbal and visual-imagery processing but not with respect to MTL connectivity with prefrontal regions.
Given that the fMRI assessment for CHR patients was performed in a prepsychotic phase of illness, on average almost 17 months before the onset of psychosis, this pattern suggests that although disruption of MTL connectivity with task-relevant extrafrontal brain regions precedes and predicts onset of psychosis, disruption of functional connectivity between MTL structures and PFC may develop and worsen as one moves from a prodromal, prepsychotic phase to a fully psychotic phase of illness. This interpretation is also consistent with the observation of progressive loss of gray matter in PFC as psychosis develops in CHR individuals (Sun et al., 2009). Given the small number of CHR-C patients (n = 9), it is possible that there was lower statistical power to detect a difference in MTL-PFC connectivity compared with control participants in this group. However, when we compared CHR-C patients with control participants, few voxels in PFC regions emerged in maps of differences in functional connectivity when statistical thresholds were reduced to zs of 1.5 and 1.0 (cluster-corrected). It is also possible that a deficit in MTL-PFC connectivity exists in CHR-C patients but is obscured by differences in age-related maturation of the PFC. In this study, control participants for the FE sample demonstrated greater MTL-frontal connectivity than did control participants for the CHR sample, which suggests that the failure of the CHR sample to replicate the frontal deficits found in the FE sample may be explained in part by limited recruitment of frontal regions by the CHR control participants. The introduction of antipsychotic medications could also account for the differential patterns of functional-connectivity deficits seen in FE patients versus CHR-C patients. Our ability to evaluate this possibility statistically was limited by small sample sizes; however, there were no consistent differences in MTL-frontal connectivity between CHR-C patients exposed to antipsychotic medications (n = 5) and those who were not exposed (n = 4).
Despite evidence of reduced connectivity of MTL with frontal, temporal, and occipital brain regions during encoding, during recognition testing, FE patients demonstrated increased functional connectivity of parahippocampal cortex with some of the same regions, especially those involved in visual perception and association (e.g., BA 18), for remember responses. Prior work has shown that activity in visual-association regions during retrieval is greater with true memory compared with false memory, thereby suggesting that these regions are important for accurate recognition (Slotnick & Schacter, 2004). The present results suggest that although patients do not engage MTL–visual association coupling to the same extent as control participants at encoding, they are able to use similar, more contextually motivated pathways to facilitate recall, thereby coactivating this circuitry at higher than normal levels. This higher than normal functional coactivation of MTL regions with visual-processing regions during remember responses may reflect a compensatory phenomenon (i.e., greater coactivation needed in patients than in control participants to produce the same behavioral output). However, given that this hyperconnectivity pattern at successful episodic retrieval was not observed in the CHR patients, regardless of conversion status, less compensation may be needed in the preonset phase of illness, an interpretation that is supported by the attenuated levels of deficits on a behavioral measure of recollective recall among CHR patients compared with FE patients.
The evidence of reduced MTL-prefrontal connectivity in FE patients during encoding could reflect differences in learning strategies. Prior studies have shown that patients do not naturally make use of strategies such as encoding and rehearsing items based on semantic content (Iddon, McKenna, Sahakian, & Robbins, 1998), although when such strategies are overtly suggested to patients, they appear able to use the strategies to improve memory performance (Ragland et al., 2003; Ragland et al., 2009). The fact that patients in the present study showed enhanced MTL–visual cortex connectivity patterns at recognition testing suggests that these regions are able to become functionally active but did not do so during the critical moment of learning, thereby supporting the notion that encoding strategy may affect associative memory deficits (Bonner-Jackson & Barch, 2011).
Although participants were instructed to remember not only the target word but also the intraitem (picture color) and interitem (paired word) features—and prior to beginning the experiment, were shown several examples of how memory for these stimuli would be tested—we did not provide participants with a specific strategy for forming associations among these elements during encoding. Thus, it remains possible that if patients were given an explicit instruction to use an imagery-based strategy at encoding, especially one that emphasizes unitization (Quamme, Yonelinas, & Norman, 2007), patients would show a normal level of functional coupling of MTL regions with imagery regions during learning. However, although it may be possible to normalize the connectivity patterns of episodic memory circuitry in patients with schizophrenia using specific instructions, it is important to keep in mind that patients do not appear to naturally use contextual information as readily during learning, which likely contributes significantly to their day-to-day memory deficits. Patients also demonstrated reduced prefrontal-MTL coupling during remember trials versus know trials at recognition testing, which suggests that the deficits in strategy or contextual organization at encoding constrain the use of these processes at recall or in postretrieval monitoring (Achim & Lepage, 2005). Again, these deficits were not observed in CHR-C patients, thereby suggesting that such deficits emerge during the development or early course of schizophrenia.
Although FE patients showed higher task-related activation of the memory circuitry than did control participants at both encoding and recognition for items remembered episodically, they showed less task-related coactivation of the MTL with task-relevant perceptual regions. FE patients also showed higher levels of coactivation of the MTL seeds during encoding with regions that were not clearly associated with the primary representational features of the stimuli and that were distributed throughout a number of regions not differentially active for remember responses versus know responses. These results may be reflective of compensation for inefficient coactivation of the MTL regions with those involved in processing the key stimulus features. CHR patients also showed higher than normal differential activation of the memory circuitry for remember responses versus know responses at recognition, although this was not accompanied by the diffuse hyperconnectivity seen in the FE patients, thereby suggesting that less compensation was necessary in the preonset phase of illness.
In this study, behavioral measures of episodic memory were not sensitive to differences between CHR-C and CHR-NC individuals, which suggests that in the preonset phase, the behavioral impairment in episodic memory may be more strongly associated with risk for psychosis rather than predictive of a psychotic disorder outcome per se. This pattern is in contrast to that seen in some prior studies (Fusar-Poli et al., 2012) in which behavioral deficits in episodic memory at baseline were more severe among patients who later converted to psychosis, although those studies used free-recall verbal memory tests rather than the remember-know recognition paradigm used in the present study. However, in this study, there was a significant association between memory performance and functioning in the CHR-C participants, thereby suggesting that these behavioral deficits may show growing relevance to the clinical picture as psychosis develops.
The results of the present study suggest several avenues for future exploration. Although this study demonstrates the potential for improved understanding of the pathophysiology of psychosis with the use of functional connectivity assessments obtained using fMRI, measures of structural connectivity would help to clarify the nature of the underlying neural disruptions. Diffusion tensor imaging has been used to show reduced white matter integrity and disrupted structural connectivity in individuals with schizophrenia (Lee et al., 2013; Yao et al., 2013) and those at risk for psychosis (Karlsgodt, Niendam, Bearden, & Cannon, 2009). These alterations have been found in fiber tracts linking the MTL to other regions (Ellison-Wright & Bullmore, 2009) and have been associated with episodic memory performance (Nestor et al., 2007). A multimodal imaging study of associative episodic memory in patients with schizophrenia would help to clarify whether performance deficits and related changes in functional connectivity reflect disruptions in the white matter tracts linking MTL and other cortical regions. Alternatively, the changes in functional connectivity could be due to differences in strategy (as discussed earlier) or task engagement (Salomon et al., 2011). For example, a failure to suppress internally focused default mode network (DMN) activity during encoding may affect patients’ memory performance (Whitfield-Gabrieli et al., 2009). Robust examination of DMN activity requires collection of fMRI data during a rest period, something that was not done in this study. However, we detected no significant differences between patients and control participants in either the FE sample or the CHR sample in terms of activity and functional connectivity in regions associated with the DMN (i.e., medial prefrontal and posterior cingulate cortex).
An additional avenue for future research is to investigate the potential of targeting these deficits for remediation. It is tempting to speculate that training patients to engage in imagery-based encoding strategies during learning may be an effective intervention to improve memory performance. However, although cognitive training is an increasingly attractive treatment modality, there remain many questions about targeting specific domains and maximizing generalizability to overall functional outcome (Wykes, Huddy, Cellard, McGurk, & Czobor, 2011). In this study, we found some evidence for a relationship between episodic memory performance and functioning and, in general, cognition as a predictor of functional outcome in schizophrenia (Green et al., 2000). However, correlational measures such as those employed here do not address whether the cognitive deficits are causally related to the functional deficits. Instead, future research must use experimental designs to determine whether training on strategic encoding can remediate behavioral deficits in memory and, in turn, whether improvements in memory performance predict improved functional outcome. In addition, there may be different primary targets for remediation depending on phase of illness.
In conclusion, these results suggest that episodic memory deficits in schizophrenia are related to failures to integrate relevant aspects of the learning context that facilitate detailed recall. These deficits are present prior to illness onset and may be associated with illness risk, although cross-sectional comparison also suggests potential neural progression from the prodromal phase to the fully psychotic phases of illness. The two primary limitations of this study are the relatively low statistical power for comparing CHR-C individuals with CHR-NC individuals and the cross-sectional design. Future studies with a larger sample size and a longitudinal design will be better able to clarify the nature of functional-connectivity abnormalities during episodic memory pre- and postonset of a psychotic disorder and the timeline of MTL-associative and MTL-frontal disconnectivity in relation to disease progression.
Footnotes
Declaration of Conflicting Interests
The authors declared that they had no conflicts of interest with respect to their authorship or the publication of this article.
Funding
This work was supported by Translational Research Center in Behavioral Science Grant P50 MH066286, Developmental Processes in Schizophrenic Disorders Grant MH037705, Grant MH082719, and gifts to the UCLA Foundation from the International Mental Health Research Organization and Garen and Shari Staglin. The initial medication, risperidone, for the patients in the UCLA Aftercare Research Program was supplied by Ortho-McNeil Janssen Medical Affairs. None of the funding agencies had a role in collection, management, analysis, or interpretation of the data or in the preparation, review, or approval of the manuscript. K. H. Nuechterlein has received research grants from Janssen Scientific Affairs, LLC; Genentech; and Brain Plasticity, Inc. and is a consultant to Genentech and Otsuka.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.