
Abstract
Previous behavioral genetic research in children has found that conduct problems in the presence of high CU traits are more heritable than conduct problems in the presence of low CU traits—a gene × trait interaction. The current study replicates and extends this finding using a sample of adolescent twins from the Texas Twin Project, who were assessed for rule-breaking and aggression. We find evidence that genetic influences on CU traits contribute to genetic liability for both rule-breaking and aggressive behavior. CU traits moderate genetic influences on aggressive behavior, such that the heritability of aggression is higher among youth with high levels of CU traits. However, we do not find evidence that CU traits moderate genetic influences on rule-breaking behavior. The continuum of callous-unemotionality and the aggression versus rule-breaking distinction continues to be meaningful and intersecting methods for characterizing heterogeneity in the etiology of antisocial behavior.
Keywords
Callous-unemotional (CU) traits are a cluster of affective and interpersonal risk factors for antisocial behavior that include shallow affect, lack of remorse, and a cruel disregard for others. An established corpus of studies indicates that antisocial behavior is unique for youth with high levels of CU traits, including distinct neurological and physiological profiles (reviewed by Frick, Ray, Thornton, & Kahn, 2014; Viding, Fontaine, & McCrory, 2012). To this end, “callousness” was included in the fifth edition Diagnostic and Statistical Manual of Mental Disorders as a personality disorder trait facet, as well as a specifier for conduct disorder (American Psychiatric Association, 2013; Krueger & Markon, 2014).
A number of quantitative genetic studies have examined the etiology of CU traits and antisocial behavior. Twin and adoption studies have found evidence that CU traits are moderately to highly heritable and index genetic liability for antisocial behavior (Hyde et al., 2016; Viding, Frick, & Plomin, 2007; Waldman et al., 2011). That is, the same genes that influence CU traits also contribute to genetic risk for antisocial behavior. For example, Hyde et al. (2016) found that CU traits in adopted children (27 months old) were significantly correlated with their biological mothers’ antisocial behavior, which provides evidence of heritable risk for CU traits in early childhood. In a sample of 7- to 9-year-old twins, Viding et al. (2007) found that the genetic correlation between CU traits and conduct problems was statistically significant and moderate in magnitude for both boys and girls (.57 and .65, respectively). In a sample of child and adolescent twins (N ~ 4,000), Waldman et al. (2011) found that additive genetic influences on low levels of prosociality (similar to high levels of callousness) accounted for 20% of the variance in symptoms of conduct disorder. Moreover, additional twin studies of youth and young adults have found evidence of substantial genetic overlap between CU traits and antisocial behavior (Larsson, Andershed, & Lichtenstein, 2006; Taylor, Loney, Bobadilla, Iacono, & McGue, 2003). Taken together, these results indicate that the phenotypic association between CU traits and antisocial behavior is driven partly by genetic factors shared by both phenotypes.
In addition to indexing genetic liability for antisocial behavior, there is also evidence that the presence of high CU traits differentiates a highly heritable form of antisocial behavior in childhood. Specifically, Viding, Blair, Moffitt, and Plomin (2005) found that antisocial behavior was more highly heritable among 7-year-old children with high levels of CU traits, compared to children with average or low levels of CU traits (i.e., a gene × trait interaction; DeYoung & Clark, 2012). This gene × CU trait interaction was replicated by the same research group in a sample of 9-year-old children (Viding, Jones, Frick, Moffitt, & Plomin, 2008), although the magnitude of the effect was less pronounced. Nonetheless, results suggest that antisocial behavior is more highly heritable among children who have high levels of CU traits, compared to children with low levels of CU traits.
It is currently unclear whether the moderating effect of CU traits on genetic risk for antisocial behavior extends to different subtypes of antisocial behavior or to antisocial behavior measured in developmental periods beyond childhood. One useful method for parsing heterogeneity in antisocial behavior centers on the distinction between aggressive behavior (e.g., acts that harm or victimize others) and nonaggressive rule-breaking (e.g., theft and truancy; Burt, 2009; Harden et al., 2015). Previous studies of whether CU traits moderate the heritability of antisocial behavior have used measurements that collapse across the aggression versus rule-breaking distinction. For example, Viding et al. (2008) used a 5-item measure of antisocial behavior that included both aggression items (“I fight a lot”) and rule-breaking items (“I am often accused of lying and cheating”). Similarly, the 5-item measure of antisocial behavior in Viding et al. (2005) included both “often fights with other children or bullies them” and “often lies and cheats.” Therefore, it is unclear from previous studies whether the observed moderating effect of CU traits is specific to aggression or rule-breaking, or general across these different types of antisocial behavior.
Indeed, there are at least three reasons to believe that a gene × CU trait interaction may differ across development, as well as between aggressive versus nonaggressive subtypes of antisocial behavior. First, the rank-order stability and mean prevalence rates of aggression and rule-breaking behaviors vary across development. As is the case for CU traits, rank-order stability of aggression is high beginning in early childhood (Burt, 2012; Frick & White, 2008). In contrast, rank order stability of rule-breaking is low in childhood and increases in adolescence. Moreover, mean levels of aggression peak in early childhood, whereas mean levels of rule-breaking increase in adolescence. Second, aggression tends to be more heritable (~65%) than rule-breaking (~48%), and is subject to little or no shared environmental influences, particularly after childhood (Burt, 2009). Conversely, rule-breaking is more susceptible to shared environmental influences (~18%; Burt, 2009). This has been interpreted as reflecting a potentially more normative trajectory of rule-breaking behaviors compared to physical aggression, an interpretation also supported by differentiated personality correlates between these subtypes (Tackett, Daoud, De Bolle, & Burt, 2013). Third, the etiology of aggression and rule-breaking differs across development. The heritability of aggression remains stable from childhood to adolescence, whereas the heritability of rule-breaking increases with adolescent age and pubertal development (Burt & Klump, 2009; Burt & Neiderhiser, 2009; Harden et al., 2015). Given that aggression and rule-breaking have distinct etiologies and show differential patterns of phenotypic expression across development, it is important to establish whether the gene × CU trait interaction previously observed in children will generalize across subtypes and developmental periods.
Goals of the Current Study
The current study uses a sample of adolescent twins to test the hypothesis that antisocial behavior is highly heritable in the presence of high levels of CU traits, as has been previously found in childhood. One limitation of previous research is suboptimal measurement of the CU construct, defined by the use of questionnaires with few items, binary response options, and poor internal consistencies (Waller, Gardner, & Hyde, 2013). In the current study we measure CU traits in a population-based sample of adolescent twins using a well-validated, 24-item questionnaire. Furthermore, to help further delineate heterogeneity in the association between CU traits and antisocial behavior, we measure rule-breaking and aggression separately using a 25-item questionnaire.
Method
Sample
Texas twin sample
The current sample consists of twins and multiples (n = 824) from families (n = 407) registered in the Texas Twin Project (Harden, Tucker-Drob, & Tackett, 2013), a school-aged twin registry that is representative of the greater Austin and Houston metropolitan areas. Twins and multiples in Grades 9 to 12 were recruited using public school rosters and invited to participate in an ongoing in-lab study, which involves a large battery of cognitive assessments, surveys, and behavioral tasks. Trained research assistants tested all siblings separately. Surveys were computerized and administered to participants privately in a quiet room. All participants either were currently enrolled in grade school or had graduated within the past 3 months, but were still living at home with their cotwin. 1
The Texas Twin sample represents a socioeconomically diverse population. Approximately one third of families reported having received a needs-based form of public assistance since the twins were born. Of the sample, 60% identified as non-Hispanic Caucasian, 25% identified as Hispanic/Latino, 10% identified as African American, and 5% identified as another race or ethnicity. Parental education varied widely with 6% of parents not completing high school, 28% completing high school or a vocational or technical program, 6% completing an associate’s degree, 36% a bachelor’s degree, and 24% a graduate or professional degree.
Zygosity
Twins and multiples were designated as monozygotic (MZ) or dizygotic (DZ) based on the results of a latent class analysis (LCA; B. Muthén, 2004), which assigns members to different subgroups in a population (e.g., MZ or DZ twins) on the basis of self- and informant-reports of physical similarity. Specifically, each twin, parents of twins, and two research assistants per twin-pair reported on questionnaire items that assessed the twins physical similarity (e.g., “Do your twins have the same color eyes?”; “How similar do the twins look in their facial appearance/hair color?”) and likelihood for being mistaken for each other (e.g., “Do family members ever mistake one twin for the other?”; “Do you have difficulty telling one twin from the other?”). Results of the LCA indicated that approximately 35% and 65% of the sample were MZ and DZ twins, respectively, and had entropy statistics equal to 0.999, which indicates that there was a high level of certainty in assigning twin-pairs to zygosity groups. Note that previous research that compared zygosity classification obtained by genotyping to LCA of questionnaire data found that the latter had a misclassification rate of less than 1% (Heath et al., 2003).
Measures
CU Traits
CU traits were measured using adolescents’ responses to the Inventory of Callous-Unemotional Traits. This 24-item questionnaire was developed to measure individual differences in callous, careless, uncaring, and unemotional dispositions (Kimonis et al., 2008). Items include “I do not care whom I hurt to get what I want,” “I do not care about doing things well,” “I am concerned about the feelings of others (R),” and “I do not show my emotions to others,” which are rated on a 4-point scale (1 = disagree strongly, 4 = agree strongly). Consistent with the results of a recent item response theory analysis (Ray, Frick, Thornton, & Cauffman, 2015), we do not decompose the ICU into subscale scores and, instead, compute a mean composite score including responses from all items.
Aggression and rule-breaking
Aggression and rule-breaking were measured using twins’ responses to externalizing items from the Youth Self-Report (YSR) version of Child Behavior Checklist (Lizotte, Chard-Wierschem, Loeber, & Stern, 1992). Based on previously published factor analytic work (Harden et al., 2015), 13 items were used to measure aggression, and 12 items were used to measure rule-breaking. All items are rated on a 3-point scale (0 = not true, 2 = very true or often true), and twins’ responses to items were summed and then divided by the total number of items to produce mean aggression and rule-breaking scores (see Table 1).
Descriptive Statistics, Correlations, and Univariate Twin Models of Callous-Unemotional (CU) Traits, Aggression (AGG), and Rule-Breaking (RB)
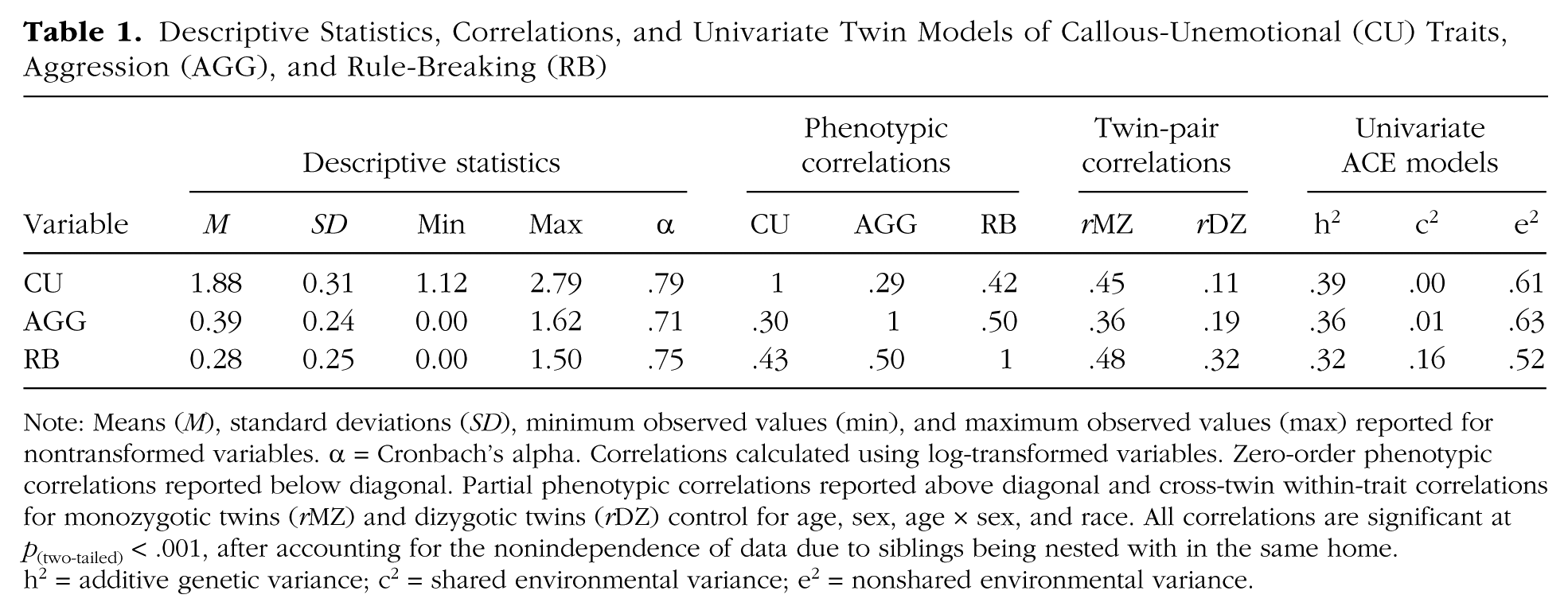
Note: Means (M), standard deviations (SD), minimum observed values (min), and maximum observed values (max) reported for nontransformed variables. α = Cronbach’s alpha. Correlations calculated using log-transformed variables. Zero-order phenotypic correlations reported below diagonal. Partial phenotypic correlations reported above diagonal and cross-twin within-trait correlations for monozygotic twins (rMZ) and dizygotic twins (rDZ) control for age, sex, age × sex, and race. All correlations are significant at p(two-tailed) < .001, after accounting for the nonindependence of data due to siblings being nested with in the same home. h2 = additive genetic variance; c2 = shared environmental variance; e2 = nonshared environmental variance.
Data analytic procedures
Internal consistencies (Cronbach’s alpha) and descriptive statistics were calculated using R version 3.1.2 (R Core Team, 2013). All inferential analyses were conducted within a structural equation modeling framework using Mplus Version 7.1 (L. K. Muthén & Muthén, 2010). Missing data were handled by estimating models using full-information maximum likelihood with robust standard errors. The CU scale approximated a Gaussian distribution and rule-breaking and aggression scales were log-transformed to correct for positive skew. To control for potential covariates, log-transformed scales and the CU scale were residualized for the main effects of Caucasian, Hispanic, and African American race/ethnicity; age; sex; and age × sex interaction (McGue & Bouchard, 1984). Finally, residual scale scores were standardized before conducting quantitative genetic analyses. All phenotypic models implemented a sandwich estimator to account for the nonindependence of data that results from siblings being clustered within the same family. The absolute fit of models was evaluated using root mean square error of approximation (RMSEA) and Tucker-Lewis index (TLI), where RMSEA values < .05 and TLI values > .90 indicate good fit to the data. Comparative model fit was evaluated using Akaike information criteria (AIC), Bayesian information criteria (BIC), sample-size-adjusted Bayesian information criteria (SSBIC), and Satorra-Bentler scaled log-likelihood chi-square difference tests (Δχ2).
Inferential analyses were conducted in three steps. First, zero-order and partial phenotypic correlations between focal constructs were estimated. Second, quantitative genetic models were used to estimate latent genetic and environmental correlations between CU traits, rule-breaking, and aggression. Finally, quantitative genetic models were extended to test whether CU traits moderated the latent genetic and environmental components of rule-breaking and aggression.
Results
Phenotypic analyses
Descriptive and reliability statistics are reported in Table 1. Zero-order correlations between CU traits and measures of antisocial behavior were similar in magnitude, .30 (with aggression) to .43 (with rule-breaking), and significantly different from zero at p(two-tailed) < .001. These correlations are reported below the diagonal in Table 1. Partial correlations controlling for the main effects of race, age, sex, and age × sex interaction are reported above the diagonal. These correlations are slightly attenuated relative to the zero-order correlations, although the pattern of correlations remains unchanged.
Bivariate quantitative genetic analyses
Based on the results of previous behavior genetic studies of CU traits (Moore et al., 2017), rule-breaking and aggressive behavior (Burt, 2009), variance in CU traits was modeled as a function of latent additive genetic (A) and nonshared environmental factors (E), and variance in antisocial behavior was modeled as a function of latent additive genetic (A), shared environmental (C), and nonshared environmental (E) factors. For each twin, we performed a Cholesky decomposition such that the second phenotype (i.e., antisocial behavior) was regressed on the latent genetic and environmental factors of the first phenotype (i.e., CU traits). This model is depicted in the top panel of Figure 1. (For details regarding the specification of twin models, see Neale and Maes [2004].)
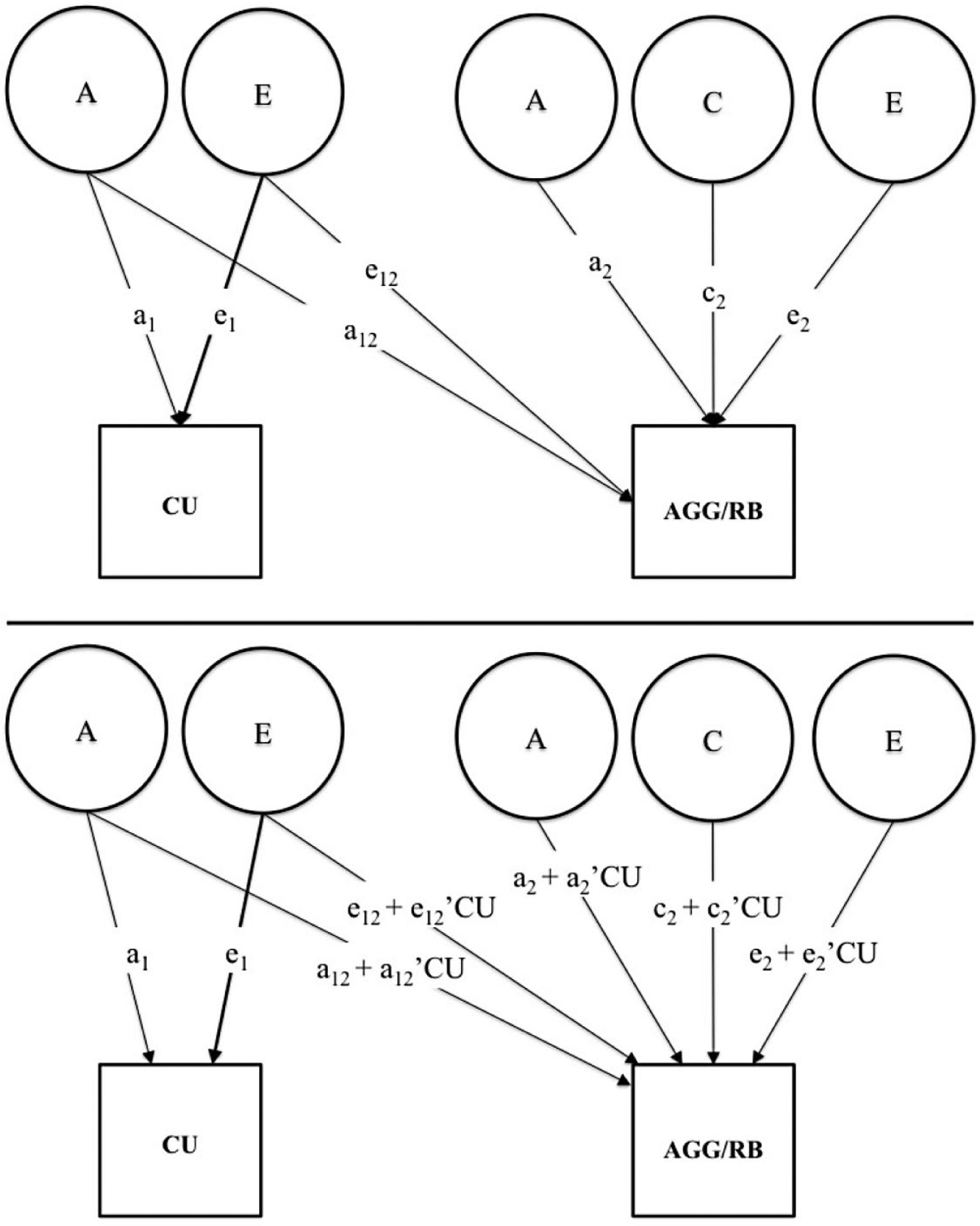
Path diagram for bivariate quantitative genetic models. Main effects models (top panel) were used to estimate latent genetic and environmental correlations. Moderation models (bottom panel) were used to test gene × trait interactions.
Parameter estimates from the Cholesky decompositions were used to compute additive genetic and nonshared environmental correlations between focal study constructs. The genetic correlation between CU traits and aggression was moderate in magnitude (rA = .38, 95% CI [.14, .62], p(two-tailed) = .002). On the other hand, the genetic correlation between CU traits and rule-breaking was large (rA = .80, 95% CI [.27, 1.33], p(two-tailed) = .003), similar in magnitude to the genetic correlation between aggression and rule-breaking (rA = .77, 95% CI [.60, .95], p(two-tailed) < .001). Nonshared environmental correlations between CU traits, aggression, and rule-breaking were comparatively small in magnitude. Nonetheless, nonshared environmental correlations between CU traits and aggression (rE = .23, 95% CI [.08, .38], p(two-tailed) = .002), CU traits and rule-breaking (rE = .30, 95% CI [.15, .45], p(two-tailed) < .001), and aggression and rule-breaking (rE = .30, 95% CI [.16, .43], p(two-tailed) < .001) were all significantly different than zero.
To test the hypothesis that antisocial behavior is more highly heritable in the presence of high levels of CU traits, compared to low levels of CU traits (i.e., a gene × trait interaction), bivariate twin models were specified to include a series of parameter constraints (see bottom panel of Fig. 1). Specifically, latent genetic and nonshared environmental paths from CU traits to antisocial behavior, as well as the genetic, shared, and nonshared environmental paths unique to antisocial behavior were specified to interact with individual differences in CU traits.
Models that included interaction terms were preferred over models with main effects only (see Table S1). Parameter estimates from gene × trait interaction models are reported in Table 2. Several results are noteworthy. First, CU traits significantly moderated (p = .001) additive genetic variance in aggression, such that aggression was more highly heritable among adolescents high on CU traits. Specifically, variance in aggression was 64% heritable for adolescents high (+2 SD) on CU traits. In contrast, for adolescents low (–2 SD) on CU traits, variance in aggression was only 9% heritable; for adolescents with average levels of CU traits, aggression was 34% heritable. Second, we found evidence that CU traits moderate shared environmental variance in rule-breaking (p < .001), such that shared environmental influences are magnified among adolescents with high levels of CU traits. For adolescents high (+2 SD) on CU traits, environments shared by siblings living in the same home (i.e., shared environments) accounted for 31% of the variance in rule-breaking. In contrast, for adolescents low (–2 SD) on CU traits, shared environmental variance in rule-breaking approximated zero. For adolescents with average levels of CU traits, shared environmental influences accounted for 18% of the variance in rule-breaking.
Parameter Estimates From Bivariate Gene × Trait Interaction Models
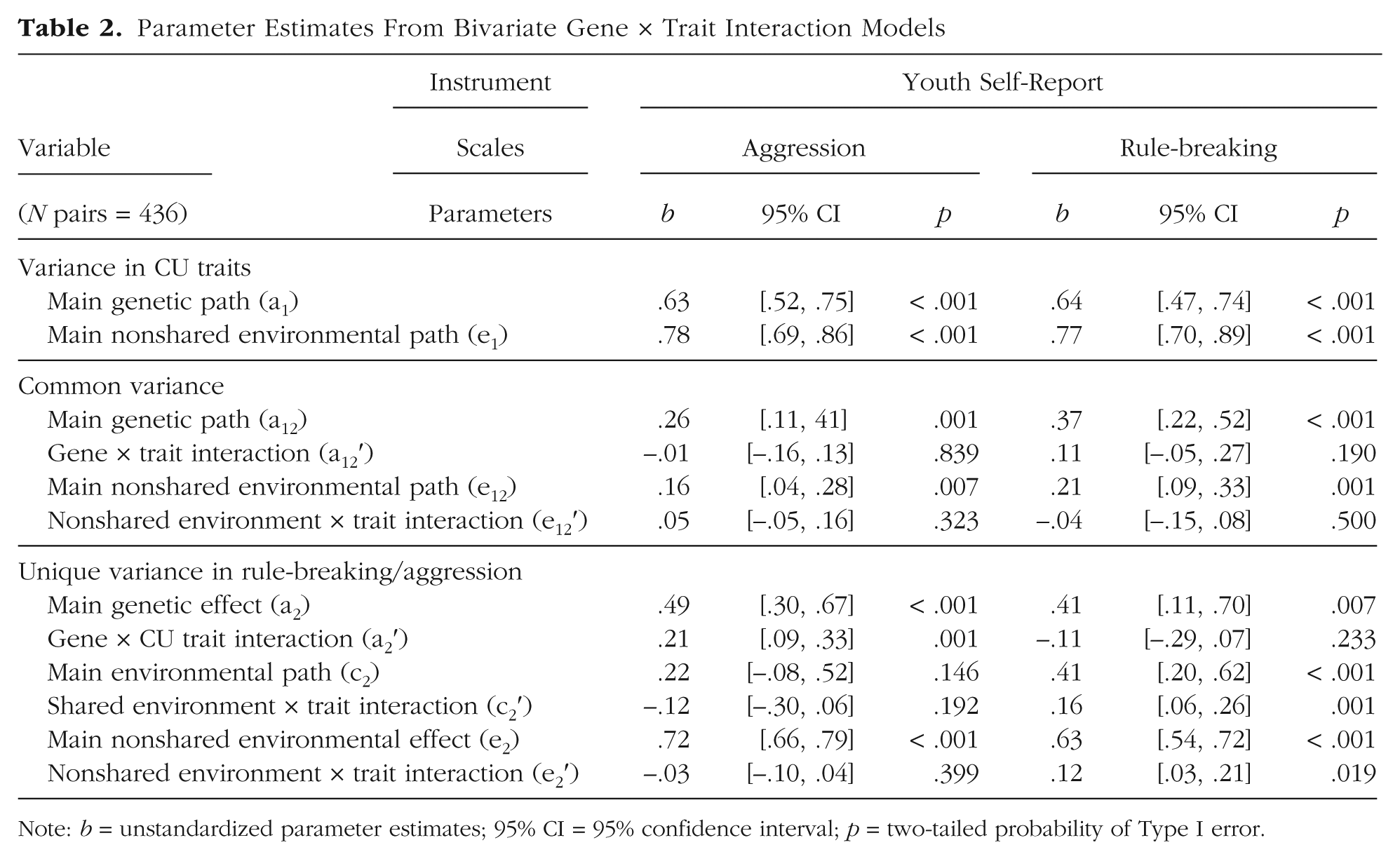
Note: b = unstandardized parameter estimates; 95% CI = 95% confidence interval; p = two-tailed probability of Type I error.
Third, CU traits moderated nonshared environmental variance in rule-breaking (p < .05), such that nonshared environmental influences are also magnified among adolescents with high levels of CU traits. Specifically, for adolescents high (+2 SD) on CU traits, environments uniquely experienced by siblings accounted for 46% of the variance in rule-breaking. On the other hand, for adolescents low (–2 SD) on CU traits, nonshared environmental influences accounted for 35% of the variance in rule-breaking. The magnitude of nonshared environmental influences for adolescents with high levels of CU traits was similar to adolescents with average levels of CU traits, accounting for 48% of the variance in rule-breaking. To help visualize moderation effects, the total additive genetic (A), shared environmental (C), and nonshared environmental (E) variance in aggression and rule-breaking were plotted across the continuous range (±2 SD) of CU traits (see Fig. 2).
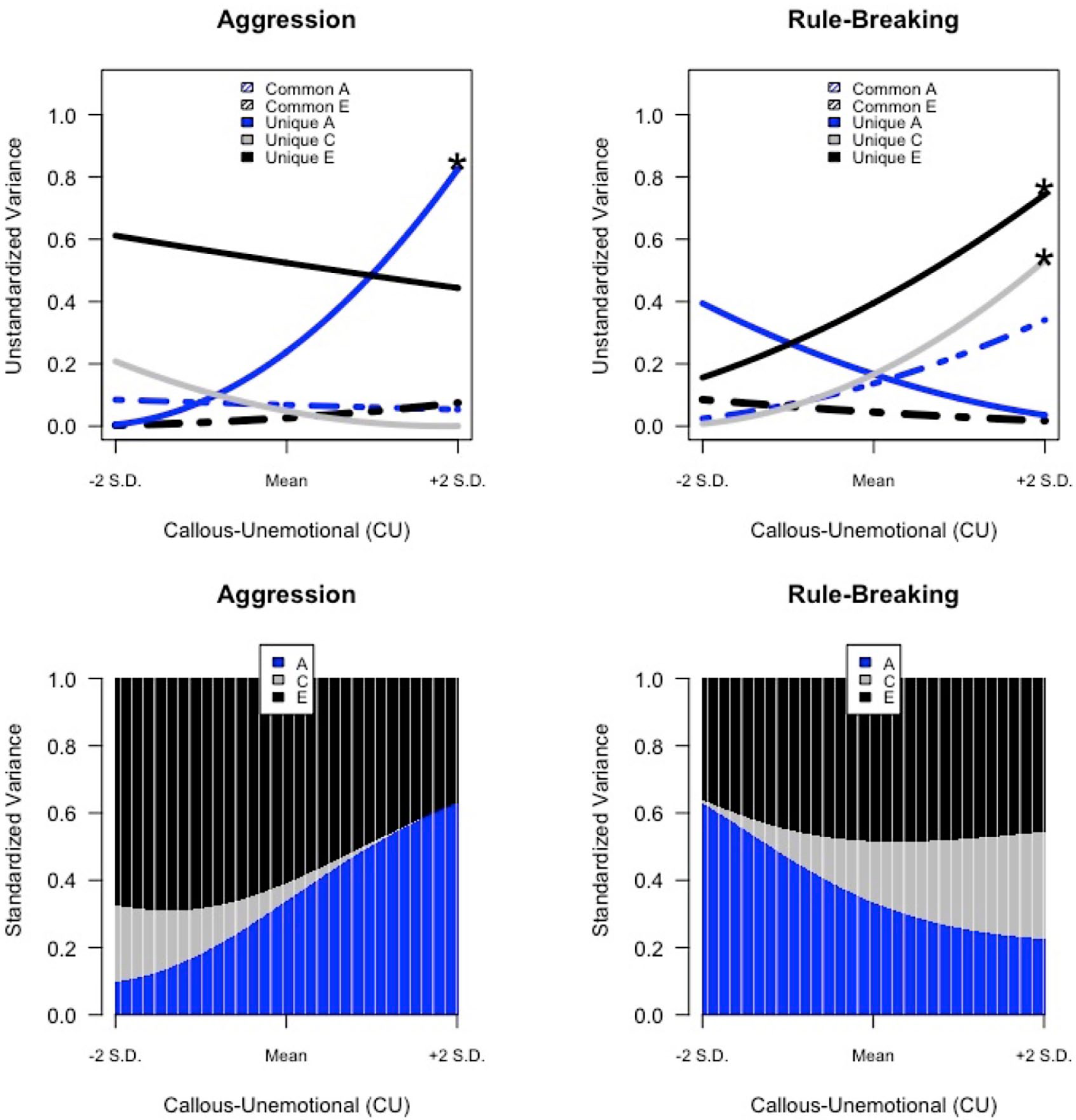
Gene × callous-unemotional trait interaction effects on antisocial behavior. Additive genetic (blue), shared environmental (gray), and nonshared environmental (black) variance in aggression and rule-breaking are plotted across the continuous range (±2 SD) of callous-unemotional traits. Top panel: dashed and solid lines represent unstandardized estimates of variance in antisocial behavior that are common to and unique of callous-unemotional traits, respectively. Asterisks mark components of variance significantly moderated by CU traits. Bottom panel: shaded regions represent portions of total variance in aggression and rule-breaking. Portions of variance calculated from parameter estimates reported in Table 2 using the following equations:
Multivariate quantitative genetic analyses
Consistent with results of the bivariate quantitative genetic models reported earlier, a previous meta-analysis of twin and sibling studies indicated that genetic influences on aggression overlap with genetic influences on rule-breaking (Burt, 2013). Also consistent with previous studies, rule-breaking, and aggression evinced a moderate phenotypic correlation in the current study (r = .50). Therefore, similar to Harden et al. (2015), we fit a multivariate quantitative genetic, common, and specific factor model to decompose variance shared between, and unique to, aggression and rule-breaking into latent biometric components (i.e., A, C, E factors). This model clarifies whether the gene × CU trait interaction documented in the current study operates on variance unique to aggression, or variance common to aggression and rule-breaking. Similarly, this model also clarifies whether the environment × CU trait interaction on rule-breaking is on variance common to, or independent of, aggression.
Parameter estimates from this model are reported in Table S2. Several results are noteworthy (see Fig. S1 in the Supplemental Material available online). Only additive genetic variance unique to aggression was significantly moderated by CU traits (b = .196, SE = .063, p = .002); additive genetic variances unique to rule-breaking, and common to rule-breaking and aggression, were not significantly moderated by CU traits (ps > .05). Shared-environmental variances unique to rule-breaking and aggression were not significantly moderated by CU traits (ps > .05). However, shared-environmental variance common to rule-breaking and aggression was significantly moderated by CU traits (b = .130, SE = .056, p = .022). Finally, nonshared environmental variance unique to rule-breaking was also significantly moderated by CU traits (b = .141, SE = .035 p < .001); nonshared environmental variances unique to aggression, as well as common to rule-breaking and aggression, were not significantly moderated by CU traits (ps > .05).
Discussion
The current study replicates and extends previous research on the association between CU traits and antisocial behavior. First, we find evidence that CU traits index genetic liability for aggressive and nonaggressive expressions of antisocial behavior in adolescence. We also find evidence that higher levels of CU traits amplify additive genetic influences on aggressive behavior, as has been previously found in samples of children using composite indices of antisocial behavior, which collapse across the aggression versus rule-breaking distinction. Moreover, results provide evidence that aggressive and (nonaggressive) rule-breaking have distinct etiologies, as CU traits did not moderate genetic influences on rule-breaking behavior. Rather, environmental influences on rule-breaking were heightened among adolescents with high levels of CU traits, such that the heritability of rule-breaking was actually lower at higher levels of CU. Results of the multivariate model, which accounted for the shared etiology between rule-breaking and aggression, were largely consistent with the results of the bivariate models in which rule-breaking and aggression were analyzed independently of each other. Both modeling procedures indicated that genetic influences on aggression (but not rule-breaking), and nonshared environmental influences on rule-breaking (but not aggression) were positively moderated by CU traits. The multivariate model did, however, reveal that shared environmental influences common to both rule-breaking and aggression were positively moderated by CU traits.
These findings are broadly consistent with the results of a meta-analysis (Burt, 2013), which indicates that genetic and environmental influences on rule-breaking are partly distinct from those operating on aggression. The current results from gene × trait interaction models suggest that CU traits further contribute to the rule-breaking versus aggression distinction. The finding that environmental influences on rule-breaking were significantly heightened among adolescents with high levels of CU traits was not predicted. Consequently, this finding should be considered tentative prior to replication. Nevertheless, to speculate on this unexpected finding, there is considerable evidence that adolescents who engage in risky behavior, including delinquency, do so in the presence of peers (Boivin, Vitaro, & Poulin, 2005; Bukowski, Brendgen, & Vitaro, 2006; Monahan, Steinberg, & Cauffman, 2009; Silva, Chein, & Steinberg, 2016). Moreover, there is evidence that adolescents who engage in minor acts of delinquency are rated as more popular than their rule-abiding peers (Allen, Porter, McFarland, Marsh, & McElhaney, 2005) and unique variance in rule-breaking, after accounting for shared variance with aggression, is actually associated with higher levels of emotional stability (Tackett et al., 2013). Conversely, hostile aggressive behaviors, which may lead to more serious antisocial offenses, are less likely to be rewarded by peers (Allen et al., 2005) and are associated with higher levels of negative affectivity and disagreeableness (Tackett et al., 2013). In sum, there appears to be a strong social component that may be unique to rule-breaking expressions of antisocial behavior. The current study suggests that adolescents with a limited capacity to express prosocial emotions are especially susceptible to environmental sources of variation in rule-breaking behavior, specifically environments that are uniquely experienced by siblings living in the same home. Individual differences in peer groups may provide a nonheritable pathway through which CU traits contribute to risk for nonaggressive expressions of antisocial behavior. Although the current study did not examine individual differences in peer groups, future research efforts may benefit from exploring these and related constructs as they relate to CU traits and the etiology of antisocial behavior.
It also remains an open question as to whether intervention programs can leverage these distinct patterns of social influence on aggressive and nonaggressive behavior to develop more effective strategies for deterring different expressions of antisocial behavior. The present study is particularly notable in identifying underlying CU mechanisms interacting with etiological factors to account for individual differences in rule-breaking behaviors, which have often been characterized as more normative than aggression. Although CU traits are typically thought to be a heritable risk factor for severe expressions of antisocial behavior, high levels of CU traits may contribute to less severe expressions of antisocial behavior by magnifying the salience of environmental pathways. Future research and intervention efforts stand to benefit from identifying the specific environmental mechanisms that account for the heightened latent environmental influences operating on rule-breaking documented in the current study.
CU traits’ magnification of genetic risk for aggressive behavior is consistent with the general consensus that CU traits “designate a particularly severe and aggressive subgroup within those with serious conduct problems” (Frick et al., 2014, p. 3). However, the results of the current study should not be interpreted to suggest that CU traits only confer risk for aggressive, as opposed to nonaggressive antisocial behavior. Rather, results highlight distinct etiological pathways, both heritable and environmental, through which CU traits confer risk for different expressions of antisocial behavior. Indeed, CU traits were as highly correlated with rule-breaking as they were with aggression—a finding that has been observed in other samples, too (Burt, 2013). This pattern of phenotypic associations appears to be invariant to important sample characteristics, including severity of antisocial offending; documented in the current study using a population-representative sample of adolescents, rule-breaking was more highly correlated with CU than aggression (|Δr| = .13, SE = .04, p = .006), similar to associations observed in a sample of adjudicated adolescents (Kimonis et al., 2008). Although CU traits are most commonly thought of in relation to cruelty or callousness toward a victim, violating societal norms and rules—even violations that do not obviously harm another person—is also facilitated by a disregard for other people. For youth with low levels of CU traits, affectively charged interpersonal concerns—such as fears about how one’s relationships with close or valued others (parents, teachers, friends) will be affected by one’s behavior—might discourage rule-breaking.
The gene × trait interaction documented in the current study may also shed light on the problem of missing heritability (DeYoung & Clark, 2012; Eichler et al., 2010; Manolio et al., 2009), which refers to the lack of correspondence between molecular genetic and quantitative genetic estimates of heritability. Molecular genetics techniques, specifically genome-wide complex trait analysis (GCTA; Yang, Lee, Goddard, & Visscher, 2011), can estimate the amount of variance in a phenotype collectively explained by thousands of measured variants across the genome. Traditional quantitative genetic techniques, alternatively, estimate heritability based on the known genetic relatedness of relatives. It is interesting that GCTA has failed to reproduce the heritability estimates of childhood behavior problems and associated traits found using quantitative genetic methodology. However, similar to gene × environment interactions, if the genetic underpinning of a phenotype varies across different levels of an associated phenotype, then efforts to identify genetic main effects will remain difficult, unless such moderating phenotypes are taken into account. Results of the current study suggest that CU traits are an example of such a phenotype.
There are a number of limitations to the current study. First, CU and antisocial constructs were measured using self-reports only. Furthermore, the sample used in the current study (N = 824) was moderate in size for quantitative genetic analyses of gene-by-trait interaction. This limitation is evident in the range of confidence intervals surrounding parameter estimates in quantitative genetic interaction models. The precision of the current results would, therefore, stand to benefit from replication using a larger sample. Although previous behavior genetic research has found evidence for a gene × CU trait interaction on antisocial behavior within tightly circumscribed ages (i.e., 7- and 9-year-old children), the current study documented a gene × CU trait interaction in a sample of adolescents ranging from 13 to 20 years old. It remains an open question whether the gene × CU trait interaction documented in the current study are further nuanced by developmental differences that manifest across adolescence.
There are also a number of strengths to the current study. First, to help overcome limitations of previous research, a well-validated questionnaire with high internal consistency was used to measure CU traits. Second, the majority of quantitative genetic studies on CU traits and antisocial behavior have analyzed racially homogenous samples. The current study used a racially and socioeconomically diverse sample, taking care to residualize for main effects of race prior to model fitting. We also used well-validated instruments to measure different subtypes of antisocial behavior and data analytic procedures that modeled a continuous relationship between CU traits and antisocial behavior. As interdisciplinary research continues to elucidate the multilevel cascade of biological processes that bridge the gap between polygenic risk and psychopathology, CU traits continue to be helpful in further organizing and delineating the constellation of physiological, neurological, and cognitive mechanisms that account for heterogeneity in the etiology of antisocial behavior.
Footnotes
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.