
Abstract
Depressed individuals exhibit biased attention to negative emotional information. However, much remains unknown about (a) the neurocognitive mechanisms of attention bias (e.g., qualities of negative information that evoke attention bias or functional brain network dynamics that may reflect a propensity for biased attention) and (b) distinctions in the types of attention bias related to different dimensions of depression (e.g., ruminative depression). Here, in 50 women, clinical depression was associated with facilitated processing of negative information only when such information was self-descriptive and task-relevant. However, among depressed individuals, trait rumination was associated with biases toward negative self-descriptive information regardless of task goals, especially when negative self-descriptive material was paired with self-referential images that should be ignored. Attention biases in ruminative depression were mediated by dynamic variability in frontoinsular resting-state functional connectivity. These findings highlight potential cognitive and functional network mechanisms of attention bias specifically related to the ruminative dimension of depression.
Cognitive models of depression posit that negative beliefs about the self are central to depressive disorders, driving negative interpretations and automatic thinking that bias goal-directed attention (Beck, 2008). Such models have received robust support and informed the development of psychosocial interventions focused on redirecting attention in the presence of negative cognitions (Eisendrath et al., 2016; Hollon & Ponniah, 2010). However, clinical research has also demonstrated a lack of precision in our understanding of cognitive or neural mechanisms of attention biases. In particular, evidence is mixed regarding the specific domains of attention that are biased in depression, qualities of negative information that bias attention, and functioning of neural systems that may be associated with a propensity toward biased attention. Furthermore, there is considerable heterogeneity in the cognitive biases depressed individuals exhibit (Everaert, Koster, & Derakshan, 2012). One source of this heterogeneity in attention biases may be heterogeneity in depression phenotypes. That is, dimensional features such as trait rumination, which vary across individuals with depression, may interact with mood to produce distinct profiles of attention bias. The current study thus examines how to gain a better understanding of the specificity and nature of attention biases related to ruminative depression, a critical step for more precisely characterizing mood disorder at an individual level.
Understanding Component Mechanisms of Attention Bias in Depression
Meta-analytic evidence indicates that depression (current or past diagnosis or elevated symptoms of depression) is associated with slowed responses when naming the ink color of emotionally negative words and speeded responses to negative targets or cues that are spatially congruent with negative words or images (Epp, Dobson, Dozois, & Frewen, 2012; Peckham, McHugh, & Otto, 2010; Winer & Salem, 2016). Neuroimaging studies provide converging evidence for attention biases in depression, showing, for example, that elevated symptoms of depression are related to increased activity and functional connectivity among prefrontal cognitive systems and midline regions involved in self-directed attention in response to negative distractors on an emotion word Stroop (Kaiser, Andrews-Hanna, Spielberg, et al., 2015). These findings provide evidence for attention biases in depression and suggest important distinctions in the impact of biases on performance (e.g., enhanced performance when negative information is consistent with task goals but impaired performance when negative information is inconsistent with goals). However, effect sizes have been inconsistent across meta-analyses and individual studies, suggesting that attention biases may be less reliable or more complex than originally suspected.
To address mixed findings for attention bias in depression, more recent theories point to established models in cognitive neuroscience emphasizing that attention is not a unitary construct but includes subprocesses such as orienting, selecting, engaging, and disengaging from stimuli (Petersen & Posner, 2012; Posner & Boies, 1971), which may be differentially associated with depression. One theory posits that attention biases in depression are specifically active at later stages of processing, such as facilitating elaboration of (and difficulty disengaging from) negative information once it has captured attention (De Raedt & Koster, 2010). This idea is supported by evidence that individuals with depression show attention biases for negative information when such information is either presented at longer (> 500 ms) but not shorter (< 250 ms) durations in the dot-probe paradigm or followed by longer (≥ 1,300 ms) but not shorter (< 250 ms) delays to a target in the exogenous cueing paradigm (Koster, De Raedt, Goeleven, Franck, & Crombez, 2005; Koster, De Raedt, Leyman, & De Lissnyder, 2010; Mogg, Bradley, & Williams, 1995; Sylvester, Hudziak, Gaffrey, Barch, & Luby, 2016). In addition, the idea that attention biases are specific to later stages of processing is supported by evidence that depression is related to increased dwell time looking at negative material but no differences in initial orienting (Caseras, Gamer, Bradley, & Mogg, 2007; Leyman, De Raedt, Vaeyens, & Philippaerts, 2011; Matthews & Antes, 1992). However, meta-analyses have yielded equivocal support for attention biases at later (elaborated) as well as earlier (orienting) stages of processing in depression or failed to find an association between attention bias and stimulus duration (Armstrong & Olatunji, 2012; Peckham et al., 2010). Thus, whereas important distinctions may exist in the attention subprocesses that are biased in depression, evidence for those distinctions is not yet conclusive.
In addition to distinguishing which domains of attention are biased in depression, a complementary goal is to distinguish the types of information that evoke such bias: What is it about negative emotional information that captures or holds attention? At least three potential answers exist for this question. One potential answer is that depressed individuals are drawn toward negative emotional content because it matches their current mood state. This mood-congruence hypothesis is supported by evidence that experimentally induced negative mood in healthy individuals can induce attention biases toward negative material that are similar to those exhibited in depression (Bradley, Mogg, & Lee, 1997; Gilboa-Schechtman, Revelle, & Gotlib, 2000; Gotlib & McCann, 1984; Isaac et al., 2012; Ridout, Noreen, & Johal, 2009). However, these effects have not been replicated consistently (Chepenik, Cornew, & Farah, 2007; McCabe, Gotlib, & Martin, 2000; Newman & Sears, 2015). In addition, attention biases in depression have been observed with other forms of negative information (e.g., anger) that are putatively unrelated to mood state (Lonigan & Vasey, 2009; Mogg et al., 1995; Oehlberg, Revelle, & Mineka, 2012; Platt, Murphy, & Lau, 2015). Together, these findings suggest that depression-related attention biases are not exclusively explained by mood congruency.
A second explanation is that depressed individuals are more sensitive to self-referential information (regardless of emotional content) and biases toward negative information are coincident to the fact that depressed individuals happen to have a self-concept that is more negative than nondepressed individuals have. Across clinical and nonclinical samples, self-relatedness of stimuli has been shown to facilitate recall and perceptual integration of information (reviewed in Sui & Humphreys, 2015) and boost activity and functional connectivity in brain systems including medial prefrontal cortex (MPFC), insula, hippocampus, and areas of anterior and posterior cingulate cortex (ACC, PCC; Craik et al., 1999; Fossati et al., 2003; Macrae, Moran, Heatherton, Banfield, & Kelley, 2004; Murray, Debbane, Fox, Bzdok, & Eickhoff, 2015; Murray, Schaer, & Debbane, 2012). These neural systems, many of which are grouped in a functional network known as the default network, show increased functional connectivity during autobiographical thinking (Young, Siegle, Bodurka, & Drevets, 2016) and other forms of self-focused attention (reviewed in Qin & Northoff, 2011). Critically, default network and frontoinsular regions also exhibit resting-state hyperconnectivity in major depression (Kaiser, Andrews-Hanna, Wager, & Pizzagalli, 2015). Although caution to avoid reverse inference is warranted when interpreting these converging patterns, one theory is that amplified activity and coordination among frontoinsular-default networks is a marker of attention biases toward self-focused thinking in depressed individuals.
A third explanation for negative attention biases in depression points to the interaction between self-relatedness and emotional valence of information: for example, that attention is biased toward positive (but not negative) self-referential information in healthy people and negative (but not positive) self-referential information in depressed people. Consistent with this assumption, research in healthy individuals has demonstrated faster judgments of and increased medial prefrontal activity in response to positive compared with negative self-referential information (Moran, Macrae, Heatherton, Wyland, & Kelley, 2006; Watson, Dritschel, Obonsawin, & Jentzsch, 2007). In contrast, depressed individuals exhibit a reversed pattern of reduced prefrontal and hippocampal response to positive self-referential information (Quevedo et al., 2016) and an amplified response to negative self-referential information (Macdonald & Kuiper, 1985; Shestyuk & Deldin, 2010). These findings complement evidence for enhanced self-focused attention and frontoinsular-default network activity in depression but suggest that the combination of self-relatedness and negative emotionality is responsible for evoking attention bias.
The outlined hypotheses have been tested only partially because although many experiments have manipulated the emotional content of stimuli, few have directly manipulated the self-referential quality of stimuli (reviewed in Wisco, 2009). Instead, researchers have commonly defined the self-referential nature of stimuli post hoc on the basis of task performance (e.g., reaction time or neural response to unselected emotional words while words are being judged on their self-referential quality; Alloy, Abramson, Murray, Whitehouse, & Hogan, 1997; Connolly, Abramson, & Alloy, 2016; Gencoz, Voelz, Gencoz, Pettit, & Joiner, 2001). Although this approach has merit, the same response biases that are of interest in this research can also confound the comparison of depressed and nondepressed participants on specific categories of self-referential information; for example, healthy individuals may endorse few negative words as self-descriptive, whereas depressed individuals may endorse many negative words as self-descriptive, yielding unbalanced sets of stimuli for further experimentation or statistical analysis (discussion in Connolly et al., 2016). In sum, research optimized for testing attention to emotionally negative (or positive) and self-descriptive (or non–self-descriptive) information is needed to understand the qualities of emotional information that evoke attention bias.
Attention Bias Across Clinical Phenotypes: Rumination and Depression
Experimental paradigms designed to unpack the neurocognitive mechanisms of attention bias are necessary to refine our understanding of how and when attention biases occur in depression. However, when such research is conducted exclusively using categorical case-control designs, it could be insufficient for understanding individual differences in attention bias. Depression is a complex and heterogeneous family of disorders, with varying symptom presentations, etiologies, and functional impairments exhibited across individuals (Kaiser, 2017). Thus, attention biases may not characterize different depressed individuals to the same extent. Here, the dimension of trait rumination, defined by the tendency toward negative and repetitive self-focused thinking (Nolen-Hoeksema, Wisco, & Lyubomirsky, 2008), could be particularly relevant. Consistent with this notion, nondepressed individuals prone to rumination exhibit attention biases that overlap with those observed in depressed individuals (Beckwe & Deroost, 2016; Hilt & Pollak, 2013), and higher levels of rumination among depressed individuals are associated with more extreme attention biases (Donaldson, Lam, & Mathews, 2007). This convergence suggests the possibility that trait rumination may explain or exacerbate attention biases associated with depression.
However, not all depressed individuals are prone to rumination, and not all individuals prone to rumination are depressed. Accordingly, it is possible that different symptom dimensions interact to produce distinct phenotypes of depression characterized by unique profiles of attention bias. In particular, whereas both depression and trait rumination have been linked to biased elaboration (or difficulty disengaging from) negative information (Joormann, Levens, & Gotlib, 2011; Joormann, Nee, Berman, Jonides, & Gotlib, 2010; Kaiser, Andrews-Hanna, Metcalf, & Dimidjian, 2015) separately, some evidence suggests that depressed ruminators also exhibit biases orienting to or selecting negative information (De Lissnyder, Derakshan, De Raedt, & Koster, 2011; Joormann, Dkane, & Gotlib, 2006; Whitmer & Banich, 2007). Thus, depressed ruminators may be characterized uniquely by both preferential attention to and elaboration of negative self-referential thoughts (related discussion in Everaert et al., 2012). Such biases match the clinical profile of ruminative depression (i.e., elevated trait rumination co-occurring with depression), in which rumination is experienced as intrusive and difficult to escape (Papageorgiou & Wells, 2001).
On the level of brain functioning, ruminative depression has been associated with increased resting-state functional connectivity (RSFC) among regions of the default network (Berman et al., 2011) and highly variable RSFC between MPFC regions of default network and anterior insula (Kaiser et al., 2016). As noted previously, the default network comprises midline, inferior temporal, and parietal regions involved in self-generated, self-focused, or autobiographical thinking (Andrews-Hanna, Smallwood, & Spreng, 2014), whereas the anterior insula is considered to be a hub of the “salience network” involved in allocating resources to other networks (including default network) on the basis of salient cues or thoughts (Menon & Uddin, 2010; Sridharan, Levitin, & Menon, 2008). Prior research suggests that variability in cross-network RSFC may be related to regulatory relationships in which key regions of one network are engaged in up- or downregulating activity in a second network at rest or in response to cognitive demands (Hutchison et al., 2013; Hutchison & Morton, 2015). Thus, increased variability in RSFC between regions of the salience (insula) and default (MPFC) networks may reflect an individual’s heightened tendency to recruit these cross-network regulatory systems, (i.e., an increased tendency for insula to be engaged to up- or down-regulate activity in default network; Sridharan et al., 2008). On the level of cognitive processing, such increased frontoinsular variability could reflect a tendency for biased allocation of resources toward self-focused thinking or efforts to regulate self-focused thinking. Together, this suggests a model in which ruminative depression is related to attention biases via altered functioning of frontoinsular and default network regions.
Present Study
The present study’s purpose is to provide insight into the cognitive mechanisms and functional network correlates of depression and, in particular, the ruminative dimension of depression. Toward this goal, we developed a behavioral task designed to manipulate the self-referential quality and emotional valence of information separately and vary the depth of elaboration for (self-referential or emotional aspects of) information (for descriptions of other tasks with partially overlapping procedures, see Elliott, Rubinsztein, Sahakian, & Dolan, 2000; Etkin, Egner, Peraza, Kandel, & Hirsch, 2006). In this task, participants judged either the self-descriptiveness or the emotional valence of words (preselected to be self-descriptive or non–self-descriptive, crossed by negative or positive emotion) while ignoring background images that were self-referential (own face) or non–self-referential (other face). To complete the task, participants must always elaborate on the content of the word, although the level of elaboration of specific features of the word (self-descriptiveness or valence) depends on the relevance of that feature to task goals (e.g., self-descriptiveness should be more deeply processed when the goal is to judge self-descriptiveness than when the goal is to judge emotional valence). Simultaneously, participants should ignore the self-referential content of the images entirely. Together, performance biases related to task-relevant word content are interpreted as evidence for later-stage attention biases (difficulty disengaging from elaborated information), whereas performance biases related to task-irrelevant word content or images are interpreted as earlier-stage attention biases (difficulty ignoring information that should not be deeply elaborated [irrelevant word content] or toward which attention should not be oriented at all [images]).
This design allowed us to test several competing hypotheses. First, regarding the type of information that evokes attention biases in depression or ruminative depression, if attention biases in this task are evoked by mood congruence, facilitated performance (faster response speed) should be observed when judging negative words regardless of word self-descriptiveness (Hypothesis 1a). However, if attention biases in this task are evoked by self-descriptiveness, facilitated performance should be observed when judging self-descriptive words regardless of word valence (Hypothesis 1b). Alternately, if attention biases in this task are evoked by the conjunction of negative emotion and self-descriptiveness, facilitated performance should specifically be observed when judging negative, self-descriptive words (interaction of word valence by self-descriptiveness; Hypothesis 1c). Second, regarding the type of attention processing that is biased in depression or ruminative depression, if attention biases are specific to later stages of processing (information that is elaborated), performance facilitation effects on this task will be strongest when task goals (e.g., judge self-descriptiveness) match the type of stimulus content that evokes attention bias (e.g., self-descriptive word) (Hypothesis 2a). In contrast, if attention biases are also evident at earlier stages of processing, performance facilitation would also be observed when bias-evoking content was task-irrelevant (e.g., judging emotional valence of self-descriptive word) or should be ignored (e.g., judging words while an image of yourself is presented in the background; Hypothesis 2b).
In addition to behavioral measures, we collected resting-state neuroimaging data to evaluate the associations between depression and the dimension of ruminative depression, attention biases, and intrinsic functional connectivity of a frontoinsular circuit linking medial prefrontal regions of the default network and areas of insula. The goals of this analysis were to replicate prior findings of increased dynamic variability in frontoinsular RSFC in ruminative depression and test the novel hypothesis that frontoinsular network functioning mediates an association between ruminative depression and attention bias (Hypothesis 3).
Method
Participants
Participants included 53 unmedicated adult women recruited from the Boston metropolitan area who either reported current major depressive disorder (MDD group n = 31) or no history of depression or other Axis I psychiatric diagnoses (healthy control [HC] group n = 22). Participants were restricted to women because of evidence that rumination is especially prevalent in women (Nolen-Hoeksema, 1994). All participants completed a Structured Clinical Interview to evaluate psychiatric history (First, Spitzer, Gibbon, & Williams, 2002). Inclusion in the MDD group required a primary diagnosis of major depression; participants were excluded from this group for lifetime history of substance dependence, psychosis, mania, anorexia, or recent history of substance abuse (past 12 months) or bulimia (past 2 years; see Table S1 in the Supplemental Material available online). Participants recruited to the HC group were excluded for any lifetime psychiatric illness. For both groups, participants were excluded on the basis of history of neurological impairment, head injury, MRI counter-indications, or cognitive or language impairments that interfered with the ability to complete behavioral testing. Groups did not differ on age, and both groups reflected the geographic region in terms of race and ethnicity and education (Table S1 in the Supplemental Material).
Procedures
The study consisted of experimental procedures spanning two sessions approximately 4 weeks apart (average = 27.47 days). In the first session, participants provided ratings of self-descriptiveness from a list of adjectives, and photographs of the participants were taken to create individualized stimuli for the self-referential information processing (SIP) task. Participants also completed a series of self-report questionnaires at this session. In the second session, participants completed the SIP task (1 MDD participant did not complete the emotion judgment condition of this task because of technical error), followed by an MRI scan. Of note, the present sample was drawn from an ongoing study with distinct, nonoverlapping experimental objectives and analyses that will be reported elsewhere (see the Supplement Material). All procedures were approved by the Institutional Review Board at Partners Healthcare and McLean Hospital and conducted in accordance with the provisions of the World Medical Association Declaration of Helsinki. Participants were reimbursed for their participation and travel and fully debriefed and provided (on request) with referral information for sliding-scale psychological services in the area.
Measures
Self-referential information processing
The SIP task includes a set of individualized stimuli consisting of self-descriptive (positive or negative) or non–self-descriptive (positive or negative) adjectives that are superimposed onto images of either the participant’s own face or a gender- and race-matched face of another person (Fig. 1). For the emotion judgment version of the SIP (SIP-EJ), the goal is to respond as quickly as possible to identify the valence of the word while ignoring the background image. For the self-descriptiveness judgment version of the SIP (SIP-SJ), the goal is to respond as quickly as possible to indicate whether the word is self-descriptive while ignoring the background image. See the Supplemental Material for replication of task effects in an independent sample.
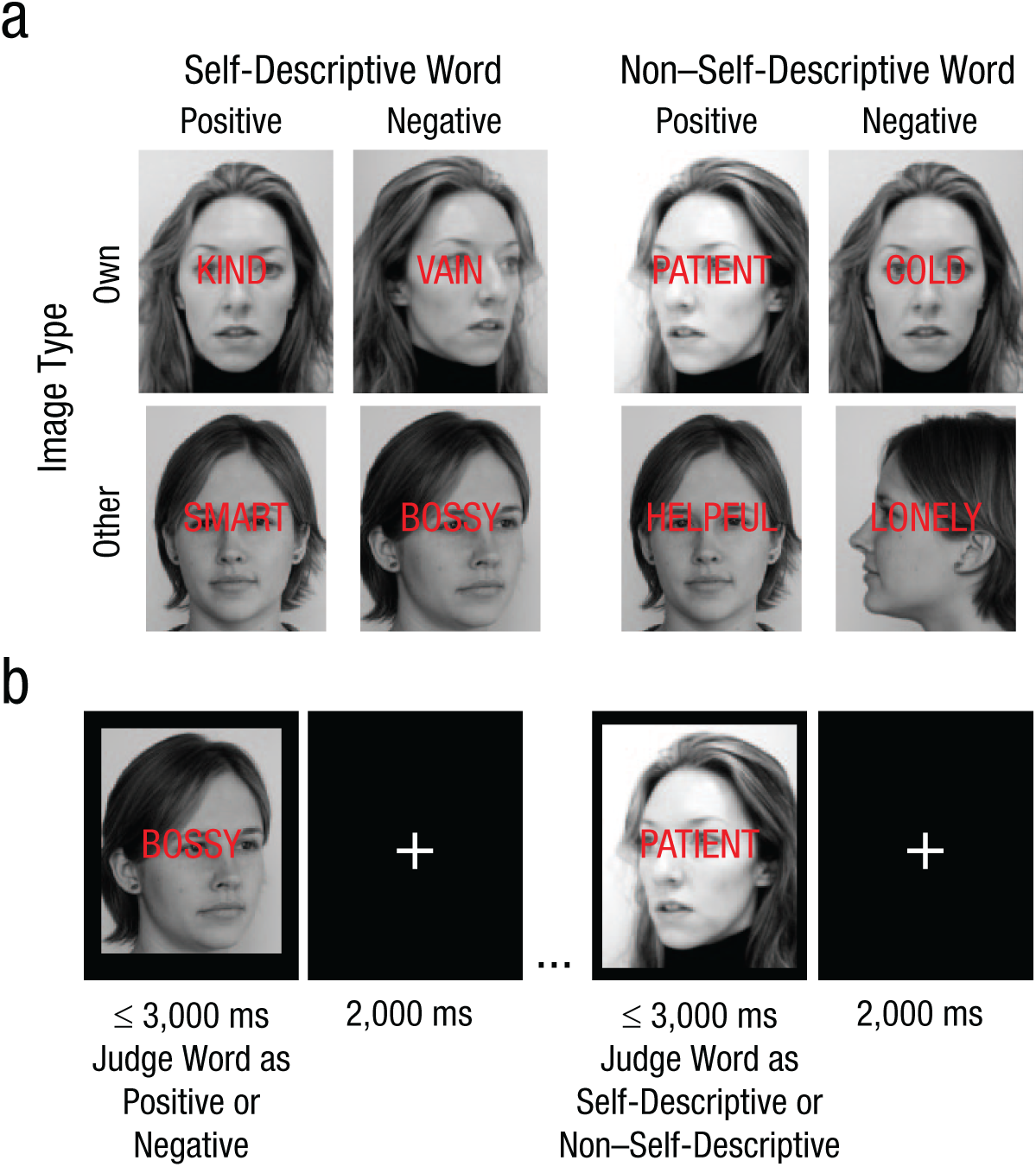
Self-referential information processing task design. The self-referential information processing (SIP) task was designed to test the effects of self-referential material on responses when judging either the emotional valence or self-descriptiveness of emotional words. Stimuli (a) were individualized to the participant and consisted of emotionally valenced self-descriptive (self-descriptive positive or self-descriptive negative) or non–self-descriptive (non–self-descriptive positive or non–self-descriptive negative) adjectives superimposed onto images of either the participant’s face (own) or a gender- and race-matched face of another person (other). Word stimuli were selected from a list of (320) positive and negative adjectives on the basis of the participant’s ratings in a previous session. There were (b) two task goal conditions. In one condition, the participant was instructed to judge the emotional valence of the word as quickly as possible; in the other condition, the participant was asked to judge whether the word was self-descriptive as quickly as possible. Thus, this task aimed to examine direct or moderated effects of self-referential or emotional content on information processing when such content is task-relevant (e.g., self-referential quality of words when judging word self-descriptiveness), task-irrelevant but attended (e.g., valence of words when judging self-descriptiveness), or task-irrelevant and should be ignored (self-referential quality of images across task conditions).
Individualized word stimuli
To create the individualized word stimulus set, we administered an adjective rating protocol to participants at the first session using E-Prime. Participants were presented serially with 320 positive and negative adjectives drawn from the Dumas word list (Dumas, Johnson, & Lynch, 2002) and rated each word on a 0 (not at all self-descriptive) to 9 (highly self-descriptive) scale. Participants were given these instructions: Rate each word on how self-descriptive it is of you, in general. Try to be as honest as possible in rating both the positive and negative traits that either do, or do not, describe you. Sometimes it helps to think about how someone who knows you well but who is as unbiased as possible would rate you—like a cousin, classmate, or work colleague.
Participants had up to 10 s to rate each word. After the participant completed the adjectives rating protocol, a subset of 96 words was selected from the list on the basis of participant ratings of self-descriptiveness balanced on valence (as determined by published norms; Dumas et al., 2002). The 96-word subset was selected to include approximately 24 words in each of four categories: self-descriptive positive words, self-descriptive negative words, non–self-descriptive positive words, and non–self-descriptive negative words (see Fig S1 in the Supplemental Material). Of note, 3 (HC) participants were unable to provide words in each category (e.g., either reported all positive words to be self-descriptive or all negative words to be non–self-descriptive) and therefore were removed from task analyses, yielding a final sample of n = 50.
Individualized image stimuli
To obtain individualized images, we took photographs of each participant’s face in five orientations to the camera (right and left 90° view, right and left 45° view, and front facing) with the participant standing 3 ft from the camera against a white backdrop. Each photograph was cropped to 336 × 425 pixels and desaturated to create five black-and-white photographic stimuli for the own image condition (own). The matched other condition (other) consisted of a standardized set of five black-and-white images of a gender- and race-matched other person in the same resolution, size, face orientation, and brightness (from the MIT CBCL Face Recognition Database, http://cbcl.mit.edu/software-datasets/heisele/facerecognition-database.html, or taken in the laboratory by the same specifications previously described).
Task administration
The SIP task was administered in Session 2, prior to the MRI scan, using E-Prime and a laptop computer. The SIP-EJ and SIP-SJ task conditions were counterbalanced in order of presentation across participants. Both task conditions included 16 trial blocks of 12 trials each, each preceded by instructions and 4 practice trials. Trial stimuli consisted of a word (self-descriptive positive, self-descriptive negative, non–self-descriptive positive, non–self-descriptive negative) superimposed on an image (own, other) for a total of six stimulus conditions (see the Supplemental Material for task validation using alternative control conditions). Each word was presented twice for the SIP-EJ and twice for the SIP-SJ, once paired with own image and once paired with other image. Trial order was counterbalanced within the SIP-EJ and SIP-SJ and across conditions. For the SIP-EJ, participants were instructed: “For the words you are about to see, please respond to each word to indicate whether it is POSITIVE or NEGATIVE. Please respond as FAST AS POSSIBLE.” For the SIP-SJ, participants were instructed: “For the words you are about to see, please respond to each word based on whether or not it DESCRIBES YOU. Please respond as FAST AS POSSIBLE.” Participants had up to 3,000 ms to respond to each stimulus using keys labeled either positive or negative or yes or no (different keys used for response mappings for the SIP-EJ vs. the SIP-SJ). Between trials, the participant was presented with a fixation cross for up to 2,000 ms.
Brooding rumination
At Session 1, participants completed the Ruminative Responses Scale (RRS; Nolen-Hoeksema et al., 2008). The RRS is a measure of trait rumination that can be decomposed into subscales that provide measures of brooding (RRS-B), reflection (RRS-R), and depression (RRS-D; Treynor, Gonzalez, & Nolen-Hoeksema, 2003). The RRS-B is believed to represent the maladaptive tendency toward negative, repetitive thinking that is distinct from depressive symptoms (captured in the RRS-D) or a more adaptive form of introspection (captured in the RRS-R). The RRS-B includes five items describing typical responses that an individual makes to stressors or negative emotions (e.g., “go away by yourself and think about why you feel this way”), and each item is rated on a scale from 1 (almost never) to 4 (almost always). (See the Supplemental Material for analyses using other subscales.)
Resting-state functional connectivity
At Session 2, participants completed a magnetic resonance imaging (MRI) scan that included anatomical scanning and a resting-state functional scan. A Siemens Tim Trio 3T scanner and 32-channel head coil were used to collect a high-resolution T1-weighted anatomical image (repetition time = 2,200 ms; echo time = 4.27 ms; flip angle = 7°; 144 slices; field of view = 230 mm; matrix = 192 × 192; voxel size = 1.2 × 1.2 × 1.2 mm) and eyes-open resting functional images (repetition time = 3,000 ms; echo time = 30 ms; flip angle = 85°; 47 slices; field of view = 216 mm; matrix = 72 × 72; voxel size = 3 × 3 × 3 mm; total duration = 6.2 min; total volumes = 124). Resting state fMRI data were collected immediately following collection of anatomical data and prior to other functional scanning.
Analyses
Behavioral analyses
The outcome variables of interest from the SIP task were reaction time (RT) or proportion of trials within each task condition that were reported to be positive versus negative (for the SIP-EJ) or self-descriptive versus non–self-descriptive (for the SIP-SJ). Behavioral data were processed using R. Reaction time analyses were conducted by calculating an average for each trial type. Incorrect trials and trials on which RTs were less than 200 ms or more than 3 standard deviations above the within-subject mean were excluded from analyses (consistent with (Henderson, Snyder, Gupta, & Banich, 2012; Kaiser, Andrews-Hanna, Metcalf, et al., 2015; Snyder et al., 2014). RTs were natural log transformed and proportion estimates were arcsine transformed to reduce the skew common to RT or proportion estimate data that violate the statistical assumption of normal distribution. All RT distributions met normality requirements (see Table S2 in the Supplemental Material); however, arcsine-transformed proportion data for several conditions were not normally distributed, suggesting possible ceiling effects and prompting the focus on RT as the outcome variable of interest. (For exploratory analyses using proportion data, see the Supplemental Material.)
Behavioral analyses were conducted using SPSS to perform mixed design analysis of variance (MANOVA) in which within-subject variables included task condition (emotion judgment, self-descriptiveness judgment), image type (own, other), word self-descriptiveness (self-descriptive, non–self-descriptive), and word valence (positive, negative) and the between-subject variables were clinical group (MDD = +1, HC = −1), trait brooding (z-transformed RRS-B score), and their interaction. Together, two MANOVAs were performed to test Hypotheses 1(a, b, c) and 2(a, b): (1) a MANOVA with task conditions as within-subject variables and clinical group and trait brooding as group-level variables and (2) the same MANOVA, adding the interaction between clinical group and trait brooding. (Of note, a simple MANOVA was performed first to clarify task effects in the absence of group-level variables.)
Analyses were also performed replacing the group variable for depression with symptom severity, as evaluated with the Beck Depression Inventory (Beck, Steer, & Garbin, 1988). The analyses yielded results that were nearly identical to those in which depression was included as a categorical variable, likely because clinical status and BDI scores were highly collinear and BDI scores in the present sample had a clear bimodal distribution. Therefore, because BDI scores were clearly bimodal, we elected to operationalize depression as a categorical variable in experimental analyses.
Resting-state functional connectivity analyses
Functional connectivity analyses were performed with parameters and processing steps identical to those described in Kaiser et al. (2016). Static (overall) and dynamic (variability over a sequence of sliding windows) RSFC was calculated between a region of interest (ROI) in MPFC (cluster defined by meta-analysis of default network RSFC abnormalities in depression; Kaiser, Andrews-Hanna, Wager, et al., 2015) and ROIs in left and right anterior insular (AI; 4-mm radius spherical ROIs at ± 34, 8, –8, defined by meta-analysis of insular RSFC and implicated in emotion processing and psychopathology; Chang, Yarkoni, Khaw, & Sanfey, 2013; see Fig. S2 in the Supplemental Material).
General image preprocessing
We discarded the first 6 s of each subject’s functional data to allow for stabilization of the magnetic field. Preprocessing of functional data was performed in SPM12 using the standard spatial preprocessing steps of slice-time correction, realignment, normalization in Montreal Neurological Institute (MNI) space, and smoothing with a 6-mm kernel.
Head motion and artifact detection
We used SPM12 to assess head motion by translation and rotation in x, y, and z directions. Next, we used Artifact Detection Tools (ART; www.nitrc.org/projects/artifact_detect/) to calculate time points of significant head motion or fluctuations in the magnetic field (> 0.5 mm motion from previous frame, global mean intensity > 3 standard deviations from mean intensity across functional scans) for each participant. Then, outlier images were modeled in each participant’s first-level general linear model (as a vector the length of the time series, with 1 for outlier time points and 0 for nonoutlier time points) to censor the influence of outlier time points on estimates of functional connectivity while maintaining the temporal structure of the data. Thus, together, motion correction included the regressing out of both residual head motion parameters (three translation and three rotation parameters, plus one composite motion parameter reflecting the maximum scan-to-scan movement) and outlier volumes (as calculated through artifact detection). Analyses were performed to test for associations between motion and experimental variables; framewise motion/outliers were not significantly associated with clinical depression, t(41) = 0.14, p = .89; depressive symptom severity, r(41) = 0.03, p = .83; or brooding, r(41) = −0.09, p = .59.
Denoising
Denoising of the time series was performed in the CONN toolbox (https://www.nitrc.org/projects/conn/; Whitfield-Gabrieli & Nieto-Castanon, 2012), which uses CompCor (Behzadi, Restom, Liau, & Liu, 2007) to estimate physiological noise from white matter and cerebrospinal fluid for each subject using principal component analysis. The first five components were then regressed out of each subject’s functional data on the first level of analysis. In addition, a band-pass filter of 0.0278 to 0.10 Hz was applied to the time series. This range was selected to remove high-frequency activity related to cardiac and respiratory activity (Cordes et al., 2001) and low-frequency activity with a period that exceeds the duration of sliding windows used in dynamic analyses (Leonardi & Van De Ville, 2015). Accordingly, the corrections performed on the time series included detrending, outlier correction, motion regression, and CompCor correction (which were performed together in a single first-level regression model), followed by band-pass filtering. These corrections produced a residual BOLD time course at each voxel that was used for subsequent analyses.
Dynamic resting-state functional connectivity analysis
Dynamic RSFC analyses were performed using the CONN toolbox (to estimate RSFC among ROIs for each sliding window) and in-house scripts (to calculate standard deviation in RSFC across windows) written in Matlab (Mathworks, Natick, MA).1 For first-level dynamic analyses, the time course was segmented into 36-s windows sliding the onset of each window by 18 s, for a total of 19 windows (see discussion in Kaiser et al., 2016; Leonardi & Van De Ville, 2015). Next, the Fisher’s z-transformed Pearson’s correlation coefficient was computed for each sliding window between the truncated time course of the MPFC and that of all other voxels, yielding a set of sliding-window beta maps for each participant. Dynamic variability in RSFC was estimated by calculating the standard deviation (SD) in beta values at each voxel. For group-level analyses, the SD in beta values was extracted from the left and right AI ROIs using REX (Duff, Cunnington, & Egan, 2007), and a MANOVA was performed in which the between-subject variables were clinical group (MDD = +1, HC = −1), trait brooding (z-transformed RRS-B), and their interaction; hemisphere (left, right) was included as the within-subject variable.
Static resting-state functional connectivity analysis
Static RSFC analyses were performed using the CONN toolbox (to estimate RSFC among ROIs across the full duration of the scan). For first-level static analyses, the Fisher’s z-transformed Pearson’s correlation coefficient was computed between the full-time course of the MPFC seed and the full-time courses of all other voxels. For group-level analyses, the beta value representing overall correlation in activity was extracted from the left and right AI ROIs using REX, and a MANOVA was performed in which the between-subject variables were clinical group (MDD = +1, HC = −1), trait brooding (z-transformed RRS-B), and their interaction; hemisphere (left, right) was the within-subject variable.
All group-level fMRI analyses were repeated including age and number of outlier images as covariates; because these variables did not significantly differ between clinical groups or correlate with RRS-B scores and controlling for these variables did not affect results, simple analyses are reported.
Mediation
To test the hypothesis that RSFC in a frontoinsular circuit linking insula with MPFC mediates an association between rumination and attention bias among depressed individuals, we used a bootstrapping approach to estimate the indirect effect of brooding rumination (z-transformed RRS-B) on task performance (RT for self-descriptive compared with non–self-descriptive words, as moderated by valence and image type) through dynamic RSFC (SD in correlated activity between MPFC and left AI and between MPFC and right AI). This analysis was designed to test Hypothesis 3. Regression analyses were performed to estimate the regression coefficients.
In sum, a total of three analyses were performed for hypothesis testing (two MANOVAs and one mediation model). Supporting analyses were performed to demonstrate basic task effects and replication of prior findings or for post hoc clarification of significant experimental effects.
Results
General task effects
To investigate overall task effects, we performed a 2 (task condition: emotion judgment, self-descriptiveness judgment) × 2 (image type: own, other) × 2 (word self-descriptiveness: self-descriptive, non–self-descriptive) × 2 (word valence: positive, negative) MANOVA. This analysis revealed significant main effects for all factors. Across the group, participants were faster to judge emotional valence of words than self-descriptiveness of words, F(1, 48) = 43.94, p < .01, η p 2 = .48; faster to judge self-descriptive than non–self-descriptive words, F(1, 48) = 12.08, p < .01, η p 2 = .20; faster to judge positive than negative words, F(1, 48) = 5.57, p = .02, η p 2 = .10; and faster to judge words presented with their own image in the background than another person’s image in the background, F(1, 48) = 18.19, p < .01, η p 2 = .28. Interactions among stimulus conditions also emerged: The facilitating effects of self-referential content (either word self-descriptiveness or background image of the participant’s own face) were stronger for positive than negative words, F(1, 48) = 26.12, p < .01, η p 2 = .35, and F(1, 48) = 4.60, p = .03, η p 2 = .09, and the strongest facilitation effects emerged for self-descriptive positive words paired with the participant’s own image, F(1, 48) = 5.50, p = .02, η p 2 = .10 (Fig. 2; see Figs. S3 and S4 in the Supplemental Material).
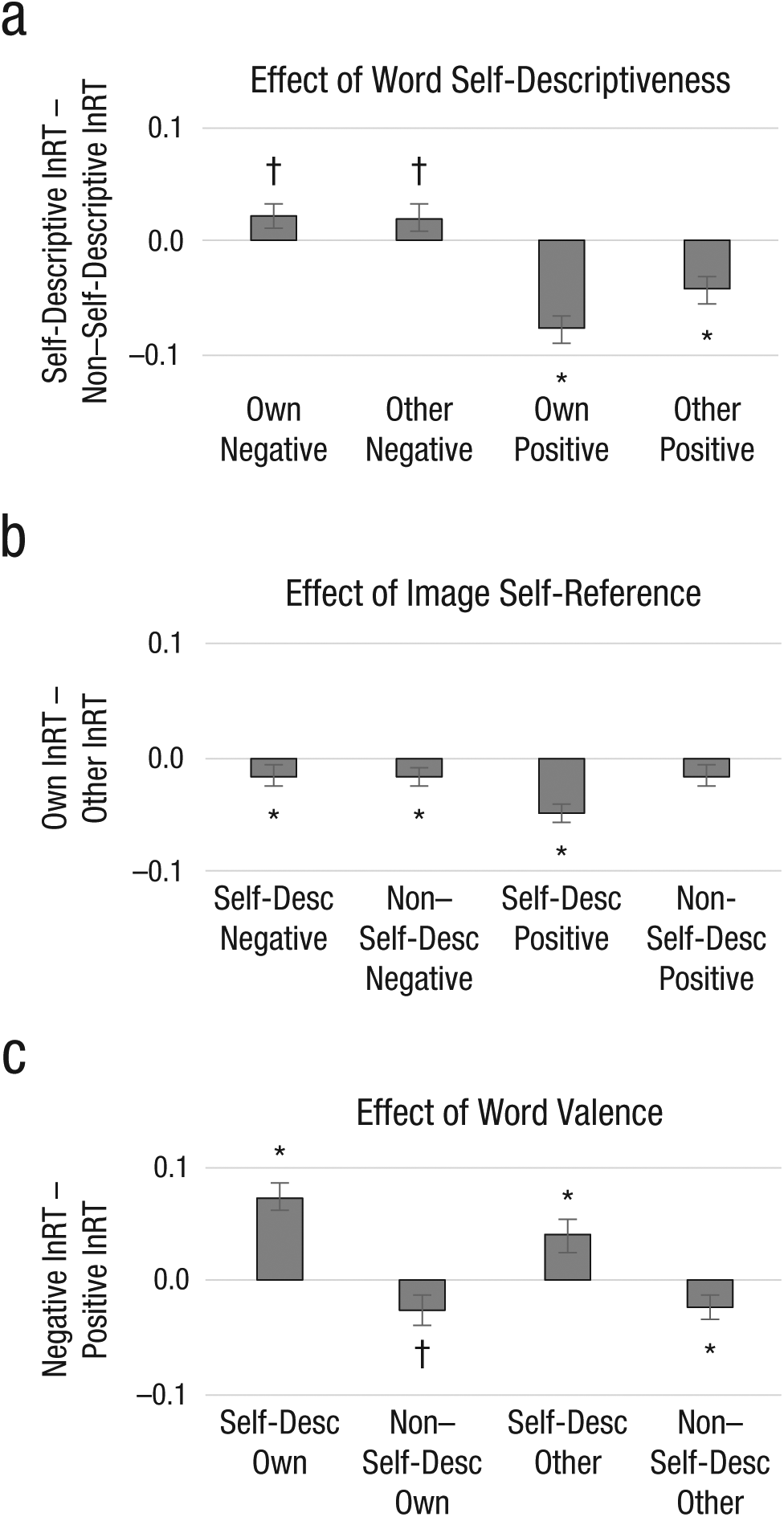
Main and moderated effects of the self-referential information processing task on reaction time (RT). On average, participants were significantly faster to judge both emotional valence and self-descriptiveness of self-descriptive (compared with non–self-descriptive) positive (compared with negative) words paired with their own (compared with someone else’s) image. Main and moderated effects of word self-descriptiveness: (a) Natural-log transformed RTs for self-descriptive – non–self-descriptive word trials are displayed. Main and moderated effects of image type: (b) Natural-log transformed RTs for own-image – other-image trials are displayed. Main and moderated effects of word valence: (c) Natural-log transformed RTs for negative – positive word trials are displayed. Task effects, †p < .10. *p < .05.
Moderating effects of clinical depression and trait rumination
We performed experimental analyses to test hypotheses by adding the group-level variables of clinical depression (HC = −1, MDD = +1) and brooding rumination (z-transformed RRS-B) to the MANOVA described previously. In this MANOVA, first, clinical group moderated the effects of task condition and word type (Group × Condition × Valence × Self-Descriptiveness Interaction), F(1, 46) = 17.40, p < .01, η p 2 = .27. Follow-up MANOVA revealed that depression effects were specific to the self-descriptiveness judgment task: When participants responded to indicate whether a word described them, healthy participants were faster to judge positive self-descriptive words and slower to judge negative self-descriptive words, but depressed participants showed faster judgment of negative self-descriptive words and also showed blunted facilitation for positive self-descriptive words, F(1, 47) = 15.23, p < .01, η p 2 = .25 (Fig. 3A; see Figs. S5 and S6 in the Supplemental Material). These depression effects are consistent with Hypotheses 1c (attention biases evoked by the interaction of self-descriptiveness and valence) and 2a (attention biases evoked at later stages of processing with information that is elaborated).
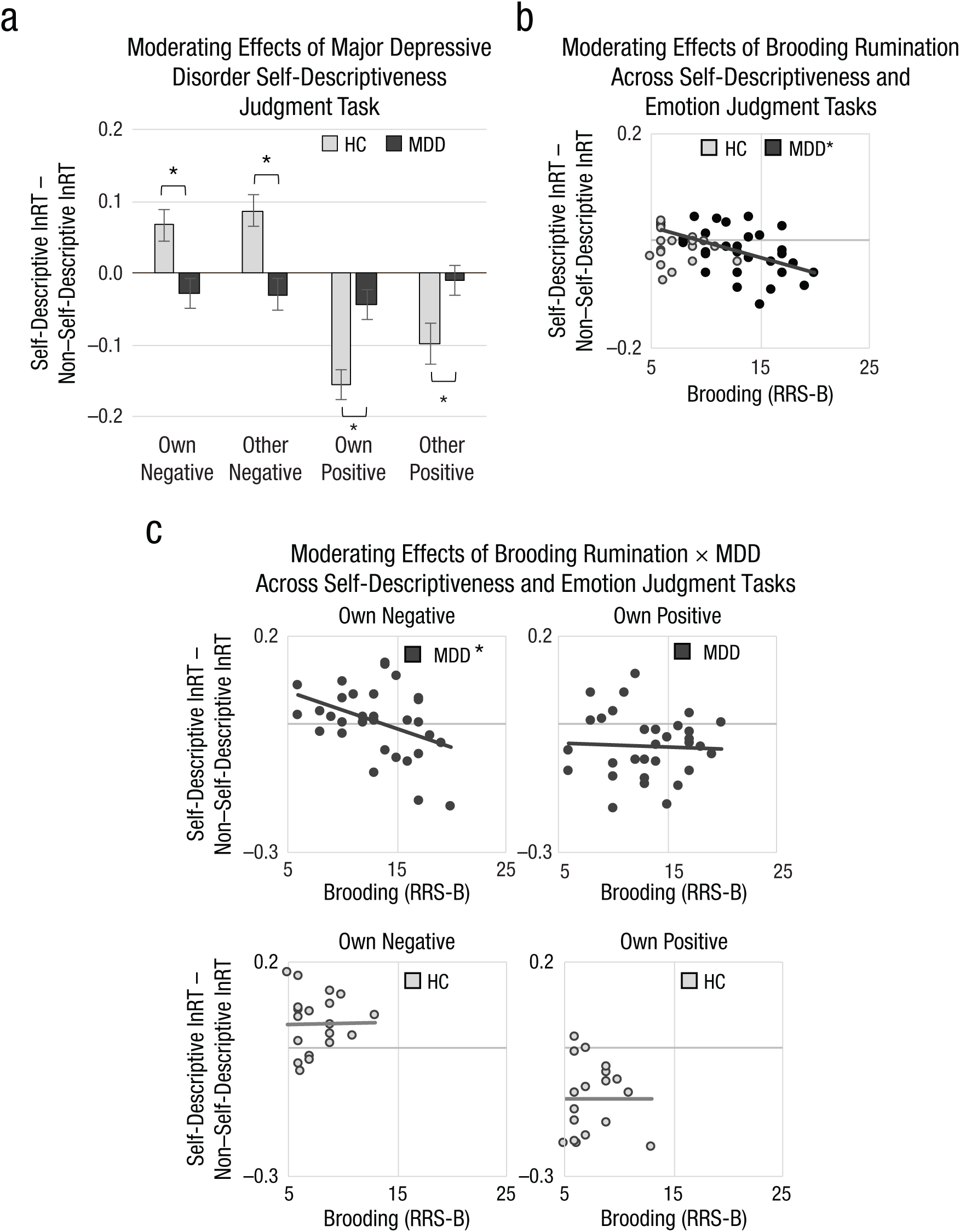
Moderating effects of depression, trait brooding, and their interaction on reaction time (RT) for the self-referential information processing task. When judging self-descriptiveness of words, (a) individuals with major depressive disorder (MDD) were faster than healthy control (HC) individuals to judge self-descriptive negative words (as opposed to non–self-descriptive negative words), but relatively slower than healthy people to judge self-descriptive positive words (as opposed to non–self-descriptive positive words). Across the group and controlling for depression status, (b) higher levels of trait brooding rumination (measured with the Ruminative Responses Scale, Brooding subscale, RRS-B) were associated with faster responses to self-descriptive (relative to non–self-descriptive) words regardless of task goals. Among individuals with depression, (b) those with higher levels of trait brooding exhibited stronger self-referential facilitation effects for negative words (i.e., faster responses to self-descriptive negative words accompanied by their own image). Group differences in task effects or significant correlations, *p < .05.
Second, trait brooding moderated the effects of word self-descriptiveness: Regardless of task goals, emotional word content, or accompanying images, participants more prone to brooding were faster to judge self-descriptive (compared with non–self-descriptive) words, F(1, 46) = 7.94, p < .01, η p 2 = .15 (Fig. 3b). Next, adding the interaction of depression and rumination to the model revealed attention bias effects specific to ruminative depression, F(1, 45) = 3.80, p = .05, η p 2 = .08. Follow-up MANOVAs conducted within the MDD and HC groups showed that there were no significant effects of trait brooding among healthy participants, but among depressed individuals, higher levels of brooding were associated with faster responses to self-descriptive words accompanied by the participant’s own image when words were negative but not positive, F(1, 28) = 4.65, p = .04, η p 2 = .14 (Fig, 3c; see Fig. S7 in the Supplemental Material). Together, these findings for ruminative depression support Hypotheses 1c (attention biases evoked by the interaction of self-descriptiveness and valence); however, they also support Hypothesis 2b (attention biases evoked at earlier stages of processing with information that is task-irrelevant and should be ignored).
(Note that replacing brooding [RRS-B] with depression-specific rumination [RRS-D] in the previously analyses failed to yield significant results; replacing brooding with reflective rumination [RRS-R] revealed overlapping attention biases to self-descriptive words, in general, but only brooding showed an interaction with clinical depression in predicting attention bias to negative self-referential material; see the Supplemental Material.)
Frontoinsular dynamic and static functional connectivity
In preparation for mediation analysis, we performed analyses to test for replication of prior findings of altered RSFC between AI and MPFC in depressed ruminators (Kaiser et al., 2016). We performed two separate MANOVAs to examine dynamic variability (standard deviation over time) in RSFC between MPFC and AI or static (overall) RSFC between MPFC and AI. In each MANOVA, the within-subject variable was hemisphere (left, right), and the group-level variables were clinical group (MDD = +1, HC = −1), trait brooding (z-transformed RRS-B), and their interaction. Analyses showed a significant interaction between depression and brooding in predicting dynamic RSFC, F(1, 39) = 5.85, p = .02, η p 2 = .13, and a trend for a main effect of brooding, F(1, 39) = 3.41, p = .07, η p 2 = .08: Higher levels of brooding were associated with increased variability in frontoinsular RSFC, and this association was significantly stronger for depressed participants than healthy participants (Fig. 4a; see Fig. S8 in the Supplemental Material). For analyses examining static frontoinsular RSFC, there were no significant main effects of clinical depression, F(1, 39) = 2.05, p = .16, η p 2 = .05, brooding, F(1, 39) = 2.06, p = .16, η p 2 = .05, or interaction, F(1, 39) = 1.53, p = .22, η p 2 = .04 (see Fig. S8 in the Supplemental Material).
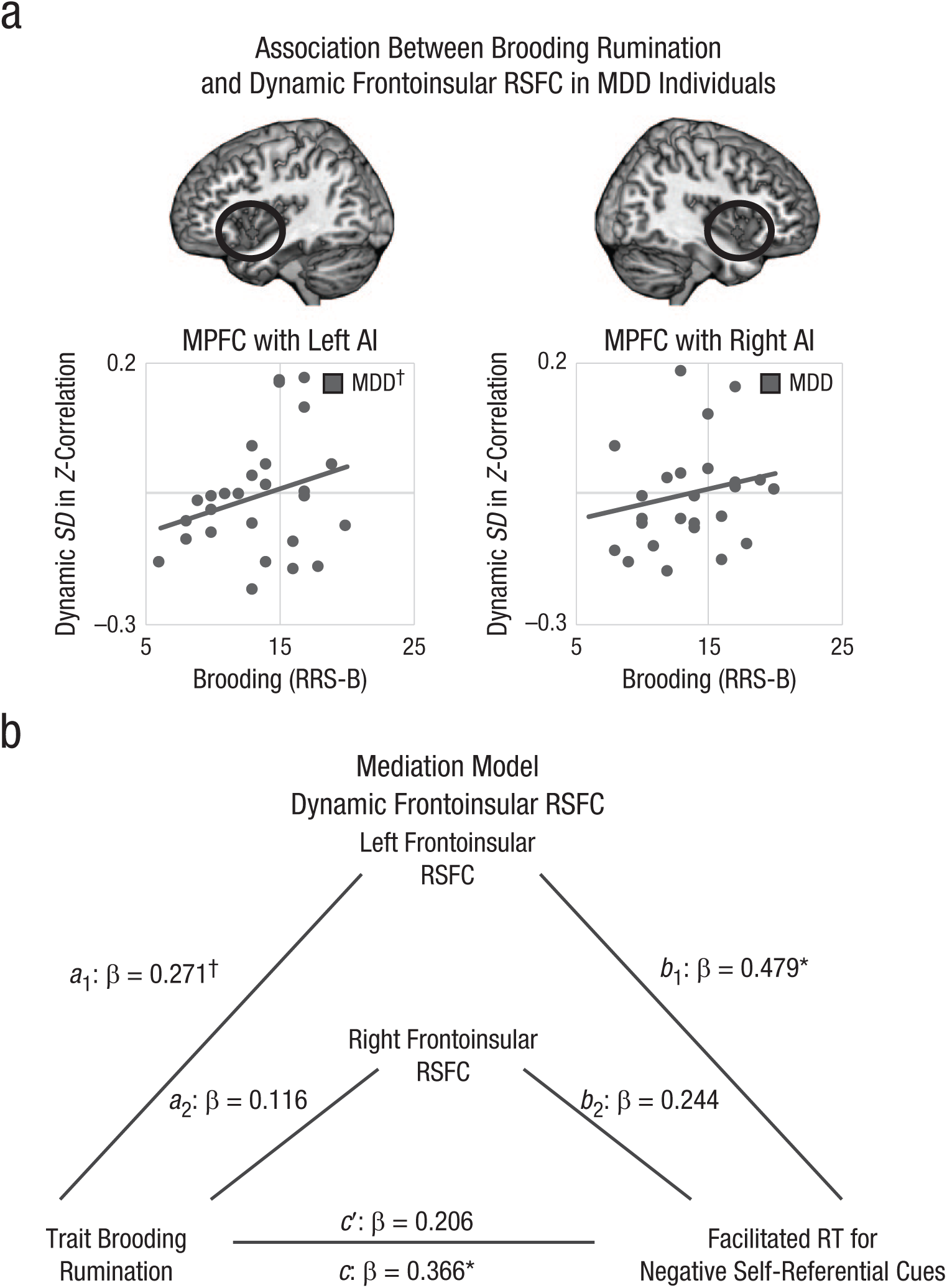
Frontoinsular resting-state functional connectivity (RSFC) and attention biases in the ruminative phenotype of depression. Consistent with prior findings in an independent sample (Kaiser et al., 2016), (a) we found that elevated trait brooding rumination (Ruminative Responses Scale, Brooding subscale, RRS-B) was related at a trend level to increased variability in RSFC between regions of interest (ROIs) in medial prefrontal cortex (MPFC) and left anterior insula (AI). Dynamic variability in RSFC (b) between MPFC and left AI significantly mediated the association between trait brooding and attention biases toward negative self-descriptive and self-referential information (indirect effect across bootstrapping = 0.01, bias-corrected 95% confidence interval, or CI = [0.0002, 0.0441]). Dynamic variability in RSFC was operationalized as standard deviation (SD) in Fisher’s z-transformed correlation coefficients over a series of sliding windows; conservative motion correction procedures were applied to the data (see Method section). Displayed are regression coefficients for each path of the mediation model, including the total effect of trait brooding on attention biases in the model without mediators (c), the direct effect of trait brooding on attention biases in a model with mediators (c′), the effects of trait brooding on dynamic variability in RSFC in left (a1) or right (a2) AI, and the effects of dynamic variability in RSFC in left (b1) or right (b2) AI when controlling for trait brooding. Correlations or paths in mediation model, †p < .10. *p < .05.
Mediation
To test the prediction (Hypothesis 3) that RSFC between AI and MPFC mediates an association between ruminative depression and attention biases, we performed a bootstrapping mediation analysis within the MDD group using the INDIRECT SPSS command (Fig 4b). Variables entered in the model included brooding (z-transformed RRS-B: X), attention bias to negative self-referential information (RT contrast corresponding with the task effect of word self-descriptiveness as moderated by valence and image type: Y), and frontoinsular RSFC (dynamic and static RSFC between MPFC and left and right AI: Ms). Consistent with the results of MANOVA analyses previously described, there was a significant total effect of brooding in predicting facilitated RT to negative self-descriptive words accompanied by the participant’s own image (c path unstandardized b = 0.032, 95% confidence interval, or CI = [0.001, 0.063], standardized β = 0.366) and a trend-level association between increased brooding and higher dynamic RSFC between MPFC and left anterior insula (left AI: a path unstandardized b = 0.018, 95% CI = [–0.006, 0.042], standardized β = 0.271; right AI: a path unstandardized b = 0.008, 95% CI = [–0.019, 0.034], standardized β = 0.116). Higher left frontoinsular dynamic RSFC was significantly associated with more extreme attention bias to negative self-referential information (left AI: b path unstandardized b = 0.671, 95% CI = [0.097, 1.245], standardized β = 0.479; right AI: b path unstandardized b = 0.335, 95% CI = [–0.207, 0.876], standardized β = 0.244). The indirect effect of brooding through left frontoinsular dynamic RSFC on attention bias was significant (average indirect effect across bootstrapping = 0.01, bias-corrected 95% CI = [0.0002, 0.0441]), and the direct effect of brooding on attention bias was no longer significant when including frontoinsular dynamic RSFC as the mediator (c′ path unstandardized b = 0.019, 95% CI = [–0.016, 0.054], standardized β = 0.206).
Discussion
This study’s purpose was to provide insight into the neurocognitive mechanisms of attention bias in depression by asking: What types of information evoke biased attention, what aspects of attention are biased, and how are attention biases reflected in the intrinsic functioning of frontoinsular-default networks? Results showed that first, clinical depression was associated with attention biases specifically toward negative information that was self-referential and elaborated, as evidenced by faster judgments by participants with depression when the self-referential quality of the stimulus was relevant to task goals (self-descriptive words judged on self-descriptiveness). Self-related content that should not be elaborated (in the form of background images or self-descriptiveness of words when judging emotional valence), or non–self-referential content, did not evoke biased performance among depressed individuals compared with nondepressed individuals. These findings support the hypothesis that it is the interaction between emotional content and self-relatedness that evokes attention bias in depression (Wisco, 2009) and are consistent with prior research suggesting that attention biases in depression are instantiated at later (elaborative) stages of attention processing, leaving intact earlier (selecting, orienting) stages of processing (De Raedt & Koster, 2010; Kircanski & Gotlib, 2015).
Second, results showed that ruminative depression was associated with attention biases that were distinct from those biases generally associated with depression: When we controlled for main effects of depression, we found that depressed ruminators were faster to judge negative self-descriptive information regardless of the task relevance of such information, in particular when such information was paired with self-referent images. These patterns are consistent with the idea that ruminative depression is related both to (earlier-stage) biases in orienting to and (later-stage) biases elaborating on negative self-referential information (Joormann, 2010; Whitmer & Banich, 2007).
Third, the present study replicates the association between ruminative depression and dynamic variability in functional connectivity of a frontoinsular circuit linking AI with MPFC (Kaiser et al., 2016) and further shows that such altered network dynamics mediate the association between ruminative depression and biased attention to self-referential negative information. These patterns support the idea that coordinated functioning of insula and midline cortical systems could underlie the tendency for attention to be captured by negative self-focused thinking. Amplified and variable resting-state functional connectivity between these regions could represent an intrinsic tendency for salient self-referential material (albeit irrelevant to present-moment goals) to activate insula to direct resources to midline brain systems and enhance processing of self-referential information (Menon & Uddin, 2010; Sridharan et al., 2008). In the present task, we observed enhanced (faster) performance; however, in more complex tasks or daily life, enhanced processing of self-referential material may also come at the cost of goal-directed action and emotional health (Koster, De Lissnyder, & De Raedt, 2013; Koster, De Lissnyder, Derakshan, & De Raedt, 2011).
Of note, there are other interpretations of the present study findings. For example, we interpreted depression-related differences in task performance as evidence for attention biases because depressed women were faster to judge self-descriptive (than non–self-descriptive) negative words, whereas nondepressed women were slowed in judging this material. However, as a group, depressed women were also faster to judge self-descriptive (than non–self-descriptive) positive words, albeit to a weaker extent than nondepressed women. Therefore, it could be argued that depression-related “biases” are defined by the absence of valence-specific biases that are robustly exhibited by healthy individuals. However, we note that among depressed women, brooding rumination was associated with the presence of increased biases toward negative self-referential material (rather than an absence of performance biases toward positive self-referential material). Thus, there may be different ways that attention biases manifest in depression and depressive phenotypes, and interpretations for attention biases are performed in consideration of relative patterns of performance between groups or across dimensions.
Several limitations to the present study warrant discussion, as do a number of questions motivated by these findings that should be addressed in future research. First, these results highlight the interactive roles of personal salience and emotional content in driving attention bias. However, stimuli were restricted to particular forms of emotional and self-referent information (adjectives, images); extending experimental stimuli to other forms of self-referent cues (e.g., words evoking autobiographical memories) could provide complementary information about attention bias. Second, attention biases with task-relevant versus task-irrelevant information are interpreted here as evidence for biases evoked at varying stages of processing; however, there may be other explanations. For example, depression versus rumination could differently influence processing of verbal versus image-based self-referential content; task variants that include other types of (image or verbal) stimuli might distinguish these possibilities. In addition, although we adapted the present behavioral task from classic attention interference tasks, other cognitive processes may influence performance. For example, this task (and indeed, many tasks requiring attention to and judgment of stimulus content) could involve incidental encoding of information, incidental retrieval of autobiographical memories, or other processing (e.g., schema activation) that could enhance or impede performance. Future research using alternative task designs that yields converging results could provide more precise evidence for biases in attention or specific attention subprocesses.
A third limitation of this study is related to sample characteristics. Here, because of evidence that ruminative depression is more common in women, we recruited a female sample. It is unknown, from this study, whether effects would generalize to other genders or if gender-specific attention biases exist. In addition, we note that a broader range of trait rumination was reported among depressed women, yielding relatively enhanced statistical power for detecting effects of rumination within this group. Studies specifically designed to assess trait rumination in nondepressed samples could better evaluate attention biases related to rumination in healthy individuals.
Fourth, the focus of the present neurobiological measures was on intrinsic frontoinsular functioning in a circuit linking AI with MPFC, but neural activity in other circuits or in response to explicit task demands could provide complementary information (Ho et al., 2014). For example, future research may compare frontoinsular activity at rest with activity in response to cognitive regulation of self-referential thinking, when attention biases may be most active. We also note that despite the encouraging replication of rumination-related abnormalities in intrinsic network dynamics, much remains unknown about the functional significance of such dynamics. Some recent theoretical and empirical work support the idea that network dynamics at rest are a reliable property of brain functioning (Hutchison et al., 2013) that may reflect cross-talk among neural circuits (Hutchison & Morton, 2015; Kaiser et al., 2016). However, other researchers have pointed out that estimates of dynamic variability are vulnerable to contamination by motion or sampling variability (Laumann et al., 2017). Although we took a conservative approach to motion correction in these analyses and sampling variability is unlikely to explain group-level effects, it is worthwhile to note that the field of network dynamics is new and rapidly evolving. Future research that investigates the convergence of multiple measures of dynamic network functioning (e.g., intrinsic network states, Calhoun, Miller, Pearlson, & Adali, 2014; co-activation pattern analysis, Chen, Chang, Greicius, & Glover, 2015) is needed.
In conclusion, this study provides evidence that attention biases in clinical depression are specific to negative self-referential material and reveals distinctions in the nature of attention biases related (generally) to depression or (particularly) the ruminative dimension of depression. Relative to nondepressed women, individuals with depression showed attention biases toward negative information that was self-referential and task-relevant, suggesting biases evoked at later stages of processing with elaborated information. Depressed ruminators were characterized by attention biases to negative self-referential information regardless of task goals and especially when paired with self-referent images, suggesting biases at both later (elaborative) and earlier (orienting) stages of processing. Rumination-related attention biases were mediated by dynamic circuit activity among regions involved in orienting attention toward self-focused thinking. These findings highlight the importance of a research approach that disentangles distinct clinical dimensions to understand psychopathology and suggest future research that asks questions such as how rumination may interact with other symptom presentations (e.g., ruminative phenotype of substance use disorder or anxiety; Nolen-Hoeksema & Watkins, 2011) or how other clinical dimensions (e.g., anhedonia) may interact with depressive symptoms to produce distinct profiles of cognitive bias. In addition, this research supports integration across multiple levels of neurocognitive functioning to understand phenotypes (or “biotypes”) of illness, which may be critical for revising and improving our theories of the active mechanisms of mood pathology. Such an integrative approach could support discovery of treatment targets: for example, interventions that target frontoinsular-default circuits (with neurofeedback or other techniques) and attention biases (using cognitive therapy techniques or attention bias modification). For example, existing attention bias modification programs teach individuals to direct attention away from negative and toward positive material using standardized computer-based paradigms and stimuli. Although the standardization of such interventions is in many ways a strength, the present results suggest that tailoring bias modification to the attention biases of a particular client—for example by incorporating personally salient negative and positive stimuli or targeting biases at multiple levels of attention processing—could enhance the impact of these treatments. Together, these dimensionally focused, multimodal approaches may support a more precise characterization of depression and related mood pathology for individual patients and enhance clinical outcomes.
Supplemental Material
Kaiser_Open_Practices_Disclosure – Supplemental material for Attention Bias in Rumination and Depression: Cognitive Mechanisms and Brain Networks
Supplemental material, Kaiser_Open_Practices_Disclosure for Attention Bias in Rumination and Depression: Cognitive Mechanisms and Brain Networks by Roselinde H. Kaiser, Hannah R. Snyder, Franziska Goer, Rachel Clegg, Manon Ironside and Diego A. Pizzagalli in Clinical Psychological Science
Supplemental Material
Kaiser_Supplemental_Material – Supplemental material for Attention Bias in Rumination and Depression: Cognitive Mechanisms and Brain Networks
Supplemental material, Kaiser_Supplemental_Material for Attention Bias in Rumination and Depression: Cognitive Mechanisms and Brain Networks by Roselinde H. Kaiser, Hannah R. Snyder, Franziska Goer, Rachel Clegg, Manon Ironside and Diego A. Pizzagalli in Clinical Psychological Science
Footnotes
Acknowledgements
A subset of analyses reported here was presented (by R. H. Kaiser) at the 2014 annual meeting of the Cognitive Neuroscience Society and the 2015 McLean Hospital Research Day.
Action Editor
Erin B. Tone served as action editor for this article.
Author Contributions
R. H. Kaiser developed the study concept. R. H. Kaiser, H. R. Snyder, and D. A. Pizzagalli contributed to the study design. Testing and data collection were performed by R. H. Kaiser, F. Goer, R. Clegg, and M. Ironside. R. H. Kaiser performed the data analysis, and R. H. Kaiser, H. R. Snyder, and D. A. Pizzagalli performed interpretation of analyses. D. A. Pizzagalli and R. H. Kaiser provided funding. R. H. Kaiser drafted the manuscript, and all other authors provided critical revisions. All of the authors approved the final manuscript for submission.
Declaration of Conflicting Interests
The author(s) declared that there were no conflicts of interest with respect to the authorship or the publication of this article.
Funding
This work was supported by the Beverley Sears Award (to R. H. Kaiser), Ted Volsky Memorial Award (to R. H. Kaiser), and National Institutes of Mental Health Grant 5R01-MH095809 (to D. A. Pizzagalli).
Notes
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.