
Abstract
The reward-hypersensitivity model posits that trait reward hypersensitivity should elicit hyper/hypo-approach motivation following exposure to recent life events that activate (goal striving and goal attainment) or deactivate (goal failure) the reward system, respectively. To test these hypotheses, we had 87 young adults with high trait reward (HRew) sensitivity or moderate trait reward (MRew) sensitivity report frequency of life events via the Life Event Interview. Brain activation was assessed during the functional MRI monetary-incentive-delay task. Greater exposure to goal-striving events was associated with higher nucleus accumbens (NAc) reward anticipation among HRew participants and lower orbitofrontal cortex (OFC) reward anticipation among MRew participants. Greater exposure to goal-failure events was associated with higher NAc and OFC reward anticipation only among HRew participants. This study demonstrated different neural reward anticipation (but not outcome) following reward-relevant events for HRew individuals compared with MRew individuals. Trait reward sensitivity and reward-relevant life events may jointly modulate reward-related brain function, which has implications for understanding psychopathology.
The brain’s reward system subserves approach motivation and pursuit of rewards and goals (e.g., Haber & Knutson, 2010). The reward-hypersensitivity theory (Alloy & Abramson, 2010; Alloy et al., 2016; Alloy & Nusslock, 2019; Depue & Iacono, 1989; Johnson et al., 2012; Nusslock & Alloy, 2017), often applied to bipolar spectrum disorders (BSDs), hypothesizes that individual differences in trait sensitivity to rewards will combine with individuals’ exposure to life events that tend to activate or deactivate the reward system to influence state levels of reward responsivity. However, to our knowledge, as yet, no work has precisely tested these hypotheses. To test the predictions of the model, in this functional MRI (fMRI) study, we examined activation in reward-related neural regions, the nucleus accumbens (NAc) and orbitofrontal cortex (OFC), as a function of recent reward-related life-event exposure among individuals with self-reported high trait reward (HRew) sensitivity compared with moderate trait reward (MRew) sensitivity. The findings from this work may contribute greater understanding of variation in neural reward responses as a function of exposure to naturally occurring reward-relevant life events. In addition, it may have implications for advancing the understanding of psychiatric disorders characterized by aberrant approach motivation and how life events can perturb reward processing implicated in these disorders.
The Reward System
Reward processing is linked to a cortico-striatal neural circuit that promotes approach motivation, goal-directed behavior, emotion, and decision-making (e.g., Haber & Knutson, 2010). Within this neural circuit, the ventral striatum is involved in coding the incentive properties of stimuli and reward prediction errors. As a core region of the ventral striatum, the NAc is particularly sensitive to reward cue and thus to anticipation of potentially rewarding outcomes (Haber & Knutson, 2010). The OFC has been implicated in reward-related decision-making and computation of reward value, which may be involved in approach toward rewards or goals. The reward system can be activated or deactivated by external events (e.g., opportunity to win a prize; Haber & Knutson, 2010). Accordingly, elevated NAc activation has been construed as a proxy for hyperresponsivity, whereas blunted NAc activation has been established as a proxy for hyporesponsivity. OFC activation is a more complicated proxy for reward-system functioning. The OFC projects input to subcortical regions, including the NAc (Brady & O’Donnell, 2004; Jackson et al., 2001), according to the perceived value and probability of reward receipt (Haber & Behrens, 2014; Phillips et al., 2008). However, subcortical regions also can guide the cortex, which highlights that the OFC may exert regulatory control of the NAc, but subcortical regions like the NAc also can modulate the OFC (Haber & Knutson, 2010; Tottenham & Gabard-Durnam, 2017). Given the bidirectional nature of connections between the OFC and NAc, OFC activation to reward cues may serve as evidence of either responsiveness to reward or regulation of reward and thus may reflect heightened or attenuated reward responsivity, depending on the nature of its functional coupling with subcortical reward regions (Haber & Knutson, 2010).
Reward Hypersensitivity and Psychopathology
A hypersensitivity to rewarding stimuli has been associated with multiple forms of psychopathology, including risk for substance use disorders (Alloy et al., 2009; Nusslock & Alloy, 2017), certain externalizing behaviors (Murray et al., 2018), and BSD. For example, according to the reward-hypersensitivity model of BSDs (Fig. 1; Alloy et al., 2016; Alloy & Nusslock, 2019; Depue & Iacono, 1989; Johnson et al., 2012; Nusslock & Alloy, 2017), people with a hypersensitive reward system excessively respond to goal- or reward-relevant cues. This reward hypersensitivity is hypothesized to lead to extreme approach-related affect (e.g., excitement, elation, or anger) and incentive motivation following exposure to specific life events. Such events include those that activate the reward system (i.e., goal striving, goal attainment, and failures or losses that can be rectified, which leads to anger, in this case) and, in turn, hypomanic or manic symptoms. Reward hypersensitivity also can lead to excessive decreases in approach-related affect and motivation in response to events that deactivate the reward system (i.e., irreconcilable failures or losses that cannot be remediated) and, in turn, bipolar depressive symptoms. In other words, a trait propensity toward extreme activation and deactivation of the reward system resulting in abnormalities in approach motivation is the theorized vulnerability to BSDs in this model (Alloy & Nusslock, 2019). Considerable self-report, behavioral, neurophysiological, and neural evidence supports the reward-hypersensitivity model of BSDs (for reviews, see Alloy et al., 2016; Alloy & Nusslock, 2019; Johnson et al., 2012; Nusslock & Alloy, 2017), but some neural studies obtain contrary results, including either no difference in reward-related brain activation or hypo-activation for individuals with BSD (Johnson et al., 2019; Schreiter et al., 2016; Trost et al., 2014; Yip et al., 2015). However, none of these studies examined neural activation following exposure to reward-relevant events, a key element in the reward-hypersensitivity model of BSDs.
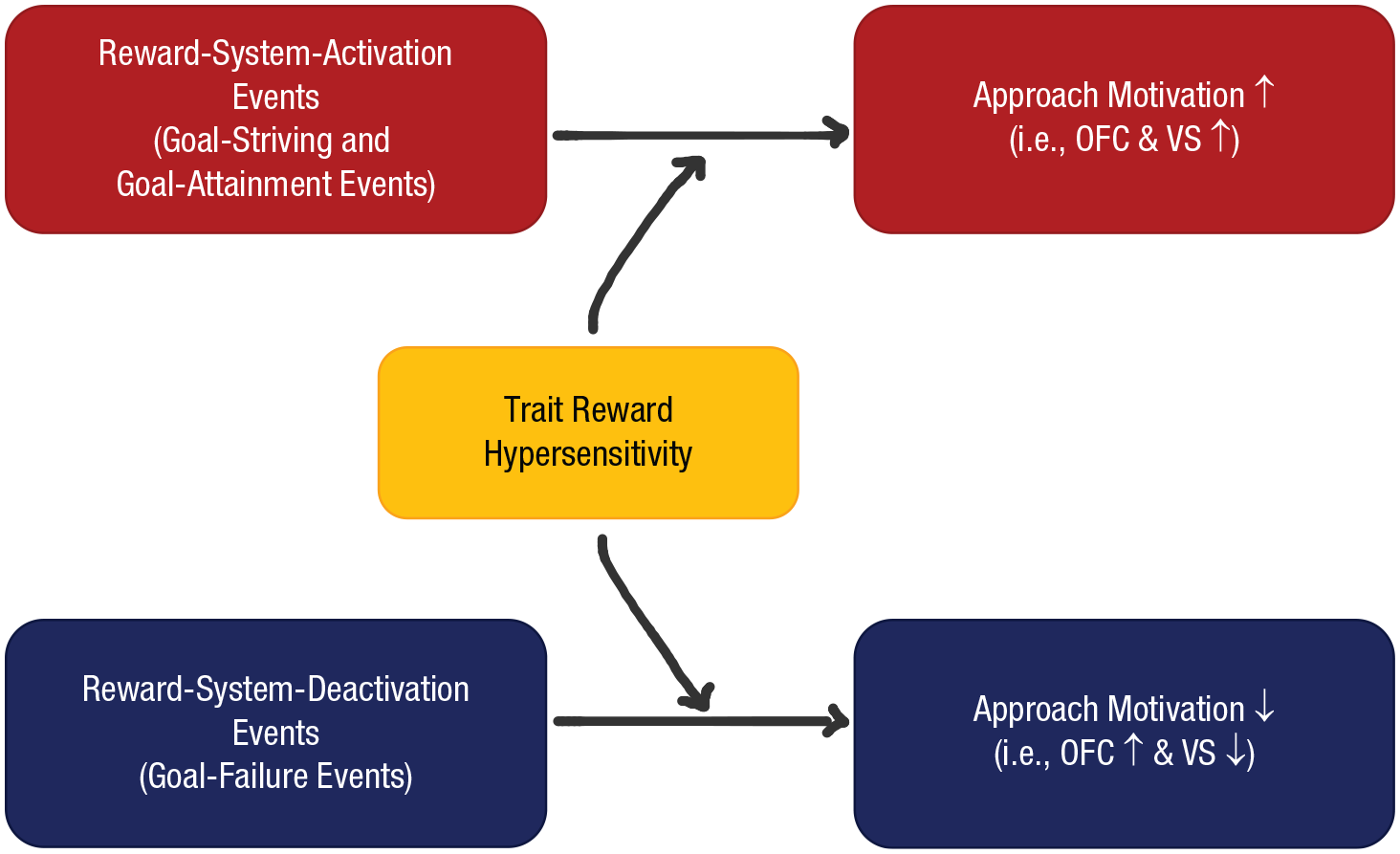
The reward-hypersensitivity model.
The Role of Reward-Relevant Life Events
A central component that separates the reward-hypersensitivity model from traditional diathesis-stress theories (Monroe & Simons, 1991) is the role of specific life events. Diathesis-stress theories propose that psychopathology emerges from the interaction of a vulnerability (diathesis) and broadly defined environmental stress (Monroe & Simons, 1991). Although the reward-hypersensitivity model does involve the concepts of diathesis (trait reward hypersensitivity) and environmental stress (reward-relevant life events), the model offers specificity with an emphasis on the differences among reward-relevant events in terms of triggering reward-system activation and deactivation rather than the “stress” posed by any kind of life events. In other words, not all life events interact with trait reward hypersensitivity in the same manner. Accordingly, individuals with a hypersensitive reward system should manifest more extreme (i.e., hyper or hypo) responses to reward cues following exposure to recent life events that activate or deactivate the reward system compared with individuals with more moderate reward sensitivity. Events involving goal striving (opportunity to work toward a goal or reward; e.g., a possible promotion) and goal attainment (actual receipt of a reward; e.g., a raise), hypothesized to activate the reward system, have been associated with increases in hypomanic or manic symptoms. Events involving irreconcilable goal failures or losses (failures or losses that cannot be remediated; e.g., being fired) have been associated with bipolar depressive symptoms in HRew individuals (Boland et al., 2016). However, there is a paucity of empirical research specifically testing the hypothesized role of such reward-relevant life events in predicting reward-system responses per se.
To our knowledge, only three studies, two with healthy samples and one with individuals with BSD, have examined the effects of an acute lab stressor on neural reward processing. Kumar et al. (2014) found that an acute lab stressor (negative performance feedback) led to increased striatal activation during reward anticipation but decreased striatal activation during reward outcome relative to a no-stress condition. Conversely, Ossewaarde et al. (2011) observed that an acute lab stressor (aversive movie clips) led to decreased medial prefrontal cortex (mPFC) activation to reward anticipation but had no effect on striatal activation. Berghorst et al. (2016) examined the threat of monetary deductions and poor performance feedback on neural reward processing in bipolar individuals. Compared with control participants, bipolar participants in a euthymic or mildly depressed state exhibited elevated amygdala activation during reward anticipation and elevated striatal putamen activation during reward outcome in the stress condition compared with the no-stress condition. Note that these acute lab stressors likely were not perceived as irreconcilable failures.
Likewise, studies have examined the effect of recent perceived life stress on neural reward processing. Among healthy participants, Treadway et al. (2013) found that greater perceived stress within the past month was associated with lower mPFC activation to reward outcome but not reward anticipation. In a sample of patients with major depression and control participants, Kumar et al. (2015) reported that participants with greater perceived life stress exhibited higher mPFC activation to reward outcome during an acute lab stress condition (negative performance feedback) than a no-stress condition, and this effect was primarily attributable to the depressed participants.
Although these findings suggest that acute negative lab stressors and recent perceived life stress influence neural responses to reward anticipation or outcome, none of these studies examined the effects of the actual frequency of recent life-event exposure or of reward-system-activating and reward-system-deactivating events specifically on neural reward responsiveness. Moreover, no prior studies have examined neural reward responsiveness following reward-relevant event exposure in HRew individuals whose reward processing should be most susceptible to such life-event exposure. In other words, it is plausible that neural reward responsiveness may be activated or deactivated following exposure to reward-relevant life events, particularly strongly among HRew individuals.
The Current Study
In the present study, we examined whether greater recent exposure to different types of reward-relevant life events is associated with specific patterns of approach motivation indexed by neural reward activation among reward-hypersensitive individuals compared with MRew individuals. Although this study did not directly test hypotheses related to mood-related psychopathology (e.g., BSD and substance use), it has implications for understanding the pathophysiology of these conditions. According to the reward-hypersensitivity theory, individuals with a trait hypersensitive reward system should exhibit elevated responses to cues signaling both the possible success and failure to obtain reward (Fig. 1). We predicted that among HRew individuals, heightened exposure to reward-system-activating events involving goal striving or goal attainment would be associated with heightened activation in the OFC and NAc (Hypothesis 1) as indicators of reward-system activation. Furthermore, we predicted that among the HRew individuals, heightened exposure to irreconcilable goal failure, involving triggers of approach cessation and deactivation of the reward system, would be associated with lower activation in the NAc (Hypothesis 2). We were less certain about our predictions for the OFC following heightened exposure to irreconcilable goal failure but suggested that reward-hypersensitive individuals may display elevated OFC activation given that they may engage the OFC in a manner that attenuates NAc signaling following failure and loss (i.e., cortico-striatal attenuation tendencies; Ng et al., 2019; Young et al., 2016; Hypothesis 2).
Because the reward-hypersensitivity model highlights the role of approach motivation, a construct that is related to reward anticipation but not outcome (Alloy & Nusslock, 2019; Chase et al., 2013; Nusslock et al., 2012), our hypotheses focused on neural activation during reward anticipation. If we obtained significant effects for reward anticipation, we also explored the effects of reward-relevant life events on activation during reward outcome to assess specificity to reward anticipation. Finally, our objective also was to determine whether any observed distinct neural function occurs specifically during reward processing but not loss processing. Thus, we examined the relationship between trait reward sensitivity, recent exposure to reward-relevant events, and neural activation to rewards above and beyond the effect of loss-related neural activation by adjusting for OFC and NAc activation during loss anticipation or outcome.
Method
Participant recruitment
Participants for the current study came from a larger longitudinal study, the Teen Emotion and Motivation project (Project TEAM), described in detail elsewhere (Alloy et al., 2012). Project TEAM recruited MRew and HRew individuals based on scores falling in the 40th to 60th percentiles (MRew) or top 15th percentile (HRew) on both of two self-report trait reward-sensitivity measures, the Behavioral Activation System Scale (BAS; cut point ≥ 43 for HRew and ≥ 37 and ≤ 39 for MRew; Carver & White, 1994) and the Sensitivity to Reward (SR; cut point ≥ 16 for HRew and ≥ 10.4 and ≤ 12.6 for MRew) subscale of the Sensitivity to Punishment Sensitivity to Reward Questionnaire (SPSRQ; Torrubia et al., 2001). Project TEAM participants completed follow-up assessments that assessed reward-relevant life events approximately every 6 months. All TEAM participants still involved in the study when the fMRI scan was introduced were invited to participate in the MRI session except participants who were excluded on the basis of the following criteria: a lifetime history of psychosis, ferrous metal in any part of the body, lifetime history of head trauma, claustrophobia, left-handedness, and pregnancy. A subset of participants (all right-handed) consented and completed an additional fMRI component approximately 26 months (SD = 28 months) after the start of their participation in Project TEAM. People who participated in the fMRI scan were screened for eligibility (standard MRI exclusion criteria were applied); they then provided informed consent and completed a set of trait and state self-report measures on the day of the fMRI scan. Participants completed the monetary-incentive-delay (MID) reward task (Samanez-Larkin et al., 2007) in the scanner. All study protocols were approved by the institutional review board at Temple University before the start of data collection.
Current study participants
Of the 133 participants who completed the fMRI scan, 26 were excluded because of excessive head motion (> 3 mm), four were excluded because of behavioral task acquisition errors, and 16 were excluded because they were missing life-event data in the 6-month time period before the scan. Thus, participants in the current study included 59 HRew and 28 MRew individuals (N = 87) who had an average age of 21.05 years (SD = 2.08, range = 18–28 years) on the day of the scan. Participants were 52% female, 54% White, 25% Black, 9% Asian, 6% biracial/multiracial, 1% Native American, and 5% other or unknown race. MRew and HRew participants differed on mood disorder history, χ2(1) = 20.704, p < .001. However, they did not differ on age at scan, t(84) = 0.868, p = .388; gender, χ2(1) = 0.386, p = .534; race, χ2(5) = 4.962, p = .421; or whether they were taking psychotropic medication at time of scan, χ2(2) = 0.134, p = .714. Furthermore, participants included in the study did not differ from participants who were excluded on mood disorder history, χ2(1) = 1.244, p = .265; age at scan, t(131) = −0.834, p = .406; gender, χ2(1) = 0.109, p = .741; race, χ2(5) = 2.777, p = .734; whether they were taking psychotropic medication at the time of scan, χ2(2) = 2.233, p = .327; or reward group χ2(1) = 0.177, p = .674. Demographic information and data characteristics by trait-reward-sensitivity group are in Table 1.
Summary of Sample Characteristics by Trait-Reward-Sensitivity Group (N = 87)
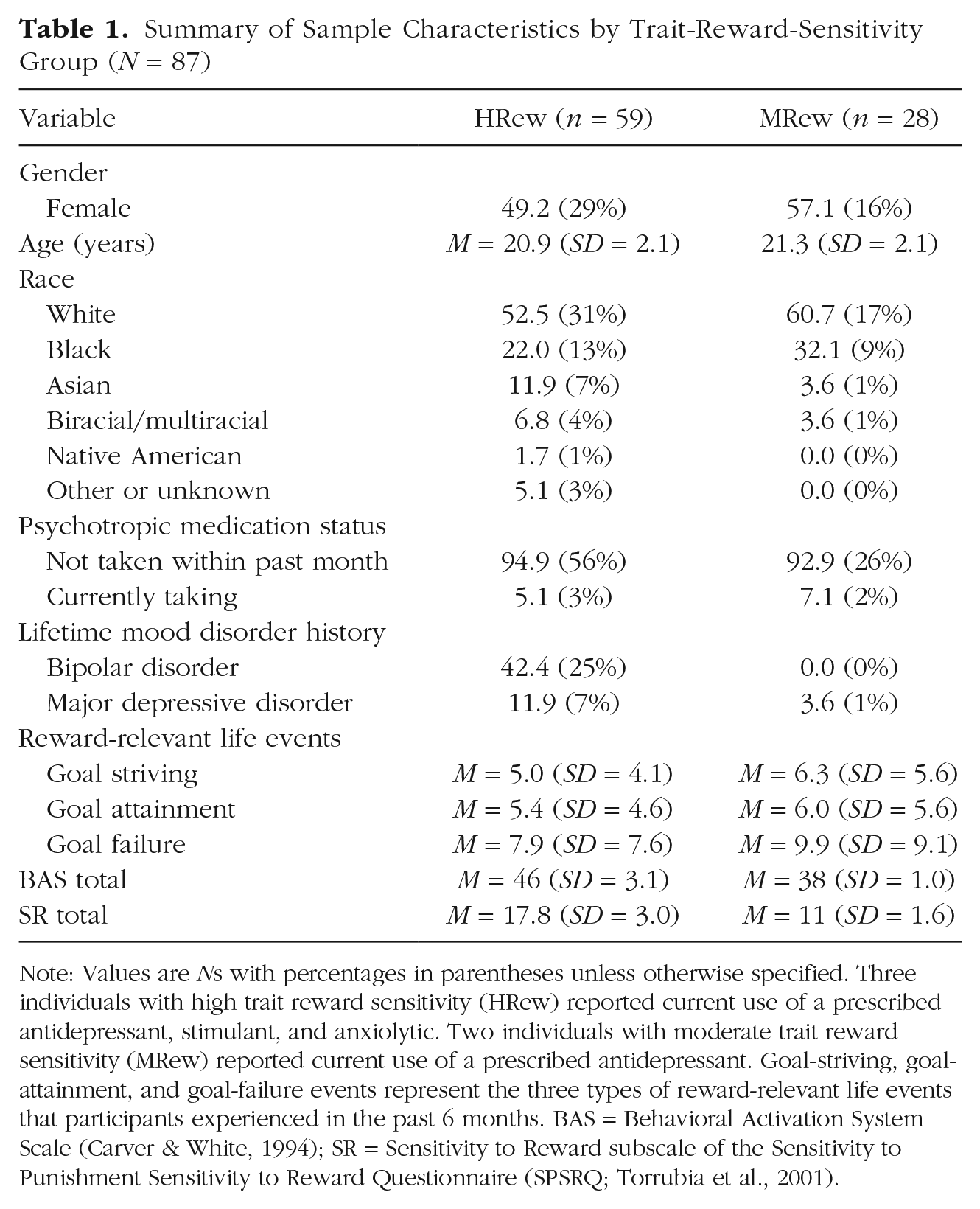
Note: Values are Ns with percentages in parentheses unless otherwise specified. Three individuals with high trait reward sensitivity (HRew) reported current use of a prescribed antidepressant, stimulant, and anxiolytic. Two individuals with moderate trait reward sensitivity (MRew) reported current use of a prescribed antidepressant. Goal-striving, goal-attainment, and goal-failure events represent the three types of reward-relevant life events that participants experienced in the past 6 months. BAS = Behavioral Activation System Scale (Carver & White, 1994); SR = Sensitivity to Reward subscale of the Sensitivity to Punishment Sensitivity to Reward Questionnaire (SPSRQ; Torrubia et al., 2001).
Measures
Behavioral Inhibition System/BAS scales
The Behavioral Inhibition System (BIS)/BAS Scale (BIS/BAS; Carver & White, 1994) is a widely used self-report questionnaire consisting of 20 items that assess trait-level indicators of the behavioral activation (reward sensitivity) and inhibition (punishment sensitivity) systems. Total BAS subscale scores were used to screen participants and determine their eligibility for the MRew and HRew groups in Project TEAM (Alloy et al., 2012). The BAS is dimensional (Liu et al., 2018) and forms a general factor of reward sensitivity (Kelley et al., 2019). Participants were asked to rate items on a scale from 1 to 4 (1 = strongly disagree, 4 = strongly agree); possible total BAS scores range from 13 to 52. The total BAS scale consists of items related to reward-sensitivity constructs such as drive (“I go out of the way to get things I want”), fun seeking (“I often act on the spur of the moment”), and reward responsiveness (“When I’m doing well at something I love to keep at it”). In the baseline TEAM sample, it had acceptable internal consistency (α = .80) and has been shown to have acceptable retest reliability (Meyer et al., 2001).
SPSRQ
Participants also were screened according to the SR subscale of the SPSRQ (Torrubia et al., 2001). The SR subscale consists of 24 yes/no items assessing reward sensitivity (i.e., “Do you often do things to be praised?”; “Do you like to compete and do everything you can to win?”). A sum score was formed from the number of “yes” item responses; possible score range is from 0 to 24. In our baseline sample, the SR subscale had acceptable internal consistency (α = .76), which is supported by the extant literature (αs = .75–.83; Torrubia et al., 2001). At baseline, the BAS-Total and SR scales correlated r = .40 with each other (Alloy et al., 2012).
Life Events Scale/Life Events Interview
The Life Events Scale (LES; Francis-Raniere et al., 2006) is a self-report questionnaire that assesses the presence of a broad range of 193 positive and negative life events that typically occur among adolescents and young adults, including major and minor events related to school, achievement, career, health, finances, family, friends, and romantic relationships. The Life Events Interview (LEI; Francis-Raniere et al., 2006) was completed by trained interviewers to confirm whether the events endorsed by the participant on the LES met the a priori event-definition criteria. Interviewers were trained via review of audiotaped interviews, live observation, and role-playing and were tested on their knowledge of the LEI manual via written exam and mock interview. Only events meeting event-definition criteria that occurred within the 6 months before the fMRI scan were included in the present study. LES events had been rated a priori by three Project TEAM investigators independent of actual interviews with participants as (a) goal-striving events (defined as opportunity to obtain goals/rewards or remove obstacles to goals/rewards; e.g., “you applied for a new job”), (b) goal-attainment events (defined as actually obtaining goals/rewards; e.g., “you won a significant award for your achievements at work”), or (c) goal-failure events involving definite failures to achieve goals or losses (defined as failure to obtain goals/rewards or loss of goals/rewards; e.g., “you received an F in an important course”; for more details, see Urosević et al., 2010). The interrater reliability of the event categorizations was α = .79 for goal-striving events, α = .91 for goal-attainment events, and α = .94 for goal-failure events (Urosević et al., 2010). All qualifying events were summed within each category; thus, scores represent the counts of the number of events within each category that occurred in the 6 months before the fMRI scan. The LES and LEI have demonstrated good reliability and validity in past research (Alloy et al., 2006; Boland et al., 2016; Francis-Raniere et al., 2006).
fMRI MID task
During the fMRI scan, participants completed the MID task (Samanez-Larkin et al., 2007; Fig. 2), a widely used and well-validated measure of neural reward function (Knutson et al., 2000). In a prescan training session, participants were instructed on how to complete the MID. They were told that they would have the opportunity to win money during reward trials (indicated by a circle cue signaling “Win $0.00,” “Win $1.50,” or “Win $5.00”) or avoid losing money during loss trials (indicated by a square cue signaling “Lose $0.00,” “Lose $1.50,” or “Lose $5.00”). The trial cue was presented for 2 s, followed by a jittered fixation and a solid white square. Participants had to press a button while the white square was on the screen to either win (reward trials) or avoid losing (loss trials) money. Next, participants received feedback (2 s) indicating the amount of money won or lost for each trial, followed by another jittered fixation as an intertrial interval. Thus, the MID task allowed us to examine separately neural activity to reward/loss anticipation (the period between presentation of the initial trial cue and presentation of the target square, 2–2.5 s) and reward/loss outcome (the feedback presentation period, 2 s). Initial target duration was calculated from the participant’s reaction time during the prescan trials and then dynamically updated to keep hit rate at approximately 66% (this was calculated separately for each trial type). Participants completed the six trial types eight times in random order, for a total of 96 trials across two MID runs.

Monetary incentive delay (MID) task. The (a) trial structure and (b) possible reward and loss cues of the MID task designed to examine neural activation during anticipation and outcome of monetary reward and loss. Adapted from C. B. Young & R. Nusslock, “Positive Mood Enhances Reward-Related Neural Activity,” Social Cognitive and Affective Neuroscience, 2016, Vol. 11, Iss. 6, pp. 934–944, by permission of the Oxford University Press. Copyright 2016.
FMRI data acquisition and analysis
Neuroimaging data were collected at Temple University Medical Center using a 3.0-Tesla Verio wide-bore MRI scanner (Siemens Medical Solutions USA, Malvern, PA) with a standard 12-channel head coil. Functional blood-oxygen-level-dependent scans were collected using the following parameters: coverage = 36 4-mm axial slices (field of view [FOV] = 236 mm), matrix = 64 × 64, voxel size = 3.7 × 3.7 × 4.0 mm, repetition time (TR) = 2,000 ms, echo time (TE) = 25 ms, flip angle = 70°, acquisition volumes = 292. Structural three-dimensional magnetization-prepared rapid acquisition with gradient echo (MPRAGE) scans also were collected in the sagittal plane with the following parameters: voxel size = 0.5 × 0.5 × 1.0 mm, TR = 1,600 ms, TE = 2.46 ms, FOV = 252, flip angle = 9°, acquisition volumes = 176.
Data were analyzed using a general linear model in SPM8 (Wellcome Trust Centre for Neuroimaging, London, England). Functional images were realigned and corrected for errors in slice-timing. Images then were spatially normalized to Montreal Neurological Institute (MNI) space and smoothed using a 6-mm full width at half maximum Gaussian kernel. We computed translational movement in millimeters (x, y, z) and rotational motion in degrees (pitch, roll, yaw) on the basis of SPM8 parameters to correct motion for the functional images in each participant. The final sample had less than 3 mm of head movement.
The hemodynamic signal was deconvolved using a general linear model identifying the six trial types during the MID anticipation and outcome phases. The anticipation phase was defined as the period after presentation of the cue indicating the possibility to win or lose money but before presentation of the target square (2–2.5 s). The outcome phase was defined as the period after presentation of the feedback (2 s). Six variables of no interest for motion were included. First-level voxel-wise t statistics were computed for each participant in which reward trials (i.e., “Win $1.50,” “Win $5.00”) were contrasted with nonreward trials (i.e., “Win $0.00”) to calculate reward anticipation and outcome; loss trials (i.e., “Lose $1.50,” “Lose $5.00”) were compared with nonloss trials (i.e., “Lose $0.00”) to calculate loss anticipation and outcome. We combined $1.50 and $5.00 trials to be consistent with previous research (e.g., Samanez-Larkin et al., 2007), for ease of interpretation, and to heighten reliability (i.e., more trials in the combined score).
Significant clusters of activation at the whole-brain level were determined at a voxel-wise height threshold of p < .001 uncorrected, and a family-wise error correction was determined at the cluster level for multiple spatial comparisons (p < .05, k = 20 voxels). All reported coordinates are in the standard MNI space.
We extracted parameter estimates (β weights) from predefined regions of interest (ROIs) for the NAc and OFC during reward and loss anticipation and outcome and exported these parameter estimates into the R software environment (Version 4.0.4; R Core Team, 2021) for analyses. For the bilateral NAc (Fig. 3a) and bilateral OFC (Fig. 4a), we used an anatomically defined bilateral ROI mask using the Harvard Oxford Atlas. The masks met a probabilistic threshold of at least 25%. The use of this OFC ROI mask provides the advantage of maximizing the balance between Type I error and Type II error. On one hand, applying multiple OFC ROIs to detect activation in smaller regions would increase risk of Type I error. On the other hand, using an ROI mask covering the entire OFC would require a large effect to detect significant associations and thus increase risk for Type II error. Although excluding portions of the supramedial OFC, the Harvard Oxford Atlas mask does cover a relatively large portion of the OFC and at the same time lowers risk for false-negative findings. Furthermore, prior literature on the link between reward-related brain function and psychiatric conditions characterized by reward abnormalities (e.g., depression, mania, substance use) often report effects in more lateral portions of the OFC (Forbes et al., 2014; Nusslock et al., 2012). This suggests that the OFC areas covered by the Harvard Oxford Atlas mask are particularly relevant to this area of research.
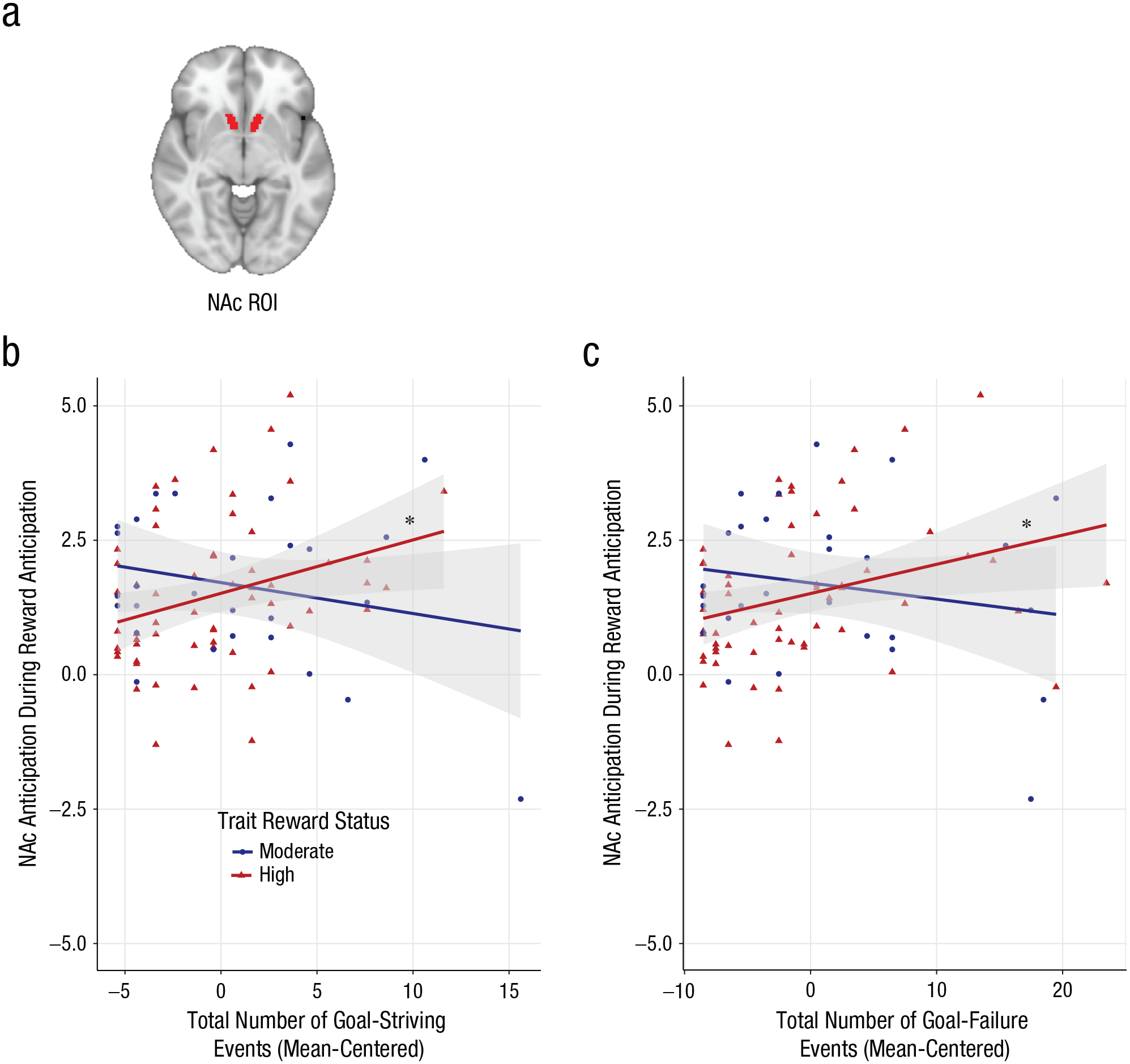
Exposure to reward-relevant life events and bilateral nucleus accumbens (NAc) activation by trait reward status. (a) Region of interest (ROI) for the bilateral NAc structurally derived with Harvard Oxford Atlas. (b) Total goal-striving events (centered) as a function of activation in the NAc during reward anticipation for individuals with moderate trait reward status compared with high trait reward status. (c) Total goal-failure events (centered) as a function of activation in the NAc during reward anticipation for individuals with moderate trait reward status compared with high trait reward status. Asterisks indicate significant simple slopes (*p < .05).
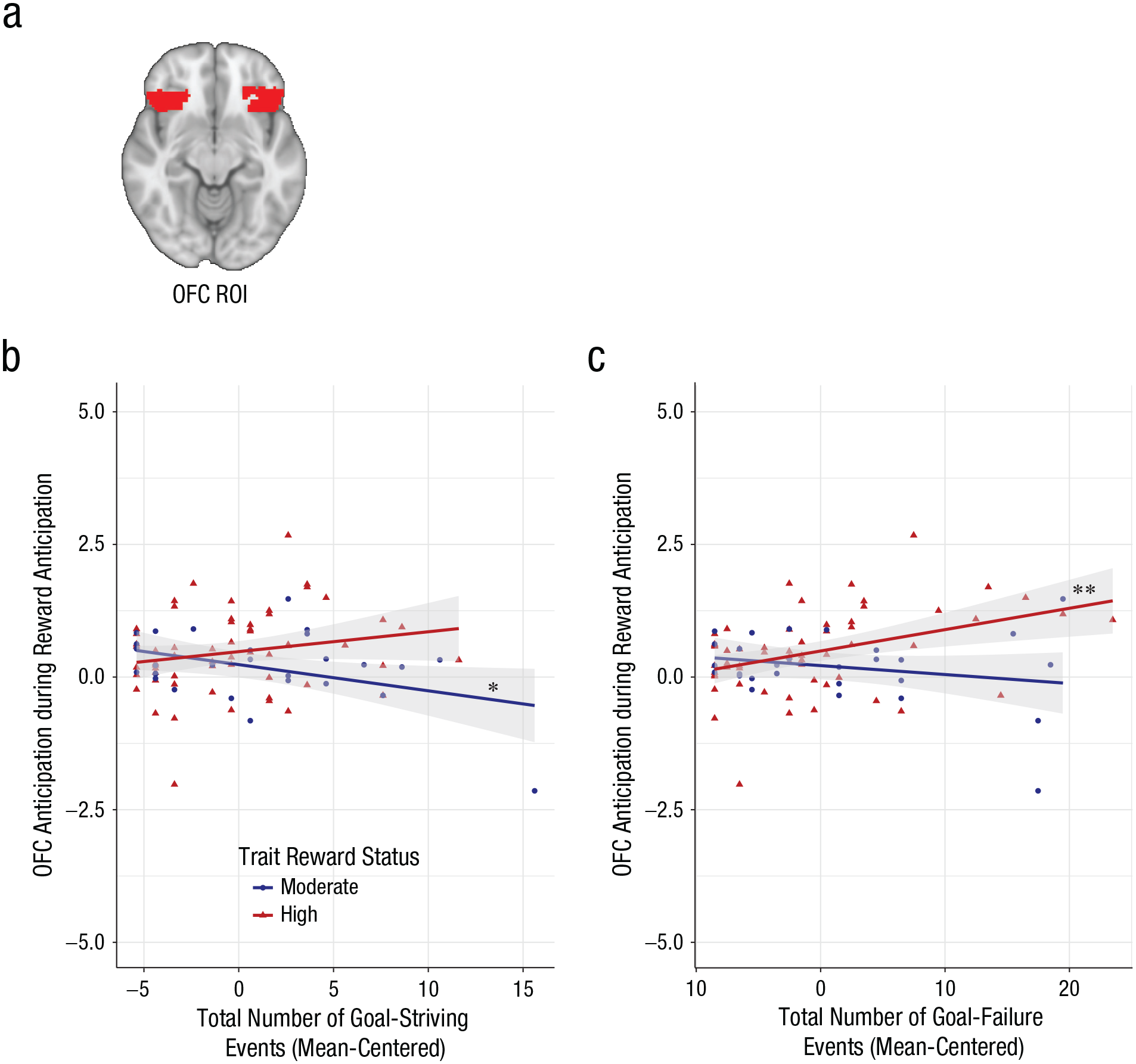
Exposure to reward-relevant life events and bilateral orbitofrontal cortex (OFC) activation by trait reward status. (a) Region of interest (ROI) for the bilateral OFC as defined by the Harvard Oxford Atlas. (b) Total goal-striving events (centered) as a function of activation in the OFC during reward anticipation for individuals with moderate trait reward status compared with high trait reward status. (c) Total goal-failure events (centered) as a function of activation in the OFC during reward anticipation for individuals with moderate trait reward status compared with high trait reward status. Asterisks indicate significant simple slopes (*p < .05. **p < .01).
Statistical analysis approach
All analyses were conducted in R using the R packages psych (Version 2.0.12; Revelle, 2020), haven (Version 2.2.0; Wickham & Miller), reghelper (Version 1.0.2; Hughes, 2021), dplyr (Version 1.0.4; Wickham et al., 2021), and ggplot2 (Version 3.3.5; Wickham, 2016). First, we conducted preliminary analyses using bivariate correlations (if the potential covariates were continuous variables) and independent samples t tests (if the potential covariates were categorical variables) to determine whether demographics or clinical characteristics should be included as covariates in the primary analyses. The variables were included as covariates if they were associated with the dependent variables (i.e., reward-related neural activation). The main effects of reward and loss during the anticipation and outcome phases also were examined to ensure that the MID task activated expected regions. Next, we employed moderation analyses to examine whether trait-reward-sensitivity group (MRew or HRew) moderated the relationship between the frequency of occurrence of goal-striving, goal-attainment, and goal-failure life events in the 6 months before the fMRI scan and neural activation during reward anticipation and outcome in the NAc and OFC. Thus, we ran three separate sets of primary analyses for each of the two ROIs (activation to reward anticipation in the NAc and OFC) with goal-striving, goal-attainment, or goal-failure events as the independent variable and reward-sensitivity group as the moderator. To protect against Type I error inflation as a result of multiple comparisons, we applied the Fisher’s protected t test, which required a significant omnibus interaction result to proceed to simple slope analyses (Cohen et al., 2013). If there were significant Group × Events interactions for neural reward anticipation, follow-up exploratory analyses were conducted for activation in the corresponding region during reward outcome to confirm specificity to reward anticipation. The first step in the regression models included any covariates found to be related to reward-related neural activation in the NAc or OFC during anticipation or outcome. In the next step, we included the mean-centered total number of goal-striving, goal-attainment, or goal-failure events and the trait-reward-sensitivity group, followed by the product term of mean-centered life events and trait-reward-sensitivity group. To determine whether the results hold above and beyond mood disorder history, we repeated the aforementioned primary analyses with the addition of mood disorder history as a covariate (see Tables S1 and S2 in Supplemental Material available online).
Results
Preliminary analyses
Preliminary analyses were conducted to examine whether there were associations between potential covariates (age at the time of the scan, gender, a history of mood disorder diagnosis, use of psychotropic medication at the time of scan, NAc activation during loss anticipation and outcome, and OFC activation during loss anticipation and outcome) and the dependent variables (NAc activation during reward anticipation and outcome and OFC activation during reward anticipation and outcome; Miller & Chapman, 2001).
Participant age at the time of the scan was not significantly associated with NAc activation during reward anticipation (r = –.004, p = .972), OFC activation during reward anticipation (r = .040, p = .715), NAc activation during reward outcome (r = .090, p = .413), or OFC activation during reward outcome (r = .119, p = .280). Gender was not significantly associated with NAc reward anticipation, t(82) = −1.074, p = .286; OFC reward anticipation, t(82) = −0.391, p = .697; NAc reward outcome, t(82) = 0.050, p = .960; or OFC reward outcome, t(82) = 0.416, p = .679. Having a history of a mood disorder diagnosis was not significantly associated with NAc reward anticipation, t(85) = 0.014, p = .988; OFC reward anticipation, t(85) = −1.660, p = .101; NAc reward outcome, t(85) = −1.138, p = .258; or OFC reward outcome, t(85) = −1.159, p = .250. Use of psychotropic medication at the time of the scan was not significantly associated with NAc reward anticipation, t(85) = −0.028, p = .978; OFC reward anticipation, t(85) = −1.040, p = .301; NAc reward outcome, t(85) = −0.955, p = .342; or OFC reward outcome, t(85) = −1.783, p = .078. Thus, none of these variables were included as covariates in the primary analyses because they did not relate to the NAc and OFC activation dependent variables.
Significant associations were found between OFC activation during anticipation of reward and during anticipation of loss (r = .282, p = .004) and between OFC activation during reward- and loss-related outcomes (r = .349, p < .001). Likewise, NAc activation during reward anticipation was associated with NAc activation during loss anticipation (r = .346, p < .001). There also was a significant association between NAc activation during reward outcome and NAc activation during loss outcome (r = .645, p < .001). Thus, these loss variables were retained as the only covariates in the primary analyses.
Finally, our sample displayed whole-brain clusters of neural activity in expected regions during the MID (see the Supplemental Material).
Primary analyses
Main effects for trait reward sensitivity and frequency of recent reward-relevant events
We did not detect any main effects for trait-reward-sensitivity group on NAc (β = 0.022, SE = 0.277, t = 0.080, p = .937, change in (∆) R2 < .001) or OFC (β = 0.206, SE = 0.151, t = 1.365, p = .175, ∆R2 = .017) activation to reward anticipation. There were no main effects for frequency of recent goal-striving life events (NAc: β = 0.021, SE = 0.031, t = 0.658, p = .513, ∆R2 = .004; OFC: β = −0.009, SE = 0.017, t = −0.500, p = .618, ∆R2 = .003), goal-attainment life events (NAc: β = 0.033, SE = 0.029, t = 1.107, p = .271, ∆R2 = .013; OFC: β = 0.005, SE = 0.016, t = 0.290, p = .772, ∆R2 = .001), or goal-failure life events (NAc: β = 0.017, SE = 0.018, t = 0.970, p = .335, ∆R2 = .010; OFC: β = 0.015, SE = 0.010, t = 1.530, p = .130, ∆R2 = .026) on ROI activation to reward anticipation. Next, we report results on the interaction between the frequency of each type of reward-relevant events and trait-reward-sensitivity group on ROI activation.
Frequency of Recent Reward-Relevant Events × Trait Reward Sensitivity interaction effects
Goal-striving life events
As predicted in Hypothesis 1, the interaction between trait-reward-sensitivity group and recent goal-striving events was associated with NAc activation to reward anticipation (β = 0.158, SE = 0.061, t = 2.581, p = .012, ∆R2 = .067; Fig. 3b). Specifically, greater exposure to goal-striving events was associated with heightened activation in the NAc among HRew individuals (β = 0.092, SE = 0.042, t = 2.207, p = .03, ∆R2 = .078) but not MRew individuals (β = −0.066, SE = 0.045, t = −1.468, p = .146, ∆R2 = .066).
The interaction of trait reward sensitivity and goal-striving events also was associated with OFC activation during reward anticipation (β = 0.088, SE = 0.033, t = 2.645, p = .010, ∆R2 = 0.074; Fig. 4b) such that greater exposure to goal-striving events was associated with decreased activation in the OFC among MRew individuals (β = −0.053, SE = 0.025, t = −2.155, p = .034, ∆R2 = .208). Contrary to Hypothesis 1, however, the association between these events and OFC activation did not reach statistical significance among HRew individuals (β = 0.035, SE = 0.023, t = 1.556, p = .123, ∆R2 = .037).
Given the significant interaction between trait-reward-sensitivity group and goal-striving events for NAc and OFC activation during reward anticipation, we conducted follow-up analyses for neural activation during reward outcome to examine specificity of the observed effects. We did not detect an interaction between trait-reward-sensitivity group and goal-striving events on NAc activation during reward outcome (β = 0.058, SE = 0.055, t = 1.063, p = .291, ∆R2 = .008) or OFC activation during reward outcome (β = 0.007, SE = 0.035, t = 0.204, p = .839, ∆R2 < .001).
Goal-attainment life events
Partially consistent with Hypothesis 1, the interaction of trait-reward-sensitivity group and recent goal-attainment events was associated with OFC activation during reward anticipation (β = 0.070, SE = 0.032, t = 2.147, p = .035, ∆R2 = .050). However, the simple slope analysis revealed that exposure to goal-attainment events was not associated with OFC activation among HRew individuals (β = 0.036, SE = 0.021, t = 1.696, p = .094, ∆R2 = .048) or MRew individuals (β = −0.034, SE = 0.025, t = −1.378, p = .172, ∆R2 = .081). The interaction between trait-reward-sensitivity group and goal-attainment events on NAc activation during reward anticipation did not reach statistical significance (β = 0.090, SE = 0.061, t = 1.494, p = .139, ∆R2 = .024).
Goal-failure life events
The interaction of trait-reward-sensitivity group and recent goal-failure events was associated with NAc activation during reward anticipation (β = 0.086, SE = 0.035, t = 2.443, p = .017, c∆R2 = .061; Fig. 3c). However, counter to predictions in Hypothesis 2, greater exposure to goal-failure events predicted heightened NAc activation among HRew individuals (β = 0.051, SE = 0.022, t = 2.272, p = .026, ∆R2 = .083). The association between goal-failure events and NAc activation was not significant among MRew individuals (β = −0.036, SE = 0.028, t = −1.296, p = .199, ∆R2 = .052).
The interaction between trait-reward-sensitivity group and goal-failure events was associated with OFC activation during reward anticipation (β = 0.055, SE = 0.019, t = 2.908, p = .005, ∆R2 = .085; Fig. 4c). The conditional effects revealed that greater exposure to goal failures and losses was associated with greater OFC activation among HRew individuals (β = 0.039, SE = 0.012, t = 3.251, p = .002, ∆R2 = .152) but not MRew individuals (β = −0.016, SE = 0.015, t = −1.106, p = .272, ∆R2 = .046).
Given the significant interaction results between trait-reward-sensitivity group and goal-failure events for NAc and OFC activation during reward anticipation, we conducted follow-up analyses for neural activation during reward outcome as well. There was no significant interaction between trait-reward-sensitivity group and goal-failure events on NAc activation (β = −0.020, SE = 0.032, t = −0.643, p = .522, ∆R2 = .003) or OFC activation (β = −0.018, SE = 0.020, t = −0.887, p = .378, ∆R2 = .008) during reward outcome.
Supplemental analyses
The results of primary analyses were largely unaffected by the inclusion of mood disorder history as an additional covariate. Summary of these statistical models compared with primary analyses is shown in Tables S1 and S2 in the Supplemental Material.
Discussion
According to the reward-hypersensitivity model, specific reward-relevant life events are posited to play a critical role in excessive activation or deactivation of a hypersensitive reward system. The results of the present analyses offer partial support for the hypothesized relationships of recent reward-relevant life events with neural activation during reward anticipation among HRew individuals compared with MRew individuals. Specifically, the findings indicate that greater exposure to recent reward-relevant life events was associated with distinct reward-related neural function among reward-hypersensitive individuals. Note that recent reward-system-activating life events (goal striving but not goal attainment) and reward-system-deactivating (goal failure) life events each were associated with elevated neural activation patterns in reward-hypersensitive individuals. Furthermore, such associations appeared specific to reward anticipation but not reward outcome.
Partially supporting Hypothesis 1, both NAc and OFC activation during reward anticipation were associated with the interaction between exposure to recent goal-striving life events and trait reward hypersensitivity. We found that among HRew individuals, NAc reward anticipation was higher for individuals exposed to a greater number of recent goal-striving life events, which is in line with prior studies finding that elevated NAc reward anticipation is a neural correlate of psychopathology (Harada et al., 2013; Nusslock et al., 2012) and the role of goal-striving events in precipitating clinical symptoms (Nusslock et al., 2007). However, contrary to our expectations, there was no difference in OFC reward anticipation across levels of exposure to recent goal-striving life events for the HRew group. Instead, greater exposure to recent goal-striving life events was associated with lower OFC reward anticipation for the MRew group. This finding may be explained by the “coasting” phenomenon (Fulford et al., 2010), in which most individuals tend to coast (i.e., not work as hard) after an extensive level of goal striving; however, individuals who are hypersensitive to goal-striving events may not coast but continue to push through. In other words, whereas MRew individuals displayed lower OFC anticipation following a high level of exposure to goal-striving events, there was no difference for HRew individuals regardless of frequency of exposure to goal-striving events. Although we detected that exposure to goal-striving events was differentially associated with NAc and OFC activation between the trait-reward groups, there were no comparable results for exposure to goal-attainment events. This may be attributable to power issues given that the current sample size was sufficiently powered (.80) to detect a small-moderate effect (i.e., r2 ≈ .12), which is higher than the observed effects. Future work should attempt to replicate these findings in a larger sample.
In examining Hypothesis 2, we found that greater exposure to recent goal-failure life events was associated with heightened OFC reward anticipation for HRew individuals but not MRew individuals. Note that among the HRew individuals, greater exposure to recent goal-failure events also was associated with greater NAc reward anticipation. One potential interpretation is that this reflected inefficiency in down-regulation of reward responsiveness among HRew individuals exposed to recent goal-failure events. For instance, there might have been a greater demand for OFC activation to provide optimal regulatory control of the NAc, which had been hyperactivated during reward anticipation. The OFC projection of inhibitory input to subcortical regions might have been inefficient and thus resulted in the observation of hyperactivation in NAc during reward anticipation. Another potential interpretation is that HRew individuals up-regulated, rather than down-regulated, reward responsiveness in the face of loss and failures, which reflects resilience and persistence. In other words, heightened OFC and NAc activation might have reflected hyperresponsiveness to reward. This may be due, in part, to the perceived nature of the goal-failure life events that were assessed in this study. Specifically, most of these life events might not necessarily reflect definitive failures or losses that could not be overcome. It is also possible that HRew individuals perceive these failures or losses as remediable challenges that can be overcome such that exposure to these events activated rather than deactivated the reward system among HRew individuals and thereby were associated with a greater level of OFC and NAc reward anticipation following frequent exposure to these events. Future research should test the relationship between reward sensitivity, reward-related brain function, and irreconcilable reward-related failures/losses compared with remediable reward-related failures/losses. Self-report questions assessing whether a reported goal-failure life event has been construed as a definitive failure compared with remediable challenge also may help clarify the nature of goal-failure events an individual experienced.
Finally, as predicted, the current findings demonstrate that exposure to reward-relevant life events specifically was associated with neural activation during reward anticipation rather than outcome. This finding is in line with the reward-hypersensitivity model, which suggests that HRew individuals should be particularly sensitive to excessive increases in approach motivation to cues that signal the chance to obtain rewards as opposed to the receipt of those rewards (Alloy & Abramson, 2010; Alloy et al., 2016; Alloy & Nusslock, 2019).
The present study and prior studies that examined the relationship between stressors and reward-related brain function yielded inconsistent results. Such discrepancies may have resulted from the heterogeneity of samples and study measurements across these studies (Berghorst et al., 2016; Kumar et al., 2014, 2015; Ossewaarde et al., 2011; Treadway et al., 2013). To our knowledge, the present study was the first to specifically investigate life events according to their reward- or goal-pursuit status when examining the link between exposure to life events and reward-related neural function. This approach allows us to identify the precise aspect of life-event exposure (i.e., the reward-related events’ ability to trigger reward-system activation and deactivation) that might have played a role in reward-related brain function but that has not otherwise been examined in prior literature (Berghorst et al., 2016; Kumar et al., 2014, 2015; Ossewaarde et al., 2011; Treadway et al., 2013). Furthermore, compared with lab-induced stressors that were administered in related research (Berghorst et al., 2016; Kumar et al., 2014; Ossewaarde et al., 2011), the assessment of naturally occurring life events in the current study allowed for greater ecological validity and understanding of naturalistic responses to the events.
The present study also was the first to include individuals varying in trait reward sensitivity when examining the associations between reward-relevant life-event exposure and reward-related neural function. The prior studies included only healthy participants (Kumar et al., 2014; Ossewaarde et al., 2011; Treadway et al., 2013) or individuals with mood disorders (Berghorst et al., 2016). Although Kumar et al. (2015) did examine both individuals with and without depression, neither that study nor the aforementioned ones examined trait reward sensitivity, a stable characteristic that might generate distinct reward-related neural function following stress exposure. In fact, our study demonstrated that the relationship between reward-relevant life-event exposure and reward-related brain function was unique among HRew individuals compared with MRew individuals. This finding supports our claim that it is critical to also consider this stable personality factor. Our data suggest that trait reward sensitivity or event exposure alone was not sufficient to explain the variance of activation in the OFC and NAc during reward anticipation, given a lack of significant main effects. In sum, the interaction between trait reward sensitivity and reward-related event exposure offers a more comprehensive picture of neural reward processing.
Our findings have implications for understanding psychiatric conditions characterized by abnormal reward function. Abnormal trait reward sensitivity, exposure to reward-relevant life events, and distinct reward brain function each separately have been associated with psychiatric conditions, including bipolar disorder, major depression, and substance use disorder (for a review, see Nusslock & Alloy, 2017). However, the prior literature yielded heterogeneous findings. This might be attributable to examination of these constructs in isolation. In line with this claim, the current study provides preliminary evidence that these constructs may modulate each other rather than contribute to psychopathology in isolated fashion. Thus, a research model that involves all three constructs may yield better understanding of the pathophysiology of psychiatric disorders. For example, assessment of trait reward sensitivity, exposure to reward-relevant life events, and reward-related neural function may lead to more precise estimation of risk level for a psychiatric condition given the current findings suggesting that trait reward sensitivity or exposure to life events alone may not be a reliable predictor. This is speculative, at best, because in the current study, we did not actually examine whether the observed distinct neural function was associated with psychiatric symptoms. However, in line with this speculation, prior work has demonstrated that altered function in the OFC and NAc may confer neural risk for mood psychopathology (Chase et al., 2013; Nusslock et al., 2012; Satterthwaite et al., 2015; Whitton et al., 2015; Zald & Treadway, 2017). Future research is necessary to examine the combination of trait reward sensitivity and exposure to reward-relevant life events in neural reward function in individuals with psychiatric disorders characterized by elevated reward function, such as BSD and substance use disorder, or blunted reward function, such as major depression, and individuals without psychiatric disorders.
The findings of the current study were bolstered by several strengths. First, the utilization of contextual life-events interviews with objective event definition criteria offered an opportunity to capture objective reward cues present in natural settings while minimizing self-report biases. Second, the study design assessed the reward-relevant life events temporally before the fMRI scan. Third, the criteria used to classify individuals into the HRew and MRew groups were based on data from a large community sample. This approach allowed us to maximize the difference in reward sensitivity between the HRew and MRew groups and established the two groups using absolute levels of moderate reward sensitivity compared with high reward sensitivity.
However, this study also had some limitations. First, the present findings cannot yield causal claims given the absence of a baseline fMRI scan visit. Despite the fact that the events assessed occurred before the fMRI scan, the temporal precedence of these effects did not allow for ruling out the possibility that reward hypersensitivity generates more exposure to reward-relevant life events. Future work should adopt a more complete longitudinal design that offers the opportunity to examine the relationship between reward-relevant life events and reward-related neural activation over time with repeated scans and repeated events assessments. Second, recall bias might occur given the retrospective nature of the life-events interview. Although other observational approaches, such as ecological momentary assessment, can reduce this bias, the life-event interview has an advantage of minimizing the interruption of participants’ daily life and time burden to participate in the study. Furthermore, the present study reduced memory bias by including only reported events that had an onset or offset date within 12 months before the interview. Third, the current results may be limited because Project TEAM excluded a low-trait-reward-sensitivity group to address the project’s primary aims. For instance, it is unclear whether individuals with low trait reward sensitivity yield distinct or share similar reward-related brain function compared with the HRew and MRew groups following exposure to reward-relevant life events.
Furthermore, because of the unequal sample size across the trait-reward-sensitivity groups, there was less information to estimate the effects of reward-relevant life-event exposure on neural activation for the smaller group (i.e., MRew) than the larger group (i.e., HRew), and the unbalanced sample size could reduce statistical power (Aguinis, 1995). Thus, future work should examine this question on a full spectrum of trait reward sensitivity with equal sample size across the subgroups. Approximately a third of our sample reported a lifetime mood disorder history, which may help to explain the heightened motion observed in our sample and why a large number of participants did not meet inclusion criteria for MRI analyses. However, our findings were comparable with or without a lifetime mood disorder history as a covariate, which offers support that the observed results were not a product of mood disorder history. In addition, although the current study reported gender, race, and ethnicity, other demographic data, such as education level and socioeconomic status, were not collected and thus limited the generalizability of the reported findings. Finally, this study focused on the presence but not intensity of reward-relevant life events. However, it is possible that intensity of events also is relevant given that related research demonstrated associations between stressor severity and reward-related neural activation (Kumar et al., 2015; Treadway et al., 2013). Future work should compare the effects of frequency of reward-relevant event exposure and intensity of reward-relevant event exposure.
Conclusions
Notwithstanding the limitations, the current study is the first to examine reward-related neural function following exposure to naturally occurring reward-relevant life events among individuals with high levels of reward sensitivity compared with moderate levels of reward sensitivity. In results partially consistent with the reward-hypersensitivity model, we found that trait levels of reward sensitivity moderated the relationship between exposure to reward-relevant life events and reward-related brain function. The present work advances the understanding of reward-related brain function by highlighting that exposure to reward-relevant life events and trait reward sensitivity jointly may moderate OFC and NAc activation specifically during reward anticipation but not outcome. Although trait reward hypersensitivity, stress exposure, and reward-related neural function each have been linked to psychopathology in isolation, the results of the present study support the need to examine reward-related psychiatric conditions using a comprehensive model that incorporates the interaction among these constructs.
Supplemental Material
sj-pdf-1-cpx-10.1177_21677026211056627 – Supplemental material for The Interplay Between Reward-Relevant Life Events and Trait Reward Sensitivity in Neural Responses to Reward Cues
Supplemental material, sj-pdf-1-cpx-10.1177_21677026211056627 for The Interplay Between Reward-Relevant Life Events and Trait Reward Sensitivity in Neural Responses to Reward Cues by Iris Ka-Yi Chat, Erin E. Dunning, Corinne P. Bart, Ann L. Carroll, Mora M. Grehl, Katherine S. F. Damme, Lyn Y. Abramson, Robin Nusslock and Lauren B. Alloy in Clinical Psychological Science
Footnotes
Transparency
Action Editor: Jennifer Lau
Editor: Jennifer L. Tackett
Author Contributions
I. K-Y. Chat, E. E. Dunning, C. P. Bart, L. B. Alloy, R. Nusslock, and L. Y. Abramson contributed to study conception and design. Testing and data collection and processing were contributed substantially by E. E. Dunning, C. P. Bart, A. L. Carroll, and K. S. F. Damme. I. K-Y. Chat, E. E. Dunning, C. P. Bart, and A. L. Carroll performed the data analysis and interpretation under the supervision of L. B. Alloy and R. Nusslock. I. K-Y. Chat, E. E. Dunning, C. P. Bart, and A. L. Carroll drafted the manuscript, and L. B. Alloy and R. Nusslock provided critical revisions. M. M. Grehl contributed substantially to data visualization and the revision of the manuscript. All of the authors approved the final manuscript for submission.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.