
Abstract
Cancer is a major cause of mortality worldwide, including in Indonesia. Busulfan, a chemotherapeutic agent for hematopoietic malignancies, is known to impair female reproductive function and induce ovarian failure. Physalis angulata L. contains phytosteroids with the potential to restore hormonal homeostasis. This study evaluated the effects of P. angulata leaf extract on estrous cycles and pregnancy outcomes in female rats exposed to busulfan. Antioxidant activity, phytosteroid, phenolic, and flavonoid contents were analyzed in vitro. In vivo experiments employed a randomized design with five groups: control, busulfan (40 mg/kgBW), P. angulata (300 mg/kgBW for 28 days), sequential busulfan—P. angulata, and simultaneous busulfan— P. angulata. Metabolite profiling was performed using LC-HRMS, while in silico analyses applied network pharmacology, molecular docking, and molecular dynamics simulations. The results showed moderate antioxidant activity attributed to phytosteroids, phenolics, and flavonoids. P. angulata extract improved busulfan-induced estrous cycle and ovarian histology disruption but paradoxically reduced pregnancy rates. Seventy-seven metabolites were identified, with Physalin F acting as an estrogen receptor 1 (ESR1) antagonist and Albanol A as an ESR1 agonist; Albanol A demonstrated superior stability in molecular dynamics simulations. In conclusion, P. angulata leaf extract exerts dual mechanisms: ameliorating reproductive cycle impairment while reducing pregnancy rates in busulfan-treated female rats.
Introduction
Cancer is the leading cause of death worldwide, causing nearly 10 million deaths in 2020 alone. Globally, there were 20 million new cancer cases and 9.7 million deaths in 2022, with an estimated 77% increase by 2050. 1 Cancer is the third leading cause of death in Indonesia, with 408,661 new cancer cases recorded in 2022 and a mortality rate of 242,099 cases. The number of cases is projected to increase by 63% between 2025 and 2040. 2
Busulfan (1,4-butanediol dimethanesulfonate) is a chemotherapy drug designed to destroy hematopoietic cancer cells. 3 Busulfan damages the DNA of cells, forcing them to stop proliferating. However, busulfan does not have a specific target cell; therefore, all cells with high proliferative activity, including granulosa cells in the ovarian follicles, are destroyed. Damage to the granulosa cells of ovarian follicles results in an imbalance in the hormones that regulate the ovulatory cycle, disrupting the cycle and leading to ovarian failure. 4 The addition of busulfan to the chemotherapy regimen caused ovarian failure in up to 100% of girls aged 4.6–17 years. This rate was significantly higher compared to the group without busulfan, which showed an incidence of only 27% (P < 0.002). 5 The clinical use of busulfan significantly increases the incidence of irreversible ovarian failure in prepubertal and postpubertal female patients, as evidenced by the low rates of spontaneous puberty (38.1% in the prepubertal group) and spontaneous menarche (10% in the postpubertal group), compared to the treosulfan group, which achieved a 100% success rate. 6 Ovarian failure accounts for 24% of all infertility cases. 7
Infertility is defined as a decrease in reproductive ability. 8 The prevalence of infertility in women worldwide from 1990 to 2021 increased by 84.44%, from 59,690,000 to 110,089,459 cases. 9 A healthy reproductive cycle in women is characterized by a regular menstrual cycle, which typically includes ovulation. Ovulation is the process of releasing mature oocytes from the female ovaries. During maturation, follicles are sensitive to various stressors, including exposure to chemical drugs. Exposure to chemical drugs can cause endocrine disorders, polycystic ovary syndrome, follicular atresia, and premature ovarian failure. 10 Ovulation cycle disorders caused by busulfan can be minimized by repairing damage to ovarian granulosa cells and balancing ovarian cycle-regulating hormones. 11
Ground berry (Physalis angulata L.) in Indonesia, known as Ciplukan, is a traditional Indonesian herb. P. angulata is commonly used as a medicine for asthma, diabetes, chicken pox, cough, fever, diarrhea, hypertension, and back pain. 12 The medicinal role of P. angulata has been reported to improve kidney damage, 13 acting as an antioxidant, anti-fibrotic agent for the liver, and an anti-inflammatory agent. 14 P. angulata leaf extract reportedly contains alkaloids, flavonoids, tannins, saponins, and steroids. 15 The phytosteroids (steroids found in plants) identified in P. angulata belong to the withanolide group, which comprises naturally occurring steroid lactones with an ergane basic skeleton. 16 Structurally, withanolides closely resemble human sex steroid hormones, which enables them to bind effectively to steroid receptors in both humans and animals. In the body, phytosteroids can mimic the role of sex steroid hormones, such as estrogen, in females. 17 These phytosteroids are expected to facilitate the stabilization of hormone levels that have declined due to ovarian failure. Consequently, the administration of phytosteroids to individuals with reproductive disorders may mitigate these impairments.
P. angulata has been tested on male reproductive organs and has been proven to increase testicular weight, seminiferous tubule diameter, and percentage of intact spermatozoa. 18 However, no studies have been conducted on female reproductive functions. This study aimed to analyze the potential of P. angulata leaf extract to improve the reproductive cycle and pregnancy success rate in female rats treated with busulfan in vivo, supported by in vitro data and in silico analysis.
Materials and Methods
Materials
P. angulata leaves were obtained from Center for Assembling and Testing of Spice, Medicinal and Aromatic Plants (former BALITTRO), Cimanggu, Bogor, Indonesia, which were planted at an altitude of 300–400 m above sea level, with a rainfall of 2000 mm/year, and an average air temperature of 27°C, with the lowest temperature of 23.3°C and the highest reaching 30.8°C. P. angulata L was identified, classified, and validated by the Herbarium Bogoriense, National Research and Innovation Agency (BRIN) (No. B-1070/II.6.2/IR.01.02/3/2026). All leaves in the habitus were used, except for the yellow ones. Busulfan was obtained from Sigma-Aldrich→. Sprague-Dawley white rats were obtained from the Indonesian Food and Drug Monitoring Agency (BPOM). The use of animal models was approved by the Animal Ethics Commission of the School of Veterinary Medicine and Biomedical Sciences (SKHB) at IPB University (No. 247/KEH/SKE/IX/2024).
P. angulata Leaf Extraction
P. angulata leaves were oven-dried at 40°C–60°C for 5–6 hours and then blended to form a powdered simplicia. Simplicia was extracted using a maceration method with 70% alcohol at a ratio of 1:10. 19 The resulting macerate was filtered using Whatman No. 1 filter paper to obtain a liquid extract of the leaves. The filtrate was then concentrated using an IKA RV 10 digital V rotary evaporator at 55°C, 130 rpm, and 150 mbar to produce a viscous extract. The thick extract was then used for in vitro, in vivo, and in silico analysis.
In Vitro Tests: Antioxidant Activity, Steroid Content, TPC, and TFC
Antioxidant activity was assessed using the 1,1-diphenyl-2-picrylhydrazyl (DPPH) assay, as previously described. 20 Phytosteroid content was determined using the Liebermann–Burchard method and expressed as beta-sitosterol equivalents. 21 Total phenolic content (TPC) testing was performed using the Folin–Ciocalteu method. Total flavonoid content (TFC) testing was carried out using the aluminum chloride (AlCl3) colorimetric method. 22
In Vivo Test: Repair of Ovarian Damage
Experimental Design
An in vivo test was conducted by preparing a solution of P. angulata leaf extract and busulfan. P. angulata leaf extract was prepared by dissolving 20 g of thick P. angulata leaf extract in 200 mL of distilled water and adding 2 g of Sodium Carboxymethyl Cellulose (CMC Na) to form a suspension. The P. angulata solution was administered orally at a volume of 0.39–0.45 mL/rat/day depending on body weight (300 mg/kg body weight (BW)). The busulfan solution was prepared by dissolving 0.4 g of busulfan powder in 0.5 mL of 10% Dimethyl Sulfoxide (DMSO) and then diluting it with 4.5 mL of phosphate-buffered saline (PBS) at 37°C. The solution was then homogenized using a vortex mixer. Busulfan was administered intraperitoneally at a dose of 40 mg/kg BW and a volume of 0.065-0.075 mL/rat. 23 The test animals used were 25 female Sprague–Dawley white rats (Rattus novergicus), 5 weeks old, weighing 130–150 g. The test animals were acclimated for 2 weeks, and then, at 7 weeks of age, the female rats were divided into five groups, each consisting of five female rats. The sample size was determined using the Federer formula, (t – 1)(n – 1) ≥15, resulting in n = 5 rats per group. A post hoc power analysis conducted using SPSS 16 software confirmed that this sample size provided adequate statistical power (b > 0.80) at a significance level of a = 0.05. The experimental groups were as follows: Group K (negative control), B (busulfan), C (P. angulata L. extract), B>C (sequential busulfan and extract administration), and B+C (simultaneous busulfan and extract administration). Group K did not received any treatment during the study period. After 14 days of treatment, estrus cycle data were collected from the rats. After 28 days of treatment, the female rats were mated with male rats to collect data on successful pregnancy. Following pregnancy, rats were euthanized on the 19th day of pregnancy to collect ovarian histology data.
Estrous Cycle
Estrous cycle data were collected after 14 days of treatment using the vaginal smear method at 06:00 and 18:00 WIB time points. The preparations were stained with 3% Giemsa stain for 30 min and observed under a microscope. The observations recorded included proestrus, estrus, metestrus, and diestrus phases. 24
Pregnancy Rate
After 28 days of treatment, female rats were mated with male rats at a ratio of one male to two females to obtain pregnancy rate data. The presence of a vaginal plug and sperm in the vaginal smears indicated mating success. When sperm were found in the vaginal smear, the rats were declared pregnant on Day 0. 25 Pregnancy success was determined by measuring BW gain, reduction in remaining feed, mammary gland morphology, and behavioral observations. 26
Ovarian Histology
Ovarian organ collection was performed on rats that had successfully conceived and those that had not conceived. If the rats successfully conceived, organ collection was performed on the 19th day of pregnancy. If the rats failed to conceive after several successful matings, organ collection was performed on the 8th day after the last successful mating. Ovarian organ collection was performed using necropsy. Slides were prepared using the embedding method and hematoxylin and eosin staining. Ovarian histology was observed using a binocular microscope (Olympus CX31; Olympus Corporation, Japan). Initial descriptive observations and comparative histology between groups were performed at objective lens magnifications of 4× to ensure the observation of whole ovarian structures and 40× for detailed cell observation. 27
In Silico Test: Prediction of the Molecular Activity of P. angulata
Metabolite Profiling
The in silico test began by profiling the metabolites in a thick extract of P. angulata leaves. Profiling was performed using a Thermo Scientific Vanquish Liquid Chromatography High-Resolution Mass Spectrometry (LC-HRMS) instrument. 28 The metabolite results were validated using the PubChem database (http://pubchem.ncbi.nlm.nih.gov/).
Network Pharmacology
Network pharmacology (NP) is an initial systemic screening tool used to identify biologically relevant protein targets and their associated signaling pathways. NP was performed by searching for proteins in the body that can be targeted by P. angulata leaf metabolites using the PubChem website (https://pubchem.ncbi.nlm.nih.gov/) and the protein targets of busulfan pathogenesis and ovarian failure on the GeneCards website (https://www.genecards.org/), OMIM, and DisGeNet using the keywords “ovarian failure,” “alkylating agent,” “estrogen signaling pathway,” and “antioxidant.” The intersection of the two protein targets was determined using a Venn diagram tool (https://www.biotools.fr/misc/venny). Protein–protein interaction (PPIs) were identified using the STRING 12.0 website (https://string-db.org/) and ranked using Cytoscape 3.10.3. 29 The ranking results were analyzed using the Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway and Gene Ontology (GO) using the ShinyGO 0.85 website (https://bioinformatics.sdstate.edu/go/).
Molecular Docking
These core NP targets (hub genes) were further validated through molecular docking (MD) to determine the binding affinities and conformational stabilities of the lead metabolites. This dual-layered strategy, which combines NP and MD, ensures that the identified compounds possess both systemic relevance and high-affinity molecular interactions.
MD was performed on estrogen receptor 1 (ESR1) and estrogen receptor 2 (ESR2) proteins. The Protein Data Bank (PDB) codes used were 1X7R and 3ERT for ESR1 and 1X7J and 2FSZ for ESR2. Receptor selection was performed according to the specified criteria. 30 Receptor selection was based on structural validation using Ramachandran plot analysis from the PDBsum database (https://www.ebi.ac.uk/thornton-srv/databases/pdbsum/). The inclusion criterion was a threshold of ≥ 90% of residues located in the most favored regions. Based on these criteria, four candidate receptors were selected with the following validation scores: 1X7R (95.5%), 1X7J (93.7%), 2FSZ (91.9%), and 3ERT (91.2%). The receptor-native ligand complex was redocked. The native ligand redocked with a root mean square deviation (RMSD) value < 2 Å was used as a comparison ligand with the test ligand. The test ligand files were downloaded from PubChem. MD was performed using PyRx 0.8. Visualization was performed using PyMol 3.1.5 (3D) and Discovery Studio 2025 (2D) applications. 31
Molecular Dynamic
The receptor–ligand complex bond resulting from MD was further analyzed for stability in a cellular environment using molecular dynamics. In this study, simulations were performed on the P. angulata metabolite with the highest binding affinity and compared with those of the native ligand. Molecular dynamics simulations were performed using the YASARA Structure software (version 19.9.17) with a trajectory of 50 ns. MD simulations were performed on the best natural and native ligand. The AMBER14 force field (assisted model building with energy refinement) was used for the MD simulations. Some of the settings implemented included 0.9% NaCl content, 310 K temperature, and 10 Å cell extension. 32 To run the MD simulations, we used the md_run macro, while the evaluation of large simulation datasets was performed using the md_analyze and md_analyzeres macros to obtain the RMSD values of Cα atoms, root mean fluctuation (RMSF), and radius of gyration (Rg). 33 MD simulation data were generated after considering the equilibration conditions specified in the macros. MD simulation data were generated using CPU and GPU-based computing.
Statistical Analysis
The research data were analyzed quantitatively for in vitro test data, estrus cycles, and pregnancy success, and qualitatively for ovarian histology data and secondary metabolite profiling. Statistical analysis was performed using a one-way analysis of variance (ANOVA) test, followed by a Duncan post hoc test (p = .05), using SPSS ver. 22 software.
Results
In Vitro Test: Antioxidant Activity, Steroid Content, TPC, and TFC
Based on the test results, P. angulata leaf extract exhibited moderate antioxidant activity, with an IC50 value of 137.09 ± 1.2 ppm. The phytochemical profile of this extract is supported by phytosterol (1.63% w/w) and TPC (69.36 mgGAE/g), both of which fall into the moderate category. The TFC was low at 3.78 ± 0.01 mg/gEQ. These data indicate that although flavonoid components were found in smaller amounts than phenolics and phytosteroids, the extract retained its functional capacity to neutralize free radicals within that concentration range.
In Vivo Test: Repair of Ovarian Damage
The estrous cycle phases observed were proestrus, estrus, metestrus, and diestrus. The average duration of each estrous cycle and its constituent phases in each group is shown in Table 1. Group K showed a normal estrous cycle duration of 4.32 ± 0.33 days. In contrast, a single treatment in Group B significantly prolonged the cycle duration to 8.28 ± 3.92 days, nearly double that of Group K. The longest prolongation was recorded in Group B>C, with an average of 10.12 ± 3.46 days, surpassing groups K, B, and C. Conversely, Group C showed a cycle duration of 5.80 ± 1.31 days, which was longer than that of Group K but significantly shorter than that of Group B. Interestingly, Group B+C produced a duration of 6.19 ± 1.0 days, which was significantly shorter than that of groups B and B>C and approached the value of Group C.
Length of Estrous Cycle in Female Rats After Administration of P. angulata Leaf Extract and Busulfan Injection.
The diestrus and metestrus phases primarily influenced changes in cycle duration. The lengthening of cycle duration in groups B, B>C, and C was driven by a significant lengthening of the diestrus and metestrus phases. The longest diestrus duration was recorded in Group B>C (5.53 ± 5.32 days) and Group B (4.54 ± 4.28 days). Normalization of the diestrus phase was clearly observed in groups B+C (2.11 ± 0.80 days), which approached the normal duration of Group K (2.42 ± 0.33 days). The duration of the proestrus and metestrus phases also substantially increased in the busulfan-treated group. The longest metestrus duration was observed in Group B>C (2.85 ± 1.36 days) and Group C (2.72 ± 1.43 days), which was longer than that in Group K (0.53 ± 0.07 days). In contrast to the other phases, the estrus phase was shortened in all busulfan and P. angulata treatment groups (B, C, B>C, and B+C), with the shortest duration in Group B>C (0.40 ± 0.42 days), which was shorter than that in Group B (0.54 ± 0.09 days). Groups B+C showed a slightly longer estrus duration (0.60 ± 0.22 days) than the other busulfan treatment groups, although it was still shorter than that of Group K (0.82 ± 0.10 days). Treatment with P. angulata extract in groups B+C improved the duration of the estrous cycle in rats disrupted by busulfan, although the values did not fully return to normal levels, as seen in the control Group (K).
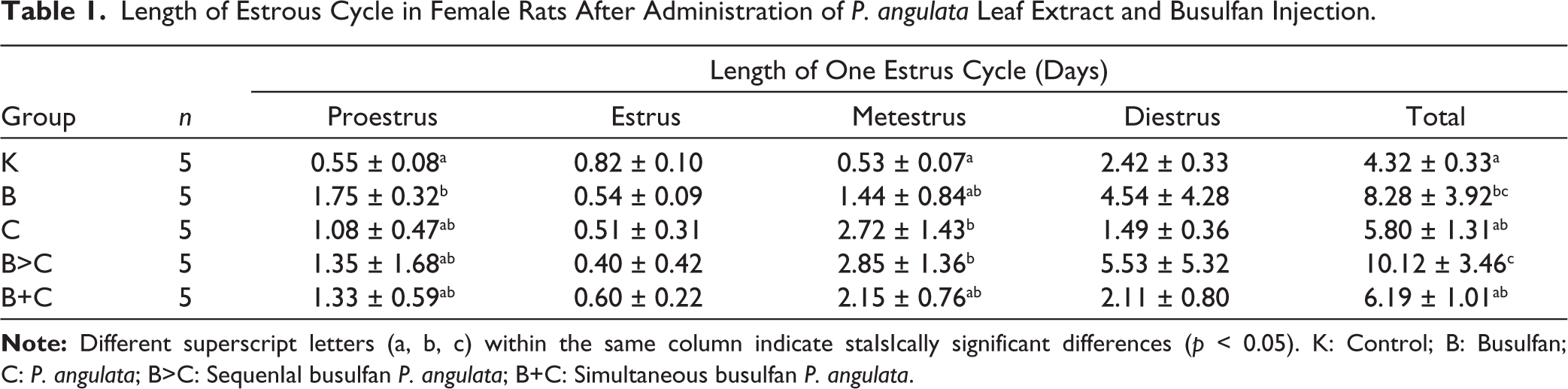
The pregnancy success rate in each group The pregnancy success rate for each group is shown in Figure 1. Group K, which had a typical estrous cycle duration (4.32 ± 0.33 days), achieved the highest pregnancy success rate (100%). Conversely, the increase in cycle duration due to busulfan (B) and C treatments was inversely related to pregnancy success. The single treatment groups B (cycle length 8.28 ± 3.92 days) and C (cycle length 5.80 ± 1.31 days) achieved only 60% pregnancy success in each Group. The most detrimental impact was observed in Group B>C, which had the most extended estrous cycle (10.12 ± 3.46 days) and the lowest pregnancy success rate of only 20%. Interestingly, simultaneous treatment in groups B+C resulted in a shorter cycle duration (6.19 ± 1.01 days) compared to groups B and B>C, accompanied by an increase in the pregnancy success rate to 40%. The pregnancy success rate in all treatment groups (B, C, B+C, and B>C) was lower than that in the control Group (K), indicating a negative correlation between estrus cycle duration and the likelihood of pregnancy.
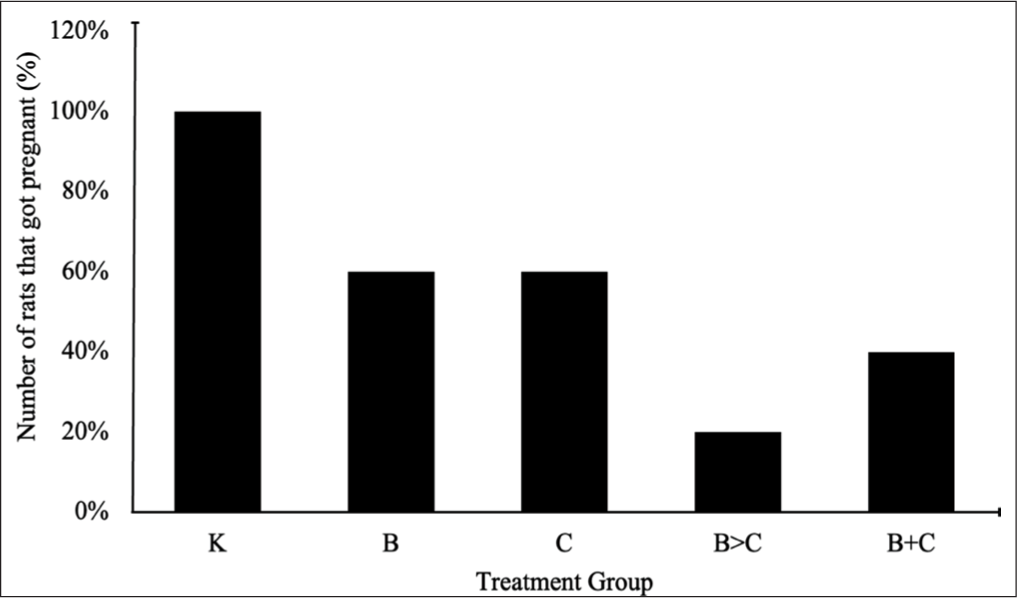
Ovarian histology provides strong structural support for the data reported on estrous cycle dysfunction and pregnancy success (Figure 2). Group K exhibited a standard ovarian architecture characterized by the presence of all stages of folliculogenesis (from primordial to antral follicles) and numerous corpora lutea. The absence of damage to the granulosa and stromal cells supported a normal estrous cycle (4.32 ± 0.33 days) and a 100% successful pregnancy rate.
Photomicrograph of Ovaries Stained with Hematoxylin and Eosin. Bar = 200 µm. (a) Group K, (b) Group B, (c) Group C, (d) Group B>C, and (e) Group B+C.
Busulfan treatment caused structural damage that was directly proportional to the prolongation of the estrous cycle and a decrease in the pregnancy rate. Group B exhibited severe damage to the underlying ovarian follicles. Secondary follicles degenerated, often leaving only empty circles in the stroma. Preantral and antral follicles displayed clear intercellular cavities and cracks in the granulosa cell layers. In Group C, the ovaries showed mild hemorrhage and vascular congestion in the medulla and around smaller follicles, as well as in intercellular granulosa cavities. Groups B>C showed a combination and amplification of the damage from groups B and C, characterized by reduced stromal density (widened intercellular spaces), intensive follicular degeneration, and mild hemorrhage. Some granulosa cells in the preantral follicles exhibited structural damage characterized by disintegrated fragments and significantly widened intercellular spaces.
Groups B+C showed significant general architectural improvement that superficially resembled that of Group K. The observed defects were a combination of those in groups B and C, but at a much lower level. Specifically, stromal damage, granulosa cell damage, inflammation, and hemorrhage were significantly reduced. This near-normal histology supported a significantly shorter estrous cycle duration (6.19 ± 1.01 days) compared to groups B and B>C. Qualitative ovarian histology data indicated that the most severe structural damage occurred in groups B>C and B, whereas Group B+C showed significant improvement in tissue architecture, approaching normal conditions (Group K).
In Silico Test: Prediction of the Molecular Activity of P. angulata
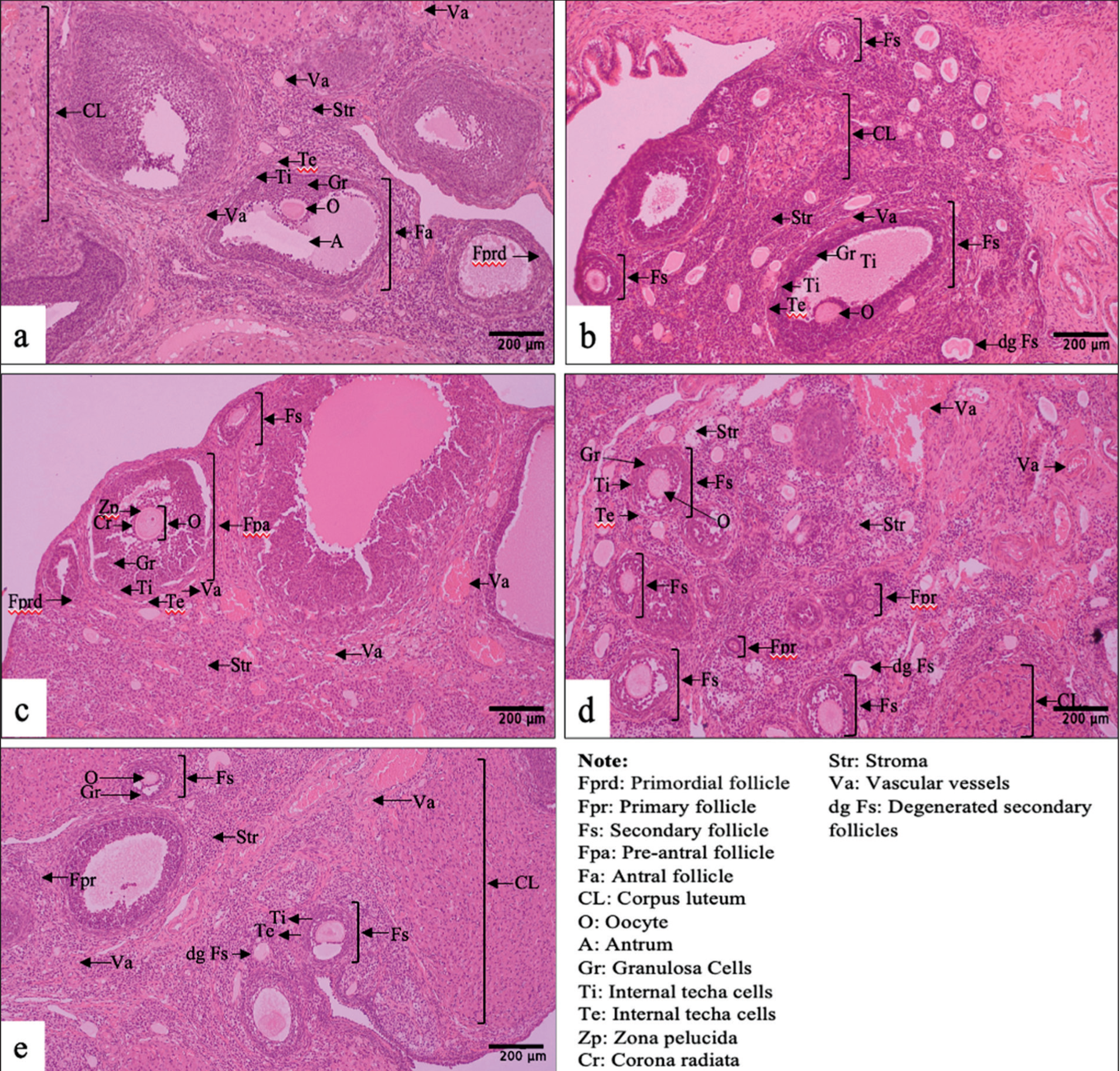
In profiling, a chromatogram was obtained, showing the highest peak intensity at retention times (RT) of 1.21, 1.45, 1.09, 6.26, 8.50, 27.01, 11.59, and 27.26 min (Figure 3). This graph identified 100 metabolites; after validation, 77 metabolites were confirmed (Table 2). The identified metabolites belong to several groups that have the potential to support pharmacological activity in the treatment of ovarian failure. These metabolites include seven phenolic metabolites: chlorogenic acid, salicylic acid, fumaric acid, 1,3,4,5-Tetrahydroxycyclohexanecarboxylic acid, 5-[(3-carboxy-4-hydroxyphenyl) carbamoylamino]-2-hydroxybenzoic acid, (E)-ferulic acid, and (2E)-3-(3,4-Dimethoxyphenyl) acrylic acid; three flavonoids: Rutin, isoquercetin, and Albanol A; and one secosteroid, Physalin F.
Chromatogram of LC-HRMS Analysis of Ethanolic Extract of P. angulata Leaves with a Wavelength of m/z 100–1500 and a Retention Time of 0–33 min. The Chromatogram Shows the Highest Peaks at Retention Times of 1.21, 1.45, 1.09, 6.26, 8.50, 27.01, 11.59, and 27.26 min.
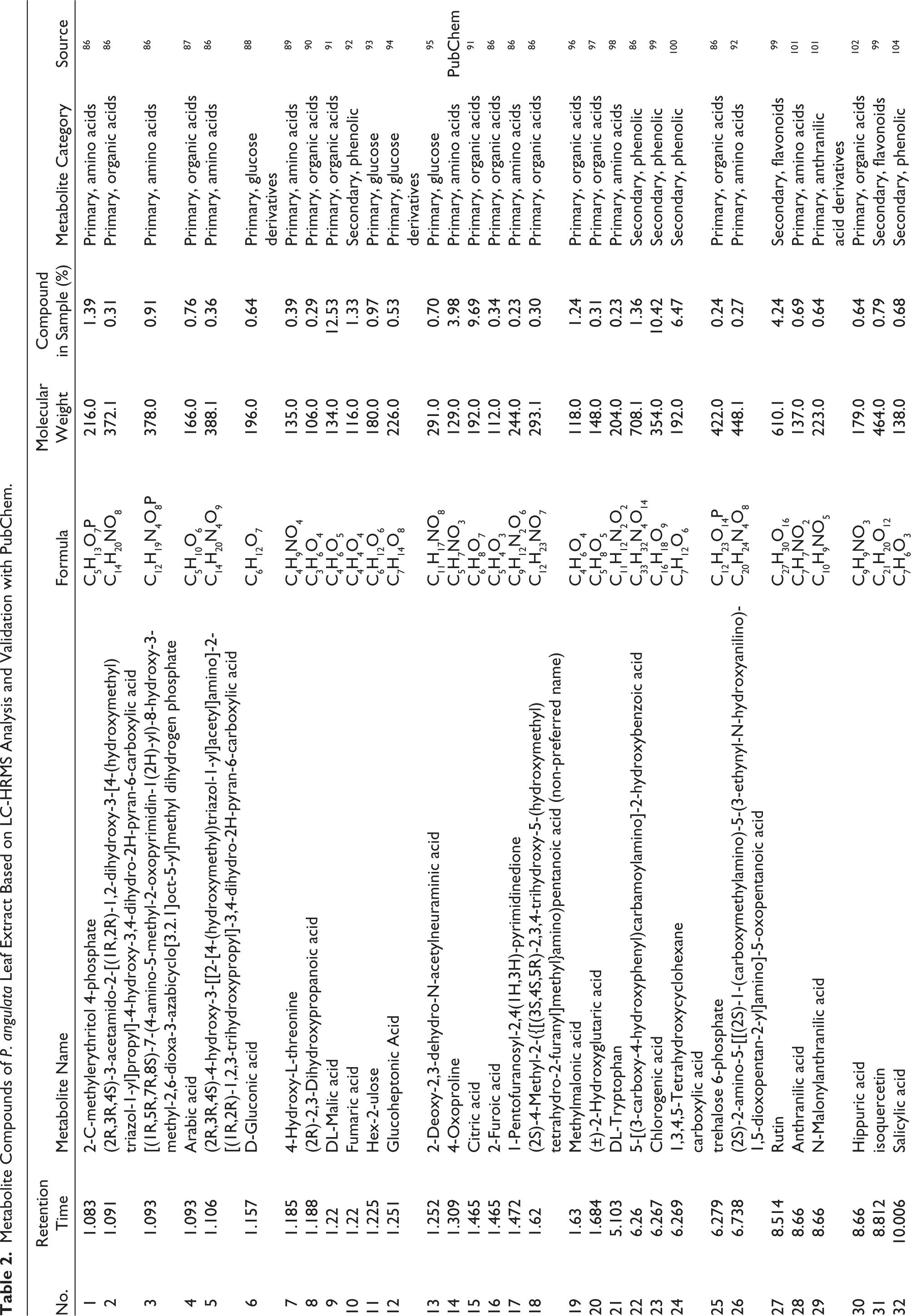
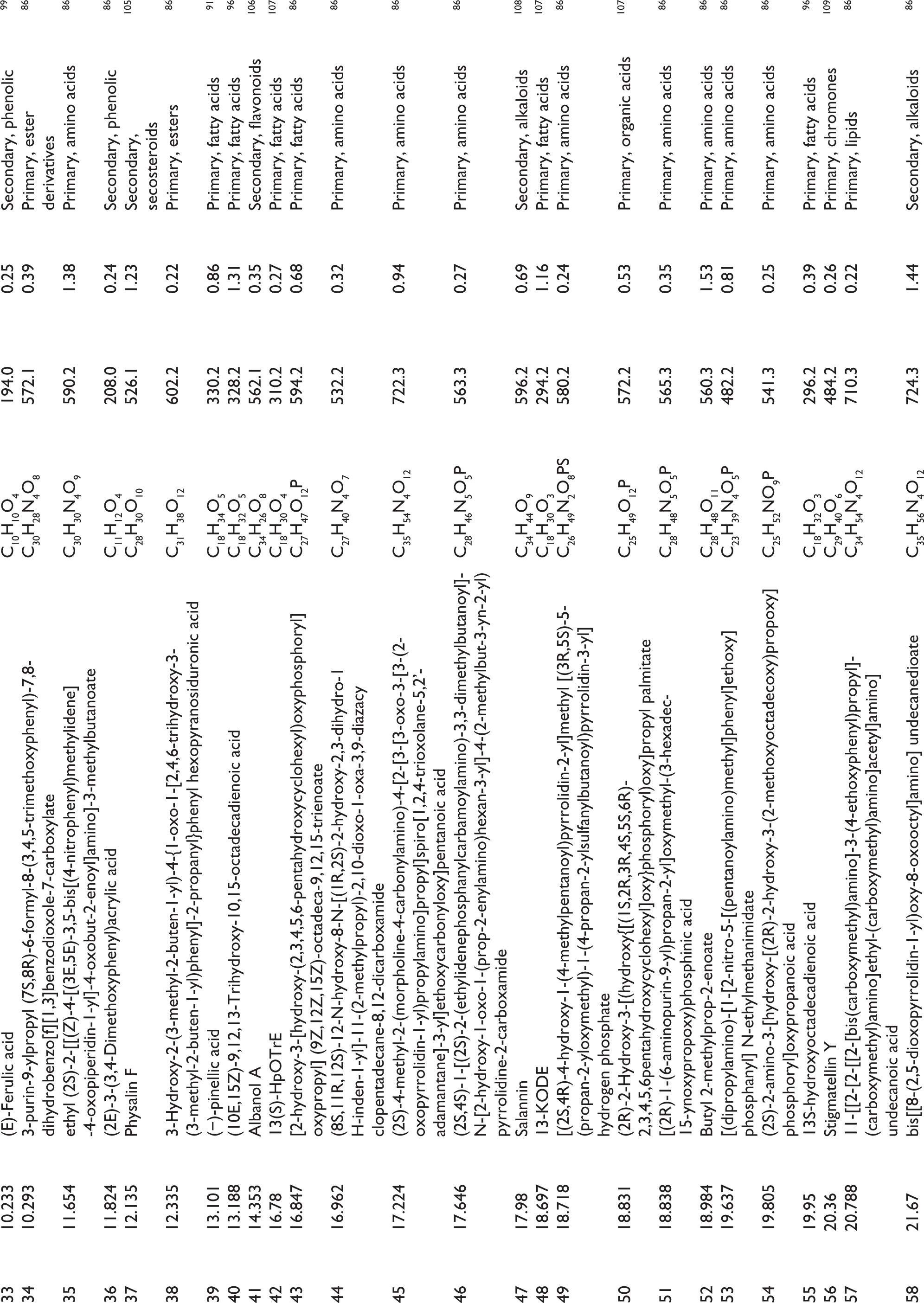
Metabolite Compounds of P. angulata Leaf Extract Based on LC-HRMS Analysis and Validation with PubChem.

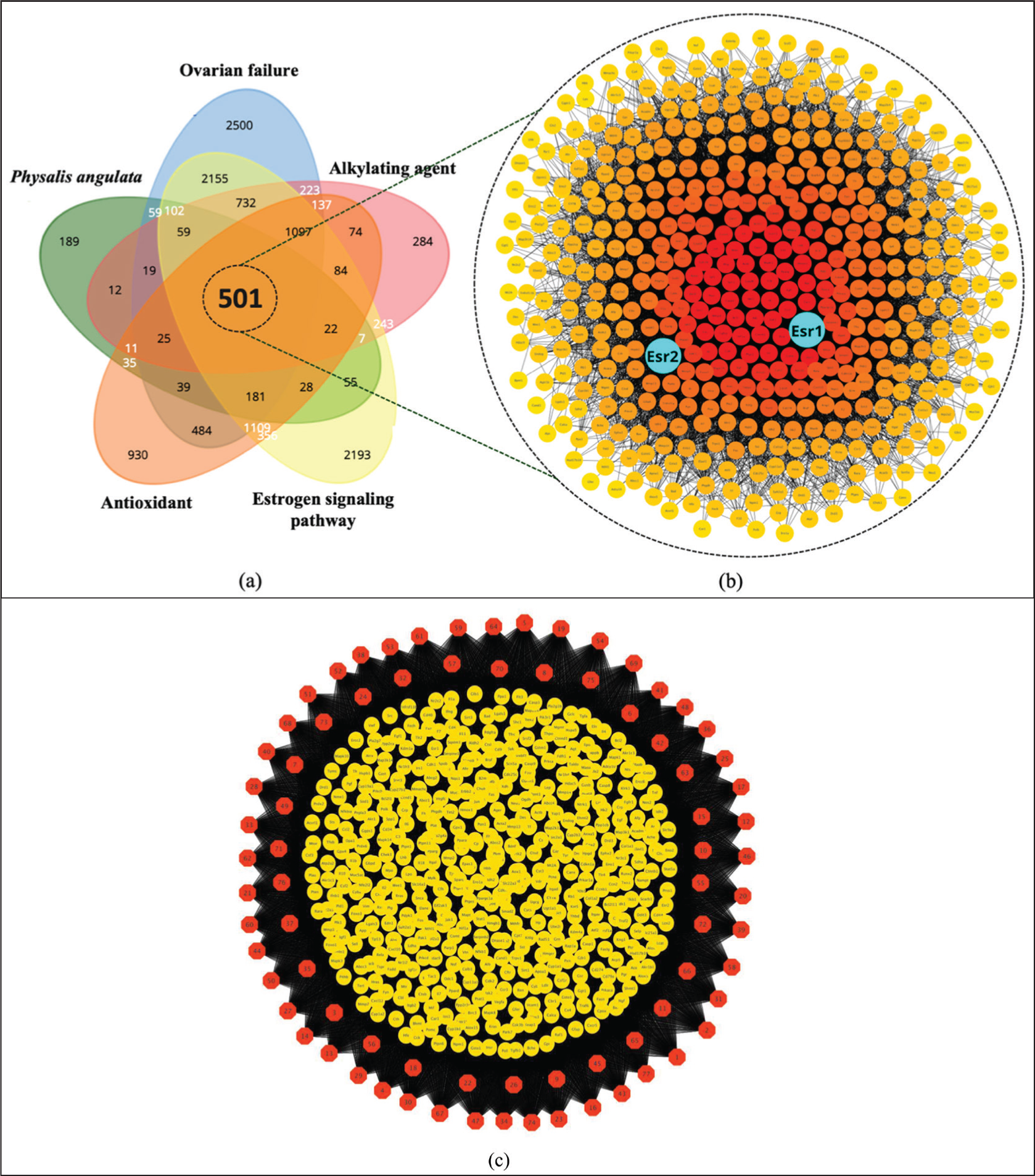
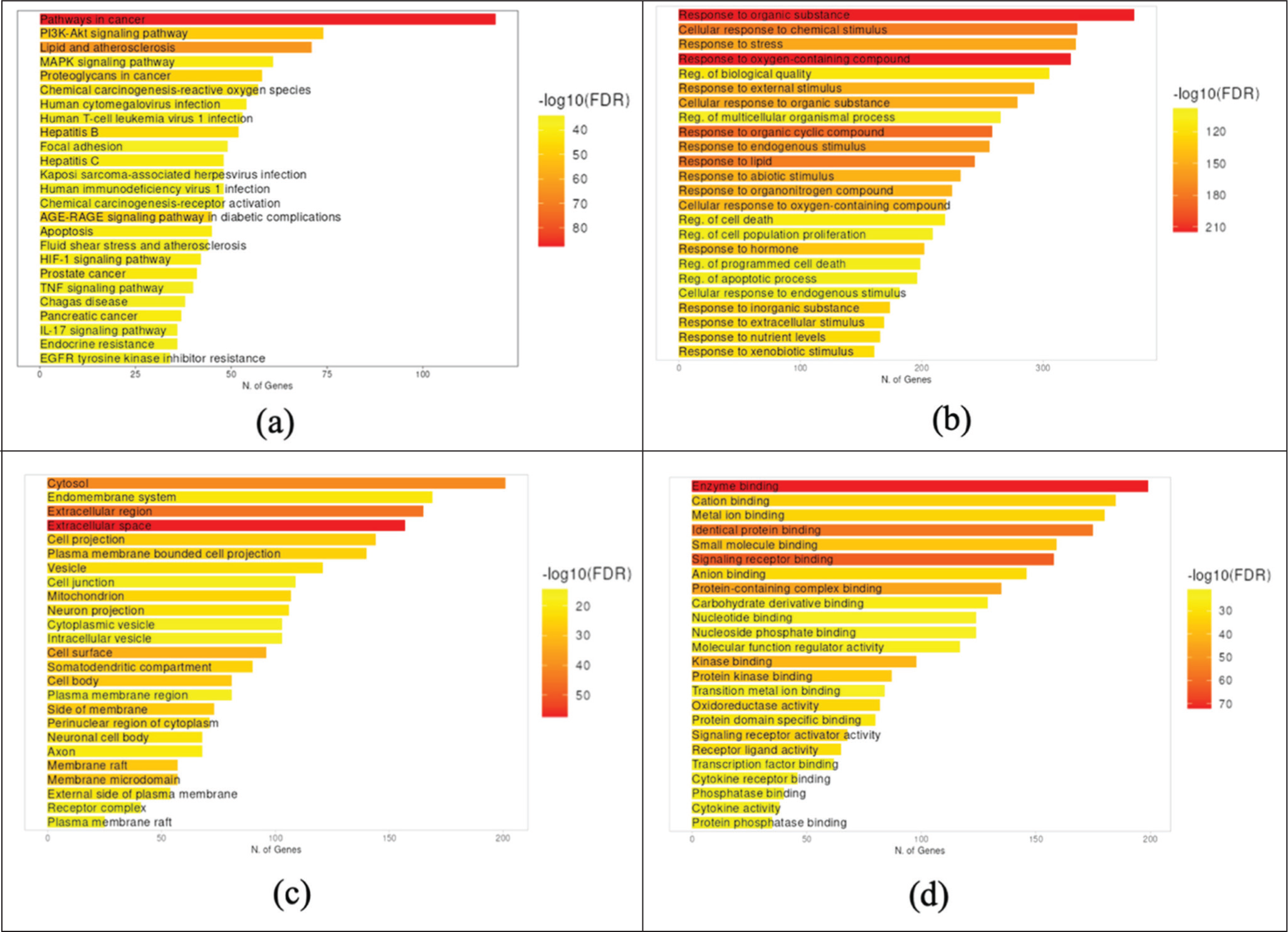
NP began by predicting protein targets from 77 metabolites, resulting in 1,344 protein targets. These targets were then intersected with four protein groups related to the pathogenesis of busulfan and ovarian failure: ovarian failure (9,208 proteins), alkylating agents (3,530 proteins), estrogen signaling pathway (8,924 proteins), and antioxidants (5,113 proteins). The intersection of the five protein target groups yielded 501 proteins (Figure 4a). Protein ranking showed that estrogen receptors (ESR1 and ESR2) were among the central target proteins, with ESR1 ranked 26th and ESR2 ranked 120th (Figure 4b), and bond compound-protein interactions (Figure 4c). KEGG pathway analysis predicted that P. angulata (Ciplukan) could regulate hormones through the endocrine resistance pathway, ranking 24th (Figure 5a). The predicted biological role of P. angulata leaf extract is its response to chemicals and oxidative stress (Figure 5b). At the cellular level, it maintained the extracellular environment of the cell (Figure 5c), and at the molecular level, it was capable of binding to signaling receptors (Figure 5d). These results indicate that P. angulata extract modulates hormones and the oxidative stress response at the molecular level, with ESR1 as the primary target, exhibiting a stronger binding affinity than ESR2.
(a) Slices of Protein Targets from the Five Protein Groups: P. angulata, Ovarian Failure, Alkylating Agent, Estrogen Signaling Pathway, and Antioxidant, Yielding 501 Proteins; (b) Protein–Protein Interactions and Rankings of the 501 Proteins (Blue Indicates ESR1 and ESR2) Show a Relatively High Ranking of ESR1, at 26th, and ESR2, at 120th; (c) Compound–Protein Bond, Red Hexagon = Compound, Yellow Circle = Protein.
Pharmacological Effects of P. angulata Leaf Extract on Ovarian Failure. (a) KEGG Pathway; (b) GO Biological Process; (c) GO Cellular Component; and (d) GO Molecular Function.
MD successfully evaluated the binding affinity energies of 77 metabolites to four estrogen receptor structures (ESR1 and ESR2); the more negative the affinity energy, the stronger the binding (Table 3). In the IX7R receptor, Albanol A exhibited an affinity energy of –10.0 kcal/mol, which was more negative than that of the native ligand Genistein (–9.5 kcal/mol). In 3ERT, three metabolites showed a stronger affinity than the native ligand 4-hydroxytamoxifen (–7.9 kcal/mol): Physalin F (–10.3 kcal/mol), Albanol A (–9.9 kcal/mol), and Rutin (–8.1 kcal/mol). In 2FSZ, Albanol A had an affinity energy of –9.9 kcal/mol, which was more negative than that of the native ligand 4-Hydroxytamoxifen (–8.5 kcal/mol). Furthermore, in 1X7J, no tested metabolite exhibited a more negative affinity energy than the native ligand Genistein (–8.5 kcal/mol). These results identify Albanol A, Physalin F, and Rutin as prime candidates with the potential to modulate the estrogen signaling pathway via ESR1 and ESR2.
Results of Molecular Docking of P. angulata Leaf Secondary Metabolites to ESR1 and ESR2.
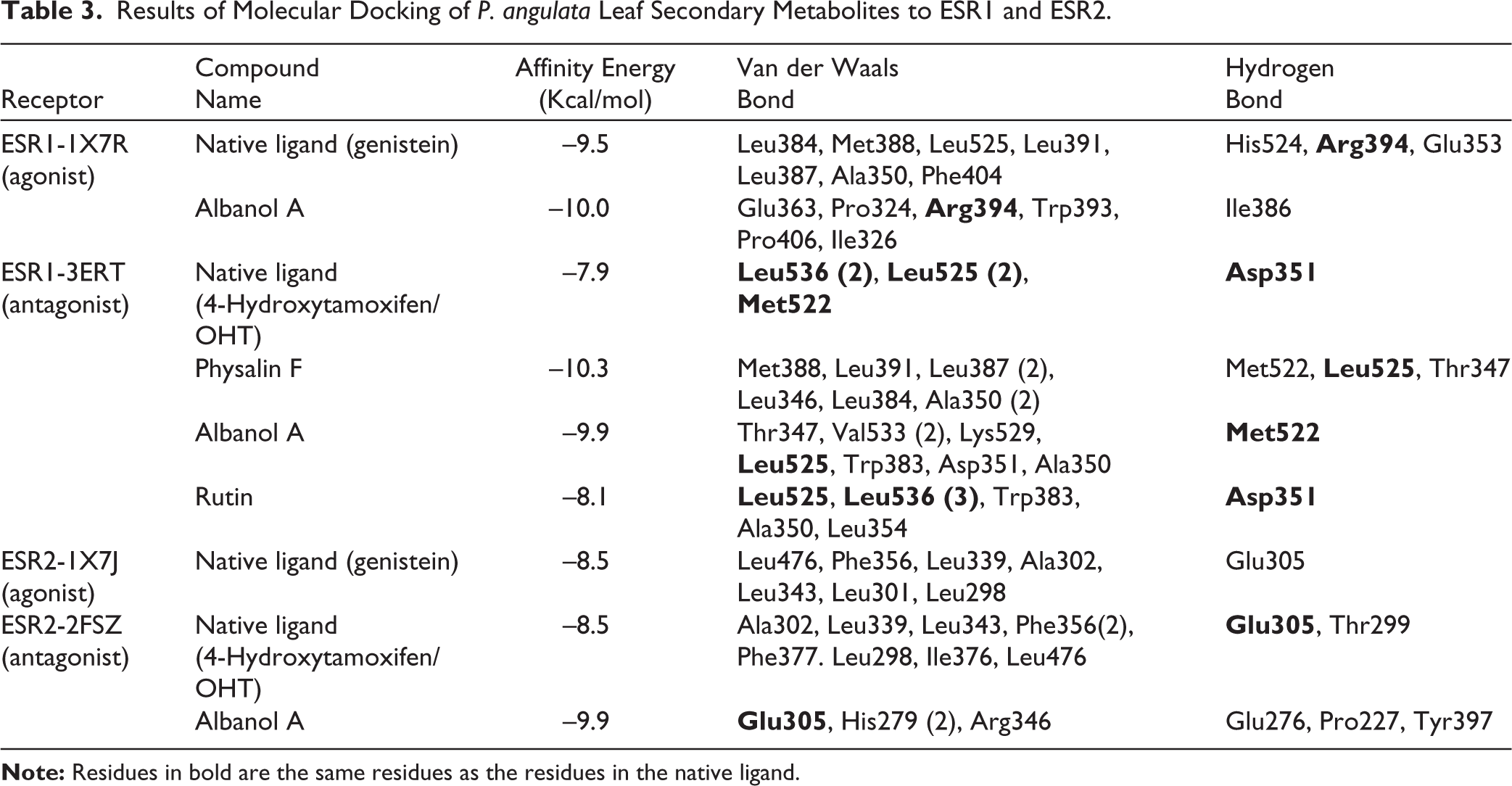
The strength of receptor–ligand binding is supported by residue interactions between the metabolites and the active site of the receptor. These interactions confirm the potential biological activities of these metabolites (Table 3 and Figure 6). The 1X7R–Albanol A complex binds to Arg394, which also interacts with the 1X7R–Genistein complex. The other binding residues included Leu384, Met388, Leu525, Leu391, Leu387, Ala350, Phe404, His524, and Glu353. The 3ERT–Physalin F complex shares the residue Leu525 with the 3ERT–native ligand 4-Hydroxytamoxifen complex. The other residues included Met388, Leu391, Leu387 (2), Leu346, Leu384, Ala350 (2), Met522, and Thr347. The 3ERT–Albanol A complex shares two residues (Leu525 and Met522) with the native ligand 4-Hydroxytamoxifen. The 3ERT–Rutin complex shares three residues (Leu525, Leu536, and Asp351) with the native ligand 4-Hydroxytamoxifen. The 2FSZ–Albanol A complex shares the residue Glu305 with the native ligand 4-Hydroxytamoxifen. However, the 2FSZ ligand redocking of the native ligand does not share the same residue as; 34 the 2FSZ–Albanol A complex binds to residues Glu305, His279 (2), Arg346, Glu276, Pro227, and Tyr397, supporting the potential interaction of the test ligands with the target protein. MD results confirmed that Albanol A, Physalin F, and Rutin were the primary candidate compounds with stronger binding affinities than natural ligands, exhibiting consistent amino acid residue interaction patterns at the active sites of ESR1 and ESR2 receptors.
2D and 3D Visualizations of the Molecular Binding of Secondary Metabolites from P. angulata Leaves to the ESR1 and ESR2 Receptors. (a) Albanol A–1X7R; (b) Physalin F–3ERT, (c) Albanol A–3ERT, (d) Rutin–3ERT, and (e) Albanol A–2FSZ.
NP and MD analyses revealed that P. angulata leaf extract had a greater effect on the ESR-1 than on ESR-2. The highest ESR-1 agonist affinity energy was observed for Albanol A, and the highest ESR-1 antagonist affinity energy was observed for Physalin F. Therefore, molecular dynamics analysis was performed on four receptor–ligand complexes derived from the MD results: Albanol A–1X7R complex compared with the native ligand (Genistein)–1X7R, and Physalin F–3ERT complex compared with the native ligand (4-hydroxytamoxifen/OHT)–3ERT.
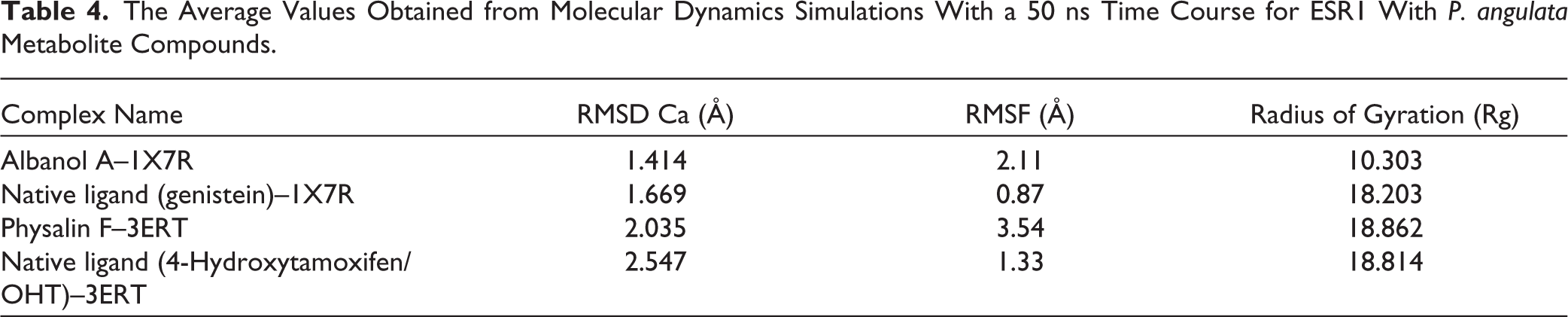
In this analysis, the parameters considered were RMSD, RMSF, Radius of Gyration (Rg), and hydrogen-bond interaction parameters, as summarized in Table 4. Based on the RMSD Ca data (Figure 7), all complexes displayed stable equilibration profiles with average values ranging from 1.414 to 2.547 Å. The Albanol A–1X7R complex recorded the lowest RMSD value (1.414 Å), indicating the highest protein backbone stability compared to the other systems, including the positive control Genistein (1.669 Å). In the antagonist conformation group, the Physalin F–3ERT complex (2.035 Å) was more stable than the 4-Hydroxytamoxifen control (2.547 Å).
The Average Values Obtained from Molecular Dynamics Simulations With a 50 ns Time Course for ESR1 With P. angulata Metabolite Compounds.
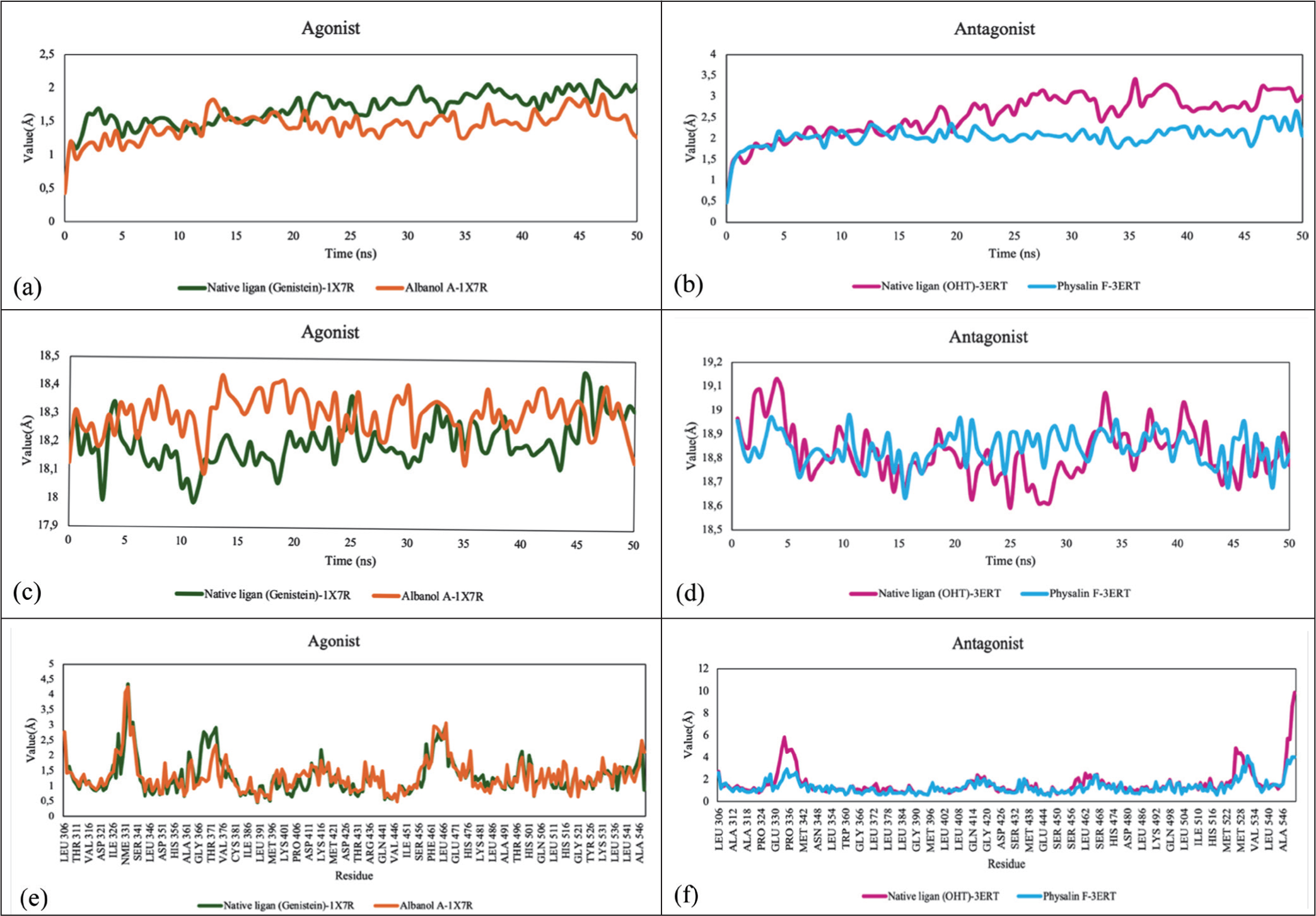
Evaluation of ligand flexibility within the binding pocket (Figure 6) showed significant variation, with Genistein having the lowest RMSF value (0.87 Å), reflecting rigid binding. Albanol A showed moderate flexibility (2.11 Å), whereas Physalin F exhibited the highest fluctuations (3.54 Å), indicating greater conformational dynamics in the antagonist state. Rg parameter analysis (Figure 7) revealed that the Albanol A–1X7R complex had the highest structural compactness with the smallest value (10.303 Å) compared to other complexes, which were in the range of 18.203–18.862 Å. These findings were supported by solute hydrogen-bond analysis, which showed consistent interactions across all systems. The Physalin F–3ERT complex exhibited the highest average number of hydrogen bonds (196.026), followed by Genistein (193.941) and Albanol A (193.931). The high hydrogen-bond density in Physalin F is suspected to be a key factor in maintaining the binding affinity despite its high fluctuations (RMSF). Molecular dynamics analysis shows that the Albanol A–1X7R complex exhibits the highest protein backbone stability and structural compactness (lowest RMSD and Rg), while Physalin F–3ERT maintains its binding affinity through the highest average hydrogen-bond density among all systems.
Molecular Dynamics Results for the ESR1—P. angulata Metabolite Complex. (a) RMSD Cα (Agonist); (b) RMSD Cα (Antagonist); (c) Radius of Gyration (Agonist); (d) Radius of Gyration (Antagonist); (e) RMSF (Agonist); and (f) RMSF (Antagonist).
Discussion
The test results showed that the P. angulata leaf extract had moderate antioxidant activity, with an IC50 of 137.09 ± 1.2 ppm, which fell within the moderate category (101–150 ppm). 35 This finding is in line with previous studies that reported similar IC50 values for the ethanol extract of P. angulata leaves, namely 190 ± 4.7 µg/mL 36 and 110.5 ± 3.2 ppm. 37 The phytosteroid content in P. angulata extract was found to be 1.63% (w/w), and falls within the reported range for several Mediterranean region plants, namely 0.1%–5.7%. 34 The test showed a phytosteroid content of 1.63% w/w, which falls within the moderate category (0.55%–2.0% w/w). 38 The phytosteroid, phenolic, and flavonoid contents of the extracts support this antioxidant activity. The TPC and TFC content were 69.36 mg GAE/g and 3.78 ± 0.01 mg/g EQ, respectively. In contrast to the classification used in other studies by Amelia et al. 39 and Chandra et al., 40 TPC was categorized as moderate, and TFC was categorized as low. Although the TPC and TFC were lower than those reported by Pillai et al. 41 (140.65 ± 3.8 mgGAE/g and 370.64 ± 4.33 mg/g EQ, respectively), the resulting antioxidant activity did not depend solely on the total quantity but also on the type and structure of specific metabolites.
Analysis of the estrous cycle data (Table 1) provided clear functional evidence of intervention-induced reproductive dysfunction and the relative effectiveness of the P. angulata extract in alleviating this dysfunction. Group K maintained a normal estrous cycle (4.32 ± 0.33 days). In contrast, Group B experienced a twofold prolongation of estrous cycle duration (8.28 ± 3.92 days). This prolongation was supported by histological findings of granulosa cell damage, which inhibited estrogen biosynthesis and delayed the negative feedback mechanisms in the hypothalamus. Functionally, this dysfunction manifested as a substantial prolongation of the diestrous phase and a shortening of the estrous phase (0.54 ± 0.09 days), indicating a failure of ovulation.
Group C showed moderate prolongation of the estrous cycle (5.80 ± 1.31 days). This functional impairment is thought to be mediated by the phytosteroid group identified in leaf extracts. Due to their structural similarity to steroid hormones, phytosteroids have the potential to influence the estrogen signaling pathway, ultimately prolonging the metestrus phase (2.72 ± 1.43 days). However, the anti-inflammatory activity of the extract successfully mitigated this impairment, preventing severe prolongation of diestrus. The most severe dysfunction occurred in Group B>C, with the longest estrus cycle (10.12 ± 3.46 days), indicating cumulative damage from busulfan exacerbated by the phytosteroid effect on the initial oxidative stress conditions. 37 This complete failure of mitigation was reflected by the maximal prolongation of the diestrus phase (5.53 ± 5.32 days).
Simultaneous treatment with B+C resulted in a clear mitigating effect, with a relatively shorter cycle duration (6.19 ± 1.0 days) compared to groups B and B>C. Co-administration is thought to allow potent antioxidant compounds (flavonoids and phenolics) in the extract to alleviate busulfan-induced oxidative stress and stabilize estrogen production. 42 Functionally, this resulted in a lengthening of the estrus phase (0.60 ± 0.22 days) and a significant shortening of the diestrus phase (2.11 ± 0.80 days); however, the metestrus transition still exhibited a slight delay compared with that of the control. This functional improvement was supported by histological findings showing the restoration of ovarian architecture resembling that of the control group.
The pregnancy success rates in female rats (Figure 1) were directly correlated with the observed hormonal stability and the duration of the estrous cycle (Table 1). Group K achieved the highest success rate (100%) owing to hormonal homeostasis, which ensures timely progesterone transition, an essential condition for endometrial receptivity and embryo implantation. 43 In contrast, single treatment in groups B and C limited pregnancy success to 60%. This decrease is functionally related to the prolongation of estrous cycle duration resulting from the inhibition of the progesterone hormonal transition. 44 This delay forces the blastocyst to enter a dormant phase; if progesterone levels are inadequate within a certain time frame, implantation will fail, and embryo degeneration will occur. 45 The worst impact was observed in the B>C group, which had the longest estrous cycle duration (10.12 ± 3.46 days) and the lowest pregnancy success rate (20%). This severe structural damage caused by busulfan was exacerbated by the phytosteroid components in the extract, which potentially affect the estrogen signaling pathway, resulting in an extreme delay in the hormonal transition. In contrast, the simultaneous treatment Group (B+C) showed increased mitigation, with a pregnancy success rate of 40%, which was 20% higher than that of the B>C group. This improvement demonstrates that the antioxidant activity of the extract, when administered concurrently, can repair granulosa cell damage and stabilize estrogen production, although it remains lower than that in the B, C, and K groups.
Pregnancy failure in the busulfan-induced model results from a complex interaction between decreased oocyte competence and disruption of maternal-fetal synchronization. Busulfan induction causes oocyte DNA damage via oxidative stress and alkylation mechanisms. Such genomic damage significantly diminishes oocyte quality, which can lead to fertilization failure in the early stages. Even if the affected oocytes are capable of ovulation, inferior cytoplasmic quality and genetic integrity hinder successful sperm penetration and fusion. 46 If fertilization occurs, genomic damage inhibits embryo development during the pre-implantation stage. Mutations in critical genes that regulate early metabolism and cell division lead to the loss of blastocyst viability. 47 Although P. angulata extract administration is hypothesized to improve endometrial receptivity through hormonal modulation, implantation will not occur if the blastocyst is genetically incompetent or undergoes early embryonic death. Successful full-term pregnancy depends on the synergy between oocyte quality (supporting fertilization), genetic viability of the blastocyst, and precise timing of the optimal endometrial window of receptivity. 48 The decreased pregnancy rate in the B+C group compared to the K, B, and C groups represents a multifactorial failure, originating from the pre-fertilization phase to synchronization failure during the implantation phase.
Histopathological examination of the ovaries confirmed the functional and hormonal dysfunctions observed in the estrous cycle and pregnancy success data (Figure 2). Group K exhibited normal ovarian morphology, characterized by folliculogenesis and a healthy corpus luteum, consistent with normal ovarian function. In contrast, Group B exhibited secondary follicle destruction and granulosa intercellular spaces. This structural damage fundamentally impedes paracrine communication between follicle cells, which is essential for estrogen biosynthesis. This led to nearly twofold prolongation of the estrous cycle duration and induced follicular atresia. 49 Group C exhibited vascular congestion and intercellular spaces, indicating that a single administration of the extract caused mild inflammatory effects and impaired granulosa cell proliferation. The most severe structural damage occurred in Group B>C (sequential), characterized by severe stromal damage and massive follicular degeneration. This cumulative damage was exacerbated by phytosteroids in the extract, which potentially affect hormonal pathways, culminating in the most severe functional dysfunction, consistent with the most extended estrous cycle duration.
In contrast, treatment with Group B+C (simultaneous) showed significant general morphological improvement, resembling the control group, although residual vascular congestion was still observed. 49 This structural improvement provides strong evidence that the simultaneous administration of P. angulata effectively mitigates busulfan-induced ovarian damage. Mitigation is achieved through antioxidant activity, which reduces oxidative stress and triggers cellular damage, and phytosteroid activity, which modulates hormonal signaling pathways. This structural improvement supports partial functional recovery, as reflected by a shorter estrous cycle duration (6.19 ± 1.0 days) compared to that observed with toxic and sequential treatments.
Profiling identified 77 metabolites, which were categorized into major groups, including phenolics, flavonoids, and secosteroids. The number of metabolites identified in this study differed from those in previous reports, which identified 43 metabolites, including 13 withanolides, 37 and 75 metabolites, including 47 withanolide derivatives. 50 This difference is thought to be caused by geographical variations, temperature, altitude of the harvesting location, and the solvent type. 51 Although the number of withanolide derivatives was lower, the presence of more phenolics than flavonoids in this extract is consistent with a previous report. 52 The composition of these secondary metabolites greatly determines the quality and internal functions of herbal medicines. 53
The metabolite profile results directly support the biological findings and serve as the basis for in silico analyses. Based on this profiling, P. angulata is suspected to have two protective roles against ovarian failure: antioxidant and phytosteroid roles. Rutin, the main flavonoid in other Physalis species, 54 plays an antioxidant role, and the phenolic compound chlorogenic acid supports the moderate antioxidant activity of the extract (IC50 137.09 ppm). This compound is hypothesized to play a role in alleviating busulfan-induced oxidative stress, as evidenced by the recovery of the estrus cycle and increased pregnancy success in the simultaneous treatment Group (B+C). The role of phytosteroids is carried out by the content of Physalin F; this group plays a role in regulating inflammatory responses and hormonal homeostasis. 55 Secosteroids are metabolized by CYP11A1 into active metabolites that effectively reduce inflammatory cytokine levels and exert corticosteroid effects.56,57 In addition, Physalin F has been reported to have immunosuppressive effects and can inhibit the production of key inflammatory transcription factors.58,59 This potent anti-inflammatory effect of Physalin F explains the moderate cycle prolongation and reduced vascular congestion observed in ovarian histology following P. angulata intervention. Although the anti-inflammatory potential of Physalin F has been tested, its role in ovarian failure and its interaction with estrogen receptors have not yet been reported. Therefore, P. angulata metabolites were further explored using NP and MD analyses to predict the specific molecular targets of action in estrogen receptor (ESR) modulation.
NP analysis predicted that the metabolites of P. angulata leaves have the potential to more strongly affect the protein encoded by ESR1 than ESR2, as indicated by the higher ranking of ESR1 over ESR2 in the PPIs analysis. These two genes encode ESRα and ESRβ proteins, which function as the main binding sites for estradiol during folliculogenesis and ovulation. 60
In the ovary, ESRα is expressed in interstitial cells, theca cells, and granulosa cells of developing antral follicles, whereas ESRβ is expressed only in granulosa cells. 61 ESRα supports follicle proliferation and survival, whereas ESRβ acts as an anti-proliferative modulator or “brake.” 62 In the pre-estrus and proestrus phases, the ratio of ESRα begins to increase over ESRβ, marking a dominant follicle ready for final maturation and ovulation. 63 Therefore, the ability of metabolites to modulate ESRα is highly relevant because busulfan-induced ovarian failure is primarily due to the failure of follicle development. Although ESR is the target, the binding affinity of metabolites to ESR is not expected to be very strong because the main steroid group found in the extract is secosteroids. 64 Secosteroids have a specific Vitamin D Receptor (VDR). However, because their basic structure (rings A–D with a break in ring B) is similar to that of steroids, binding is possible, although with lower affinity than that of endogenous steroids. This interaction is important for modulation.
NP analysis revealed that the mechanism of action of P. angulata leaf metabolites is multifactorial, with the majority playing a role in cancer pathways and a minority playing a role in hormonal pathways through endocrine resistance. GO analysis predicted that the extract responds to ovarian damage caused by alkylating agents through oxidative stress response and chemical stress (GO: biological function) (Figure 4). This role involves balancing extracellular oxidants and antioxidants 65 through the direct scavenging of Reactive Oxygen Species (ROS), inhibition of ROS formation, and upregulation of antioxidant enzymes. 66 This antioxidant role is mainly modulated by phenolic and flavonoid metabolites. 67 In addition, metabolites target estrogen receptor-binding signaling (GO: molecular function) and maintain extracellular components of cells (GO: cellular component). P. angulata metabolites (phenolics, flavonoids, and secosteroids) are associated with improvements in in vivo studies. Antioxidants respond to oxidative stress (explaining granulosa cell repair and shorter diestrus), whereas estrogen receptor (phytosteroid) signaling mediates the modulation of folliculogenesis (explaining the normalization of estrus and increased pregnancy success).
The MD results support the hypothesis that P. angulata metabolites mitigate ovarian damage by modulating estrogen receptors (ESRs). The analysis showed a stronger metabolite-binding affinity for ESR1 (ESRα) than for ESR2 (ESRβ), consistent with the protein ranking findings in NP. ESRα is a key mediator of granulosa cell proliferation and follicle development. 62
Selective antagonism of ESRα (ESR1) inhibits growth signaling pathways and suppresses granulosa cell proliferation. 68 Although ESRα inhibition normally impairs folliculogenesis, in the context of alkylating agent toxicity, this antagonism is interpreted as a protective mechanism against ovarian failure. Busulfan toxicity results in the premature depletion of the follicular reserve, as follicles are forced to rapidly develop into atretic follicles. 69 By suppressing the uncontrolled rate of follicular development and delaying granulosa cell proliferation, 70 this metabolite minimizes cellular damage and preserves the ovarian reproductive potential.
The strongest antagonistic binding to ESR1 is mediated by Physalin F (–10.3 kcal/mol), exceeding that of the native ligand 4-Hydroxytamoxifen (–7.9 kcal/mol). This binding strength is attributed to the presence of more hydrogen bonds in the 3ERT–Physalin F complex than in the native ligand complex. 64 As a seco-steroid, Physalin F is thought to exhibit selective antiestrogenic effects, which aligns with literature findings that withanolides have anti-proliferative and antiestrogenic roles.71,72 These antiestrogenic and pro-apoptotic properties have the potential to disrupt the estrogen signaling pathway, contributing to ovarian failure and inflammation.
Physalin F’s activity is biphasic, acting as a selective estrogen receptor modulator (SERM) that depends on endogenous estrogen levels. 73 When endogenous estrogen levels are high, as may occur during proestrus, phytosteroids act in an antiestrogenic (antagonistic) manner by competing with endogenous estrogen for binding to the ESR. Conversely, under conditions of low endogenous estrogen levels, they can act estrogenically (agonistically) to help maintain hormonal cycles. 74 The combined effects of the P. angulata extract provided dual protection. ESR1 antagonism by Physalis F protects against excessive ESRα activation and maintains the follicular reserve. Partial agonism and antioxidant activity by flavonoids, namely Albanol A (–9.9 kcal/mol) and Rutin (–8.1 kcal/mol), also showed a strong affinity for ESR1. This mechanism stimulates a protective estrogen response while reducing oxidative stress (an antioxidant), which triggers ovarian damage.
The structural stability of the Albanol A–1X7R complex, characterized by its lowest RMSD value (1.414 Å), provides strong mechanistic evidence for the effectiveness of this compound as an ESR1 agonist. In molecular dynamics simulations, a low and stable RMSD value reflects a highly harmonious binding affinity, in which the ligand minimizes protein backbone fluctuations. Albanol A’s ability to maintain an agonist conformation with greater stability than the natural ligand, Genistein (1.669 Å), explains why the administration of 40 mg/kg BW P. angulata leaf extract significantly contributes to improvements in ovarian histology and estrous cycle duration, similar to the control group. Energetically, this stability supports the activation of estrogenic signaling pathways required for ovarian structural recovery after busulfan-induced damage. The most striking finding was the exceptionally low radius of gyration (Rg) for Albanol A (10.303 Å), indicating very tight protein structure packing. This structural compactness functionally reinforces the role of Albanol A in maintaining cellular integrity against oxidative damage, enabling functional recovery at the level of granulosa cells.75–77
In contrast, the estrogenic selectivity mechanism of P. angulata also manifests an antagonistic role mediated by Physalin F. Although the Physalin F–3ERT complex exhibits relatively high ligand fluctuations (RMSF) at 3.54 Å, its stability is maintained through the highest density of hydrogen bonds (196.026). These conformational dynamics suggest that Physalin F actively occupies the ESR1 antagonist pocket, which physiologically explains why ovarian improvement is not accompanied by increased pregnancy success. This antagonistic role triggers a functional blockade that is different from the direct DNA damage pathway caused by busulfan. While busulfan causes permanent damage to granulosa cells, the presence of Physalin F binding to ESR1 is suspected to create a temporary protective mechanism by halting cell proliferation to avoid busulfan cytotoxicity, but at the same time sends a “pseudo signal” to the hypothalamus.77,78
The systemic effects of this dual role cause inconsistent reproductive responses. In the short term, the pseudofeedback mechanism resulting from ESR1 inhibition by Physalin F can help normalize the estrous cycle through hormonal homeostasis, which is consistent with the data showing cycle recovery in the test subjects. However, the long-term stability of this antagonistic interaction disrupts the hormonal balance necessary to achieve the hormonal surge required for fertilization.79,80 Although the ovaries structurally appear to improve via the agonist pathway of Albanol A, functional failure—such as a decrease in conception rates—still occurs due to the dominance of the antagonistic mechanism of Physalin F in other target tissues, such as the uterus. The integration of these MD data confirmed that P. angulata metabolites act as SERMs, in which clinical efficacy is determined by the dynamic competition between the agonist stability of Albanol A and the antagonistic persistence of Physalin F at the estrogen receptor.
The integration of estrous cycle data, ovarian histology, and computational simulations confirmed that the efficacy of P. angulata leaf extract is determined by the dynamic competition between two major metabolites, Albanol A and Physalin F. Conformational analysis via molecular dynamics simulations indicated that Albanol A is the most stable agonist candidate at the ESR1 receptor, with a superior equilibrium profile evidenced by the lowest RMSD value (1.414 Å) and the most compact structure (Rg 10.303 Å). The high thermodynamic stability of Albanol A in the solvent environment provides a strong mechanistic explanation for the improvement in ovarian architecture and normalization of estrus cycle duration in the simultaneous treatment Group (B+C), where Albanol A maintains the estrogenic signaling necessary for granulosa cell regeneration following busulfan toxicity.
However, this study also revealed that ovarian structural recovery of the ovaries was not necessarily followed by a significant increase in pregnancy success. 81 This phenomenon is strongly suspected to be related to the role of Physalin F, which acts as a persistent ESR1 antagonist. Although Physalin F had a higher ligand fluctuation (RMSF 3.54 Å), the stability of its binding at the antagonist pocket was supported by the highest hydrogen-bond density (196,026). The presence of Physalin F creates a pseudofeedback mechanism capable of helping to normalize the hormonal cycle at the ovarian level; however, the persistence of this antagonist in target tissues, such as the uterus, can inhibit endometrial maturation and embryo receptivity.82,83
Although Physalin F contributes to low fertility rates through functional hormonal blockade, Albanol A is the superior component owing to its ability to maintain ovarian structural integrity and prevent ovarian failure. The stability of Albanol A in molecular dynamics simulations demonstrates its potential as a stable gonadotoxic protective agent, effectively mitigating the destructive effects of alkylating agents at the cellular and tissue levels.84,85
Limitations
This study had several limitations. First, the sample size per group (n = 5) was relatively small, although it fulfilled the minimum requirements of the Federer formula and demonstrated adequate statistical power in our post hoc analysis. A larger sample size in future studies could further strengthen the generalizability of these results. Further research incorporating more in-depth molecular signaling assays, such as western blotting or qPCR for ER-α and ER-β expression, would be highly beneficial in confirming the precise molecular pathways and long-term stability of the observed interactions.
Conclusion
The extract of P. angulata L. leaves contains secosteroids, phenolic, and flavonoid compounds with moderate antioxidant activity. Administration of P. angulata leaf extract together with busulfan has the potential to improve the estrous cycle and ovarian histological structure, but decreases pregnancy success rates in female rats experiencing ovarian damage due to busulfan treatment. Nevertheless, the observed restorative effect is not yet optimal. In silico studies showed that the metabolites in P. angulata leaf extract had a stronger binding affinity for ESR1 than for ESR2. Molecular dynamics analysis confirmed the role of Albanol A as a strong agonist with high stability and a highly compact protein structure, supporting the restoration of ovarian cellular integrity. Conversely, although Physalin F was flexible, it was identified as a strong antagonist that maintained binding stability through a superior hydrogen-bond density, thereby inhibiting the functional reproductive response.
Further research is warranted by adding a pretreatment group before busulfan injection to examine the effectiveness of P. angulata leaf extract as a preventive agent for ovarian damage. In addition, more in-depth studies are necessary to investigate the potential of P. angulata extract as a herbal contraceptive, considering the observed decrease in pregnancy success rates in this study, which was molecularly validated by the stability of the antagonistic interaction of Physalin F with estrogen receptors.
Footnotes
Acknowledgements
The authors would like to thank the Animal Laboratory Management Unit (UPHL) of the School of Veterinary Medicine and Biomedical Sciences (SKHB) IPB University for the experimental animal management facilities, the Advanced Laboratory of IPB University for the LC-HRMS facilities and in vitro tests, and Gadjah Mada University for the in vitro test facilities.
Authors’ Contribution
All authors made substantial contributions to conception and design, acquisition of data, or analysis and interpretation of data; took part in drafting the article or revising it critically for important intellectual content; agreed to submit to the current journal; gave final approval of the version to be published; and agreed to be accountable for all aspects of the work. All the authors are eligible to be authors as per the International Committee of Medical Journal Editors (ICMJE) requirements/guidelines.
Consent to Participate
Not applicable.
Consent for Publication
Not applicable.
Data Availability Statement
All the data are available with the authors and shall be provided upon request.
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Ethical Approval
The study protocol was approved by the Animal Ethics Commission of the School of Veterinary Medicine and Biomedicine (SKHB), IPB University, Indonesia (Approval No.: 247/KEH/SKE/IX/2024).
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: The author would like to thank the Education Fund Management Institution, Indonesia, for funding this research on behalf of YS (Contract number: SKPB-7572/LPDP/LPDP.3/2023) and IPB University BIMA fundamental research on behalf of ASS Contract number: 027/E5/PG.02.00.PL/2024.
Informed Consent
Not applicable.
Publisher’s Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of the publisher, the editors and the reviewers. This journal remains neutral with regard to jurisdictional claims in published institutional affiliation.
Use of Artificial Intelligence-assisted Tools
The authors declare that they have not used artificial intelligence (AI)-tools for writing and editing of the manuscript, and no images were manipulated using AI.