
Abstract
Background
The latest bladder cancer (BC) urine tests based on multiple genomic and/or epigenomic markers detect BC with high sensitivity and specificity. The GALEAS Bladder (GB) gene panel covers several actionable mutations, including in FGFR3.
Objective
To modify GB to detect FGFR3 fusions as well as single nucleotide variants and assess its potential as a urine-based companion diagnostic. We also study tissue-urine concordance and associations between FGFR3 genomic alterations and FGFR3 expression.
Methods
Probes were added to GB covering the 3’-region of FGFR3. The new panel was used to analyse genomic DNA extracted from 158 frozen primary BCs. Mutation concordance was also analysed in 107 tumour-urine pairs. Associations between FGFR3 mutations and expression were analysed in 608 FFPE BCs with the Decipher Bladder transcriptome assay.
Results and limitations
FGFR3-TACC3 fusions were found in 3/3 positive controls and 4/158 frozen BCs. Fusions were also detectable in genomic DNA from FFPE tumour and urine cell pellet DNA. All fusions were corroborated by PCR and Sanger sequencing. FGFR3 mutation status in urine was 94.4% concordant (95% CI 87.7–97.7%); GB detected mutations in urine with 92.1% sensitivity (95% CI 81.7–97.0%) at 97.7% specificity (95% CI 86.5–99.9). All FGFR3 mutations were associated with increased FGFR3 expression. This is a proof-of-principle study, rather than definitive evidence of utility as a companion diagnostic, with the need for fusion detection validation in larger patient cohorts.
Conclusions
In addition to BC detection, a modified version of GB demonstrates feasibility for tissue and urine-based detection of FGFR3 fusions.
Introduction
Fibroblast growth factor receptor (FGFR3) is a frequently mutated oncogene in bladder cancer (BC). 1 Activating single nucleotide variants (SNVs) are found in up to 70% of low-grade non-muscle-invasive BCs (NMIBCs) and 10–20% of high-grade NMIBCs and muscle-invasive BCs (MIBCs), predominantly at codons 248, 249, 370, 373 and 650.1,2 Additionally, 2–4% of BCs harbour FGFR3 gene fusions, most commonly with TACC3, which results in constitutively active FGFR3-TACC3 fusion proteins.3,4 Due to their frequent occurrence, FGFR3 mutations are useful components of gene panels for the non-invasive detection of bladder cancer.5–8 FGFR3 inhibitors can be effective treatments for patients whose BCs harbour FGFR3 mutations and FGFR3 mutations are therefore useful as both diagnostic and predictive biomarkers.9,10 Theoretically, a diagnostic urine test which includes FGFR3 mutations could also be used to select patients for anti-FGFR therapies (at least for cancers within the urinary tract) without the need for further testing, leading to savings in both time and cost.
The recently developed GALEAS Bladder (GB) test sensitively identifies tumour derived SNVs in urine DNA enabling non-invasive BC detection.7,8 GB is based on the capture and deep sequencing of 23 BC-associated genes (all exons of TP53 plus mutation hotspots in an additional 22 genes). GB includes all FGFR3 mutation hotspots and we now evaluate the addition to the panel of capture probes to target FGFR3 gene fusions. By adding capture probes to cover all of FGFR3 from exon 16 onwards (covering previously reported break-points3,4) we aimed to capture and sequence DNA molecules covering FGFR3-fusion breakpoints and identify all FGFR3 fusions regardless of where they occur in the genome. Because the assay does not require primers or probes for multiple potential fusion partners this represents a minimal change to the panel design: the panel remains small, economical to run and easy to analyse, and the test is not limited to specific fusion partners. We used this extended version of GALEAS Bladder (termed “GB-F”) to search for FGFR3 fusions and mutations in genomic DNA from 158 frozen BC tissues and subsequently in the FFPE tissue and urine of patients whose tumours harboured FGFR3 fusions. Additionally, we report novel data on the association of individual FGFR3 mutations, FGFR3 gene expression and gene signature activity in a cohort of 608 BC patients analysed with a clinical-grade transcriptome assay.
Methods
Patients and samples
All biospecimens were collected as part of the West Midlands Bladder Cancer Prognosis Programme (BCPP, ethical approval ref: 06/MRE04/65) between 2004 and 2011. 11 Participants were recruited following cystoscopy indicating the presence of primary bladder cancer and gave informed signed consent. Tumours were graded by pathologists using the 1973 WHO grading system which was used in the UK NHS at the time of patient recruitment. Voided urine was collected prior to TURBT and FFPE and fresh-frozen tissue were collected at initial TURBT. Genomic DNA was extracted from fresh-frozen tissue using DNeasy Blood and Tissue kits (Qiagen), FFPE tissue using QIAamp DNA FFPE Advanced kits (Qiagen), and from urine pellets using Quick DNA Miniprep Plus kits (Zymo); all DNA was quantitated using the dsDNA high sensitivity Qubit reagents.
The “discovery cohort” of 158 frozen tumours analysed with GB-F in this study were selected on the basis of sample availability and maximising overlap with existing paired urine data. The partially overlapping “urine-tissue” (n = 107) and “expression” (n = 608) cohorts are shown in Table 1 and Figure S1. GB data for the urine pellet DNAs has been published previously 7 and GB-F data was additionally generated for urine in 2 cases where the paired tumour harboured FGFR3 fusions and 4 wild type cases. Two bladder cancer cell lines previously reported to contain FGFR3 fusions and a bladder tumour in which we had previously identified an FGFR-TACC3 fusion by RNA sequencing were used as positive controls.
Patient information. Numbers in parenthesis are percentages.

Fusion detection
The GB-F assay was run according to the GALEAS Bladder manufacturer's instructions (Nonacus Ltd). The additional probes covered the 3’-region of FGFR3 (chr4:1,806,530-1,809,203, hg38). Briefly, 25 ng of genomic DNA was enzymatically fragmented, end repaired and A-tailed before adapter ligation. This was followed by PCR amplification, pooling, overnight capture with biotinylated probes, washing and a further limited-cycle PCR before 2 × 147 bp PE sequencing to a read depth of ∼30,000x (urine) or ∼500x (tissue). For consistency with previous publications7,12 SNVs were called if found at variant allele frequencies >0.5% in urine or >2.5% in tissue. Fusion events were identified by overlaying Genecode annotation with the aligned reads and defining fusion candidates by either shared individual reads between 2 genes (‘split reads’) or read pairs (‘discordant reads’). Fusions were called if supported by at least 50 reads and 2% read fraction (computed as the number of reads supporting the fusion divided by the average depth of the FGFR3 regions on the panel). Fusions were confirmed by PCR and Sanger sequencing of genomic DNA for 5 cases and cDNA sequencing for 4 cases where RNA was available.
Mutations and FGFR expression/signature
Gene expression analysis was performed for 608 FFPE BCs from the BCPP cohort with known FGFR3 single nucleotide variant (SNV) status. 12 RNA was extracted from macrodissected FFPE tissues and genome-wide gene expression analysed using the Decipher Bladder Genomic Subtyping Classifier (GSC) oligonucleotide microarray assay in a CAP/CLIA laboratory (Veracyte, Inc San Diego, CA). Data were SCAN normalized 13 and FGFR1-4 gene expression was extracted and signatures of FGFR3 co-expressed genes 14 and lncRNA-based FGFR3 activity 15 generated using locked classifiers available in the GRID bioinformatics platform (v3.1, Veracyte, Inc).
Statistical analysis
Gene expression or signature levels between patient groups were compared using Mann–Whitney U tests and mutation frequencies compared using chi-squared tests. Where gene expression was compared across multiple patient groups, Benjamini–Hochberg adjusted p-values (P-adj) are presented. A multivariable linear regression model was used to test the effects of FGFR3 mutations, stage and grade on FGFR3 expression. All statistical analyses were conducted using R (version 4.4.1).
Results
FGFR3 SNVs and fusions in BC tumour DNA
The GB-F panel was used to analyse tumour genomic DNA from the fresh frozen primary tumours of 158 BC patients, as well as genomic DNA from RT4 and SW780 cell lines and a fresh frozen NMIBC tumour previously reported to harbour FGFR3 fusions (Patient E). 16 A single FGFR3 SNV was found in 98/158 tumours, 3 tumours harboured 2 SNVs, and 57 tumours did not contain FGFR3 SNVs (Table 1).
Three samples known to harbour fusions were used as positive controls for the new method. All 3 positive controls (RT4, SW780, and Patient E) and 4 of the fresh frozen tumours (all without FGFR3 SNVs) were found to harbour FGFR3 fusions by GB-F. The cell line fusions were exactly as previously reported (RT4: FGFR3-TACC3 and SW780: FGFR3-BAIAP2L1). 4 In the tumours, all detected fusions were between FGFR3 and TACC3: data are shown in Figure 1, Figure S2 and Table S1. Of the 4 fusion-positive frozen tumours, in patient A (G1pTa) the fusion was between FGFR3 intron 17 (chr4:1806985) and TACC3 intron 10 (chr4:1738164); in patient B (G3pTa) the fusion was between FGFR3 intron 17 (chr4:1807052) and TACC3 intron 13 (chr4:1743693); in patient C (G3pT1) the fusion was between FGFR3 exon18 (chr4:1807144) and TACC3 intron 9 (chr4:1737442); in patient D (G3pTa) the fusion was between FGFR3 intron 17 (chr4:1806969) and TACC3 intron 10 (chr4:1738086); in patient E (G3pT1) the fusion was between FGFR3 intron 17 (chr4:1806987) and TACC3 intron 9 (chr4:1737513). Fusions were confirmed by Sanger sequencing of genomic DNA in all cases. Sanger sequencing of cDNA from the 4 newly discovered fusion cases (patients A-D) identified expressible FGFR3-TACC3 mRNAs with exon 17 of FGFR3 reading through into exon 10, 11 or 14 of TACC3 (Figures 1 and S2 and Table S1). The fusions appear to be most common in G3 NMIBCs; however, the numbers are small, the cohort is 90% NMIBC, and we did not find a significant association between fusion frequency and stage or grade.

FGFR3-TACC3 fusions identified in genomic DNA using GB-F and confirmed by Sanger sequencing of cDNA. NGS was performed on tissue DNA using GB-F. Sanger sequencing was performed on amplicons made with primers complementary to sequences in the exons adjacent to the genomic breakpoints identified by GB-F. Data shown for patients A& B. Data for patients C & D are shown in Figure S2.
For patients B and C (but not A and D), FFPE tumour tissue and urine cell pellet DNA were available: GB-F detected identical FGFR3-TACC3 fusions in genomic DNA extracted from fresh frozen and FFPE tumour tissue and urine cell pellets for both patients (Figure S2). FGFR3-TACC3 fusion detection was supported by over >300 breakpoint-spanning reads in urine DNA from patients B and C. We also used GB-F to analyse urine from 4 patients without fusions in their tumours and did not detect a single FGFR3-TACC3 fusion supporting read. As GB typically sequences >3000 unique urinary DNA molecules for each biomarker in the panel, the low background for fusion reads suggests that GB-F should be able to detect FGFR-TACC3 fusions even in urine DNAs with a low tumour fraction.
Concordance between FGFR3 SNV detection in urine and tissue
For 107 of the patients whose frozen tumours were analysed by GB-F, previously generated original GALEAS Bladder urine test data were available. The tissue FGFR3 status of the 107 tumours by GB-F was 44 wild type and 63 mutated (61 with SNVs and 2 with fusions). Overall, the FGFR3 mutation status of paired tumour-urine samples was the same for 101/107 patients (94.4% agreement, 95% CI 87.7–97.7). Considering both SNVs and fusions, FGFR3 alterations were detected in urine with 92.1% sensitivity (95% CI 81.7–97.0%) at 97.7% specificity (95% CI 86.5–99.9) (see Table S2).
In patients with wild type tumours, 43 out of 44 of the paired urines were also wild type; one urine sample was positive for FGFR3 SNVs (S249C, K650E, K650M, VAF<2%) that were not detected in the paired tumour. In patients with FGFR3-mutant tumours, the same SNV detected in the tumour was also detected in the urine in 56/61 (92%) of cases; 1 tumour also harboured a second, lower VAF mutation which was not detected in the paired urine sample. There were 5 false negative urines (urine samples where the tumour mutation was not detected). All false negative urines were from patients with stage pTa BC: 3 with G1pTa and 2 with G2pTa disease.
Effects of FGFR3 mutations on FGFR3 expression across stages and grades of BC
Previously acquired FGFR3 SNV data 12 and microarray-based genome-wide expression data were available for 608 BCs. FGFR3 was the most highly expressed member of the FGFR family (p < 0.001, Figure 2). FGFR3 expression significantly decreased with increasing stage of disease and is high in grade 1 and grade 2 disease (and not significantly different, p > 0.05) but significantly lower in grade 3 disease (p < 0.001).
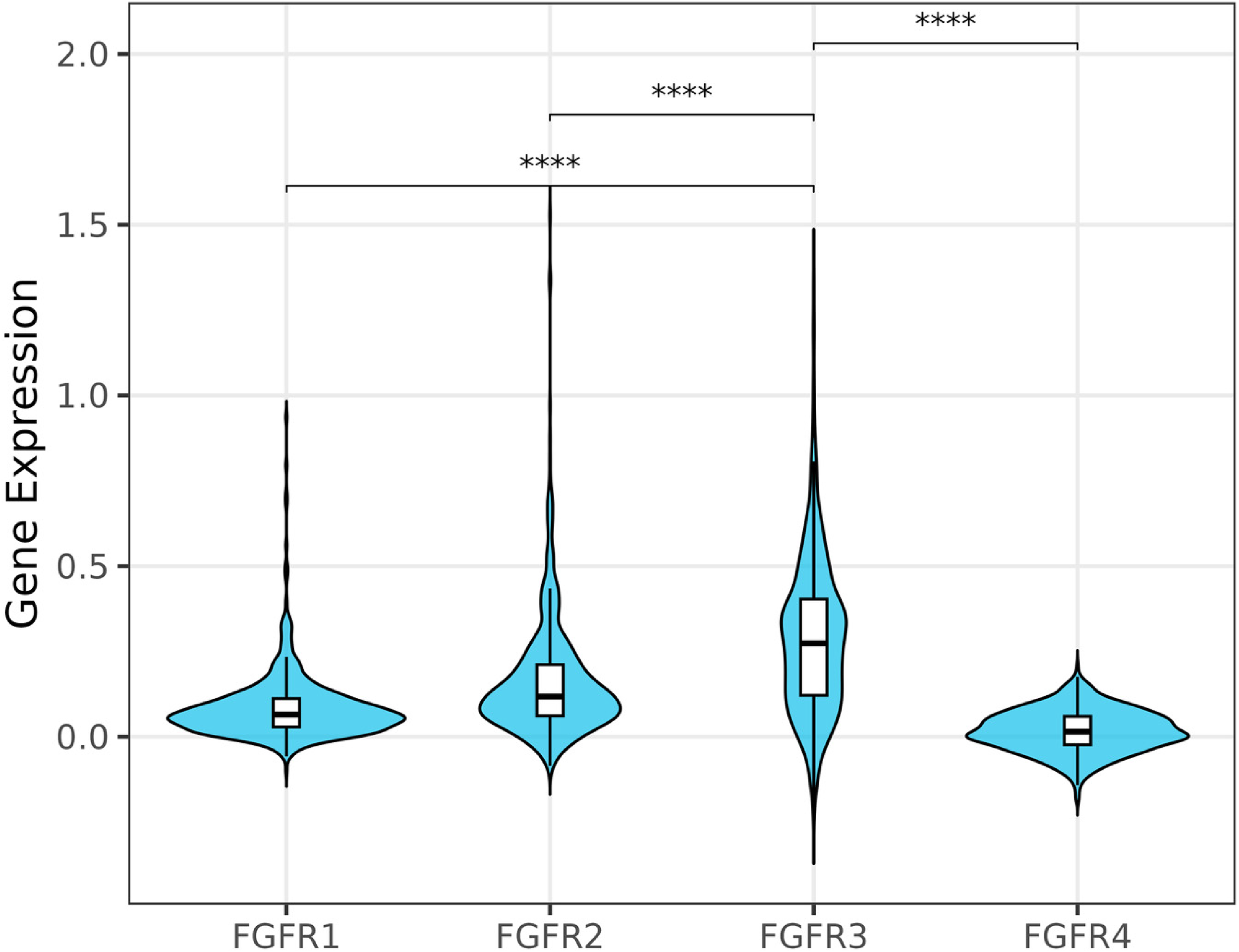
FGFR expression across 608 primary bladder cancers. Further details of FGFR1-4 expression across stages and grade of disease are shown in Supplemental Information. The y-axis represents normalised gene expression extracted from microarray data. **** P-adj < 0.0001.

FGFR3 expression across stages and grades of bladder cancer. Data is shown for FGFR3 wild type tumours in pale blue and FGFR3 mutated tumours in dark blue. The y-axis represents normalised gene expression extracted from microarray data. P-adj values shown for FGFR3 expression in wt vs mutant BCs in each stage or grade of disease.
As others have reported, FGFR3 is mutated in up to 70% of grade 1 & 2 NMIBCs and the frequency decreases with stage and grade to 15% in MIBC (Figure 3). FGFR3 mutation frequency was not significantly associated with gender (male = 44% mutated, female = 45%, p = 0.75), age (<71 years old = 48%, ≥71 years old = 41%, p = 0.08), smoking (never = 48%, ex = 45%, current = 45%, p = 0.87) but was associated with tumour size (<3 cm = 51%, >3 cm = 38%, p = 0.0005 (3 cm = median size)).
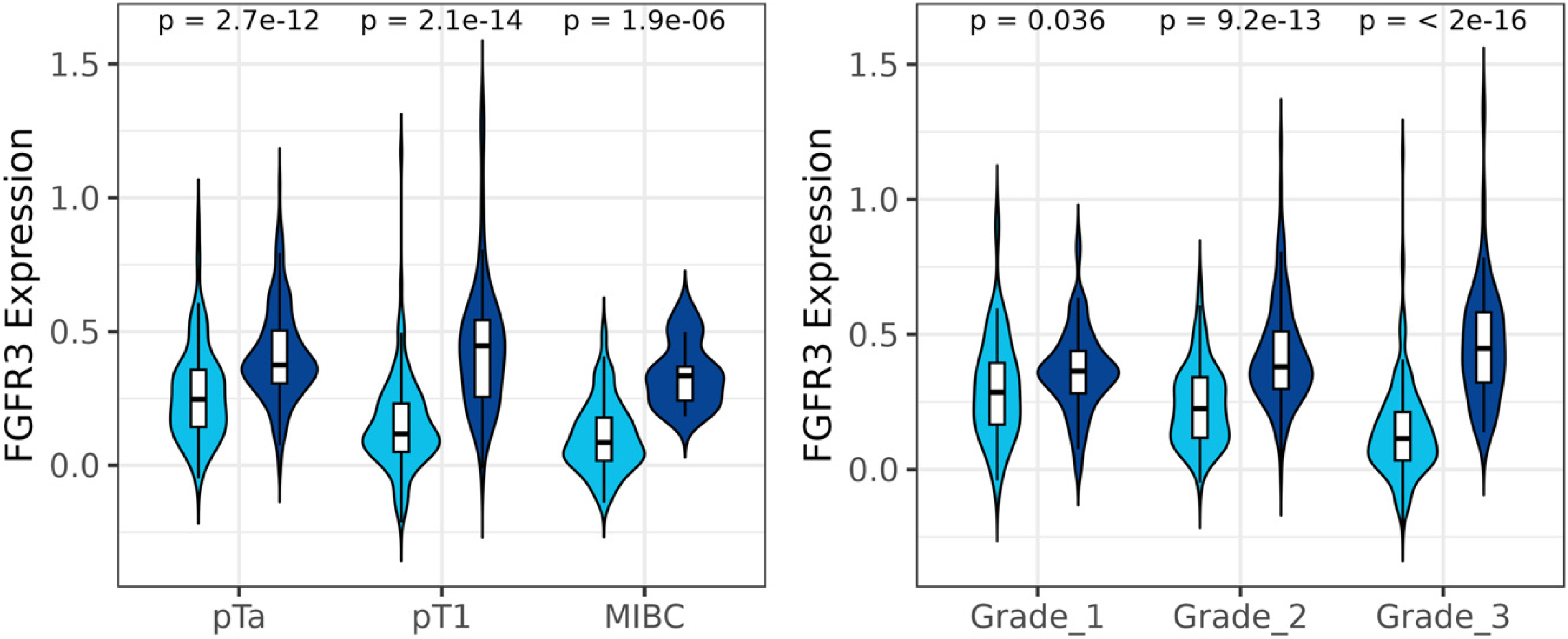
Across the whole cohort, FGFR3 was more highly expressed in BCs with FGFR3 mutations and this was also significant within each grade and stage of disease (Figure 4). The influence of FGFR3 mutations on FGFR3 expression was confirmed by multivariable statistical analysis including stage and grade (Figure S4). Mutations at all 5 hotspots are associated with significantly increased FGFR3 expression (Figure 5). The effect of FGFR3 mutation on expression is most marked in high grade disease. FGFR3 mutations were also significantly associated with increases in previously reported FGFR3 and lncRNA-FGFR3 activity signatures (Figure S5).14,15 As previous work used a cut-off on the continuous lncRNA-based activity score to identify luminal favourable tumours harbouring elevated FGFR3 signalling activity, 15 we compared FGFR mutations between luminal favourable and non-luminal favourable BCs. Luminal favourable BCs were significantly enriched for FGFR3 mutations (60% cases mutated versus 35% of cases, p < 0.0001).

The effect of individual FGFR3 SNVs on FGFR3 expression. Data are shown for all cases (pale blue) and for grade 1 and 2 combined (darker blue) and for grade 3 (darkest blue). Expression data for the single G370C mutant tumour in G3 patients has been omitted. Statistical testing was applied to FGFR3 expression in mutant versus wild type versus FGFR3 mutant cases for each mutation for the whole cohort and in grade 1 and 2 disease and in grade 3 disease in all cases where sufficient data existed for statistical testing (which excludes G370C and K650E/M/T in grade 3 where n = 1 and 2 respectively). **** indicates P-adj <0.0001, *** < 0.001, ** < 0.01. The y-axis represents normalised gene expression extracted from microarray data.
Expression data were available for 4 of the 5 tumours identified as having FGFR3-TACC3 fusions and all 4 BCs expressed FGFR3 at higher levels than the median level for FGFR3 wt BCs (p = 0.019). Across the 4 cases, levels of FGFR3 and TACC3 expression were correlated (r2 = 0.983) consistent with fusion-gene expression (Supplemental Figure 6).
Discussion
We have shown that a modified version of GALEAS Bladder with additional FGFR3 probes (GB-F) can detect FGFR3 fusions as well as SNVs in genomic DNA extracted from fresh frozen tumour, FFPE tumour and urine cell pellets. FGFR3 mutation status was concordant in 101/107 tumour-urine pairs (94.4% agreement). The most common disagreement between urine and tissue was false negative results from urine; we assume this to be due to low tumour DNA fraction and was only seen with grade 1 and 2 NMIBCs. Activating FGFR3 alterations and, in particular fusions,17,18 are known to confer sensitivity to the FDA-approved FGFR selective inhibitor Erdafitinib (Balversa). To be eligible for treatment with Erdafitib, patients must undergo a companion diagnostic test which adds complexity and cost to the patient pathway. Such tests use either next generation sequencing or reverse transcription-quantitative PCR to analyse archived FFPE-tumour tissue.19,20 Hence, both tissue availability and testing capacity may delay or limit access to a treatment with proven efficacy in intermediate and high-risk NMIBC 21 and muscle-invasive and metastatic BC.10,21 Our proof-of-concept study suggests that if, in the future GB-F urine testing becomes routine during diagnostic workup, then FGFR3 status would be known for all BC patients and this could avoid the need for additional companion diagnostic testing for anti-FGFR3 treatment. Thus, GB-F could serve not only as a detection test for bladder cancer, but potentially as a liquid biopsy companion diagnostic for anti-FGFR therapies, similar to how plasma ctDNA is already used to select lung cancer patients for anti-EGFR treatments. 22 Whilst urine may be the ideal liquid biopsy for NMIBC and organ-confined disease, a recent study in metastatic BC has reported 90% concordance between tissue and plasma FGFR3 alterations. 23
In addition, we have explored FGFR3 expression in the presence and absence of SNVs and find that FGFR3 SNVs result in higher FGFR3 expression in all stages and grades of disease. This effect is not just seen with the very common S249C mutation, but also with mutations at the other 4 hotspots. FGFR3 is expressed in all mutated tumours, supporting the notion that inferring FGFR3 inhibitor sensitivity directly from genomic DNA (rather than cDNA) is a valid approach. Van Rhijn et al. reported that FGFR3 is overexpressed not only in 73% of mutated tumours but also in 22% of wt tumours. 24 More recently, Eisner et al. 25 reported that 89–97% of patients with FGFR3-altered tumours expressed a “FGFR predictive response signature” (FGFR-PRS). The FGFR-PRS could potentially identify additional patients likely to respond to FGFR3 inhibitors and could become the tissue-based companion diagnostic of choice.
FGFR3 fusions were found in 4/158 bladder tumours, and the tumours with fusions all lacked other FGFR3 alterations. The fusions with TACC3 are likely to be driver events as they are in-frame fusions and result in the expression of mRNA containing at least exons 1–17 of FGFR3 and exons 14–16 of TACC3 (coding for the coiled-coil dimerization region of TACC3); these exons of TACC3 are the minimal requirement for constitutively active FGFR3-TACC3. 26
We have presented a simple method to detect FGFR3 fusions in urine DNA potentially enabling the use of urine as a liquid biopsy for BC treatment selection. Further work is required to determine the limit of detection and robustly determine sensitivity for FGFR3 fusion detection before clinical use. However, this study does illustrate the potential of gene panel urine-based diagnostic tests such as GALEAS Bladder to provide molecular information on BC in addition to a yes/no answer or probability of BC being present.
Supplemental Material
sj-docx-1-blc-10.1177_23523735261447112 - Supplemental material for Development of an extended version of GALEAS bladder: Detection of FGFR3 fusions in urine and associations between genomic alterations and gene expression
Supplemental material, sj-docx-1-blc-10.1177_23523735261447112 for Development of an extended version of GALEAS bladder: Detection of FGFR3 fusions in urine and associations between genomic alterations and gene expression by Jessica L Neil, Naheema S Gordon, Anshita Goel, Ayse N Myumyun, Benjamin Tura, Joep J de Jong, Elai Davicioni, Maurice P Zeegers, Kar Keung Cheng, Nicholas D James, Mario Neou, Roland Arnold, Richard T Bryan and Douglas G Ward in Bladder Cancer
Footnotes
Acknowledgements
Biospecimen collection was funded by Cancer Research UK. Sequencing costs were met by philanthropic donations to the University of Birmingham Bladder Cancer Research Centre. This work used the CaStLeS computing infrastructure at the University of Birmingham. Sequencing was performed at Genomics Birmingham.
Ethical approval and informed consent statements
All patients provided written signed informed consent. This study was conducted under UK ethical approval 06/MRE04/65 (East Midlands - Derby - Ethics Committee).
Author contributions
Data acquisition: JLN, NSG & DGW, data interpretation: AG, ANN, BT, JJJ, RA & MN, study design: DGW, RTB, MPZ, KKC & NDJ, manuscript writing and reviewing: ED, JJJ, RTB, DGW, RA.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: The BCPP study was funded by Cancer Research UK (C1343/A5738). Sequencing costs were met by philanthropic donations to the University of Birmingham Bladder Cancer Research Centre.
Declaration of conflicting interests
The author(s) declared the following potential conflicts of interest with respect to the research, authorship, and/or publication of this article: DGW and RTB co-developed the GALEAS® Bladder urine test with Nonacus Ltd (UK). RTB is a paid consultant for Cystotech ApS (Denmark) and an unpaid charity trustee for Action Bladder Cancer UK (UK). JJD is a paid consultant and ED an employee of Veracyte Inc, makers of the Decipher Bladder Genomic Subtyping Classifier tissue test (USA). MN is an employee of Nonacus Ltd.
Data availability statement
Data is available from the corresponding author upon reasonable request.
Supplemental material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.