
Abstract
Background
Social isolation significantly heightens the risk of dementia among the elderly. Maintaining social engagement has been proposed as a potential strategy to attenuate age-related cognitive decline.
Objective
This study investigates whether social interaction with young conspecifics could improve cognitive function in aged mice with chronic social isolation.
Methods
Twenty-month-old male C57BL/6 mice with long-term social isolation were randomly assigned to either cohousing with 3-month-old young mice or continued isolation for two months. Cognitive function was assessed using Y-maze spontaneous alternation and fear conditioning tests. Synaptic integrity and pathological markers were evaluated through western blotting and immunofluorescence.
Results
Cohoused mice exhibited significantly enhanced novel arm exploration in the Y-maze and improved contextual fear memory compared to isolated controls. Molecular analyses revealed increased synapsin-1 expression and decreased tau phosphorylation at Ser214 in the cohousing group. Immunofluorescence demonstrated greater astrocyte density in the dentate gyrus of cohoused mice.
Conclusions
Our findings demonstrate that social interaction with young counterparts can rescue isolation-induced cognitive deficits in aged mice, potentially through mechanisms involving tau phosphorylation regulation and synaptic protein restoration. These results provide preclinical evidence supporting social intervention strategies for preventing cognitive decline in socially isolated elderly individuals.
Keywords
Introduction
Aging is accompanied by a plethora of inevitable biological processes, leading to a range of gradual physiological and pathological alterations. 1 Within the central nervous system (CNS), aging escalates vulnerability to several neurodegenerative disorders and cognitive decline. 2 Approximately 10% individuals aged 65 years or older experience dementia or mild cognitive impairment (MCI), with prevalence increasing with advancing age. 2 Notably, prolonged social isolation has been shown to exacerbate these age-related cognitive deficits in both humans and animal models. Despite these well-established associations, effective interventions for isolation-induced cognitive impairment in the elderly remain limited.3,4
Social relationships constitute a fundamental human need across the lifespan, with particular importance in aging populations. 5 Social isolation detrimentally affects mental and physical health, causing depression, anxiety, insomnia, hypertension, and elevated cardiovascular risk.6,7 Epidemiological studies have identified that solitary living in older adults as a significant risk factor for accelerated cognitive decline and Alzheimer's disease (AD) pathogenesis. 8 While companion-based interventions have shown as an effective strategy to mitigate dementia risk,9,10 the underlying mechanisms remain incompletely understood. Previous studies using AD animal models have verified that social isolation promotes hallmark pathological features including amyloid-β deposition and neurofibrillary tangle formation. 3 11–14 However, the influence of social interaction on cognitive impairment and associated neuropathology in non-transgenic, naturally aging subjects remains largely unexplored.
In the present study, by Y-maze spontaneous alternation and fear conditioning tests, we found two months of social cohousing with young conspecifics significantly improved cognitive performance in aged mice. Molecular biological assessments revealed an elevated level of the presynaptic protein synapsin-1 and a reduction in tau phosphorylation at Ser214. Furthermore, cohousing increased astrocyte density in the dentate gyrus. Our findings provide compelling evidence that social interaction can ameliorate age-related cognitive decline and associated neuropathological changes, suggesting novel avenues for developing non-pharmacological interventions targeting socially isolated elderly populations.
Methods
Animals
18-month-old male C57BL/6J mice were individually housed under conditions preventing physical interaction. Following two months of social isolation (until 20 months of age), the aged mice were randomly assigned to two experimental groups: (1) a cohousing group, where each mouse was paired with a 3-month-old female conspecific, and (2) a control group that maintained individual housing. Mice were housed in a standard condition (12 h light/dark cycle, 50–60% relative humidity and keep 25°C). Following 2 month, mice underwent a series of behavioral assessments, followed by euthanasia through isoflurane gas for biochemical evaluations. All animal handling protocols adhered strictly to the NIH guidelines for the care and use of laboratory animals (NIH Publications No. 8023, revised 1978) and were approved by the Medical Ethics Committee of Jianghan University.
Y-maze test
The Y-maze was used to test the spatial exploration and memory capabilities. 15 In brief, mice were placed in the starting arm of the Y-maze and allowed to explore for 5 min, while one arm remained accessible, and the other was closed. Following a designated exploratory period, mice were reinstated in the starting arm, both arms were opened, and the time spent in the new arm versus total exploration time (5 min) was recorded.
Fear conditioning
Fear conditioning was used to evaluate associative learning and memory. 16 On the first day, mice were given 2 min to acclimate in the conditioned chamber in order to establish a baseline freezing rate. An acoustic cue was presented for 10 s, culminating in a 2-s foot shock. A second round was conducted after a 60-s interval. On the following day, mice were placed in the platform, and the freezing response was recorded as contextual freezing. Following a change of the platform's shape without shock, cued freezing was assessed via the sound stimulus alone.
Western blot
Upon euthanasia, brain tissues were promptly extracted from the mice. RIPA lysis buffer (Beyotime, Nantong, China) was utilized to extract total protein from isolated hippocampal and cortical tissues. Sodium dodecyl sulfate-polyacrylamide gel electrophoresis was performed for protein analysis, followed by transfer to nitrocellulose membranes (Millipore, Burlington, MA, USA). Membranes underwent a blocking procedure with 5% nonfat milk, followed by overnight incubation with primary antibodies (detailed in Table 1). After washing, secondary antibodies were applied, and protein expression was visualized using an enhanced chemiluminescence method (Bio-Rad, Hercules, California, USA).
The primary and secondary antibodies used in this study.

mAb: monoclonal antibody; pAb: polyclonal antibody.
Immunofluorescence
Following fixation, brains were sliced into 30 μm sections using a vibratome (Leica VT1000S, Wetzlar, Germany). After blocking with a solution of 0.1% Triton X-100 and 5% bovine serum albumin, the sections were incubated with primary antibodies overnight (detailed in Table 1) and subsequently secondary antibodies for 1 h. Nuclei were stained with Hoechst dye (Invitrogen, Waltham, MA, USA) for 10 min. Imaging was conducted using a laser scanning confocal microscope (Leica SP8, Germany).
Data analysis
Statistical analyses were performed using GraphPad Prism 8 software (GraphPad Software Inc., MA, USA). Differences between two groups were analyzed using two-tailed unpaired Student's t-tests, with significance determined as *p < 0.05. Results are expressed as means ± standard error.
Results
Social cohousing ameliorates cognitive deficits in aged mice with prolonged social isolation
Previous studies have indicated that 8-week social isolation induces cognitive impairment in 18-month aged mice. 4 Building upon this evidence, we established an experimental model using 18-month-old male C57BL/6J mice that were individually housed under conditions preventing physical interaction. Following two months of social isolation (until 20 months of age), the mice were randomly assigned to two experimental groups: (1) a cohousing group, where each aged mouse was paired with a 3-month-old female conspecific, and (2) a control group that continued individual housing. Behavioral observations confirmed increased social interaction between aged and young mice in the cohousing condition. After two months of cohabitation, the aged mice were subjected to Y-maze and fear conditioning assessments (Figure 1A). The Y-maze test revealed that cohousing mice exhibited significantly greater exploration of the novel arm compared to isolated controls (Figure 1B), suggesting enhanced spatial memory retention. Fear conditioning tests further demonstrated superior memory performance in cohoused mice, as evidenced by significantly increased freezing responses to both contextual and cued fear memory tests (Figure 1D, E), with no observable differences in baseline freezing behavior (Figure 1C). These findings collectively indicate social cohousing with younger conspecifics effectively reverses isolation-induced cognitive deficits in aged mice.

Co-housing improves cognitive performance in aged mice following social isolation. (A) Schematic of the experimental design and behavior tests. (B) New arm ratio of two groups of animals explored the in the Y-maze. (C-E) Freezing ratio of baseline, cued induced and contextual induced fear conditioning test in two group mice (n = 10–15 per group, *p < 0.05, **p < 0.01, ***p < 0.001, two-tailed unpaired t-test). Data expressed as mean ± SEM. The cartoon in panel A comes from www.biorender.com.
Social cohousing attenuates hippocampal synaptic deficits in aged mice following social isolation
Synaptic integrity is a crucial determinant of cognitive function. 17 To investigate whether social cohousing modulates synaptic protein expression, we analyzed key pre- and postsynaptic protein in the hippocampus. Western blot analysis revealed that while levels of postsynaptic density proteins (PSD93 and PSD95) remained unchanged between groups, the presynaptic protein synapsin-1 was significantly upregulated in cohoused mice compared to isolated controls (Figure 2A, B). This finding was further corroborated by immunofluorescence staining, which demonstrated enhanced synapsin-1 expression in the dentate gyrus and CA1 of cohoused mice (Figure 2C). These findings suggest that the co-housing effectively rescues synaptic impairments in the hippocampus, which may contribute to the remission of cognitive deficits of the isolated aged mice.
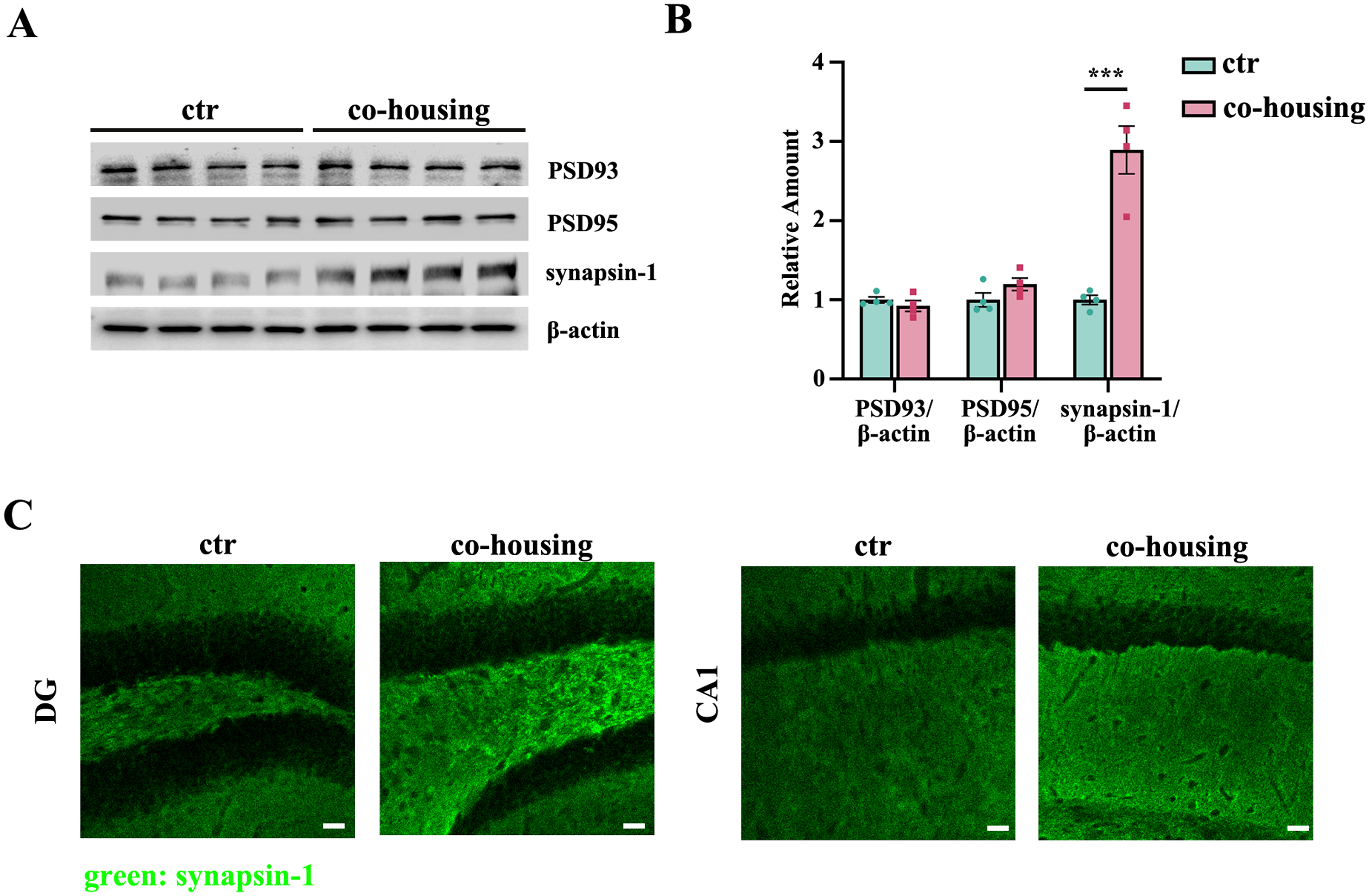
Co-housing alleviates synaptic damage in aged mice following social isolation. (A) Immunoblotting and (B) quantification of PSD93, PSD95, and synapsin-1 in hippocampus of two group mice (n = 4 per group, ***p < 0.001, two-tailed unpaired t-test). (C) Representative immunofluorescence images of synapsin-1 in DG and CA1 (green: synapsin-1, scale bar = 30 μm). Data expressed as mean ± SEM.
Social cohousing attenuates tau hyperphosphorylation in the aged mice
Tau hyperphosphorylation is a crucial hallmark of several neurodegenerative diseases. 18 Western blot analyses revealed substantial reductions in tau phosphorylation at Ser214 in the social cohoused aged mice compared to controls (Figure 3A, B). This finding was further supported by immunofluorescence staining, which revealed decreased p-Tau (Ser214) immunoreactivity in multiple brain regions, including the cortex, dentate gyrus, and CA1 (Figure 3C, D).

Co-housing decreases tau phosphorylation in aged mice following social isolation. (A) Immunoblotting and (B) quantification of phospho-tau (Ser 199), phospho-tau (Ser 214), total tau, CDK5 and p25/p35in hippocampus of two group mice (n = 4 per group, *p < 0.05, **p < 0.01 two-tailed unpaired t-test). (C) Representative immunofluorescence images of phospho-tau (Ser 214) in cortex, DG and CA1 (green: phospho-tau (Ser 214), scale bar = 50 μm). (D) Quantification of phospho-tau (Ser 214) in cortex, DG and CA1 (n = 3 mice per group, **p < 0.01, two-tailed unpaired t-test). Data expressed as mean ± SEM.
To elucidate potential mechanisms, we investigated the CDK5 signaling pathway, a key regulator of tau phosphorylation. Although total CDK5 expression remained unchanged between groups, cohousing significantly reduced the p25/p35 ratio, a critical determinant for CDK5 activation (Figure 3A, B). These results suggest that social interaction may mitigate tau pathology through modulation of CDK5 activity in the brain of socially isolated aged mice.
Social cohousing promotes hippocampal astrogenesis in aged mice following social isolation
Glial cells are essential for maintaining CNS homeostasis and supporting neuronal function.19–21 While microglia, the brain's resident immune cells, undergo characteristic morphological changes (e.g., adopting an amoeboid shape) during neuro-inflammatory states, 22 morphological analyses of microglia showed no significant alterations in microglial surface area, volume, or sphericity between groups (Supplemental Figure 1A-D). These findings suggest that the beneficial effects of social cohousing may be independent of microglia.
Notably, we observed profound effects on astrocyte populations. Given that astrocyte is essential for synaptic plasticity, neuroprotection and metabolic support for neurons, and given the well-established association between astrocyte depletion and dementia progression,23,24 we quantified GFAP-positive cells in the hippocampus. Strikingly, cohoused mice demonstrated a dramatic 7-fold increase in astrocyte density within the dentate gyrus compared to socially isolated controls, and an approximately 3-fold increase in CA1 (Figure 4A-C). Considering astrocytic neurotrophic functions and neuroprotection, this robust gliogenesis may underlie the cognitive improvements observed in cohoused animals.

Co-housing increases the number of astrocytes in aged mice following social isolation. (A) Representative immunofluorescence images of GFAP+ astrocytes in DG and CA1 (blue: Hoechst, green: GFAP, scale bar = 50 μm). (B and C) Quantification of number of GFAP+ astrocytes in DG and CA1 (n = 3 mice per group, *p < 0.05, two-tailed unpaired t-test). Data expressed as mean ± SEM.
Discussion
The growing prevalence of dementia in aging populations underscores the urgent need for effective preventive strategies. Epidemiological studies have identified social isolation as a significant modifiable risk factor for cognitive decline and dementia.5,8,9,25 Our study has demonstrated that social cohousing can effectively mitigate isolation-induced cognitive impairments in aged mice through multiple neuroprotective mechanisms. The CNS's glial cells, including microglia, astrocytes, and oligodendrocytes, play integral roles in dementia pathogenesis.19,21,26,27 While our study found no significant microglial activation, we observed remarkable astrocyte proliferation in cohoused animals, particularly in hippocampal regions critical for learning and memory. This finding aligns with previous reports that social enrichment enhances brain-derived neurotrophic factor release, promotes neurogenesis, and modulates neuro-inflammatory responses in AD models.28,29 In addition, our current findings have indicated that social interaction ameliorates tau pathology and improves synaptic integrity. In conclusion, our study demonstrates that social cohousing effectively rescues isolation-induced cognitive deficits in aged mice through coordinated improvements in synaptic function, tau pathology, and glial support. These results underscore the potential of non-pharmacological, socially-based approaches for dementia prevention and highlight new directions for therapeutic development.
Neurofibrillary tangles, composed of hyperphosphorylated tau protein, are one of the neuropathological hallmarks of AD. 30 Tau hyperphosphorylation at Ser214, considered a pivotal phosphorylation site, has been linked to neurodegeneration, underscoring the necessity of regulating tau phosphorylation to mitigate AD progression.31,32 Through western blot and immunofluorescence analyses, we demonstrated that social cohousing significantly reduced tau hyperphosphorylation at Ser214 in aged mice. Cyclin-dependent kinase 5 (Cdk5) has been identified as an important kinase that upregulates tau hyperphosphorylation at Ser214. P35/P25 was identified as the Cdk5 regulatory subunit and was found to be necessary for Cdk5 activity. Because P25 binds directly to Cdk5 and has a 5- to 10-fold longer protein half-life than p35, it prolongs Cdk5 activation.33,34 In present study, we found that co-housing lowers p25/p35 ratio and attenuates tau phosphorylation in the aged mice.
CNS glial cells, including microglia, astrocytes, and oligodendrocytes, are integral in sustaining cerebral homeostasis. 19 Microglia serve multiple roles in CNS, including participating in neuronal circuit formation and debris clearance. 22 In AD, the inflammatory response mediated by microglia could exacerbate related pathologies, including neurofibrillary tangles and plaques. Prior inquiries indicate that social interaction mitigates microglia-driven inflammation in murine models. 35 Our study surprisingly found no evidence of reduced microglial activation following cohousing. This suggests that the cognitive benefits of social interaction in our model may operate through microglia-independent mechanisms. Instead, we identified a robust increase in astrocyte proliferation among cohoused animals. Astrocytes perform essential neuroprotective functions including trophic support for neurons, regulation of lipid metabolism, and maintenance of neuronal excitability. 19 Notably, since astrocytic apoptosis and pyroptosis have been implicated in AD pathogenesis,23,36 the observed astrocyte proliferation may represent a crucial mechanism by which social interaction confers neuroprotection in the aging brain.
Recent study has shown that the social activities of animals are regulated by neurons in the CA2 region of the hippocampus. 37 The excitatory or inhibitory neurons in this region are regulated by surrounding perineuronal nets (PNNs).38,39 To further investigate whether social interaction affects the expression of PNNs in the CA2 region of mice, we stained the PNNs in the mouse hippocampus with FITC-labeled wisteria floribunda lectin (FITC-labeled WFA, Invitrogen L32481, USA). However, the results showed that there was no significant difference in the expression intensity and area of PNNs in the CA2 region of mice (Supplemental Figure 2A-C). This result reveals that the influence of co-housing behavior on the cognition of elderly mice may not be mediated through the regulation of PNNs.
To maintain experimental consistency, this study exclusively utilized aged male C57BL/6J mice. And we employed young female mice for cohousing to prevent aggression-related confounding factors, as male-male cohousing often leads to territorial conflicts that could compromise welfare and experimental outcomes. While this design ensured controlled variables, it necessarily raises important questions regarding potential sex-specific responses to social interaction, as suggested by previous studies demonstrating sexual dimorphism in stress responses and social behavior.40,41 Future investigations should systematically examine these gender differences to provide a more comprehensive understanding of social intervention effects. In summary, our experiments provide compelling evidence that social interaction can effectively ameliorates isolation-induced cognitive deficits in aged mice. The demonstration that non-pharmacological social intervention can simultaneously target multiple pathways of neurodegeneration suggests promising avenues for developing community-based strategies to prevent or delay cognitive decline in elderly individuals.
Supplemental Material
sj-docx-1-alr-10.1177_25424823251394408 - Supplemental material for Social interaction mitigates cognitive impairments and tau pathology in socially isolated aged mice
Supplemental material, sj-docx-1-alr-10.1177_25424823251394408 for Social interaction mitigates cognitive impairments and tau pathology in socially isolated aged mice by Jiayu Zeng, Zhenzhen Bao, Wanbing Huang, Adan Fu, Zheng Liang and Hongmei Zhu in Journal of Alzheimer's Disease Reports
Footnotes
Acknowledgements
The authors thank the Hubei Province health and family planning scientific research project (Grant No. WJ2023F039) and the Research Fund of Jianghan University (Grant No. 2022SXZX29).
Ethical considerations
The animal study protocol was approved by the Ethics Committee of Jianghan University (approval JHDXLL2022-072) for studies involving animals.
Consent to participate
Not applicable
Consent for publication
Not applicable
Author contribution(s)
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by the Research Fund of Jianghan University, Hubei Province health and family planning scientific research project, (grant number 2022SXZX29, WJ2023F039).
Declaration of conflicting interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Data availability statement
The data support the findings of this study are available from the corresponding authors upon reasonable request.
Supplemental material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.