
Abstract

CRISPR-Cas9 has had a tremendous impact on research and biotechnology due in part to its simple programmability. A single guide RNA (gRNA) can direct Cas9 to its complementary DNA target within a large and complex genome to enable precise genome engineering. Although the activity of this Cas9-gRNA complex can be influenced by the local chromatin context,1–3 most CRISPR systems do not directly sense or respond to specific epigenetic marks. As a result, this key regulatory layer of the genome is unaccounted for by the targeting mechanism of current CRISPR approaches. Writing in Nature, Roth et al. introduce ThermoCas9, a methylation-sensitive Cas9 variant that can leverage this regulatory information, expanding the transformative CRISPR toolkit and offering an opportunity for epigenome-guided genome engineering. 4
ThermoCas9 behaves like a conditional editor. Its activity is gated on the methylation status of the cytosine in the fifth position within its protospacer adjacent motif (PAM) sequence 5′-NNNNCNR-3′. When the PAM contains an unmethylated CpG or CpC motif, ThermoCas9 can bind and cleave DNA as expected. However, when that cytosine is methylated, ThermoCas9 binding is disrupted, and no DNA editing can occur. This feature is a notable distinction from the authors’ previously described methylation-sensitive system AceCas9, which can maintain binding to methylated DNA but fails to initiate the conformational change required for cutting. 5
The gating of ThermoCas9 activity at the DNA binding step could have profound implications for the applications of this technology. The modern CRISPR toolkit generally achieves its specificity of action by relying on the programmable DNA binding of Cas proteins rather than modulating their associated catalytic activity. Nuclease-deactivated Cas9 (dCas9) is often fused to effector domains with their own catalytic activity to achieve various forms of editing, including epigenetic modification, 6 single-base modification with base editing, 7 or complex genetic writing with prime editing. 8 ThermoCas9 is poised to enable methylation-specific activity across these diverse technologies.
To reveal the mechanism of methylation sensitivity, the authors performed cryo-EM of ThermoCas9 in complex with its DNA substrate. Insight into the structure of a molecular system is often a key bridge between observing how the natural system works and informing how it might be engineered for translational applications. In this case, the authors identified extensive interactions between three amino acids (Arg1035, Asp1017, and Ser1019) within the PAM-interaction domain that contact the C-G base pair and seemingly provide little room to accommodate an additional methyl group.
Beyond its compelling mechanism, ThermoCas9 poses many exciting applications (Fig. 1). Perhaps one of the most immediate opportunities lies in cell-type-specific activity, exploiting the distinct methylation profiles across cell and disease states. The authors focus on this application in the context of cancer, where widespread changes in methylation are common and can be a distinguishing feature from healthy cells. 9 In principle, one could design gRNAs targeting loci that are hypomethylated in tumor cells but methylated in normal tissues. ThermoCas9 would then selectively bind and edit cancer cells while sparing healthy ones. Such a strategy could be used to edit cancer-causing genes or knock-in toxic payloads. While conceptually appealing, translation of this strategy will depend on overcoming some practical hurdles, including achieving efficient delivery of CRISPR components to tumor cells.
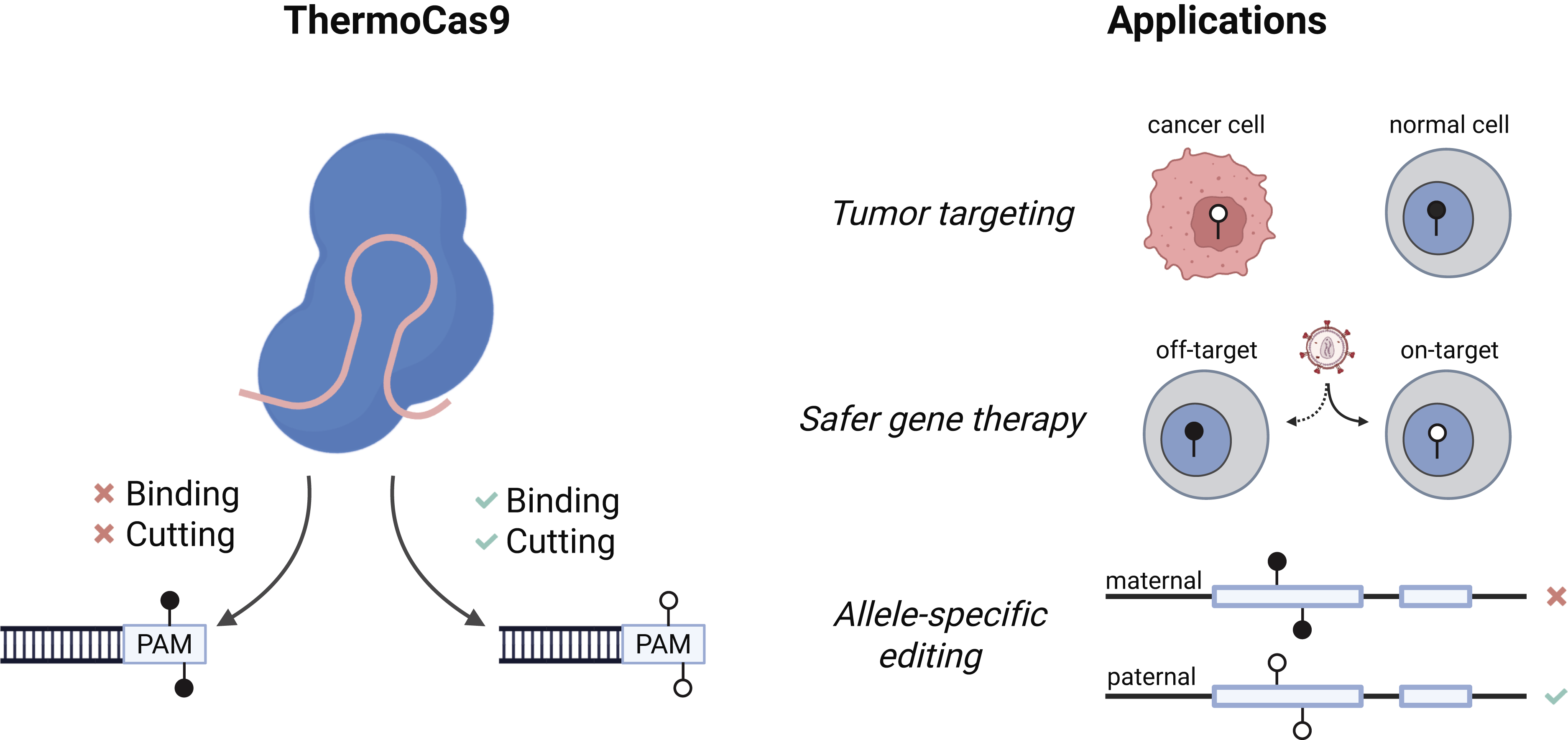
Methylation-dependent DNA targeting with ThermoCas9. (Left) ThermoCas9 activity is gated on the methylation status of a cytosine within its protospacer adjacent motif (PAM). At unmethylated PAM sites, ThermoCas9 can bind and cut DNA, whereas methylation of the PAM cytosine disrupts binding and prevents editing. (Right) This methylation-dependent activity supports exciting applications in biotechnology and therapeutic development. In cancer, ThermoCas9 could preferentially target hypomethylated tumor cells while sparing methylated healthy cells. In gene therapy, it may provide an additional safeguard by restricting editing to intended cell types while limiting activity in off-target tissues. Finally, ThermoCas9 could achieve allele-specific targeting at differentially methylated loci, such as in imprinted regions, which could enable novel therapeutics or improve disease modeling. Created in Biorender.
ThermoCas9 could more broadly provide an additional layer of specificity to in vivo gene therapies. The most optimized, cell-type-specific delivery vehicles often still can distribute to unintended tissues. As one example, recent advances in targeted delivery have enabled the generation of CAR-T cells directly in vivo, but these systems can have measurable delivery to off-target tissues such as the liver. 10 In a recent study, S. pyogenes Cas9 (SpCas9) was used to achieve CAR knock-in into the TRAC locus directly in vivo. 11 Notably, the TRAC locus is relatively demethylated and accessible in T cells but often remains methylated in most other cell types, 12 presenting an opportunity for ThermoCas9 to act as an additional safeguard by restricting editing to the intended cell type. To extend this further, ThermoCas9 could even enable selective editing of specific T cell subsets defined by their unique DNA methylation patterns. 13
Another compelling application lies in allele-specific targeting. Genomic imprinting, where one allele is epigenetically silenced, presents a natural testbed for ThermoCas9. The ability to selectively edit the unmethylated allele while leaving the methylated counterpart untouched could enable more precise control of imprinted loci with potential applications both therapeutically and to facilitate the development of cellular and animal models of imprinting disorders.
As a last example, the low-cost and point-of-care diagnostic space may also benefit from this technology. Methylation-sensitive restriction enzymes have long been used to detect DNA methylation for cancer diagnostics, but they are limited in scope by fixed recognition sequences. 14 ThermoCas9 offers programmability and multiplexing with gRNAs to direct Cas9 to virtually any locus of interest, providing an opportunity for more customizable and targeted detection systems.
Despite these exciting prospects, several key questions and challenges remain before ThermoCas9 can be broadly adopted. First, the current study examined a relatively small number of genomic targets. A more comprehensive dataset is needed to understand how sequence context and local chromatin state influence ThermoCas9 activity and methylation sensitivity. High-throughput methods, such as pooled CRISPR screens targeting thousands of sites, could provide valuable insights into the generalizable rules governing ThermoCas9 activity.
Additionally, ThermoCas9’s PAM requirements constrain the targetable sites. The preferred motif (5′-NNNNCGAA-3′) only occurs a few times per kilobase, which will limit flexibility and broad application. Expanding the targeting range of ThermoCas9, either through directed evolution or rational engineering, as has been demonstrated with SpCas9, 15 will be a critical step toward making this system broadly useful.
ThermoCas9 represents an important step toward integrating epigenetic information into genome engineering. This work also raises an intriguing possibility: Could we engineer the inverse system? A Cas9 variant that preferentially binds methylated DNA would enable complementary avenues, particularly for diseases driven by aberrant hypermethylation. More advanced systems in the future may even take advantage of other locus-specific information, such as histone modifications, nearby transcription factor binding, or chromatin conformation, for modulating CRISPR activity. The structural mechanisms of ThermoCas9 methylation sensitivity revealed here provide an exciting foundation for further engineering of CRISPR systems to achieve orthogonal epigenetic-dependent functions.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
No funding was received for this work.