
Abstract
Purpose:
This study investigates two electromagnetic field (EMF) modalities on cancer cells: near-infrared pulsating light (NIR) and hyperpolarized light (HPL).
Methods:
PC3, MCF7, and HeLa were exposed to Bioptron (HPL, 40 mW/cm2) and VieLight NeuroPro (NIR, 4.59 mW/cm2, 810 nm, 10 Hz) for 10–60 min, with intermittent breaks. Viability, cytoskeletal and mitochondrial morphology, ATP, and lactate levels were assessed.
Results:
NIR primarily enhanced mitochondrial ATP production, while HPL induced early morphological changes. Both modalities showed exposure dependency on viability.
Conclusion:
The analysis reveals distinct cellular responses to NIR and HPL, underscoring the importance of optimized protocols for effective EMF therapies.
Introduction
The human body naturally generates endogenous electromagnetic fields (EMFs) through essential physiological processes such as ion transport and cellular metabolism, establishing an electromagnetic baseline that is fundamental to homeostasis.1,2
In the field of oncology, the therapeutic application of EMFs is well established, most notably through radiotherapy, which employs high-energy ionizing radiation to induce DNA damage and apoptosis in cancer cells.3,4 More recently, non-ionizing modalities have gained attention for their therapeutic potential. For instance, photobiomodulation (PBM) utilizes red and near-infrared light (600–1100 nm) to stimulate cellular processes.5–8 Discovered accidentally by Endre Mester in the 1960s,5,9 PBM operates primarily through cytochrome C oxidase activation, triggering retrograde mitochondrial signaling that modulates gene expression and cellular metabolism.10–14 Its therapeutic efficacy exhibits a biphasic dose-response relationship, necessitating careful parameter optimization.15–18 Among PBM devices, the Vielight NeuroPro (https://www.vielight.com/; Vielight, Toronto, Canada) operates in the near-infrared (NIR) range (810 nm, 4.59 mW/cm2, 10 Hz frequency sweep) and has been investigated for its effects on cellular structures, including the disassembly of microtubules and enhancement of cellular functions in vitro.9,19 Hyperpolarized light therapy represents another innovative approach. The Bioptron Brewster optical unit (https://www.bioptron.com/) generates a unique Fibonacci-sequenced photon pattern through a C60 fullerene interface, 20 with preliminary evidence suggesting immune modulation and tumor-suppressive effects, potentially through mechanisms involving immune modulation, increased oxidative stress, or disruption of cancer cell metabolism.21–24 However, the precise mechanisms by which hyperpolarized light affects cancer cells remain inadequately understood. This study aims to directly compare these two illumination regimes on cancer cells by systematically varying exposure parameters to: characterize dose-dependent metabolic responses; assess morphological changes; and elucidate mechanistic differences between their therapeutic actions. The findings will provide insights for optimizing non-ionizing EMF therapies in oncology, particularly for regimens targeting cancer cell metabolic vulnerabilities.
Materials and Methods
Cell culture
The prostate cancer cell line PC3 was purchased from ATCC. The HeLa cell line and MCF7 cell line were obtained by Dr. Godbout’s laboratory at the Cross Cancer Institute. Cells were cultured in Dulbecco’s Modified Eagle Medium supplemented with 10% (V/V) fetal bovine serum (FBS) and 1% antibiotics (100 U/mL penicillin and 100 μg/mL streptomycin). Standard culture conditions were set at 37°C with 5% of CO2.
Sample preparation and irradiation
Cells were plated into µ-Dish 35 mm high-wall glass-bottomed dishes with a cell density of 25 × 104, following incubation overnight. Then, serum starvation for 24 h to synchronize them to the G0/G1 phase to monitor possible delays in the mitotic stage.
For the NIR radiation, the VieLight device NeuroPro (810 nm) Module A was utilized, with an output power density of ∼4.59 mW/cm2 at the distance of 125 mm and a frequency sweep of 10 Hz. Regarding HPL, the power density of the Bioptron device was 40 mW/cm2. Both devices have tunable exposure time. Irradiation was performed at ambient temperature (21.5°C). A summary of the specifications is reported in Tables S1 and S2.
Cell viability
Cell viability was assessed 24 h post-irradiation using the Alamar Blue assay. Fluorescence was measured with a FLUOstar Omega plate reader (BMG LABTECH, Germany) at 544 nm excitation and 590 nm emission. Experiments were performed in triplicate in 96-well plates, with fresh medium used as a blank.
Immunofluorescence
Gelatin-coated coverslips were placed in each dish and allowed to dry. Cells were then fixed, permeabilized, and stained to visualize actin, tubulin, and mitochondria. Confocal imaging was performed using a Zeiss 710 microscope (Carl Zeiss AG, Oberkochen, Germany). Detailed protocols are provided in the Supplementary Data.
Glycolysis-Oxphosphorylation
In total, glycolytic and mitochondrial ATP and the lactate production in cells were assessed by using the Glycolysis/OXPHOS Assay kit (Dojindo Molecular Technologies, Tokyo, Japan), following the manufacturer’s specifications.
Data analysis
Normality was assessed with the Shapiro–Wilk test. Multiple unpaired t tests with Holm-Šídák correction were employed to compare each group. Experiments were conducted in triplicate, and results are expressed as mean ± SD. Statistical significance was determined by setting a threshold of p < 0.05. Data analysis was performed using GraphPad Prism (version 10.0).
Results and Discussion
The potential effects of PBM therapy on cellular processes have fueled an ongoing debate regarding its efficacy in cancer treatment. 10 To explore cell line-specific characteristics and irradiation parameters, our study aimed to report novel insights on the application of two light-emitting devices: NeuroPro 1 (NIR) and Bioptron Pro 1 (HPL), on cancer cells (PC3, MCF7, and HeLa), exploring their effects on the viability and metabolism. Devices specification (Supplementary Tables S1, S2) and relative structural changes (Figs. S1, S2) are reported in detail in the Supplementary Data.
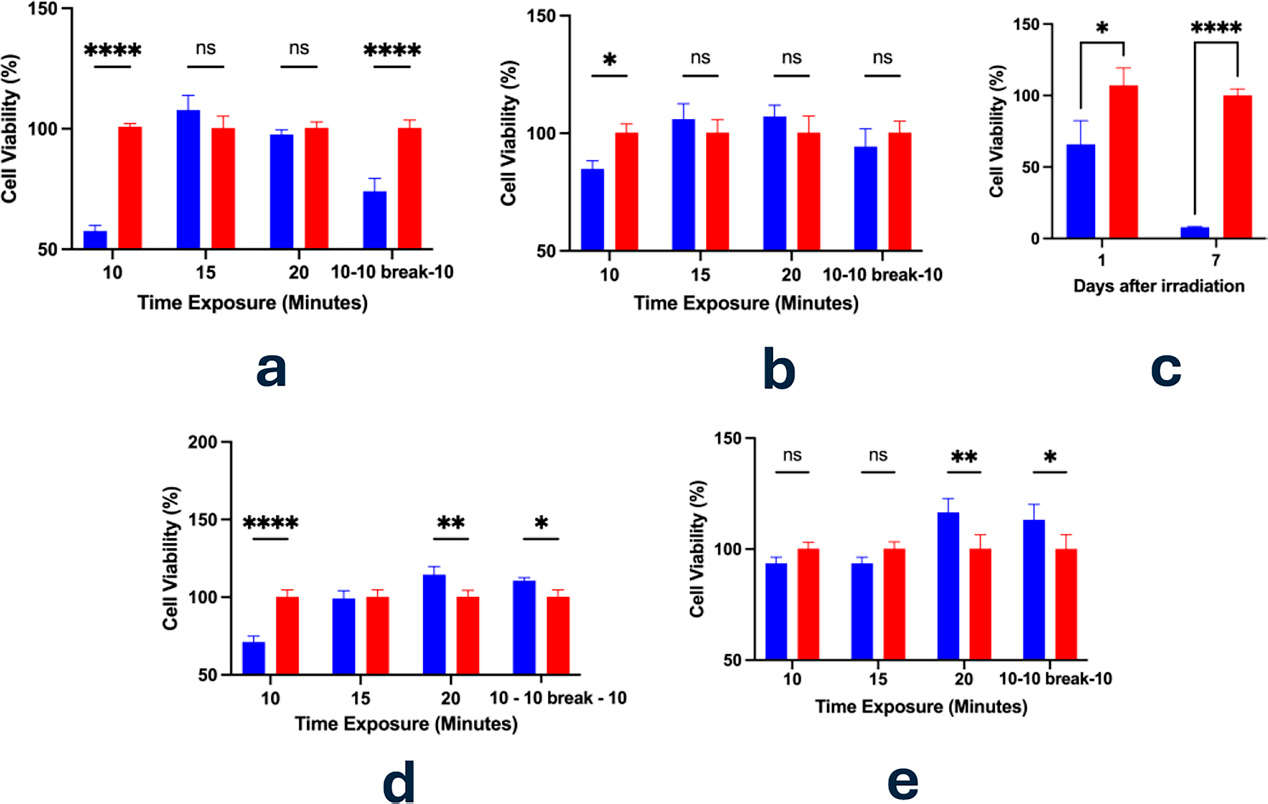
Cellular Viability assessed through Alamar Blue Assay after 24 h from the Bioptron exposure (hyperpolarized light, 40 mW/cm2). 440 µM resazurin was added to cells at a 1:10 dilution and cells were incubated for 2 h. Bar plots represent three experiments reported as mean ± standard deviation. Figure 1a reports the results obtained while testing Bioptron on PC3 cells and medium; Figure 1b shows the results after 24 h, testing solely medium that was subsequentially inserted in the dishes where the PC3 cells were cultured. Figures 1c reports PC3 cellular viability assessment after the first and the seventh day of exposure. In particular, an exposure time of 10 min was tested. Figures 1d and 1e report the impact of the irradiation MCF7 and HeLa cells respectively. Statistic significance is considered as ns (p > 0.05); (*) p ≤ 0.05; (**) p ≤ 0.01; (***) p ≤ 0.001; (****) p < 0.0001. Blue = treated; red = control.
Hyperpolarized light
HPL emitted cells for 10, 15, 20, and 20 min with 10 min of intermission. The 10-min interval showed decreased cellular viability, however significant only on PC3 and MCF7 cells (Fig. 1a—1d). When extending the time exposure to 15 or 20 min, a positive reaction is triggered with a significant increase in the cell viability in the treated cells after 20 min for MCF7 and HeLa (Fig. 1a-d-e). In addition, after the intermittent exposure (20 min of irradiation with 10 min of break in between), only PC3 cells appear to be consistent in the trend, whereas both MCF7 and HeLa cells show promoting proliferation. This suggests that the total dose given is mainly responsible for the cellular response, together with the specific cell-line characteristics.
Plain medium was also exposed to the same conditions tested previously and subsequently placed in the dishes where cells were plated (Fig. 1b). Again, there is a significant decrease in cellular activity after 10 min exposure, which is then increasing over time, even if not significantly. Perhaps, understanding more deeply the level of involvement of water and soluble solutions with respect to cells is essential, in combination or not with the specific wavelength.
Taking into consideration the above-mentioned results, 10 min of exposure was also tested in a longer experiment. PC3 cells were exposed on the first and the sixth days and tested accordingly in terms of viability. We can see that multiple irradiations have significantly decreased cellular metabolic activity (Fig. 1c).
For assessing the metabolic response, ATP has been measured after 10 min of irradiation for PC3 cells. Surprisingly, the repetition of these experiments led to three different results: (a) an initial switch to glycolytic activity with evident damage to mitochondria; (b) an increased mitochondrial activity as well as the production of the glycolytic ATP; and (c) a decrease in glycolytic ATP followed by a 10-fold increase in the mitochondrial energy production (Fig. 2). This suggests a synergistic upregulation of both glycolytic and mitochondrial pathways, potentially reflecting a metabolic adaptation strategy to meet the increased energy demands associated with cellular repair and recovery processes.
Near-infrared pulsating light
The same cancer cell lines have been tested with NIR pulsating light at different intervals. For PC3 cells, 10 and 15-min of irradiation resulted in a decrease in viability compared to non-irradiated controls but only significant in the second case (Fig. 3a). However, extending the irradiation time to 20 min led to a statistically significant reduction in viable cells, suggesting that prolonged light exposure may exert detrimental effects on cell health or survival (Fig. 3a). The 15-min exposure was also applied to PC3 cells on both day 1 and day 6 to assess the longevity of the effect. As shown in Figure 3c, a significant reduction in viability remained evident even 1 week post-treatment, indicating a sustained cellular response.
Regarding the MCF7 and HeLa cells exposure instead, we decided to maintain the time point of 15 min of exposure but to include two extended time points in order to investigate the time dependency. In doing so, we tested 2.07, 4.54, and 8.26 J/cm2. The decrease in viability due to a longer exposure was significantly confirmed in the case of the MCF7 cells (Fig. 3d), but not in the HeLa cells (Fig. 3e). Evidence shows a potential apoptotic effect with an energy dose of about 3 J/cm2.13 However, here the pulsating NIR irradiation had not confirmed the result.
Plain medium was also exposed to the same conditions tested previously (10, 15, 20, and 20 min with 10 min of intermission) and subsequently placed in the dishes where PC3 cells were plated (Fig. 3b). As previously reported, solute–solvent interactions may absorb electromagnetic radiation, potentially reducing EMF’s efficacy. Supporting this, cell viability was significantly higher with medium pre-exposed for 10 min compared to direct irradiation. Moreover, medium exposures of 15 and 20 min showed a time-dependent decline in viability. These findings suggest that the medium may exert a transient screening effect within the first 10 min of NIR exposure (Fig. 3b).
Metabolic assays highlighted a dynamic shift in energy production pathways following Vielight irradiation. Figure 4a shows that there was a decrease in glycolytic ATP, and Figure 4d shows a decrease in lactate levels, suggesting that light exposure may potentially reverse the Warburg effect by promoting a more oxidative phenotype. Indeed, mitochondrial activity appeared to be activated (Fig. 4c), leading to ATP production that was about 5-fold greater than glycolytic ATP.
Limitations
Considering the sample area and the beam spot of the Vielight device, one important limitation of the study resides behind the different uniformity of the irradiation between the two devices. The bystander effect illustrated in Supplementary Figure S3 of the Supplementary Data is assumed 25 in the NIR-treated samples, but a proper setup for a precise quantification is needed to discern between EMF effects and non-EMF-induced effects. Moreover, the same amount of dose should be considered for an accurate comparison.
Conclusion and Future Perspectives
The observed dichotomy in viability patterns across different exposure durations, wavelengths, and cell lines emphasizes the complexity of their mechanisms of action and their interaction with the heterogeneous nature of cancer. Shorter exposure intervals with HPL appeared more effective, whereas the opposite is true for pulsating NIR. Future research should systematically investigate the interplay between irradiation parameters (wavelength, dose, exposure time) and cell line-specific characteristics, including more cell lines, genetic profiles, and metabolic phenotypes. Indeed, the mitochondria activation requires a more extensive assessment to determine the metabolic reprogramming rate in terms of oxygen consumption, transmembrane potential, and electron transport chain. Addressing these areas will deepen our understanding of EMF’s mechanisms and facilitate the development of more effective cancer treatments.
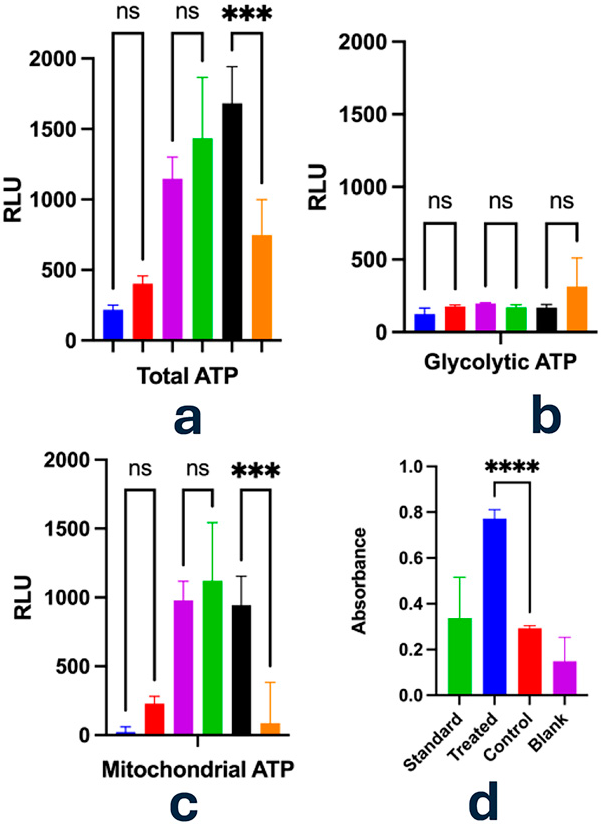
Evaluation of ATP levels and lactate production in PC3 cells following exposure to Bioptron Pro 1 (hyperpolarized light; 40 mW/cm²; 10 min) by Glycolysis/OXPHOS assay. ATP content was measured using a luciferase-based luminescence substrate. Mitochondrial ATP was calculated by subtracting glycolytic ATP from total ATP. Glycolytic ATP was determined after treatment with oligomycin (1.25 μmol/L) following a 3-h incubation at 37°C.
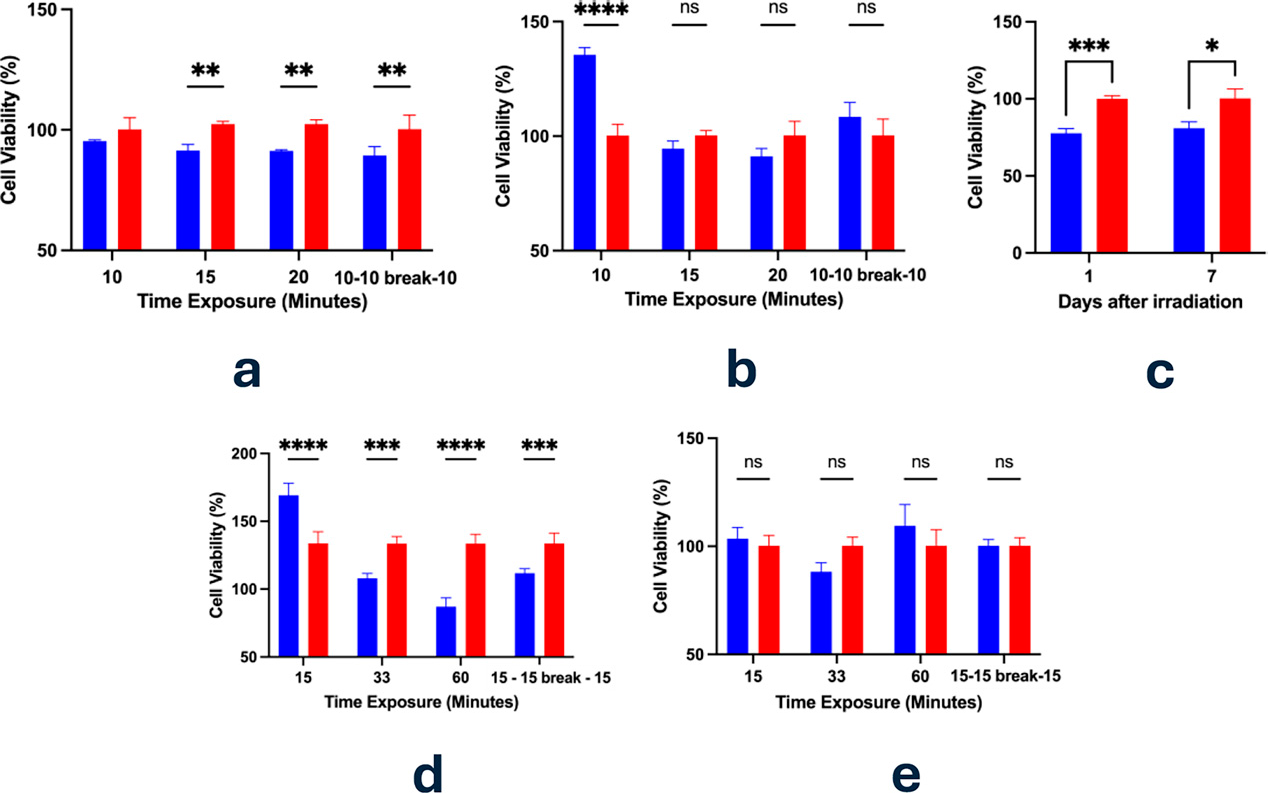
Cellular Viability assessed through Alamar Blue Assay after 24 h from the Vielight exposure (810 nm, 4.59 mW/cm2). 440 µM resazurin was added to cells at a 1:10 dilution and cells were incubated for 2 h. Experiments were run in triplicates and reported as mean ± standard deviation. Figure 3a reports the results on PC3 cells and medium; Figure 3b shows the results on PC3 after 24 h, testing solely medium. Medium was subsequentially inserted in the dishes where the PC3 cells were cultured. Figures 3c reports PC3 cellular viability assessment after the first and the seventh day of exposure. In particular, 15 min of exposure were tested. Figures 3d and 3e report the impact of the irradiation MCF7 and HeLa cells respectively. Statistical significance is considered as ns (p > 0.05); (*) p ≤ 0.05; (**) p ≤ 0.01; (***) p ≤ 0.001; (****) p < 0.0001. Blue = treated; red = control.
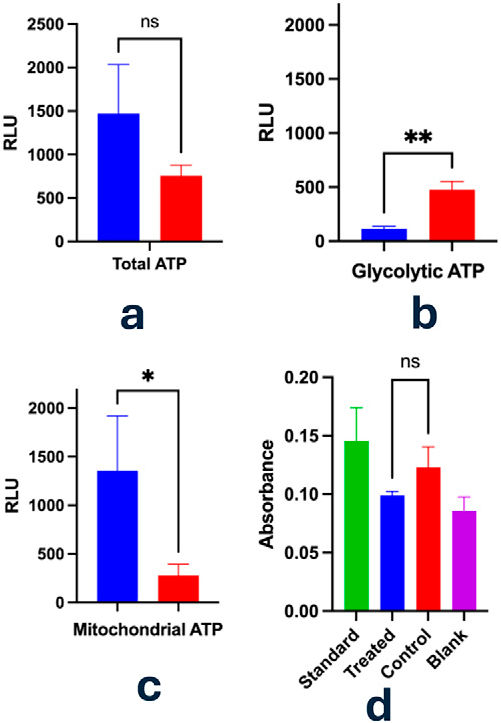
Analysis of ATP generation and lactate output in PC3 cells following treatment with Vielight NeuroPro (810 nm; 4.59 mW/cm² 15 min, 10 Hz sweep) by using Glycolysis/OXPHOS assay. Intracellular ATP concentrations were quantified using a luciferase-based luminescence method. Mitochondrial-derived ATP was estimated by subtracting glycolytic ATP from the total ATP. Glycolytic ATP production was measured after the addition of oligomycin (1.25 μmol/L) post a 3-h incubation at 37°C. Panel
Footnotes
Authors’ Contributions
J.A.T., D.B., and B.T. designed the study and the experiments. B.T. and S.C. ran the experiments, collected the data provided, and performed the formal analysis. J.A.T. supervised the entire study. All the authors reviewed the article and collaborated in the writing stage.
Availability of Data and Materials
The data obtained and analyzed in the current study are available from the corresponding author on reasonable request.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
No funding was received for this article.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.