
Abstract
Background:
Cannabinoid-based medicines (CBMs) have garnered attention due to their anti-inflammatory potential in people with HIV (PWH), whose comorbidities are driven by chronic inflammation. The expanded endocannabinoid system (or endocannabinoidome, eCBome) is an important target of cannabinoids that cross talks with gut microbiota and regulates many homeostatic processes and inflammation. In a prospective, pilot clinical trial, PWH on antiretroviral therapy (ART) were randomly assigned to receive cannabidiol (CBD) ± Δ9-tetrahydrocannabinol (THC) capsules for 12 weeks, titrating doses as tolerated, to examine the impact of cannabinoids on plasma eCBome mediators and gut microbiota.
Methods:
Ten individuals were randomized, five to the CBD+THC arm and five to the CBD-only arm. Eight individuals completed the study. Plasma was collected at each visit and measured in batches by liquid chromatography (LC)-mass spectrophotometry (MS). The eCBome mediators were measured at each visit by LC-MS-MS, whereby fecal microbiota composition was assessed by 16S rDNA sequencing at the initiation and end of treatment.
Results:
Plasma concentrations of THC and CBD metabolites varied throughout the course of the study. Capsule administration resulted in a significant decrease in monoacylglycerols 2-eicosapentaenoylglycerol (2-EPG) and 2-oleoylglycerol (2-OG) after treatment. No changes were observed in the levels of other mediators measured. PWH in the distinct treatment arms had different fecal bacterial taxa at baseline. These differences persisted through the course of the study and were not altered by cannabinoid administration. However, Coprobacillus and Lachnospiraceae UCG001 relative abundance was lower, while Collinsella was higher, in the THC/CBD compared with the CBD arm.
Conclusion:
2-EPG and 2-OG were both reduced following cannabinoid administration. No changes in fecal bacterial taxa were observed following 12 weeks of treatment. Larger studies are needed to understand if these changes reflect adaptation of the eCBome to the beneficial effects of CBM in PWH.
Trial Registration:
ClinicalTrials.gov Identifier NCT0355035.
Keywords
Introduction
HIV is characterized by reduced gut barrier integrity, intestinal damage, microbial translocation, 1 and gut dysbiosis that contribute to a proinflammatory state and a higher burden of age-related comorbidities compared with HIV-negative individuals.2,3 Therefore, many approaches to reduce the comorbidity gap between those people with and without HIV infection are based on reducing chronic systemic inflammation.4–7
Cannabinoid-based medicines (CBMs) are widely used by people with HIV (PWH) and are of interest due to their potential to reduce inflammation.5,8 Cannabinoids impact physiological processes mostly by modulating the expanded endocannabinoid (eCB) system or “endocannabinoidome” (eCBome). 9 This system consists of the G protein-coupled receptors (GPCRs), cannabinoid receptor type 1 (CB1R) and 2 (CB2R), and their endogenous ligands. It also consists of the endocannabinoids anandamide (N-arachidonoylethanolamine; AEA) and 2-arachidonoylglycerol (2-AG)10–12 and eCB-like lipid congeners from, among others, the N-acylethanolamine (NAE) and the monoacylglycerol (MAG) families, their GPCRs, as well as nuclear receptors and ligand-activated ion channels.10–12 AEA, 2-AG, and their congeners exert principally prohomeostatic properties.11–18 The eCBome plays a role in several biological functions, including inflammation, lipid metabolism, and gut permeability,19,20 among others.11,12,16,18,21,22
PWH have a high abundance of specific proinflammatory bacteria, such as Catenibacterium and Prevotella, which correlates with accelerated biological aging. 3 Moreover, PWH have a lower intestinal abundance of bacteria producing anti-inflammatory short-chain fatty acids, such as Subdoligranulum and Erysipelotrichaceae, which are associated with slower biological aging. 3 Correlation networks reveal significant relationships among specific microbial genera in the colon and ileum and increased aging; a rise in proinflammatory microbe-related metabolites, such as those involved in tryptophan metabolism; and a decrease in anti-inflammatory metabolites such as hippuric acid. 3 The gut microbiota may also contribute to differential immune responses to antiretroviral therapy (ART). 23
eCBome mediators and receptors impact microbiota composition, and various bacteria and fungi, in turn, impact the eCBome.20,24,25 In participants in the Canadian HIV and Aging Study (CHACS) cardiovascular substudy, our team showed that PWH with preclinical atherosclerosis had altered eCBome profiles, including reduced levels of AEA and some of its NAE congeners, and increased levels of MAGs. There were also alterations in several fecal bacterial taxa, compared with HIV-negative controls. 26 Thus, HIV infection may modulate plasma eCBome mediator metabolism in PWH. Although one study reported that cannabis use did not impact eCBome lipid levels in PWH, 27 the interplay between gut microbiota and circulating eCBome mediators in the context of cannabidiol (CBD) and Δ9-tetrahydrocannabinol (THC) administration has not been examined.
Insight into alterations of circulating eCBome mediators and gut bacteria in PWH receiving CBM may offer new avenues for patient management. Alterations may either mediate some of the effects of CBM or be explained as an adaptive mechanism following CBM’s beneficial effects. We showed that oral cannabinoid administration with ART may result in reduced plasma levels of inflammatory markers and cellular inflammation in PWH. 28 In the current study, we examined the dynamics of plasma levels of eCBs and their congeners, along with gut microbiota composition, in PWH randomized to receive oral capsules containing THC/CBD or CBD alone for 12 weeks in the CTN PT028 pilot clinical trial.
Methods
Blood and stool sampling and processing
At enrollment, fasting blood was collected by venipuncture to measure circulating soluble markers, including eCBs. Plasma was isolated via centrifugation and stored at –80°C to be analyzed in one batch at study end. Participants also provided a stool specimen that was immediately aliquoted and stored at –80°C until batch analysis.
11-OH-THC, 11-COOH-THC, 7-OH-CBD, and 7-COOH-CBD in human plasma
Measures were performed by the Pharmacokinetics core facility of the Centre de Recherche, Centre hospitalier de l’Université de Montréal by mass spectrophotometry.
Microbiota characterization
DNA was extracted from fecal samples and used to produce 16S rDNA (V3-V4) libraries for sequencing on an Illumina MiSeq System as previously described. DNA was extracted from feces using the QIAmp PowerFecal DNA Kit (Qiagen, Hilden, Germany) according to the manufacturer’s instructions, and concentrations were fluorometrically quantified with the Quant-iT PicoGreen dsDNA Kit (Thermo Fisher Scientific, MA, USA) and used to produce sequencing libraries according to the Illumina 16S ribosomal RNA gene V3-V4 region amplicon preparation protocol for the Illumina MiSeq System using primer pairs F (5′-TCGTCGGCAGCGTCAGATGTGTATAAGAGACAGCCTACGGGNGGCWGCAG-3′) and R (5′- GTCTCGTGGGCTCGGAGATGTGTATAAGAGACAGGACTACHVGGGTATCTAATCC-3′) in conjunction with the QIAseq16S/ITS 384-Index I (Sets A, B, C, D) Kit (Qiagen, Hilden, Germany). The 16S metagenomic libraries were eluted in 30 µL of nuclease-free water, and 1 µL was qualified with a Bioanalyzer DNA 1000 Chip (Agilent, CA, USA) to verify the amplicon size (expected size ∼600 bp) and quantified with a Qubit (Thermo Fisher Scientific, MA, USA). Libraries were then normalized and pooled to 2 nM, denatured, and diluted to a final concentration of 8 pM. Sequencing (2 × 300 bp paired-end) was performed using the MiSeq Reagent KitV3 (600 cycles) on an Illumina MiSeq System. Sequencing reads were generated in less than 65 h. Image analysis and base calling were carried out directly on the MiSeq. Data were processed using the DADA2 pipeline according to the recommended workflow, 29 and ASV sequences were assigned taxonomy using the most recent SILVA taxonomic database (SILVA SSURef 138.1 NR, March 2021) as a reference dataset using default parameters. 30 To deal with differences in sampling depth, the data were rarefied to a depth of 18,000 reads before data filtering (10% of features with the lowest variance based on interquartile range), analysis, and figure production utilizing the Microbiome Analyst web-based platform. 31
Statistical analysis
Refer to Supplementary Data S1.
Results
Participants
Among the eight individuals who completed the study, the median age was 57.5 [interquartile range (IQR) = 55.5, 62) years, the median time from HIV diagnosis was 21.5 (IQR = 15, 25) years, six individuals (75%) were male, and six individuals (75%) were Caucasian. Median CD4 count at study enrollment was 439 cells/mm3 (IQR = 419, 559), and all were virally suppressed on ART. 32
Impact of CBD ± THC capsules on THC, CBD, and metabolites
Plasma levels of metabolites over the course of study visits are depicted in Supplementary Table S1 and Supplementary Figure S1. Overall, there was significant variability within and between participants in concentrations of cannabinoid metabolites from visits 2 to 10. Interestingly, we found that some participants in the CBD-only arm had THC-COOH detectable in their plasma. Potential reasons include exposure to secondhand smoke. Under acidic conditions, CBD can turn into THC, and it has been suggested that gastric fluid can transform CBD into THC. 33 However, that this conversion may actually occur in vivo has been recently deemed very unlikely. 34 All participants indicated they only consumed cannabinoids as provided for use in the study. However, the possibility that participants consumed exogenous products without disclosure cannot be entirely excluded.
Impact of CBD ± THC capsules on endocannabinoid levels
eCBs and eCB-like lipid levels in participants along the course of treatment are depicted in Table 1. Overall, between visit 2 (week 0) and the treatment interruption at visit 9 (week 12), there was a trend towards decreased levels of several MAG levels in study participants following CBM treatment (Table 1). Notably, the decreased trend at the end of treatment (visit 9) was close to significant for plasma levels of 2-eicosapentaenoylglycerol (2-EPG) and 2-oleoylglycerol (2-OG) (Fig. 1).
Endocannabinoids and Endocannabinoid-Like Lipid Levels in Clinical Trial Participants Along the Course of Treatment

AEA, N-arachidonoyl-ethanolamine; EPEA, N-eicosapentaenoyl-ethanolamine; LEA, N-linoleoyl-ethanolamine; DHEA, N-docosahexaenoyl-ethanolamine; PEA, N-palmitoyl ethanolamine; OEA, N-oleoyl-ethanolamine; stearoyl-EA, N-stearidonoyl-ethanolamine; DPA-EA(n-6), N-docosapentaenoyl-ethanolamine n-6; DPA-EA(n-3), N-docosapentaenoyl-ethanolamine n-3; 2-AG, 2-arachidonoyl-glycerol; 2-EPG, 2-eicosapentaenoyl-glycerol; 2-DHG, 2-docosahexaenoyl-glycerol; 2-LG, 2-linoleoyl-glycerol; 2-DPG, 2-docosapentaenoyl-glycerol n-3; 2-OG, 2-oleoyl-glycerol; 2-SDG, 2-stearidonoyl-glycerol; SDA, stearidonic acid; EPA, eicosapentaenoic acid; DHA, docosahexaenoic acid; AA, arachidonic acid; DPA-w3, docosapentaenoic acid n-3; DPA-w6, docosapentaenoic acid n-6; LA, linoleic acid; PUFA, polyunsaturated fatty acids; IQR, interquartile range.
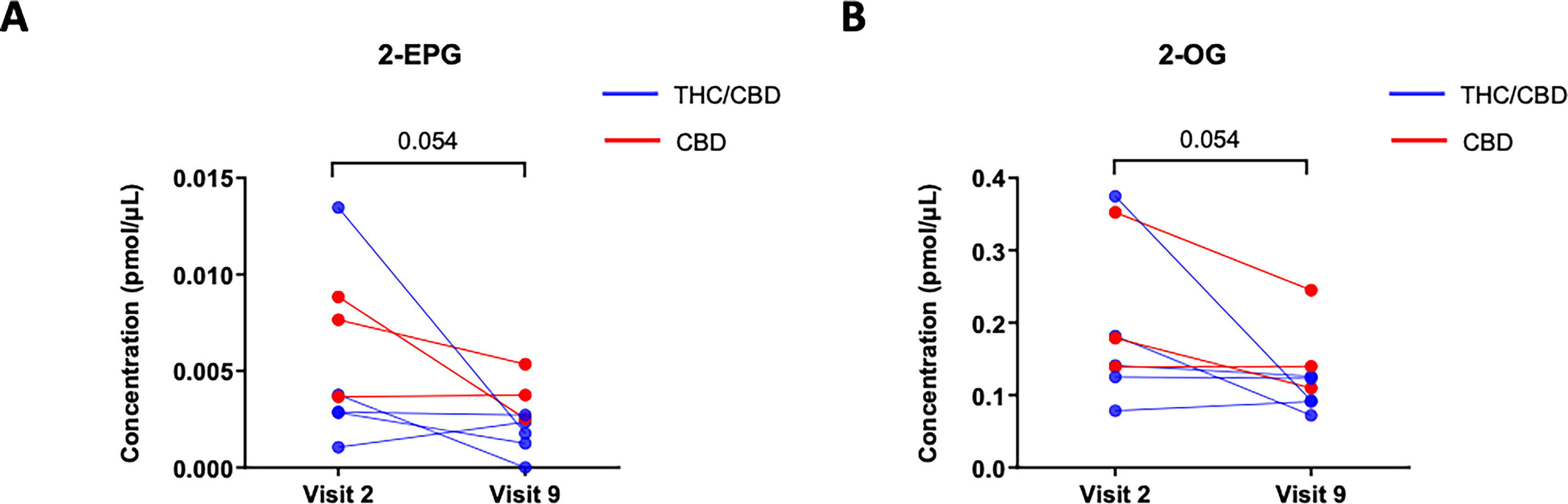
Decrease of monoacylglycerol (MAG) levels on study participants after treatment.
Impact of CBD ± THC capsules on gut microbiome
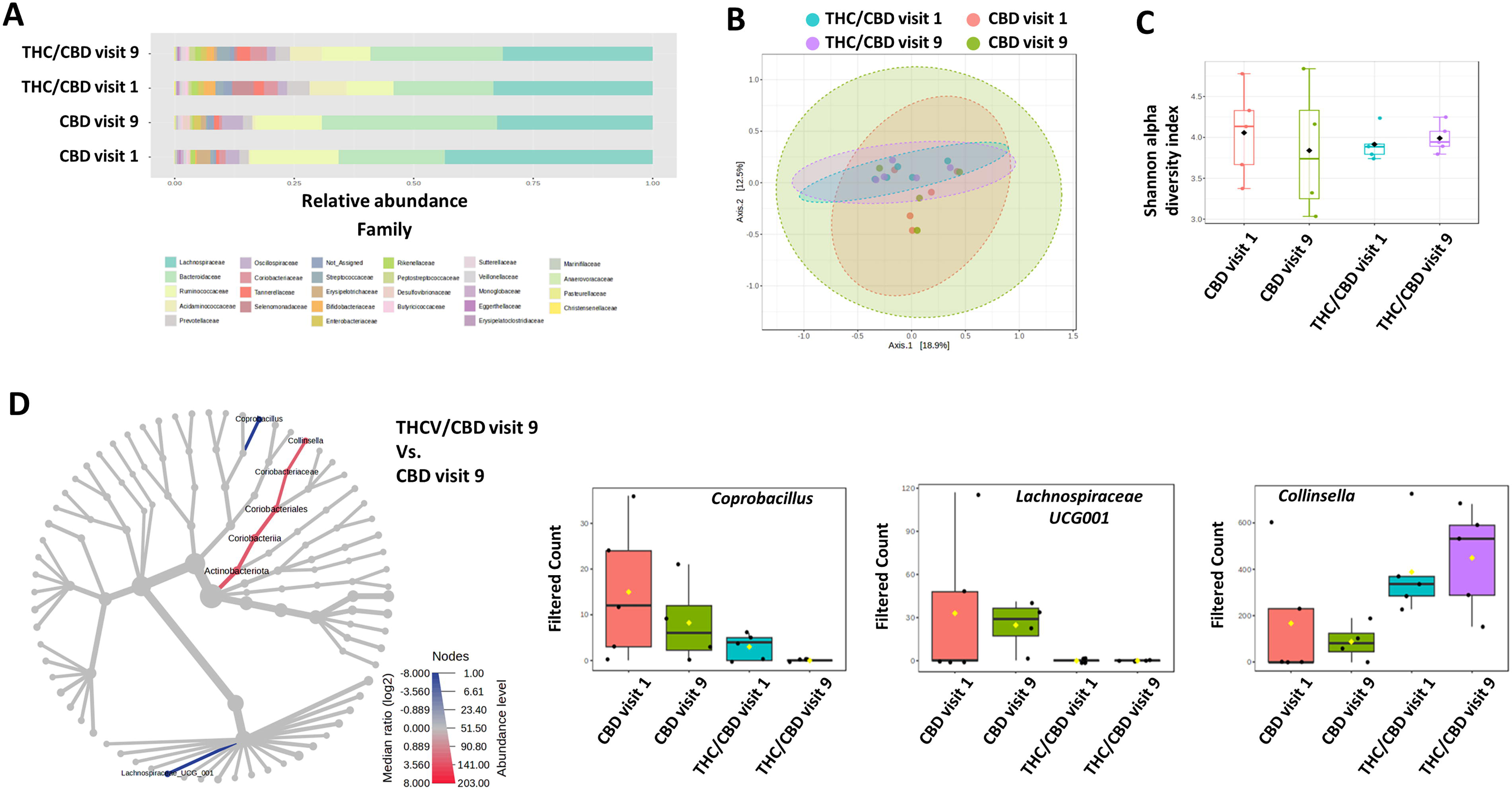
Bacterial families with the highest relative abundances across all groups were Lachnospiraceae, Bacteroidaceae, and Ruminococcaceae, with Acideaminococcacocase also being relatively abundant within the THC/CBD group (Fig. 2A). Taxonomic composition of stool community, at the family level, did not differ significantly between visits 1 and 9. This is supported by the principal coordinate analysis to assess beta diversity, which shows that neither the THC/CBD nor the CBD treatment altered the overall community architecture and that while diversity of the THC/CBD and CBD arms separate slightly from each other, PERMANOVA identified no difference between the groups (F value: 0.83236; R2: 0.14271; p value: 0.82; Fig. 2B). Furthermore, as shown in Figure 2C, Shannon’s alpha-diversity was not different between the arms and did not change significantly within each treatment arm between visits 1 and 9 (statistical significance: p value: 0.92; [ANOVA] F value: 0.17). We then assessed taxonomic differences between microbial communities of the THC/CBD and CBD arms at visit 9 using “heat tree analysis.” 35 This did not reveal any differences in abundance between the THC/CBD and CBD arms at week 1 or any effect of THC/CBD or CBD over time, that is, week 1 versus week 9 (data not shown). However, when comparing the THC/CBD and CBD arms at week 9, the analysis identified that Coprobacillus and Lachnospiraceae UCG001 abundance was lower in the THC/CBD arm, while Collinsella was higher (Fig. 2D, left). While the subsequent Kruskal–Wallis analysis did not identify any statistical differences in genera between the groups, there were trends for decreased Coprobacillus and increased Collinsella abundance in both the THC/CBD and CBD arms from visits 1 to 9, while Lachnospiraceae UCG001 abundance was unaltered (Fig. 2D, right).
Discussion
We examined levels of plasma THC and CBD metabolites and endocannabinoids, along with changes in gut microbiota composition, in PWH on ART at baseline and following 12 weeks of CBM. First, we found that concentrations of THC and CBD metabolites varied throughout the study course. Consumption of relatively low doses of cannabinoids, coupled with low bioavailability of capsules, may explain the relative lack of impact on the microbiome parameters examined. In some cases, individuals only took a single capsule of CBM at night for tolerability reasons, and these doses may not have been high enough to result in detectable metabolites. Bioavailability of oral THC and CBD is relatively low. 36 THC’s oral bioavailability is 6% versus smoked and inhaled bioavailability, which is 25% and 10–35%, respectively.36,37 CBD also has poor bioavailability due to first-pass metabolism and erratic absorption, in contrast with intranasal CBD, which has a bioavailability of 34–46%.37–39 Furthermore, research study visits, and hence blood draws, occurred at different times of day based on participant and study staff availability.
Second, we found that capsule administration resulted in a significant decrease in two MAGs, that is, 2-EPG and 2-OG. No changes were observed in other eCBome mediators. This finding was consistent with a published study showing that a dose of 800 mg of CBD increased AEA and other NAE levels in less than 3 h (without affecting 2-AG), but no effects were observed at the lower dose of 600 mg of CBD or 20 mg of THC. 26 The reduction in MAGs after treatment may be considered potentially protective, when these act on targets that exacerbate inflammation, for example, coronary artery disease and obesity.40–43 Alternatively, the reduction in MAGs may represent an adaptation to the beneficial effects of CBM, when these mediators instead are elevated to counteract the condition being treated. In particular, 2-OG has recently been found to contribute to the development of metabolic dysfunction-associated steatohepatitis in conjunction with, and as a result of, changes in the gut microbiome,44,45 which has a high prevalence in PWH. 46 Accordingly, in the CHACS, which involved individuals 40 years and older with well-controlled HIV on ART and duration of HIV infection for at least 15 years, we documented a relationship between increased MAGs and plasma triglycerides. 26 We also found that an inverse relationship between NAEs and MAGs may be predictive of subclinical coronary artery disease in this population. 26 In another observational study, altered eCBome profile and elevated levels of inflammatory mediators, concomitant with reduced levels of AEA and some of its NAE congeners, were impacted by HIV status. 27 However, 2-AG levels were not impacted by HIV status. 27 These findings suggest that HIV has a differential impact on circulating eCBome mediator metabolism in PWH. 27 Our data are in line with the observed decreases in plasma levels of 2-OG (at least in the liver through hepatic stellate cell activation). 45 It should be noted, however, that when acting in the gut on the same receptor (GPR119), 2-OG appears to be metabolically beneficial via increased GLP1 production. The molecular target of 2-EPG is not known. However, NAEs and MAGs derived from omega-3 polyunsaturated fatty acids usually play anti-inflammatory actions, as in the case of N-eicosapentaenoyl and N-docosahexaenoylethanolamine.47,48 2-EPG has been associated with beneficial eating habits in healthy individuals. 49
Our previous data on the same study participants showed that oral cannabinoids also reduced inflammatory soluble markers IFN-γ, IL-1β, sTNFRII, and gut mucosal damage marker REG-3α, in addition to decreased cellular inflammation. 28 Although our findings are promising, the results need to be interpreted with caution given the small sample size. Furthermore, it is likely that any intervention that modulates the ECS through reduction of MAGs would likely need to be taken for many years before having an impact on risk reduction of clinical outcomes such as coronary events. 50
We documented heterogeneity in microbiome composition across participants at baseline. Heterogeneity in microbiome composition persisted throughout the study and was not impacted by CBM capsules in either arm. The lack of impact may be related to the relatively small doses of cannabinoids consumed. We also observed that bacterial families with the highest relative abundances were Lachnospiraceae, Bacteroidaceae, Ruminococcaceae, and Acidaminococcaceae (in the THC/CBD arm), which are recognized as having high relative abundance in the gut. 51 These families are all main producers of short-chain fatty acids, which, while generally considered as beneficial mediators of inflammation and metabolism, have an impact on host physiology that is not always consistent across studies,51,52 and we did not detect significant changes in their relative abundances here. 52
In immune responders and immune nonresponders (defined as persons with CD4+ T counts ≥500 and <200 CD4+ T cell counts/µL after 2 years of HIV-1 viral suppression, respectively), the taxa Faecalibacterium and Ruminococcaceae were depleted versus healthy controls. 23 Faecalibacterium was positively correlated with CD4/CD8 ratio and negatively correlated with the inflammation markers and Lipopolysaccharide (LPS), while Ruminococcaceae was positively correlated with the current nadir CD4+ T cell counts and negatively correlated with the CD8+ and CD57+ T cell counts. 23 In our previous work with the cohort studied here, we reported a transient decrease in the frequencies of CD28−CD57+ senescent CD4 and CD8 T cells between weeks 1 and 8. 28 This is noteworthy given the interest of CBM as a potential strategy to mitigate processes associated with aging,53–55 although we could not associate these changes with alterations in either Faecalibacterium or Ruminococcaceae. However, gut microbiota dysbiosis may contribute to different immune responses and treatment outcomes to ART, with Fusobacterium, Alistipes, Ruminococcaceae, Faecalibacterium, Escherichia-Shigella, Roseburia, and Blautia having been found to be potential genera contributing to these different responses. 23 In our study, most individuals had CD4 T cell counts above 500 cells/mm3, and none had CD4 T cells below 200 cells/mm3, which is a threshold sometimes used to define an immune nonresponse.
Our study has important limitations. Notably, the small sample size, single study site, and a predominance of White male participants preclude us from generalizing our results to other populations. The older age of our participants (55–62 years) suggests the presence of some gut dysbiosis, with a decrease in immune-supportive bacteria and a lowered gut integrity at baseline.56–58 Although our participants consumed phytocannabinoids with over 98% purity in oil, 32 studies on cannabinoids should be cognizant of potential impact of the “entourage” effect, whereby complementary or synergistic effects of other medicinal cannabis constituents, particularly terpenes, may occur concerning their therapeutic potential. 59 Doses of cannabinoids taken by participants in our study (Supplementary Table S1) also varied throughout the course of the study, as participants titrated the doses consumed based on their tolerability. 60
Gut microbiota varies among individuals across different geographical locations globally. In a recent study, the geographical location had a greater effect than HIV infection status, regardless of whether the individuals were on ART. 61 By directly comparing large cohorts from the United States, Botswana, and Uganda, Rocafort et al. found few common bacterial changes with HIV infection between individuals in these regions, and no HIV-associated bacterial changes were shared among all three geographic cohorts. 61 Gut microbiota is also influenced by host genetics and exercise and has notable interpersonal variations. 62 Furthermore, polymorphisms in genes for cannabinoid receptors, enzymes, and transporters impact pharmacokinetics and therapeutic effects. 39 Recent studies indicate gender-specific differences in immunity, with the gut microbiota shaping and being concomitantly shaped by the hormonal milieu governing differences between the sexes. 63
We did not collect dietary information, and thus, given the ability of gut microbiota and lipid mediators to respond rapidly to dietary alterations, 64 we cannot discount the possibility that alterations observed here were not influenced by the participants’ food choices before fecal and plasma sample collection. Although most participants were men who have sex with men, the sample size was likely too small to see differences in the microbiota, as receptive anal intercourse can affect the composition of the microbiome, especially with regard to Prevotella species.65,66
Conclusions
Preclinical data suggest that cannabinoids may play a crucial role in reestablishing the balance and symbiosis between gut microbiota and the immune system, restoring homeostasis and reducing inflammaging.24,25 However, due to safety and tolerability considerations, dosages of CBD ± THC that could be administered may have prevented us from observing potentially beneficial impacts. We found a reduction in some MAGs that could potentially reduce the risk associated with premature CAD but treatment did not impact on the microbiome. Our study raises important considerations for the design of future studies where a clinical outcome is the primary end-point. Furthermore, the duration of time participants would need to continue treatment to impact CAD progression is likely several years. Future studies using oral cannabinoid capsules to examine impact on the eCBome and gut microbiome composition should ensure that doses given to participants are both safe and well-tolerated and that doses are sufficiently high to induce noticeable changes in microbiome composition.
Authors’ Contributions
Conceptualization and study design: C.T.C. and M.A.J. Participant recruitment and data acquisition: C.T.C. and J.P.R. Methodology: G.G., C.M., H.K., C.T.C., M.A.J., N.F., C.S., and V.D. Validation: G.G., C.M., H.K., C.T.C., M.A.J., N.F., C.S., and V.D. Formal analysis: R.S.M.B., K.F.B., G.G., C.M., H.K., M.A.J., and C.T.C. Investigation: R.S.M.B., K.F.B., G.G., C.M., H.K., C.T.C., M.A.J., N.F., C.S., and V.D. Resources: C.T.C., M.A.J., N.F., C.S., and V.D. Data curation: C.S., G.G., S.M., K.F.B., M.A.J., and C.T.C. Writing—original draft preparation: C.T.C. Writing—review and editing: All authors. Supervision: C.T.C., M.A.J., N.F., C.S., and V.D. Project administration: C.T.C. and M.A.J. Funding acquisition: C.T.C. and M.A.J. Article approval: All authors.
Footnotes
Acknowledgments
The authors thank the participants for their participation in this study. The authors also thank the staff at the CTN, including Judy Needham (study manager), Nisha Shewaramani and Elisa Lau (database managers), Jayamarx Jayaraman (study monitoring), and Alison Müller (communications), and all the CVIS/MUHC research and clinical staff, especially Suzanne Samarani, Florien Beaubien and Claude Vertzagias. The authors are also grateful to the Tilray staff, including Philippe Lucas, Catherine Jacobson, and Gosia Grzyb, as well as Dr. Carolyn Baglole and the Research Centre for Cannabis at McGill University. The authors also thank Ms. Fleur Gaudette, manager of the Plateforme de pharmacogénétique at the CR-CHUM, for overseeing measurements of the cannabinoid metabolites.
Author Disclosure Statement
C.T.C. has served on advisory boards for ViiV Healthcare and Gilead and received grant support from Merck, Gilead, ViiV, and Tilray, Inc. She has also received travel support to attend conferences from Gilead and ViiV Healthcare.
Funding Information
This work has been supported by a team grant from the Canadian Institutes of Health Research (CIHR) (# CC1-177334), CIHR Canadian HIV Trials Network CTN PT028 study, and the Lotte & John Hecht Memorial Foundation. Tilray provided study capsules as an in-kind service but did not provide study funding. R.S.M.B. is supported by a CIHR postdoctoral fellowship. J.-P.R. holds the McGill University Louis Lowenstein Chair in Hematology and Oncology. M.A.J. holds the tier 2 CIHR Canada Research Chair in Immuno-Virology. C.T.C. is a recipient of an FRQ-S Senior Chercheur Boursier Clinicien career award. V.D. holds the Canada Excellence Research Chair on the Microbiome-Endocannabinoidome Axis in Metabolic Health, which is funded by the Federal Tri-Agency.
Presentations
This work has been presented at the 34th Canadian Conference on HIV/AIDS Research (CAHR 2025) in May 1–4, 2025, at Halifax, Nova Scotia, Canada.
Supplemental Material
Supplemental Material
Abbreviations
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.