
Abstract
This article examines the effects of biotic and abiotic influences on trichome development in cannabis and how these influences may shape the ultimate cannabinoid and terpene profiles in cannabis cultivars. As cannabis agriculture focuses primarily on either high tetrahydrocannabinol or cannabidiol production, the result has been an attenuation of genetic and chemical diversity observed in contemporary cannabis cultivars. Techniques for modulating trichome development remain an active area of commercial importance. The clinically therapeutic role of varied cannabis metabolite profiles, which include the wider range of cannabinoids and terpenes, also ultimately tied to trichome development. This review will analyze how different stressors can modify phytochemical ratios and alter trichome development.
Keywords
Introduction
In this review, we explore various biotic and abiotic factors influencing the phytochemical profiles of Cannabis sativa L., using the levels of cannabinoids and terpenes found in the floral trichomes as fulcrum for analysis. Comparative analysis of mechanisms identified in other medicinal plants is crucial as it allows us to apply existing literature in better researched species to contextualize salient questions regarding similar mechanisms in cannabis, which are currently less well characterized. By examining the influence of environmental factors on trichome development and secondary metabolite production in other species, we hypothesize potential parallels in cannabis. This framework aligns with the experimental techniques that underpin the development of most existing cultivation strategies, and, while focused on cannabis, references to other species with similar trichome and hormonal responses are essential for developing a model of how biotic and abiotic stresses account for trichome variation and potential phytochemical variability in C. sativa.
C. sativa is a flowering plant in the family Cannabinaceae. The plant has long been used medicinally and for fiber production. 1 C. sativa is an annual, wind pollinated, dioecious, flowering herb. It possesses a laterally branched taproot (often underdeveloped on clonally propagated specimens). The plant’s stems are erect (ridged) furrowed and hollow. The plant’s height varies from 0.2 to 6 m (usually 1–3 m) tall. C. sativa leaves are palmately compound with serrated, lanceolate leaflets. They usually have an odd number of leaflets (3–13), with the upper leaves being smaller and simpler than the lower ones. The leaves are oppositely arranged on the stem during the early growth stage, transitioning to an alternate arrangement as the plant matures (Fig. 1). The yellow to light green staminate (male) inflorescences are small (2–4 mm), axillary or terminal, erect, appearing in a lax panicle or a compound cyme, and lack trichomes.

Variations in Cannabis sativa (Cannadoc NB130, NB 140, NB150).
Pistillate (female) inflorescences exhibit a wide range of coloration from green to purple to red and/or mottled or streaked, sessile. Bract proximal upper surfaces are densely covered by capitate stalked trichomes, with serrate or entire margins enveloped in bracts and covered with resinous glandular trichomes (Fig. 2). Cannabis achenes (fruits) are solitary and usually green–brown but also white or gray. Several different types of trichomes exist on the cannabis plant, with the glandular trichomes of upper leaves and unfertilized female inflorescences the primary source of cannabinoids storage. 2 Sessile, glandular, and stalked trichomes are present on bracts and bracteoles surrounding female flowers (Fig. 2). The presence of nonglandular trichomes on the upper leaf surfaces and slender noncystolithic trichomes on both upper and lower surfaces is characteristic of C. sativa. 3 C. sativa is diploid, with 10 pairs of chromosomes (2n = 20) and is characterized by an XY/XX chromosomal sex-determining system, with a genome size of about 830 Mb. 4

Closeup of cannabis female inflorescence. Permission to reproduce material from other sources. Permission by author. Upton, R. (2014). Cannabis Inflorescence: Cannabis spp.:Standards of identity, analysis, and quality control. American Herbal Pharmacopoeia.
The systematization of cannabis has been controversial. Historical representation of a formal taxonomy of C. sativa and C. indica Lamarck has, in modern times, been conflated with vernacular descriptions in the modern marketplace. Ernest Small and Arthur Cronquist 5 proposed only one cannabis species, C. sativa, which is divided into subspecies sativa (fiber type) and indica (drug type). Furthermore, the subspecies ruderalis has been proposed to include plants that flower independent of photoperiod, but this trait is consistent with the escape and subsequent intermingling of cultivated fiber-type cannabis. 6 The result of attempting to reconcile the vernacular and formal nomenclatures results in the observation that ‘‘Sativa’’ is most likely indica, ‘‘Indica’’ is “Afghanica,” and ‘‘Ruderalis’’ is usually sativa. All three are considered varieties of one species, C. sativa. 6
Earlier phytochemical and genetic research supported the taxonomic separation of traditional landraces “Sativa” and “Indica,” but distinguishing between them has been characterized by modern taxonomic researchers as an “exercise in futility.” 6 Thus, contemporary attempts at speciation have become impossible because of extensive crossbreeding. Furthermore, the traditional landraces of “Sativa” and “Indica” are becoming extinct through introgressive hybridization. 6 Modern genetic sequencing has identified four distinct genetic groups: basal (feral) cannabis, hemp-type, drug-type feral, and drug-type cultivated.4,7 Genomic and molecular analyses increasingly support the hypothesis that these genetic groups are best characterized as diverse monotypic species. 8
When subjected to modern phytochemical analysis, sativa and indica secondary metabolite profiles did not differ in their average content of the major cannabinoids. Slight cannabinoid differences between sativa and indica have been noted, such as the content of CBC and Cannabigerol (CBG) being higher in the sativa group and significant differences in terpene composition between them. 9 It is likely that anthropogenic selection and extensive hybridization have led to a coevolution of plant morphology and terpene profile given the role of those compounds defining the sensory signature. 9 Vernacular labels do not accurately reflect genetic relatedness and specimens with the same vernacular name are often as genetically and chemically distant from each other as pairs of specimens with different vernacular names. 10 A more nuanced technique for discussing variation within C. sativa is chemotype, which can be defined as infraspecific variation in secondary metabolite profiles where the constituents of note come from the same biosynthetic pathway in all chemotypes, and these varying constituent profiles are dependent on several genetic and environmental factors.11,12
The two main cannabinoids found in the plant, cannabidiol acid (CBDA) and trans-Δ9-tetrahydrocannabinol acid (THCA) are produced via CBDA or THCA synthase encoded in the genes CBDAS and THCAS that compete for the same substrate, cannabigerolic acid (CBGA). Analysis indicates that both synthases exist in ancestral and feral type samples, whereas the drug-type genetic grouping has a complete coding sequence for THCAS and two CBDAS pseudogenes, and the hemp-type grouping possessing a complete sequence for CBDAS, with some exceptions. 4 One theory is that the loss of full THCAS in hemp-type cultivars and CBDAS in drug-type cultivars occurred during domestication due to strong selection for crop phenotypes. 4 Two models of that theory have been tested as an explanation for varying trans-Δ9-tetrahydrocannabinol (THC): cannabidiol (CBD) ratios with neither providing fundamental support for the theory—one where THCA gene copies influence the ratio,13,14 and a second proposing nonfunctional CBDAS alleles in the homozygous state alter the ratio toward Δ9-THC. 15
The synthesis of cannabinoids takes place largely in the trichome structure of the flowers. Researchers profiled 14 cannabinoids and 47 terpenoids in cannabis flowers, leaves, stem barks, and roots in three chemovars reporting 15–20% cannabinoids and 1–2% terpenoids in the flowers, 1–2% cannabinoids and <0.3% terpenoids in the leaf, and none of either in stem, bark, or root. 16 This suggests that trichomes are useful for detecting biotic and abiotic influences on their secondary metabolite production in varied ecosystems.
Clarke and Merlin 17 reported that the original habitats for wild-type cannabis chemotypes are exposed riverbanks, freshwater shorelines, and the border regions between forests and cultivated or naturally disturbed land, which may be further described as an edge ecosystem, defined as the boundary between two adjacent land cover types. Wild-type variants originally diverged from two geographical centers with equal parts CBD and trans-Δ9THC in an evolutionary neutral state, since, in its wild state, Δ9-THC is not a significant metabolite of cannabis. Anthropogenic trait selection for different utility characteristics (fiber, seed oil, and medicinal applications) resulted in morphological variations in seed size, coloration, nutrient density, loss of seed dehiscence, reduction in seed perennation variance, inflorescence density and resistance to fracture, stalk thickness, plant height, and trichome size and production as well as the cannabis composition of the trichomes.
Lessons learned from other cultivated medicinal plants
Published research on medicinal plants provides support for the supposition that producing a greater variety of ratios of medicinally active compounds may lead to different clinical outcomes. Akula and Ravishankar 18 reviewed environmental influences on a range of medicinally important plants, finding that the diversity of secondary metabolite profiles was for the most part stimulated when plants were exposed to varying conditions. American ginseng (Panax quinquefolius L.) wild populations produced greater variation in chemotypes than cultivated plants. 19 Clinically, the ratio differences of woods (wild) grown versus cultivated American ginseng suggest a possible explanation for greater brain neuronal activity inhibition produced by the former.20,21
Zandkarimi et al. 22 used an in vitro CB1 receptor system to quantify receptor activation by 16 individual cannabis terpenes, Δ9-THC alone, and Δ9-THC-terpenes mixtures. When combined with Δ9-THC, β-pinene and geraniol responses were lower than the sum of their individual contributions. The researchers noted that the limonene, borneol, and sabinene increased the activation by Δ9-THC at the CB1 receptor by several fold, reflected a synergistic effect, and that it occurred at very low terpene concentrations as well as at terpene/cannabinoids ratios closer to those found in existing strains. Clinically, this may result in reduction of the need for higher Δ9-THC doses, thus minimizing the THC-related adverse effects for sensitive groups such as geriatric and pediatric populations. 23
This article will focus on understanding the role of both abiotic and biotic ecological influences in altering the ratios of phytochemicals in cannabis trichomes. How might reintroduction of these influences impact the formation of secondary metabolites, which ultimately inform clinical application and efficacy?
Trichomes
Because of the nature of metabolic pathways, secretory glands contain a series of structurally related hydrophobic compounds such as fatty acids, flavonoids, cannabinoids, or terpenes. Variations in enzymatic pathways lead to chemical diversification and are evolutionarily driven.24–26 Terpenoid biosynthesis in cannabis is through two independent but interactive pathways: the 2-methyl-D-erythritol-4-phosphate (MEP) pathway is responsible for monoterpenoids and some sesquiterpenoids (Fig. 3), whereas the mevalonate pathway is mostly responsible for sesquiterpenoids. The MEP pathway generates geranylpyrophosphate (GPP), the monoterpenoid precursor of cannabinoids. In addition, there is a strong positive correlation between transcription factor CsMIXTA expression and the expression of both geranylpyrophosphate synthase (GPPS) and olivetolic acid cyclase enzymes, which synthesize the necessary precursors (GPP and olivetolic acid [OA], respectively) for both cannabinoids and, in the case of GPPS, monoterpenes (Fig. 3).
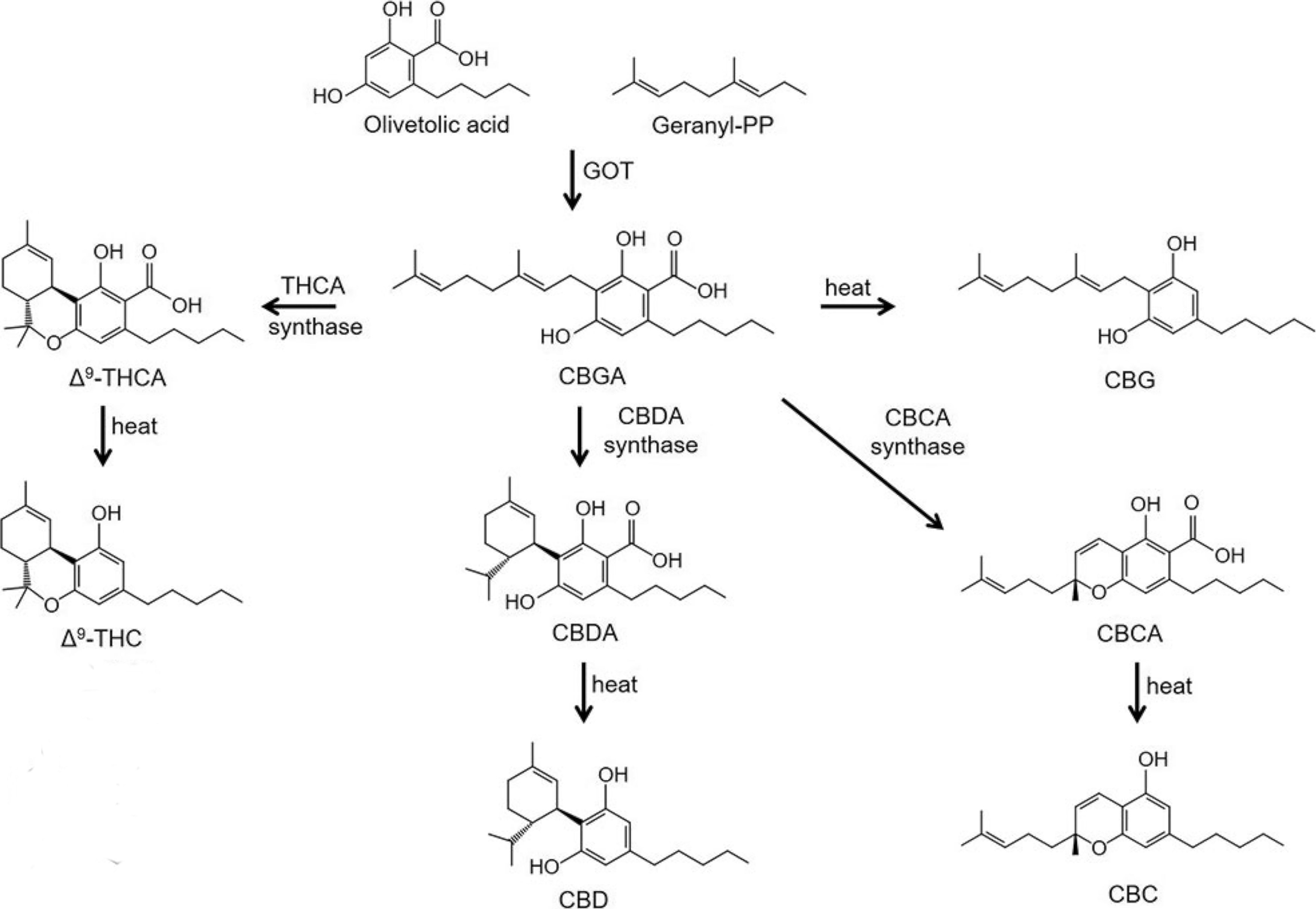
Cannabinoid biosynthetic pathway, leading to the two major phytocannabinoids, Δ9-THC and CBD. Walsh et al. 27 . CBD, cannabidiol; Δ9-THC, tetrahydrocannabinol.
Levels of cannabinoid synthases do not change during floral development,28,29 with their accumulation in the trichomes correlating with cannabinoid content. The ratio of THCA to CBDA synthases also remains constant during flowering13,30 for both cannabinoid pathway genes in chemotype I high Δ9-THC and chemotype II equivalent ratios of Δ9-THC and CBD. During this same period of florogenesis, genes involved in the production of the precursor compound for both terpene and cannabinoid pathways, OA, were downregulated. 31
Glandular trichomes act as production and secretory sites for acidic cannabinoids and terpenes, whereas the glandular structure acts as a storage mechanism to protect the plant since cannabinoids may cause mitochondrial permeability, DNA degradation, and ultimately apoptosis in plants.32,33
Structure and development
Trichomes are fine outgrowths of plant epidermis (Fig. 4). They can occur as scales, papillae, or hairs. Trichome hairs can be branched or unbranched and glandular or nonglandular. Nonglandular trichomes are distributed throughout the plant kingdom in angiosperms, gymnosperms, lycophytes, ferns, and bryophytes.34–37 The functionality of the trichome type is apparent from the development shape of the trichomes. Glandular trichomes enclose secretory cells that contain volatile compounds that need to be kept in a liquid state, released only after trichomes are damaged. For trichomes with resinous, nonvolatile compounds, such as acylsugars or diterpenoids, capitate trichomes act as storage agents, 37 with the compounds exuded onto the trichome surface.27,38

Cannabis female inflorescence trichomes at varying degrees of magnification.
Based on the absence of phytochemical production and/or storage, nonglandular trichomes act as nonelicited structural (physical) defenses. Of the nonglandular trichomes, noncystolithic and cystolithic types are located on the lower side of vegetative leaves and pistillate bracts. Glandular trichomes produce a rich mélange of phytochemicals. Both the bulbous and capitate-sessile (unstalked) glandular trichomes are found on vegetative leaves, whereas capitate-sessile (stalked) forms occur on the bracts and floral leaves.39–48
Floral trichomes in cannabis contain more secretory disc cells than nonfloral (leaf and stem) trichomes.38,49 These excretory cells secrete resin into a subcuticular storage cavity.50,51 Stalked glandular trichomes are primarily concentrated on the calyces and bracts52,53 of female flowers. The resin heads of high Δ9-THC types are larger than glandular trichomes from low Δ9-THC industrial hemp. 50 Thus, marketplace traits correlated with psychoactive potency include larger inflorescences, an associated increase in trichome gland size and increased resin production.
Mature cannabis flowers have stalked glandular trichomes with 12–16 secretory disk cells, which are consistent across hemp and drug type chemotypes. 54 These metabolically active cells in cannabis contain extensive cytoplasmic bridging across the cell walls, with production organelles lining up next to the apical surface where metabolites are secreted.
As the plant matures, cannabis floral glandular trichome heads progress in color from clear to milky to brown, with milky heads being considered a signal for the optimal harvest time and brown heads being a sign that the potency may have been degraded by age.55,56 Sutton et al. 57 suggested this characteristic as a visual progression of change is related to secondary metabolites since fluorescence images of the glandular trichomes associated with secondary metabolites also change in the brown heads.
Trichomes contain membrane-bound, catalytic enzymes necessary to produce complex cannabis oleoresin molecules within vacuoles along with membranes shed from the endoplasmic reticulum of trichome disc cells. Interestingly, these enzymes remain catalytically active under hydrophobic conditions. Additional hydrophilic and amphiphilic compounds have been detected in trichome exudate proposed to create an aqueous microenvironment necessary for biocatalysis. 58
Klyachko et al. 59 suggested that other compounds in the exudate created reverse micelles that would help create an environment of high catalytic activity, an emulsion of cell wall polysaccharides according to Livingston et al. 54 Hydrophilic floral nectars contain carbohydrates, amino acids, and organic acids50,60 and represent microenvironments where enzymes could remain catalytically active61,62 by providing a hydration shell in limited water conditions.
Two types of organelles in the plant cells at the base of the trichome’s head work to combine their products.47,63–65 Vacuoles contain the amphiphilic and hydrophilic constituents described above. The internal pH of the vacuoles is acidic, which helps concentrate secondary metabolites such as phenols, optimize degradative enzyme function, and increase turgor pressure involved in plant structure. 66 The other organelle, the plastid, specializes in producing essential oils and other terpenes. Since cannabinoids and monoterpenes utilize the same precursor GPP to produce their final metabolites in the glandular trichomes, the enzymes for cannabinoid production will be in proximity within the trichome. Cannabinoid biosynthesis catalyzed by THCAs is localized in the cell-surface wall across from the extracellular storage cavity. 67
Biosynthesis of amphipathic terpene precursor occurs in the plastid stroma with the GPP and hydrophobic terpenes mixing freely. 67 The plastid surface in the cytosol is the catalytic site for an aromatic prenyltransferase to biosynthesize the highly lipophilic cannabinoid CBGA, from GPP and OA. Within cannabis glandular trichomes, this congregation of apposed membranes between plastids acts as a nonpolar metabolite channel from plastids to the plasma membrane. 67
Selection pressure effects
Density concentration and distribution of the secretory glands are subject to selection pressure. Samples of inflorescences of medical chemotypes possessed 4 × the volume of glandular trichomes with larger secretory heads compared with cultivated hemp and wild-type chemotypes. 50
This form of selection pressure can be considered heteroblasty, an adaptive mechanism used by plants in response to altered environmental conditions. 68 Since cannabis has had several evolutionary origin points from varying ecologies, as well as selection pressures from domestication, such adaptation would be expected.6,69,70 In other plants, the impact of heteroblasty on trichomes appears to be developmentally regulated, influenced by photoperiod 71 and hormones.72–80
Plant development and growth are controlled by genetic regulatory factors, including transcription and translation, in response to temporal and ecological signals. 81 Regulation of these processes is driven by transcription factors recognizing and binding to promoter regions upstream of the genes of interest.82–84 These transcription factors have multiple binding sites, allowing them to influence multiple gene expression scenarios85,86 pointing to a role activating or inhibiting the expression of specific genes. 87
The subfamily IV of homeodomain leucine zipper (HDZ) transcription factors (HD-ZIP IV) is associated with regulating plant epidermal structure development. 88 Analysis of the HDZ IV gene family was carried out in varied plants including Arabidopsis thaliana (L.) Heynh, Oryza sativa subsp. japonica cv Nipponbare, Zea mays subsp. Mays L., Glycine max (L.) Merr., and Cucumis sativa L.89–93 This transcription factor is expressed in epidermal cells of plant tissue and in the shoot apical meristem. Of note, in these model plant species, the expression patterns of HDZ IV transcription factors appear to regulate the initiation of leaf stomata, trichomes, and root hairs. 94
Promoter analysis of the cannabis HDZ IV transcription factor domains found light regulatory elements occurring most frequently and that the HDZ IV genes also contain multiple elements responsive to plant hormones. 95 The nine HDZ IV genes isolated from cannabis are a comparatively fewer amount than other plants, such as A. thaliana (thale cress) and O. sativa (rice). In contrast, the cannabis genome size of ∼830 Mb 4 is larger than those same plants, which may point to a multifunctional regulatory role in trichome development.
Chemical Ecology
The critical conditions for optimal cannabis growth include influence from abiotic characteristics such as water scarcity,96–99 cold acclimatization,100,101 light intensity, quality and photoperiod, 102 storage temperatures and humidity, 103 and fertilization.104,105
Abiotic factors
Water scarcity
Preliminary observations indicated that cannabis growing under water scarcity correlated with increased trichome density with observed densities of twice the amount in water stressed subjects compared with nonwater stressed subjects. 96 But more research is needed to isolate this mechanism from genotypic variation. Subsequent studies indicate that while water restriction did not influence the inflorescence yield per se, the yields of major cannabinoids that are exclusively produced in the trichomes increased significantly. 97 Similar results occurred during water stress on CBD production in hemp chemotypes. 106 Drought stress increases both acidic and neutral forms of Δ9-THC and CBD.
Cold acclimation
This biological adaptation is a complex, multigenic process that has been estimated to involve modulating the expression of approximately 10% of all genes in a plant’s genome107–109 and involves the induction of multiple genes,109,110 collectively referred to as cold-regulated genes (COR genes). DNA methylation is an epigenetic process that has been shown to play a role in cell differentiation, genome imprinting, development, defense, chromatin activation, and gene expression in cold acclimatization.111,112
The 2015 study by Mayer et al. 101 is the first to characterize cold acclimation response in hemp and to demonstrate that most of the hemp chemotypes tested exhibited the capacity to acclimate to cold by increasing their tolerance to freezing with the range of capacities distributed by chemotype. Their research reported a correlation of DNA methylation profiles to the hemp chemotypes able to respond to a cold acclimation treatment. Data for medical or recreational chemotype are lacking.
Light effects
Early greenhouse work reported that supplemental UV-B increased Δ9-THC measured in flowers of a drug-type chemotype suggesting that cannabinoids act as UV protectants.113–115 However, recent controlled studies using different levels of UV photon fluxes (UV-A = 315–400 nm; UV-B = 280–315 nm; UV-C =100–280 nm) found little to no increase in inflorescence cannabinoids.116–118 In planta, cannabinoids are predominantly in acidic form (THCA and CBDA). Spectral measurements show neutral THC/CBD and their acids have strong absorbance maxima near ∼210–221 nm (UVC-), with weaker bands that extend into the UV-B/UV-A region.119,120 The balance of primary evidence does not demonstrate that either cannabinoid acids or their decarboxylated counterparts function as the plant’s primary UV-protective pigments. Minimal cannabinoid upregulation under UV occurs, whereas leaf flavonoids/phenolics, considered classical UV-screening compounds, appear more responsive in cannabis under UV exposure. 118
Other research suggests that CBD and other cannabinoids may protect cells against DNA-damaging UV radiation, thus supporting the geographical chemotype hypothesis. 121 Higher elevation increases CBDA, terpenes (β-myrcene, α-humulene, and trans-caryophyllene) and cannflavin content,113,122 all of which would potentially be elicited by increased UV-B exposure associated with higher elevation. More than 20 UV-light protectant flavonoids have been identified in cannabis, including quercetin and kaempferol 123 and the more abundant cannflavins A, B, and C unique to cannabis.
Langenheim 124 reported that long-day exposure to sunlight at moderate temperatures led to an increase in trichome density, whereas low temperatures at the same light exposure decreased the density. 125 In general, the composition of terpenes is determined by genetic rather than environmental factors. 126
The predominant wavelengths from high-pressure sodium lamps, which comprise a preponderance of the lighting systems in the greenhouse production of cannabis, are yellow/orange. Magagnini et al. 127 combined blue light and UV-A to increase levels of CBG. Using blue light emitting diodes (LEDs), researchers were able to increase total cannabinoid content by 66% compared with high-pressure sodium vapor light. 128 Specifically, terpene synthesis increased with blue light exposure over the last 3 days of flower development. It is unclear whether the sensory profile was altered.
Canopy position
Light penetration into the canopy is an additional influence that needs to be considered when reviewing light effects on cannabis metabolites. Thus, flowers high in the cannabis plant canopy get full high intensity full spectrum light. Lower down light intensity is reduced and becomes enriched in green wavelengths (reflected by chlorophyll) as well as red light depleted (absorbed by chlorophyll). Studies of Malus domestica (Suckow) Borkh., 1803, nom. cons. (aka: Malus x domestica), 129 Silene littorea Brot., 130 and a series of west African rain-forest plants 131 revealed intracanopy variation in light penetration as a contributing factor to high degrees of plasticity in secondary metabolite production. The position of cannabis tissue within the canopy effects the concentration of cannabinoids, but the underlying mechanism is not fully elucidated, although light concentration and wavelengths are altered by vegetative density. 132
Analysis of cannabis metabolites identified in floral samples from the terminal bud and upper portion of the canopy has higher levels of Δ9-THC than other regions. 133 Additional analysis identified similar intraplant variation in concentrations of THC, CBD, CBG, CBC, and THCV based on vertical position in the plant with increased levels in the higher parts of the plant canopy105,132 (Bernstein et al., 2019a; Stack et al., 2022).
Hawley et al. 134 experimented with the use of subcanopy lighting (SCL) using red–blue (RB) or red–green–blue (RGB) LED lighting (Table 1). While both lighting regimens significantly increased Δ9-THC and Δ9-THCA, they did not impact CBD, CBDA, or CBG. Their findings suggest that subcanopy positioning did influence CBGA, however. The RGB lighting regimen increased several important terpenes.
Effects of RGB LED Lighting as well as Subcanopy Lighting on Cannabinoids and Terpenes

Data from Hawley et al. 134
Student’s t test = 0.05.
CBD, cannabidiol; CBGA, cannabigerolic acid; RB, red–blue; RGB, red–green–blue; THC, tetrahydrocannabinol; THCA, tetrahydrocannabinol acid.
Clones of a C. sativa hemp chemotype (Kompolti variety) grown in Italy at elevation (1.2 km above sea level) exhibited three times greater quantities of CBD, CBDA, and CBGA, a higher total amount of terpenes β-myrcene, trans-caryophyllene, and α-humulene, as well as greater amounts of cannflavins A, B, and C compared with the same clones grown at 130 m above sea level. 122
Biotic factors
What is less well understood, nor is it explored as extensively in production growth methods are biotic elicitors, including phytohormones,135,136 microbiome influences, 137 insects, herbivory,138–141 and edge effects. The following will investigate how biotic factors affect cannabis growth and phytochemical production. Note the multitrophic plant defense interactions in Figure 5. The focus of this discussion will be on herbivory and microbial interactions.
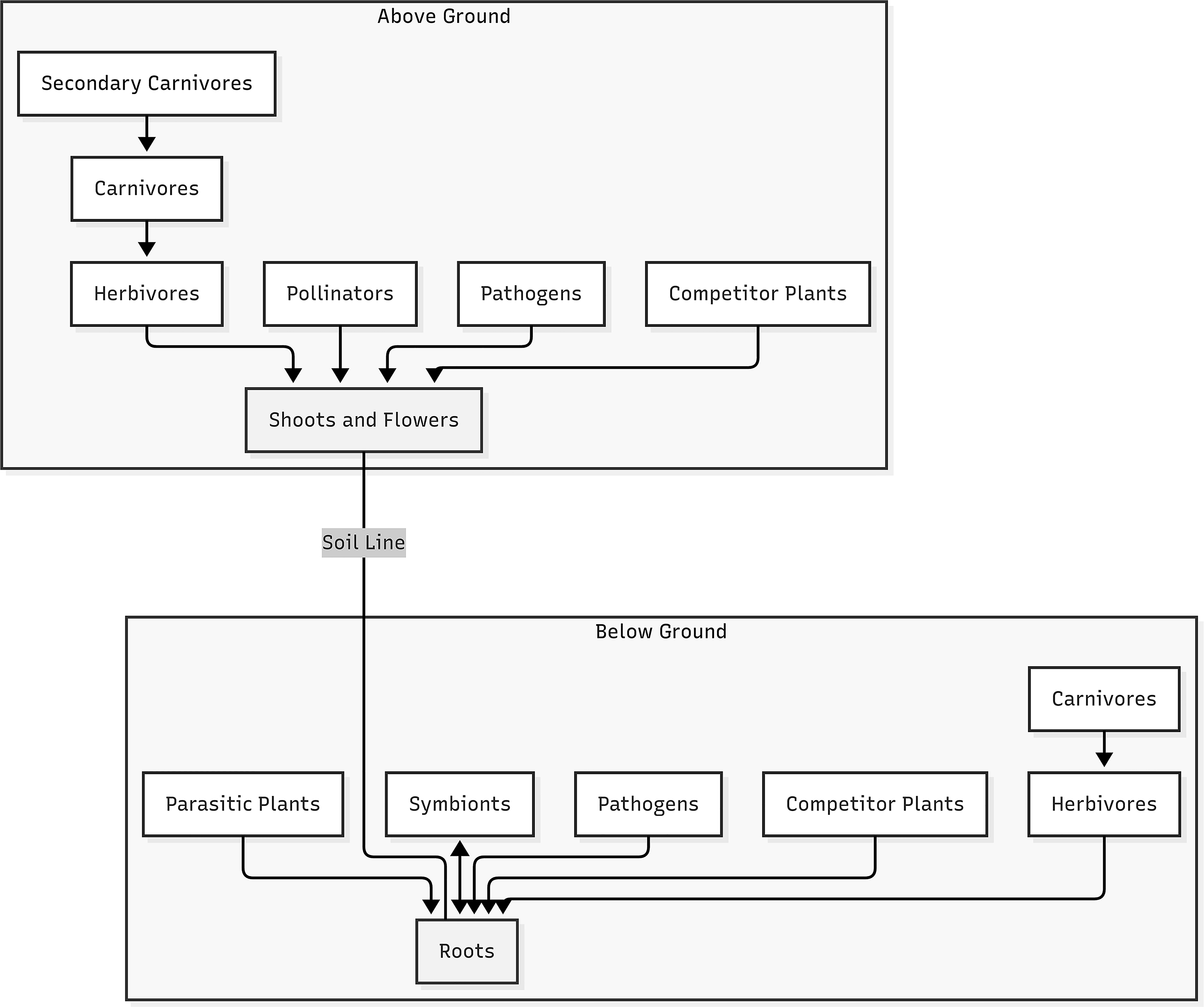
Multitrophic interactions in plant defense. Modified from Bruinsma and Dicke. 142
Herbivory
Plant defensive metabolites are defined as either phytoanticipins, which are produced and stored constitutively, or phytoalexins, which are elicited in response to pathogenesis or plant attack,142,143 with the latter considered an induced resistance. Allelopathic plant metabolites are released constitutively into the surrounding environment (usually soil) and inhibit germination or growth of other plants. There is a larger body of evidence for cannabinoids and terpenes acting as phytoanticipins and phytoalexins. Evidence that cannabis metabolites act as allelopathic agents is lacking. However, Shikanai and Gauge 144 reported that the hemp chaff (“skin” of the stem) residue may be an effective allelopathic agent for the control of certain weeds.
The impetus for induced resistance is influenced by biotic factors. Three models for the utility of induced metabolic responses have been proposed: (1) the metabolic costs of defense mechanisms are conserved until the risk of herbivory is elevated; (2) metabolite variability reduces the nutritive quality of the host plant to the herbivore; and (3) intracommunity signaling (alarm) modulates greater resilience in surrounding plants.145,146 In addition, the actual magnitude and effect of elicitation and alarm signaling on constituent profiles are likely species specific and highly variable. 147 In plants producing a range of secondary metabolites, different pathways may be characterized as constitutive or elicited, depending on the plant response to stressors. In Nicotiana tabacum trichomes, the phytoalexin sesquiterpenes are synthesized only in response to a pathogen attack, whereas a constitutive pool of terpenoid precursors using diterpene synthesis is present, even absent elicitation events. 49
Evidence exists that in a wide array of plant species, alarm signaling upregulates secondary metabolite production including terpene synthase pathways.148–150 Several plants have exhibited the elicitation of secondary metabolites by intraplant and cross organism signaling due to biotic factors such as insect feeding, plant wounding, and exposure to exogenous chemicals. 147 Only limited results for biotic factors have been published on cannabis.
Cannabis plants produce several classes of phytochemicals (Table 2), many of which appear in studies showing that these compounds chemically mediate plant defense against insects via pest repellence. 169
Cannabis Terpenes and Exemplar Plants
The utility of terpenes specifically acting as defense compounds has been previously elucidated, 170 and increased terpene expression in plants due to defensive signaling has been identified in several species.171,172 In cannabis, increased terpene concentrations are associated with inflorescence maturity, 173 which could be an adaptive mechanism to resist herbivory and lends credence to the theory that terpene composition would be modulated by defensive signaling.
Evidence of increased cannabinoid production in Δ9-THC dominant chemotypes in response to stress has repeatedly been demonstrated.140,174 Six plants with mature flowers infested with the spider mite Tetranychus urticae Koch produced significant increases of all terpenes and of neutral cannabinoids Δ9-THC, CBC, and CBG in flowers; in leaves, THCA, CBDA, and Cannabinolic Acid also rose. 141 Table 3 provides the relative distribution of the main cannabis phytochemicals, which points to a strategic allocation of cannabinoids, terpenes, and flavonoids at plant tissue most likely to need defense from herbivores, insects, and airborne microbial pathogens.
Phytochemical Concentration (%) of Cannabinoids, Terpenes, Flavonoids, Sterols, and Triterpenoids in Different Tissue
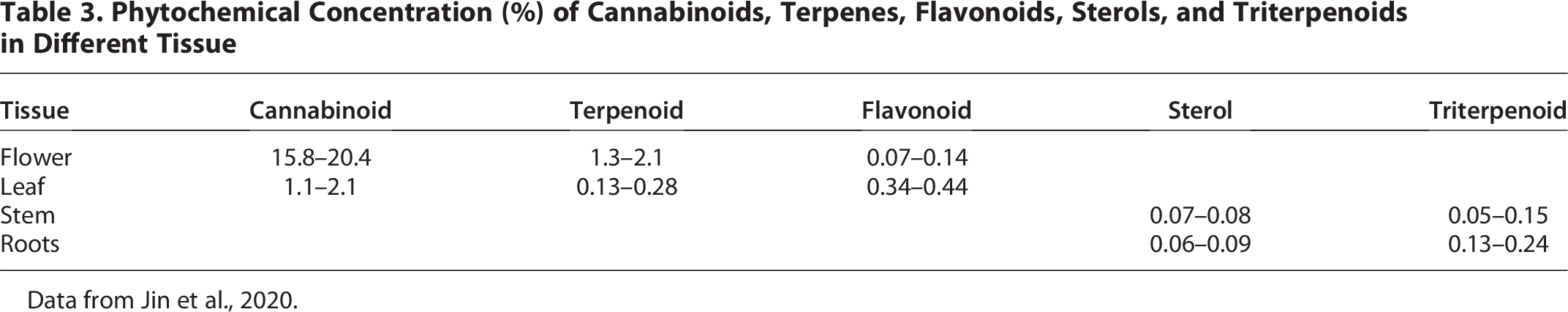
Data from Jin et al., 2020.
Limited examples from cannabis research provide evidence of increased terpene concentrations resulting from stressors, including evidence from aquaponic cultivation. 175 Winnacker and Rieger 176 reported that the essential oil purified from the floral whorls of cannabis proved effective in killing aphids, mosquito larvae, tobacco cutworms, and houseflies. The essential oil comprised of 45% beta-caryophyllene, 25% myrcene, 18% α-pinene, 8% humulene, 5% β-pinene, 5% ocimene, and 3% farnesene. Only 3% of beneficial insects, such as ladybug larvae, were killed at the highest concentration.
Floral terpenes, such as those found in cannabis trichomes, have been shown to repel insects 139 and attract pollinators and insect predators of potential insect herbivores of the plant. 45 In cannabis, the lower fan leaves express higher concentrations of bitter sesquiterpenoids to act as antifeedants for grazing animals, domestic,177,178 and wild.179–181 Another major group of phenolics found in cannabis includes stilbenoids such as dendinobin that act as insecticides and antibacterials. Many secreted terpenes also protect plants from herbivores and microbial pathogens.182–185
Plant/soil microbe interaction
Soil microbes provide numerous ecosystem services to plants—nitrogen fixation, production of growth stimulants, improved water retention, and suppression of root diseases,186–189 which occur in the rhizosphere.190,191 Both bacterial and fungal endophytes alter the phytochemical makeup of their hosts and shape the terroir of the plants, as shown in wines.149,150,169
Experiments with numerous medicinal plant species have shown that inoculation with bacterial endophytes increases secondary metabolite synthesis and accumulation, as well as trichome density.192–194 Ginseng is a salient example, considering the widespread use of clonal propagation in farmed crops. This propagation method is used extensively in cannabis agriculture. Song et al. 195 demonstrated enhanced ginsenoside production in adventitious root cultures of Panax ginseng C.A. Meyer when inoculated with a chemotype of Bacillus spp. (LB-53) isolated from field grown ginseng plants. Increases in both biomass and ginsenoside accumulation were greater with LB-35 inoculation than previously identified elicitors such as methyl jasmonate (MeJA). The endophytic inoculation in combination with MeJA led to reduced biomass and ginsenoside accumulation potentially due directly to toxic effects of MeJA in culture. These results may also represent the presence of exogenous MeJA acting in a negative feedback loop with the endophytic production of the same hormone.
Plant growth-promoting rhizobacteria and endophytes produce specific phytohormones, which influence plant growth and development—auxins, 196 gibberellins (GAs), abscisic acid (ABA), and ethylene (ET). They can also have a direct effect on the plant by modulating the plant’s endogenous phytohormone levels.197,198
Results in modifying cannabinoid content following direct inoculation of cannabis with bacterial endophytes have been mixed. A mixture of four bacterial endophytes significantly increased CBD and Δ9-THC contents199,200 finding that CBD concentration was associated with root endogenous bacterial communities, whereas Δ9-THC was influenced by temperature, pH, and salinity of soil. A bacterial biostimulant called Mammoth P™ boosted aerial biomass by 16.5%, but no cannabinoid enhancement was detected. 201
Analysis of surface-sterilized plant tissue from Bedrocan BV cannabis vouchered specimens identified 30 different fungal endophytes. 202 The degree of fungal endophytic association was dependent on geographical origin.202–205 It is probable that hemp selectively established relationships with mycorrhizal fungi to counteract abiotic stress through symbiosis. 206
Nitrogen uptake in plants occurs in the root zone, and assimilation occurs in actively growing tissue, particularly where bacteria are most active. This region turns out to be rich in carbohydrates. Micci et al. 207 noted that efficient nitrogen transfer via endosymbiosis occurred in glandular trichomes, in both hops (Humulus lupulus L.) and hemp.
Using plant hormone to alter trichomes
Plant hormones are molecules produced in response to pulsed changes in plants, due both to internal development and external elicitation. Egamberdieva et al. 208 provided evidence that ecosystem influences elicit a more robust response than the experimental application of hormones.
Mansouri et al. 135 noted that GA treatment increased the number of cannabis trichomes and reduced the amount of Δ9-THC produced. This suggests a strategic use in growing hemp to reduce Δ9-THC levels.
Cytokinin (CK) induces floral induction and trichome formation in cannabis, which are intimately connected through similar genetic pathways. 209
Jasmonic acid (JA) is involved in the formation of plant memory of herbivory with population level damage cues creating heterogeneity in JA bursts. 210 The hormone m increases the number of multicellular trichomes or trichome size 211 and density.212,213 It also increases terpene production.214,215 A related hormone, MeJA, increased Δ9-THC by 21% when applied. 216
Salicylic acid (SA) increased THCA expression, while decreasing CBDA and metabolic precursors for terpenes. The hormone acts antagonistically to JA stimulation by reducing trichome formation. 136
Direct application of ABA increased Δ9-THC levels. 135
ET, a gaseous plant hormone, is produced in response to herbivore attack dramatically exceeds that produced by simple mechanical wounding.195,196,217 This speaks to an herbivore saliva-based signaling for this specific hormone, which when present elicits ET to modify the accumulation of defense molecules, including phenolics 218 and release of volatile terpenes thought to function as indirect defenses.219,220
Garrido et al. 221 found that the foliar application of SA, MeJA, and GABA increased leaf and inflorescence production of CBDA and THCA. They did not report any significant increase in trichome density. Weyers and Paterson 222 suggest a focus on the “whole plant” aspects of hormonal control and a need for clear delineation of spatial and temporal activity of these wide ranges of physiologically active plant hormones. Plant systems react to multiple hormonal signals, and each hormone appears to impact multiple systems, 223 a process that allows plants to respond within the existing balance of these hormones present in specific tissue. 224 In opposition to the design of many experiments, concentration of these compounds does not always drive plant response, rather changes in plant physiological activity appeared to be based on spatial differences or temporal changes in the tissue’s responsiveness to a set of plant hormones. 225
We see cross talk between plant hormones serving an oscillatory function, balancing multiple trophic influences on plant development and phytochemical expression patterns. ET modulates other defense signals, including JA.219,226 GAs and CKs work synergistically to increase density of trichomes, 78 whereas SA can reduce trichome formation, antagonistic to JA stimulation. 136
Detailed time-course studies of secondary metabolite induction in glandular trichomes under stress remain scarce. When comparing plant response characteristics between abiotic versus biotic stress, research showed that as herbivory led to glandular trichomes rupture metabolites being released in milliseconds. 227 Gene expression-based secondary metabolism, such as JA-induced pathways, is activated within minutes to hours. 228 Based on abiotic stress, hormone-regulated glandular trichome pathways (ABA, ET, and SA) are activated over hours to days, with metabolite accumulation typically measured at later timepoints. 229 Detailed time-course studies of secondary metabolite induction in glandular trichomes under stress remain scarce.
Edge effects
Growing cannabis in complex ecosystems not only exposes it to other plants but also creates the transition regions between two biological communities. Edge effects are characterized by significant changes in the complex interactions between the different populations or communities230,231 including greater diversity of pollinators and herbivores as well as increased complexity in the soil plant energy matrix. 232 This technique can likely significantly modulate the expression and heritability of traits that modulate trichome phytochemical levels.
Data from Golubkina et al. 233 indicate edge effects lead to greater resistance mechanism via the trichomes of the aromatic plant Artemisia annua L. that responded to environmental stress by producing more levels and variations of essential oils. Given the role of terpenes found in plant essential oils as both attractant and defense mechanism, this trichome protection strategy aids in creating ecological niches and alters relationships between ecosystem organisms. 233
While the edge effect has yet to be formally quantified in cannabis, growers in established legal markets have argued that more complex organoleptic signatures result for cannabis grown in permaculture/regenerative agriculture settings.234–236 This is corroborated by anecdotal reports of sophisticated Central Mid-Atlantic region hobbyist growers who are leveraging permaculture principles and regenerative farming techniques (Personal communication; Courie, June 16, 2023). The influence of light spectral quality on cannabis (Cannabis sativa L.) development is not well defined. It stands to reason that tailoring light quality to the specific needs of cannabis may increase bud quality, consistency, and yield. In this study, C. sativa L. “WP:Med (Wappa)” plants were grown with either no supplemental SCL (control) or red/blue (‘‘Red-Blue’’) or red–green–blue (‘‘RGB’’) supplemental SCL. Both RB and RGB SCL significantly increased yield and concentration of total Δ9-THC in bud tissue from the lower plant canopy. In the lower canopy, RGB SCL significantly increased concentrations of a-pinine and borneol, whereas both RB and RGB SCL increased concentrations of cis-nerolidol compared with the control treatment. In the upper canopy, concentrations of a-pinine, limonene, myrcene, and linalool were significantly greater with RGB SCL than the control, and cis-nerolidol concentration was significantly greater in both RB and RGB SCL treated plants relative to the control. RB SCL yielded a consistently more stable metabolome profile between the upper and lower canopy than RGB or control treated plants, which had significant variation in CBGA concentrations between the upper and lower canopies. Overall, both RB and RGB SCL treatments significantly increased yield more than the control treatment, RGB SCL had the greatest impact on modifying terpene content, and RB produced a more homogenous bud cannabinoid and terpene profile throughout the canopy. These findings will help to inform growers in selecting a production light quality to best help them meet their specific production goals. 130
Germplasm diversity
Reversing the impact of introgression and hybridization on cannabis can be accomplished by using the reproductive approach of pollination and subsequent epigenetic changes. Plant adaptations to dynamic environment changes lead to epigenetic changes captured in seed germplasm.160,161
The prevalent industrial production model of cannabis crops for both medicinal and recreational markets is characterized by attempts to achieve uniformity in chemotype genetic profile through clonal propagation. This approach has manifested as frequently automated and too often attenuates most of the biotic stressors outlined in this article.237–240 The idea of reintroducing biotic influences works against production efficiency. Cannabis growers remove male plants, use clonal propagation, or sow feminized seed to produce only female plants with the goal of increasing the number of flower buds and cannabinoid potency. 241 This represents a loss of potentially favorable clinically useful heritable traits from the male parents. The germplasm from commercially driven agricultural techniques has reduced cannabis diversity. 17 Without biotic influences and open pollination in a complex ecosystem, the diversity of the cannabis genome is at risk.
Open-pollinated sexual reproduction provides the possibility for evolutionary influence.
Historically, as humans spread into new geographical regions, C. Sativa seed stock went with them subsequently being exposed to new and challenging environments. 17 Reintroduction of biotic influences on cannabis and prioritizing wind pollinated propagation methods provides an opportunity to diversify germplasm and with it the variance in phytochemical ratios necessary to create new therapeutic options based on nuanced and complex ecological influence.
Future Work
More research is required to fully understand ecosystem influences on both trichome structure and the ratio of secondary metabolites from abiotic factors such as water scarcity, cold acclimatization, light intensity, and photoperiod. Even less well understood are biotic elicitors, including phytohormones, microbiome influences, insect and herbivory predation, and edge effects.
Other important questions that need investigation include, how do signals from ecosystem interactions alter the accumulation of cannabinoids in trichome storage cells? In the case of herbivory, do these external signals alter the rate of catalytic function necessary for cannabinoid and terpenes production? If trichome cellular disruption occurs by apoptosis, herbivory, pathogenesis, or physical damage resulting in lysed organelle membranes, does the respective mixing of contents change the chemistry of trichome content?
Instead of linear progression and homogenous effectors in the growing process, ecosystem influences generate a pulsed environment that creates a diversity of phytochemical end-points. This approach creates some difficulty in accounting for simultaneous and nonsimultaneous multiple influences on the biological expression of genes in plant systems and subsequent phytochemical profiles. With the advent of robust machine-aided statistical systems, such as large language models and principal component analysis, the complexity can be analyzed along with the influence of individual variables on each other.
Experiments are needed to verify whether increased heterogeneity of cannabis plants that reseed via pollination will lead to changes in trichome structure or phytochemical makeup. The questions driving such an investigation should include whether specific combinations of stressors prime the plant for specific ratios? Are the changes to phytochemical ratios and/or trichome structure in response to external stimuli predictable? And as we follow the succession process of a biotically more complex system, how do we define when and where to intervene? Is there a point where the lack of genetic variability limits the emergent or evolutionary adaptation of the plant? How does one stabilize traits to create consistent ratios associated with a clinical end-point? Or are we generating freeze frame data from an ongoing experiment?
Technical improvements will be needed in real-time measurement of trichome phytochemistry in the field using Raman-based handheld devices for Δ9-THC and CBD content242,243 to provide more discrete data on both temporal and ecosystem input effects on trichome metabolite formation.
Authors’ Contributions
M.T. and J.C. jointly contributed to the conceptualization and methodology of the project. M.T. prepared the original article draft and supervised the overall process. J.C. compiled and formatted all references, figures, and tables and contributed to article writing and editing. Both M.T. and J.C. were involved in review and editing of the text. J.M.B. provided critical review and feedback throughout the writing process.
Footnotes
Acknowledgments
The authors would like to thank Maryland University of Integrative Health for the time to complete this article.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This research received no specific grant from any funding agency in the public, commercial, and not for profit sectors.