
Abstract
Introduction:
Although classified in the United States as Schedule I drugs, medical and recreational cannabis and Δ9-tetrahydrocannabinol (THC) are becoming increasingly consumed by pregnant women. Determining the potential adverse effects following exposure during early development is essential for risk communication.
Materials and Methods:
Following exposure to 0.08, 0.4, or 1 µM THC for 6–96 h post fertilization (hpf) in wild-type zebrafish, latent behavioral effects caused by THC were evaluated at 3 weeks post fertilization (wpf) (juvenile), 11 wpf (onset of sexual maturity), and 24 wpf (adult) and in F1 offspring. Bioenergetics were measured in F0 adult brain mitochondria.
Results:
Open field tests conducted at 3 wpf and 11 wpf indicated concentration-dependent hyperactivity (increased velocity) and increased thigmotaxis (anxiety-like wall-hugging) at the two highest THC concentrations. The 5D F1 generation of zebrafish (exposed to THC as germ cells) also demonstrated significant dark-phase hyperactivity at 120 hpf in the larval photomotor response. Brain mitochondrial bioenergetics in adults developmentally exposed to THC showed that oxygen consumption rate was significantly increased at the two highest THC concentrations for females and the highest concentration for males.
Discussion:
Collectively, these results show that exposure to THC during a critical period of development caused behavioral and mitochondrial alterations that persisted into adulthood and across generations.
Introduction
Cannabis is the most commonly used illicit drug by pregnant women,1–3 and the incidence of usage has more than doubled in the past decade, coinciding with a dramatic increase in potency 4 and an increase in availability due to changes in legalization. In the United States, 10–23% of pregnant women use cannabis to anecdotally manage symptoms of pregnancy such as nausea and sleeplessness.5,6 Thus, a significant number of children are gestationally exposed to cannabinoids each year, while the developmental adverse outcomes and their persistent impacts remain unclear.
Recent meta-analyses on the effects of prenatal cannabis exposure have confirmed lower body weight at birth, increased chance of placement in neonatal intensive care, and increased incidence of neonate morbidity or death.7–11 There are established persistent psychopathologies of prenatal cannabinoid exposure.12,13 Yet, these results are mixed, and outcomes may be influenced by confounders in self-reporting, trimester (timing of developmental exposure), and/or route of maternal use. 14 Furthermore, sex-dependence of behavioral pathologies resulting from exposure are appreciated in animal models15,16 and in humans.17–20
Mitochondrial modulation is often a key event in toxicant adverse outcome pathways.21,22 Regulation of mitochondrial bioenergetics plays a key role in fundamental cellular processes, including energy production, nucleotide biosynthesis, iron metabolism, and apoptosis. During brain development, mitochondrial function is critical, not only for Adenosine Triphosphate (ATP) production but also for the regulation of ROS and cellular death. 23 Adverse developmental outcomes (e.g., developmental delay, induced cardiovascular abnormalities, gastrulation arrest, and mortality 24 ) and behavioral disorders (e.g., impaired memory, disordered food intake, and anxiety 25 ) are commonly linked to disrupted mitochondrial function and ATP deficit. 21 Both endocannabinoids and cannabinoids alter mitochondria-dependent signal transduction and mitochondrial respiration.26,27
Zebrafish (Danio rerio) are a well-established animal model for high-throughput studies of development.28,29 The characteristic external fertilization, rapid development, and embryonic transparency provide unique opportunities, not available in mammal models, to raise large numbers of animals for the study of developmental cannabinoid exposure and persistent generational effects. Zebrafish have conserved endocannabinoid signaling,30,31 and behavioral assessments have both face and construct validity32,33 and can be overlaid on the NIMH Research Domain Criteria framework of human functioning domains. Changes in mitochondrial bioenergetics caused by other stressors are also correlated with altered zebrafish behaviors.34–36 The authors and others have used zebrafish to study the consequences of cannabinoid exposure, including developmental defects.37–41 This study expands the utility of the zebrafish model by highlighting the impact of a developmental Δ9-tetrahydrocannabinol (THC) exposure on growth, behavior, and bioenergetics and considers sex-related differences that persist into adulthood.
Methods and Materials
Fish care and handling
Zebrafish (5D strain; kindly provided by Dr. Robyn Tanguay at Oregon State University) were used in this study and raised under the approved Institutional Animal Care and Use Committee protocol (21-010). The fish were maintained in Aquatic Habitats ZF0601 Zebrafish Stand-Alone System (Aquatic Habitats, Apopka, Florida) at a constant pH (7.0–7.6, 760 µS), temperature (25–28°C), and 14:10 light–dark cycle. The fish were fed twice daily with Gemma 300 micro food (Skretting USA, Utah). Male and female sexually mature fish without any deformities or signs of disease were selected as breeders. Their fertilized embryos were collected and placed into petri dishes containing embryo water (pH 7.6, 60 ppm Instant Ocean, 0.5% methylene blue, 28°C) until 6 h post fertilization (hpf). After 6 hpf, fertilized embryos were transferred to embryo water without methylene blue. All the experiments and exposure protocols were in accordance with approved Institutional Animal Care and Use Committee guidelines and recommendations.
Exposures
The exposure and subsequent developmental assessments are depicted in Figure 1. Zebrafish were exposed to solvent control (0.05% Dimethyl sulfoxide (DMSO)) and 0.08, 0.4, or 1 µM THC (obtained from NIDA Drug Supply Program) 6–96 hpf in glass scintillation vials (n = 5 vials per treatment; 20 embryos per vial; 0.6 mL water per embryo). During exposure, the fish were screened daily for survival and hatch. At 96 hpf, exposure water was removed and nonexposed embryo water was added to each vial until 120 hpf. At 120 hpf, developmental deformities were assessed, and then the fish were raised to adulthood (n = 20 fish per 3-L tank, 5 tanks per treatment). Behavior and growth were assessed at 3, 11, and 24 weeks post fertilization (wpf). Fecundity was assessed at 18/19 wpf and 52 wpf. At 55–60 wpf, the fish were euthanized and brain mitochondrial function was assessed, as described below.
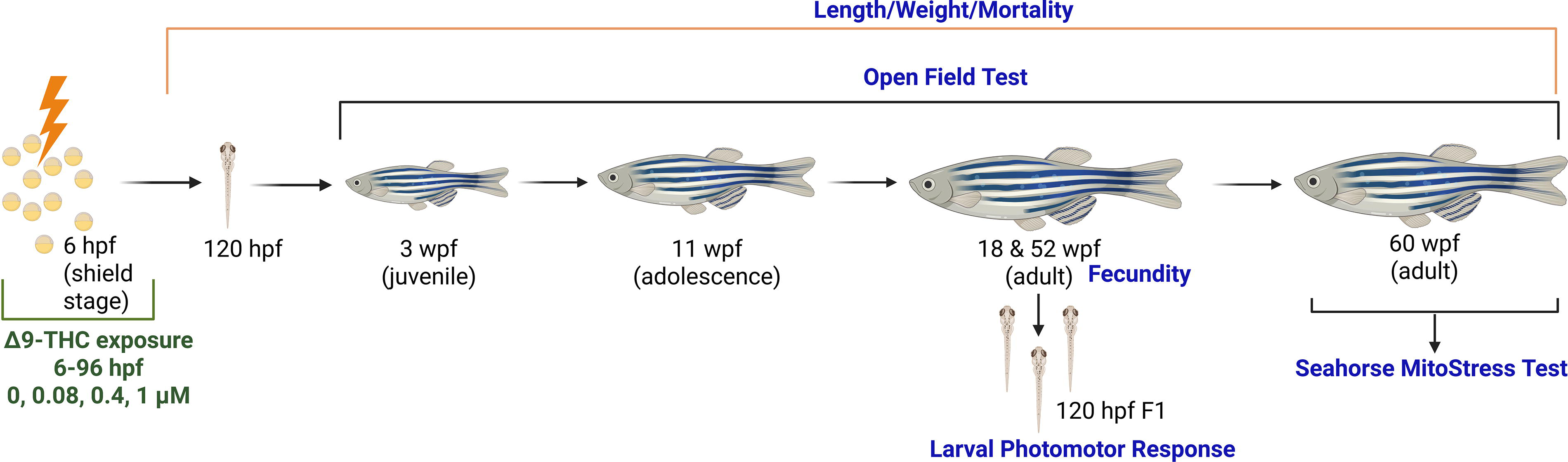
Experimental plan indicating initial 6–96 hpf Δ9-tetrahydrocannabinol (THC) exposure and subsequent behavioral and developmental assessments as fish matured in clean conditions. Figure made with bioRender. Created in BioRender. Willett, K. (2025) https://BioRender.com/xr76ghy.
THC quantification
At 0 h post exposure, water samples were collected to quantify THC water concentrations, as described in Carty et al. (2018). 37 In brief, deuterated THC (THC-d3; Sigma Aldrich; Saint Louis, Missouri) was added to 2 mL of water in addition to 1 mL 2 M NaOH and then extracted twice with hexane:ethyl acetate (9:1). Samples were evaporated to dryness with nitrogen, derivatized with N,O-Bis(trimethylsilyl)trifluoroacetamide with trimethylchlorosilane BSTFA:TMCS (100 µL for 60 min at 90°C), evaporated to dryness with nitrogen, and reconstituted in iso-octane. Samples were then quantified via gas chromatography/mass spectrometry (GC/MS), as previously described. Retention times and ions [quantitative;qualitative] for quantifying THC-d3 and THC were as follows: 8.142 min [374; 389 m/z], 8.167 min [386; 371 m/z], respectively. Actual THC concentrations for the 0.4 and 1 µM THC exposures were 0.32 ± 0.08 and 0.85 ± 0.02 (80–85% of nominal), respectively.
Survival/growth
Survival was recorded daily from 0 to 7 days post fertilization (dpf), every other day from 8 to 14 dpf, and then monthly until 6 months. The length and wet weight of the fish were recorded at 3 (length only), 11, and 24 wpf.
Open field behavior
Open fields were utilized to determine whether exposure affects locomotor activity and thigmotaxis. The behaviors were evaluated in zebrafish at 3 wpf or 11 wpf (n = 24 fish per treatment) and 6 months post fertilization (n = 12 fish per sex per treatment) (exposure paradigm defined above). The arena size was based on the fish life stage. At 3 wpf, juvenile zebrafish were tested in a 12-well plate filled with 5 mL of water. At 11 wpf, the zebrafish at the onset of sexual maturity were tested in a 2-gallon bucket that was 24 cm in diameter, filled with 4 L of water. For adult zebrafish, a 5-gallon bucket that was 28 cm in diameter and filled with 10 L of water was used. All arenas were filled with system water from the primary zebrafish culture unit in a temperature-controlled room. Each arena contained an inner and an outer zone, where the inner zone and the periphery were equal in area. For adult fish, tanks were separated into males and females prior to testing. The fish were given a 5-min acclimation period to adjust to temperature and lighting conditions. To minimize diurnal variation in them, treatment tanks were randomized, and testing was only conducted between 2 and 5 pm. One at a time, the fish were transferred into the arena and assessed for 5 min. After each test, the fish were returned to their original tanks. Following each tank, the water was changed in the bucket. EthoVision was used to track velocity and time spent in the periphery of the arena. The bucket was recorded from above with a color GigE camera. The fish that did not move (were frozen for >280 sec of the 300 sec test) were excluded from the data analysis; this did not occur for 3-wpf fish but occurred for 5 fish at 11 wpf (2x control fish and 3 × 1 µM THC) and 4 fish at 6 months post fertilization (2x control and 2 × 1 µM THC).
Fecundity
At 18 wpf and 52 wpf, fecundity was assessed by setting up breeding tanks (n = 2–4 breeding tanks per treatment) 3 consecutive days for 3 consecutive weeks. On the evening prior to egg collection, males and females (6 males and 6 females) were separated per breeding tank. At the onset of light, the divider was removed, and males and females were allowed to breed for 1 h. Adult fish were returned to their original tank, and eggs (F1) were collected and counted per tank.
F1 larval photomotor response assay
At 120 hpf, F1 larvae from developmentally exposed parents were transferred to a 96-well plate (1 larva per well; 300 µL embryo water per well), and behavior was assessed in the larval photomotor response assay (LPR). The fish were acclimated in light (8000 Lux) for 10 min and then 10 min alternating dark–light phases [0–10 min light; 10–20 min dark (0 Lux); 20–30 min light; 30–40 min dark; 40–50 min light]. Total distance traveled was measured in 2-min intervals in the Viewpoint Zebrabox in each of the lighting phases.
Mitochondrial function
At 55–60 wpf, the fish were euthanized in buffered MS-222, brains were isolated, and mitochondria were purified following a modified protocol. 42 Briefly, brains (n = 4 replicates per treatment per sex; 3–5 brains per replicate) were homogenized in extraction buffer (125 mM sucrose, 250 mM mannitol, 10 mM HEPES, 10 mM EGTA; pH 7.2) with 0.01% bovine serum albumin and 1x protease inhibitors by slowly twisting the pestle 20 times up and down in a borosilicate mortar. To isolate the mitochondria, homogenized brains were centrifuged twice at 700 × g (g-force) for 10 min (4°C) to remove any remaining tissues. The supernatant obtained was centrifuged at 10,000 × g for 15 min (4°C). The mitochondrial pellet was rinsed with extraction buffer and centrifuged again at 10,000 × g for 15 min (4°C). The pellet was resuspended in 50–150 µL of extraction buffer. Mitochondrial protein was quantified using the Nanodrop 2000 (ThermoFisher). Mitochondria (50 or 150 µg for female and male, respectively) were added to 96-well plates in four technical replicates for each biological sample. The plate was centrifuged at 2000 × g for 20 min (4°C) to attach the mitochondria to the bottom of the wells, buffer was removed from each well, and 180 µL Dulbecco’s Modified Eagle’s Medium (DMEM) complete (DMEM plus 1 mM pyruvate, 10 mM glucose, and 2 mM glutamine) were added to each well. After loading the samples, the Cell Mito Stress Test protocol (Agilent Technologies) was followed. In brief, the respective metabolic activators and inhibitors [oligomycin (1.5 µM; a complex V/ATP synthase inhibitor), carbonyl cyanide 4-(trifluoromethoxy) phenylhydrazone (FCCP; 1 µM; a mitochondrial uncoupler), rotenone and antimycin A (0.5 µM; complex I/III inhibitors)] were loaded into the cartridge. For each condition (baseline, oligomycin, FCCP, rotenone/antimycin A), oxygen consumption rate (OCR) was measured 3 times over the course of 18 min (3-min mixing, 0-min waiting, and 3-min measuring) in a Seahorse XFe96 Flux Analyzer (Agilent Technologies).
Statistics
Survival and growth were assessed via repeated measures analysis of variance (ANOVA) and two-way ANOVA, respectively. Fecundity was assessed via ANOVA. Open field dependent measures (total distance traveled, freezing duration) were assessed via one-way ANOVA (immature fish) or two-way ANOVA (sexually mature fish). Differences in oxygen consumption rate were analyzed by ANOVA within each Mito Stress Assay treatment. Group differences were determined via Student-Newman-Keuls (SNK) post hoc tests. For all tests, p ≤ 0.05 was considered significant. As this study was conducted in two batches, the controls from each batch were pooled as they were not statistically significantly different. Sample sizes per endpoint are provided in Supplementary Table S1. All statistical analyses were conducted using the SYSTAT software package in SigmaPlot 14.0.
Results
Survival and growth
Survival was recorded during the exposure and also following the end of the exposure. THC was not acutely toxic (during the exposure) at any of the concentrations tested; however, by 12 dpf (exposure ended at 96 hpf), 1 µM THC had significantly increased mortality by almost double compared to controls (54 ± 16% and 26 ± 5%, respectively) (Fig. 2).
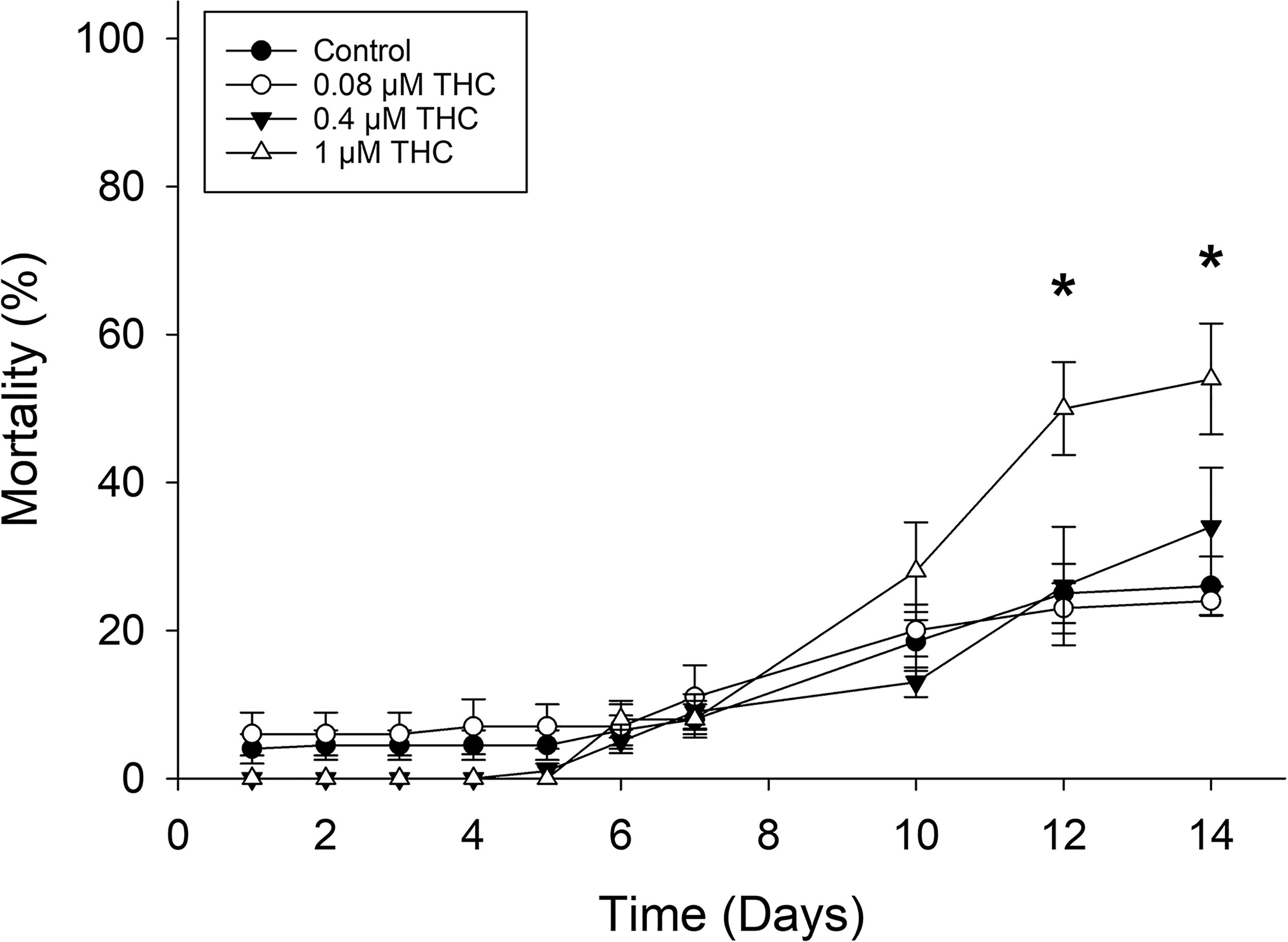
Zebrafish were exposed to control, 0.08, 0.4, and 1 µM THC from 6–96 hpf. Survival was recorded daily from 0 to 7 dpf and every other day from 8 to 14 dpf. By 12 dpf, the 1 µM THC exposure was significantly more lethal than control (n = 10 controls, 5 treatments, two-way repeated measures ANOVA, both time and treatment and their interaction were statistically significant). *Indicates statistically significant difference from the control.
In addition, growth was measured at 3, 11, and 24 wpf. By 3 wpf, animals exposed to THC showed a dose-dependent decrease in length, which was significantly decreased relative to the control in the highest concentration of 1 µM (Fig. 3A–B, D). By 24 wpf, length was not significantly different (Fig. 3C); however, weight was significantly different at this stage (Fig. 3E). Body weight of male and female zebrafish exposed to THC exhibited a biphasic response, with those exposed to the lowest concentration (0.08 µM) weighed significantly less than controls, while the fish exposed to the highest concentration (1 µM) weighed significantly more than controls (Fig. 3E).
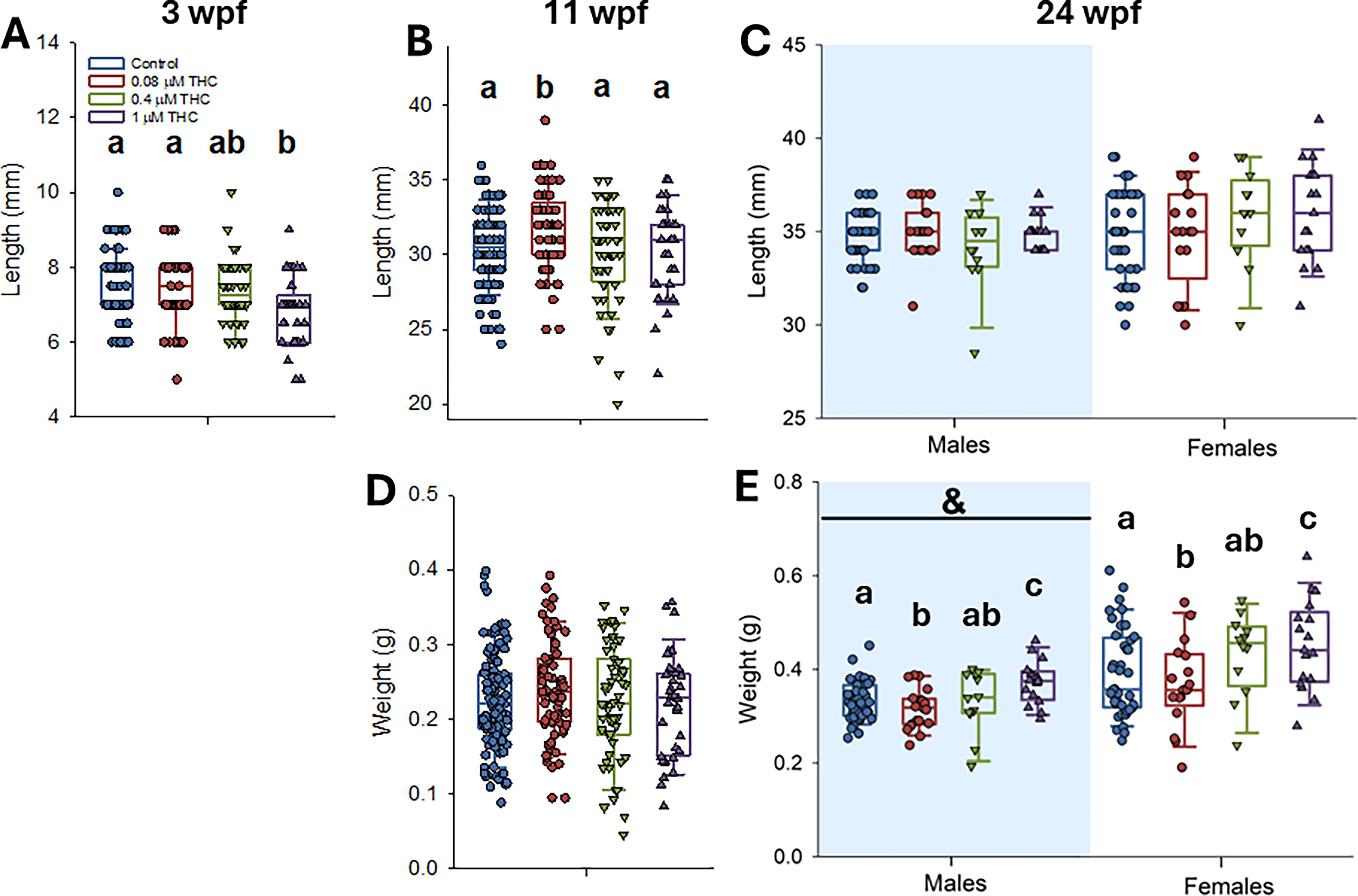
Zebrafish were exposed to control, 0.08, 0.4, and 1 µM THC from 6 to 96 hpf. Growth (length A–C and weight D–E) was measured at 3 (length only, n = 37 −139), 11 (n = 36–122), and 24 (n = 12–42) wpf. There were more controls than treated as exposures were conducted in batches; controls were not significantly different between trials and were pooled. For 3 and 11 wpf, data were assessed by one-way ANOVA with Dunnett’s post hoc (treatment as the factor) or one-way ANOVA on ranks with Dunn’s post hoc (if the data were not normally distributed/equal variance) and at 24 wpf by two-way ANOVA (treatment and sex as factors). Letters not in common indicate a significant difference between treatments, an & indicates that there was a significant difference between males and females.
Behavior
Behavior was assessed at 3, 11, and 24 wpf in an open field test (Fig. 4). Hyperactivity, measured as significantly increased velocity following exposure to 0.4 and 1 µM THC, persisted throughout the entire study, and was last measured at 24 wpf to be significantly increased by 34% and 83% in females, and 90% and 185% in males, exposed to 0.4 and 1 µM, respectively (Fig. 4A–C). In addition to overall activity, thigmotaxis was also assessed. At 3 wpf, the fish developmentally exposed to 0.4 and 1 µM THC spent significantly more time in the periphery (Fig. 4D). This anxiety-like phenotype persisted at 11 wpf for those exposed to 1 µM THC (Fig. 4E) but was not detected by 24 wpf (Fig. 4F). Similar to the effects on growth, there was a detectable biphasic response at 24 wpf, as fish exposed to the lowest concentration of THC (0.08 µM) had statistically significantly lower time in periphery than control (Fig. 4F).
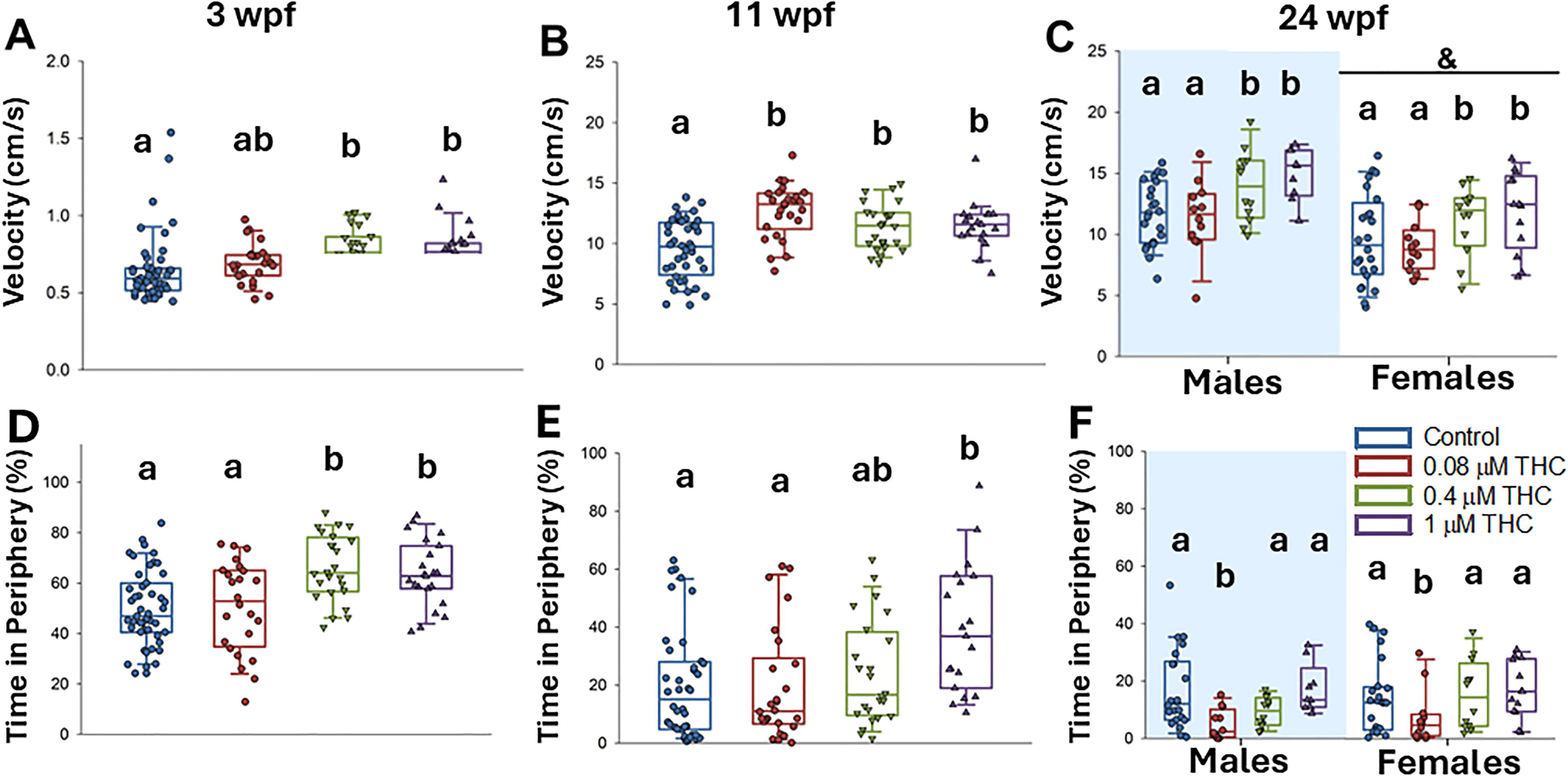
Zebrafish were exposed to 0, 0.08, 0.4, and 1 µM THC from 6 to 96 hpf. Behavior was assessed in an open field test measuring velocity and time in periphery at 3 (n = 24–48), 11 (n = 19–40), and 24 wpf (n = 10–22 per sex). For 3 and 11 wpf, data were assessed by one-way ANOVA with Dunnett’s post hoc (treatment as the factor) or one-way ANOVA on ranks with Dunn’s post hoc (if the data were not normally distributed/equal variance) and at 24 wpf by two-way ANOVA (treatment and sex as factors). Letters not in common indicate a significant difference between treatments, an & indicates that there was a significant difference between males and females.
Fecundity
At 18 wpf and 52 wpf, fecundity was assessed to determine if developmental THC exposure had reproductive impacts. Reproductive success was not significantly altered at either time point following THC exposure (Supplementary Table S2).
F1 LPR
Because hyperactivity was noted in THC-exposed fish (F0 generation) at all stages evaluated, locomotor response was also assessed in their offspring (F1 generation) (Fig. 5). Interestingly, F1 offspring from parents exposed to either 0.4 or 1 µM THC demonstrated a hypoactive response during baseline acclimation (25% and 23% less total distance traveled relative to control, respectively). However, in the dark phase of the LPR assay, they demonstrated a hyperactive response (35% and 45% more relative to control, respectively) (Fig. 5).
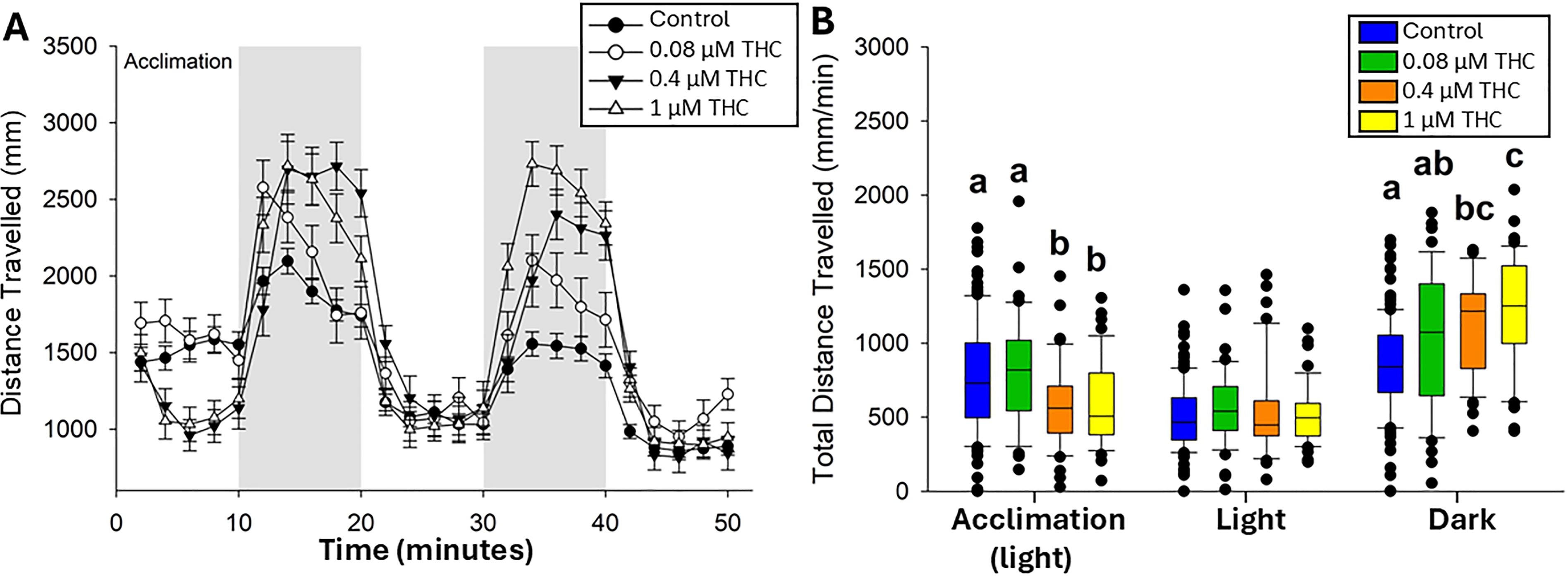
Larval photomotor response (LPR) in 120 hpf F1 larvae.
Mitochondrial function
At 55–60 wpf, the Cell Mito Stress Test was used to determine mitochondrial function following a developmental THC exposure. In females, basal respiration, ATP-linked respiration, maximal respiratory capacity, and reserve capacity were significantly increased following developmental exposure to 0.4 and 1 µM THC by 37% and 81%, respectively (Fig. 6A–B). In males (Fig. 6C–D), only 1 µM developmental exposure caused increased OCR on these measures, which was more modest with an average increase of 27%. However, the percent change compared to baseline OCR was not significantly altered; the increases stemmed from an increased basal respiration (data not displayed).
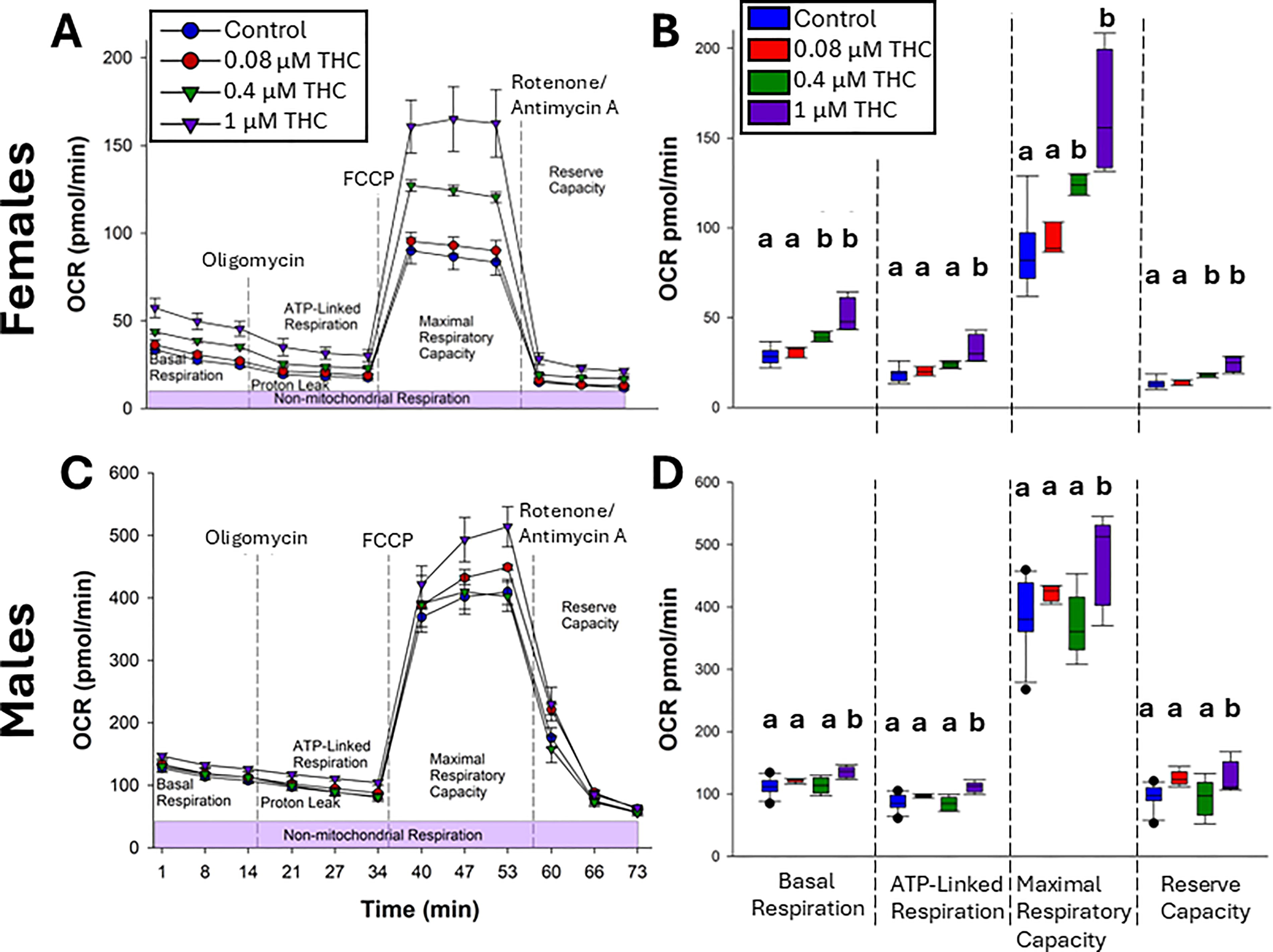
Mitochondrial bioenergetics. Brain mitochondria isolated from 55–60 wpf females (50 µg protein,
Discussion
This study explored the developmental origins of health and disease and multigenerational impacts of THC exposure during a critical period of development (6–96 hpf) by assessing adverse outcomes on growth, mitochondrial function, and behavior in both F0 adults and F1 offspring. Zebrafish developmentally exposed to THC from 6 to96 hpf demonstrated behavioral alterations that persisted into adulthood. Similar to previous studies conducted within this research group, there was acute toxicity only at the highest concentration of THC exposed (1 µM), which reached a plateau by 14 dpf.37,38 After the initial toxicity caused by the developmental exposure over 2 weeks, there was no significant mortality observed for any exposures over the course of the 13–14 month study. In addition, there was a dose-dependent decrease in length of the animals when measured at 3 wpf, which is similar to decreases in size detected by other studies at 96 hpf.38,43 However, changes in growth (length and weight) of the animals developmentally exposed to THC was biphasic by 24 wpf, with the animals being larger at the highest dose (1 µM) and smaller at the lowest dose (0.08 µM).
Zebrafish are well established as a model to study the effect of cannabinoid receptor 1 & 2 modulators such as THC.44,45 In humans, exposure during pregnancy to cannabis has been linked to altered behavioral responses of offspring, such as psychotic-like experiences, externalizing and social problems, 12 increased startle response, 46 impulsivity, hyperactivity, inattention, 47 and negative effects on memory and verbal outcome measures, with persistent adverse behavioral outcomes (e.g., delinquency and depression48,49) through middle childhood and young adulthood,13,50,51 as are the contrasting effects associated with acute versus prolonged or repeated exposure. 52 The hyperactivity and potential effects on anxiety-like behavior in zebrafish exposed during a critical developmental window in this study support effects observed in other animal models/humans.
Previous studies conducted exposing zebrafish to THC that measure behavior at earlier timepoints, such as 96 or 120 hpf, typically measure a decrease in locomotor function, which may be an effect of direct exposure and toxicity of THC. Conversely, in this study, there was a dose-dependent increase in the velocity of fish developmentally exposed to THC, which was detectable at 3 wpf and persisted into adulthood. This increase in locomotor activity was also observed in F1 generation of these fish when measured at 120 hpf. Conversely, there was a dose-dependent increase in thigmotaxis (an anxiety-like behavior) that did not persist into adulthood. In fact, similar to growth, by adulthood there was significantly reduced thigmotaxis in fish exposed to the lowest concentration of THC (0.08 µM). Importantly, in general, adult female zebrafish have lower open field activity, so considering sex as a biological variable is essential; yet, we have found developmental THC exposure (at both 0.4 and 1 µM) causes persistent hyperactivity in both sexes. Behavioral decrements may be more pronounced in males, because in other animals, androgen-mediated regulation of the endocannabinoid system was linked to behavioral alterations at later life stages 53 and reviewed in Bara et al. (2019). 54
Mitochondrial function is fundamental for development and normal physiology. While developmental studies assessing the neural effects of cannabinoids on mitochondrial function in whole-animal models are rare, RNASeq data from 96 hpf zebrafish larvae exposed to THC from 6 to 96 hpf revealed many of the pathways associated with energy metabolism were altered, 38 informing our interest in prolonged impact on mitochondrial function. In this study, for the first time, we measured altered mitochondrial function of fish developmentally exposed to THC. Specifically, we showed a dose-dependent increase in mitochondrial activity for all four parameters measured (increased basal respiration, ATP-linked respiration, maximal respiratory capacity, and reserve capacity). Such altered mitochondrial function may represent a consequence of fetal programming in response to gestational stress and may partly explain how animals were able to maintain a hyperactive state into adulthood, relative to controls. Indeed a similar mitochondrial profile, characterized by greater basal oxygen consumption rate and maximal/spare respiratory capacity, was observed in circulating immune cells collected from adults that were born pre-term. 55 Similar to the present work, effects were greater in females (not reaching statistical significance in males). 55 Another report found maternal stress during pregnancy to be associated with greater mitochondrial content and bioenergetic capacity in their children’s leukocytes. 56 These effects may represent an adaptive response to gestational challenge.
While the exposure design of this study (a short 6–96 h) was to measure developmental toxicity and multigenerational impacts relevant to human health, these findings have important implications for environmental exposures as well. THC and a metabolite THC-COOH have been detected in concentrations of up to 2.6 µg/L in untreated wastewater, 0.169 µg/L in treated wastewater, and 0.080 µg/L in surface water. 57 Our study exposed zebrafish to 0.08–1 µM (75–3750 µg/L), concentrations that are higher than found in the environment, but for a much shorter exposure period (as the fish in the wild would be exposed chronically over their lifespan to cannabinoids found in wastewater treatment effluent). Thus, there is potential for fish adverse outcomes in the environment, which could be explored by chronic studies.
Collectively, these results show that exposure to THC during a critical period of early development caused behavioral alterations that persisted into adulthood and across generations. Notably, some effects like hyperactivity and altered mitochondrial function occurred in a dose-dependent manner, while others, such as growth or anxiety-like behavior occurred in a biphasic manner. The zebrafish model provides a useful approach to further probe the molecular and cellular mechanisms and developmental windows of susceptibility for the persistent neurobehavioral impacts of THC exposure.
Footnotes
Acknowledgments
The authors would like to thank University of Mississippi undergraduate students Victoria Jackson, Morgan Appenrott, Jenna Cripe, Su Hahm, Ava Stegal, and Marissa Skulsky for helping with exposures, data collection, and data analysis. Dr. Courtney Roper (UM) helped with THC GC/MS detection methods. The authors thank Fakhri Mahdi for her assistance with the Seahorse assay. They would also like to thank Dr. Robyn Tanguay from Oregon State University for kindly providing the 5 D zebrafish.
Author Disclosure Statement
The authors declare that there is no conflict of interest.
Funding Information
This work was supported by the National Institute on Drug Abuse Centers of Biomedical Research Excellence program, funded by the National Institute of General Medical Sciences (COBRE-NIGMS) P20GM104932 and R01DA057317. The content is solely the responsibility of the authors and does not necessarily represent the official views of the National Institutes of Health.
Supplemental Material
Abbreviations
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.